Introduction
Prostate cancer (PCa), also known as carcinoma of
the prostate, is the development of cancer in the prostate gland.
The mortality of PCa is the second leading cause of cancer-related
mortality in the United States (1).
PCa grows slowly and cell proliferation depends on the androgen
levels in the early stage. Conventional anti-androgen therapy can
control the progression of the disease at this stage. Yet, the
cancer becomes difficult to cure when it progresses into
castration-resistant prostate cancer (CRPC) (2). Thus, patients who suffer from CRPC
have an extremely poor prognosis. Thus, elucidating the mechanism
of CRPC is critical to prevent PCa progression. The proliferation
and survival of prostate cells depend on the essential hormone
androgen and the androgen receptor (AR), which is the principal
receptor responsible for mediating the physiological effects of
androgens. The AR plays a critical role in the progression of
androgen-sensitive to castration-resistant PCa. PCa activity of the
AR is augmented and may be activated by other stimuli or adaptive
signaling pathways, during the progression to castration-resistant
PCa (3,4). Many drugs that are applied in the
clinic may block the binding of androgens to ARs attenuating the
activity of AR and inhibiting the proliferation of PCa cells
(5). However, the AR may become
more sensitive to androgens and may be activated by adaptive
pathways as PCa becomes resistant to castration or anti-androgen
treatment (6,7). Therefore, development of new drugs
that directly target the AR protein is urgently needed.
Capsaicin, a pungent spicy ingredient extracted from
red peppers, has been recently demonstrated to induce growth
inhibition in many types of tumors including PCa (8). Sánchez et al demonstrated that
capsaicin enhanced apoptosis by induction of reactive oxygen
species (ROS) generation, a decrease in the mitochondrial inner
transmembrane potential and activation of caspase-3 in prostate
cancer cells (9). However, the
relationship between capsaicin and AR and the relevant mechanisms
remain largely unknown.
microRNAs (miRNAs) are a class of small non-coding
RNAs (ncRNAs) that regulate gene expression by repressing
translation and have been proven to be involved in the regulation
of critical processes, such as proliferation, differentiation and
apoptosis in various types of cancer (10). Among the miRNAs, miR-449a functions
as an important tumor suppressor in many types of tumors by
targeting different genes. In PCa, Noonan et al found that
miR-449a caused cell cycle arrest by suppressing Rb phosphorylation
(11) and by directly targeting
HDAC1 (12). By regulating the
activation of Rb, miR-449a also induced growth arrest and
senescence in an Rb-dependent manner (11). Previous studies demonstrated that
capsaicin may influence the expression of miRNAs or synergize with
miRNAs to enhance its tumor-suppressive function (13,14).
However, whether capsaicin treatment regulates miR-449a expression
and thereby changes the activation of downstream genes remains
unknown. In the present study, we explored the function of
capsaicin on miR-449a expression and further researched the
downstream effects of miR-449a overexpression induced by capsaicin,
aiming to elucidate capsaicin/miR-449a interaction and its
influence on PCa.
Materials and methods
Reagents
Capsaicin (M2028) was purchased from Sigma-Aldrich
(St. Louis, MO, USA) and was diluted into a 200 mmol/l stock
solution with DMSO. Anti-AR rabbit polyclonal antibody (N20) and
anti-glyceraldehyde-phosphate dehydrogenase (GAPDH) mouse monoclone
antibody were purchased from Santa Cruz Biotechnology (Santa Cruz,
CA, USA). Anti-cyclin D1 mouse monoclonal antibody was obtained
from Proteintech (Chicago, IL, USA).
Cell culture
Human C4-2 and LNCaP cells were obtained from the
American Type Culture Collection (ATCC; Rockville, MD, USA). The
two cell lines were cultured at 37°C in humidified air containing
5% of CO2. Both cell lines were cultured in RPMI-1640
medium with 10% fetal bovine serum (FBS) (Gibco, Carlsbad, CA, USA)
containing 1% of penicillin-streptomycin.
Lentiviral infection
C4-2 and LNCaP cells were seeded in a 10-cm dish and
a lentivirus that overexpressed miR-449a was transfected into 293T
cells. Forty-eight hours after transfection, viral supernatant
fractions were collected and then infected into the C4-2 and LNCaP
cells along with 10 μg/ml Polybrene. Twenty-four hours after
infection, the medium was replaced with fresh medium.
Cell proliferation assay
Cell viability was analyzed by MTT assay. Cells
(8×103/well) were seeded in 96-well culture plates and
incubated overnight. Then, the cells were treated with capsaicin
(0, 50, 100, 150 and 200 μM) for 2 days. After being washed
once, 0.5 mg/ml of MTT was added and incubation was carried out at
37°C. Four hours later, the culture medium was removed carefully
and dimethyl sulfoxide (DMSO) was added to solubilize the formazan
crystals. Finally, the absorbance was measured at a wavelength of
490 nm using a microplate autoreader (Bio-Tek Instruments Inc.,
Winooski, VT, USA). Independent experiments were repeated in
triplicate.
Flow cytometric analysis
Flow cytometric analysis was applied to evaluate the
cell cycle progression and apoptosis. After the indicated capsaicin
treatments for 2 days, the cells in each group were harvested and
washed twice and then 70% ethanol was used to fix the cells at
−20°C for at least 24 h. For the apoptosis assay, the cells were
washed twice and stained with propidium iodide (PI) and Annexin
V-FITC (BD Pharmingen, San Diego, CA, USA) at appropriate
concentrations and time according to the manufacturer’s protocol.
For cell cycle detection, the cells were washed twice and incubated
with 50 μg/ml PI and 50 μg/ml RNase A at room
temperature for 30 min. Then, flow cytometry was performed using a
FACSCalibur system with CELLQuest software version 3.3 (both from
Becton Dickinson, San Jose, CA, USA), and the cell cycle
distribution was calculated using ModFit LT software (version 3.0;
Verity Software House, Topsham, ME, USA).
Total RNA isolation and reverse
transcription
Total RNA for each group was extracted using Fast
200 kit (Feijie Biotechnology, Shanghai, China) and was reverse
transcribed using PrimerScript™ RT Master Mix (Takara Biotechnology
Co., Ltd., Dalian, China) according to the manufacturer’s protocol.
5X PrimeScript RT Master Mix (2 μl), RNase
free-dH2O (7 μl) and total RNA (1 μl) were
mixed. Then, the protocol was followed: 37°C for 16 min and 85°C
for 5 sec.
miRNA isolation and reverse
transcription
miRNA in the capsaicin-treated groups of the C4-2
and LNCaP cells was extracted using TRIzol and quantified at an
absorbance of 260 nm. RNA (5000 ng) was reverse transcribed using
PrimerScript™ RT Master Mix according to the
manufacturer’s protocol.
Quantitative real-time polymerase chain
reaction (qPCR)
qPCR was carried out using the Bio-Rad CFX96™
real-time system (Bio-Rad, Hercules, CA, USA) and the SYBRR Premix
Ex Taq™ II system (Takara). The SYBRR Premix Ex Taq II(12.5
μl), 1 μl primer (10 μM), 2 μl cDnA
solution and 8.5 μl Rnase-free water were mixedtogether,
followed by the second stage of pre-degeneration at 95°C for 30
sec, one repeat and PCR reaction, 95°C for 5 sec followed by 60°C
for 30 sec, 40 repeats and the third stage of dissociation at 95°C
for 15 sec followed by 60°C for 30 sec and at 95°C for 15 sec. The
data were collected. gAPDH was used as the internal control.
Primers used for amplification were as follows: AR sense,
5′-CCAGGGACCATGTTTTGCC-3′ and AR antisense, 5′-CG
AAGACGACAAGATGGACAA-3′; PSA sense, 5′-GTGTGT GGACCTCCATGTTATT-3′
and PSA antisense, 5′-CCACTC ACCTTTCCCCTCAAg-3′. The primer for
miR-449a was purchased from Takara.
Western blot analysis
After capsaicin treatment for 48 h, proteins from
the C4-2 and LNCaP cells were harvested with RIPA buffer [50 mM
Tris (pH 8.0), 150 mM NaCl, 0.1% SDS, 1% NP-40 and 0.5% sodium
deoxycholate] containing 1% cocktail and 1 mM PMSF. The cellular
lysates were centrifuged at 15,000 rpm for 15 min at 4°C, and the
supernatants were harvested and mixed with loading buffer. The
protein content was quantified using the Enhanced BCA Protein Assay
kit (Beyotime Institute of Biotechnology, Jiangsu, China).
Then, the protein lysates were separated by SDS-PAGE
on 10% gel and transferred to nitrocellulose membranes. After
blocking in 5% non-fat dry milk in TBS, the membranes were probed
with specific primary antibodies overnight at 4°C, washed three
times with TBS Tween-20 and then incubated with HRP-conjugated
secondary antibodies at room temperature for 1 h. Then, the
membranes were washed with TBS Tween-20 and incubated with
secondary antibodies (supplied by Licor, Rockford, IL, USA) coupled
to the primary antibody at room temperature in the dark for 1 h,
followed by washing as above in the dark, drying with neutral
absorbent paper and scanning by Odyssey detection system (Licor).
GAPDH was used as the loading control.
Statistical analysis
Data are expressed as the mean ± SE from three
independent experiments. Statistical analysis was carried out using
a Student’s t-test and SPSS 15.0 software (SPSS Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant result.
Results
Capsaicin inhibits the proliferation of
AR-positive PCa cells
Capsaicin has been proven as an antitumor drug due
to its effect on promoting growth inhibition in various types of
cancer. In order to verify whether capsaicin affects the
proliferation of AR-positive PCa cells, we analyzed the effects of
capsaicin on two PCa cell lines: C4-2 and LNCaP. The cell lines
were grown in RPMI-1640 medium with 10% FBS and treated with
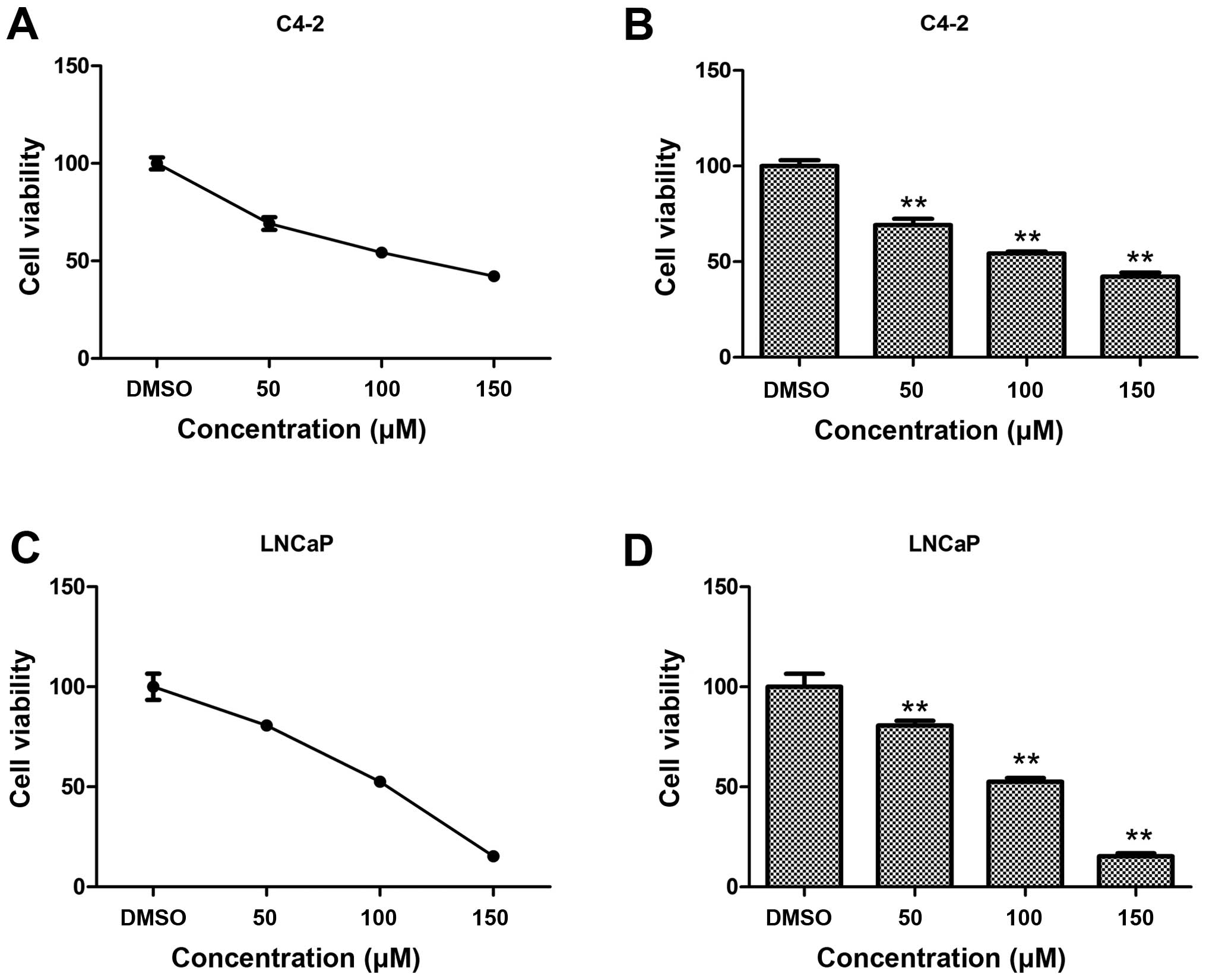
capsaicin at different concentrations for 48 h. Cell viability was
then examined by MTT assay. Capsaicin induced a dose-dependent
inhibition of cell proliferation in both PCa cell lines. At the
concentration of 100 μM, the cell viability of the C4-2 and
LNCaP cells was 53 and 52%, respectively (Fig. 1). The MTT assay showed that
capsaicin inhibited cell proliferation in a dose-dependent manner.
These results demonstrate that capsaicin inhibits the proliferation
of PCa cells.
Capsaicin induces
G0/G1 cell cycle arrest, but has little
influence on apoptosis in AR-positive PCa cells
Anticancer drugs suppress cancer growth mainly
through two modes: apoptosis and cell cycle arrest. According to
previous studies, capsaicin induces apoptosis and cell cycle arrest
leading to cancer growth suppression. Based on this, we further
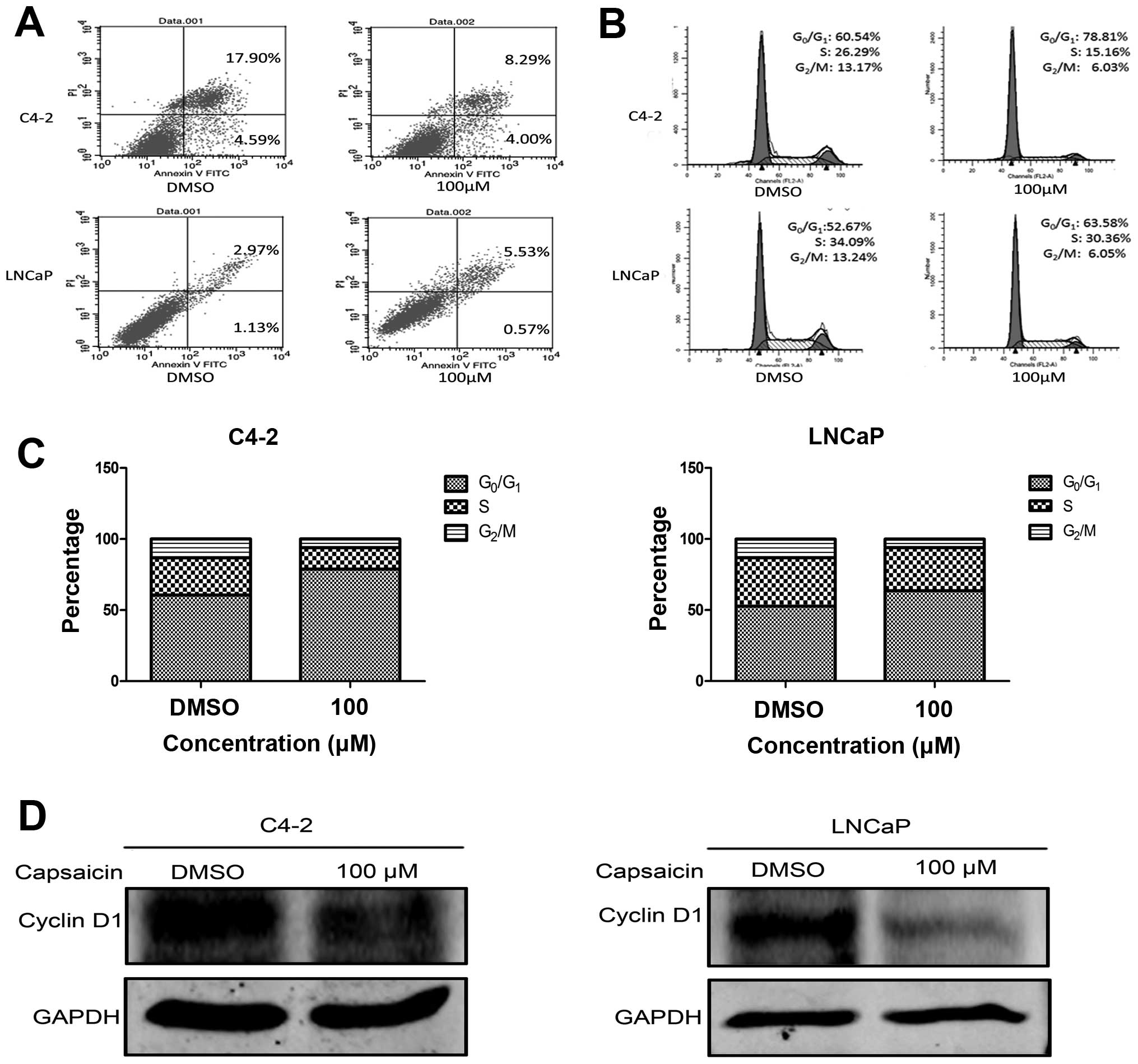
investigated the effects of capsaicin on apoptosis and cell cycle
progression by flow cytometry. C4-2 and LNCaP cells, in a
logarithmic growth phase, were treated with 100 μM capsaicin
for 48 h. As shown in Fig. 2A and
B, the percentage of C4-2 cell apoptosis was 12.29% following
treatment with 100 μM capsaicin for 48 h, compared with the
control group (22.49%). Concerning the LNCaP cells, 100 μM
capsaicin treatment led to a 5.49% cell apoptosis, while the
control group was 4.1%. Thus, capsaicin did not lead to significant
apoptosis in the C4-2 and LNCaP cells. Yet, a significant increase
in the percentage of cells arrested in the
G0/g1 phases in both cell lines was observed
(Fig. 2C). In parallel, there was a
reduction in the percentage of cells in the S and G2/M
phases. To confirm these data, western blot analysis was used to
examine the expression of cyclin D1. Following treatment with
capsaicin, cyclin D1 was obviously reduced both in the C4-2 and
lnCaP cells (Fig. 2D).
Collectively, these results demonstrated that capsaicin affected
the expression of key proteins involved in the cell cycle and
induced G0/g1 cell cycle arrest in the PCa
cells.
Capsaicin inhibits AR activation and
decreases the AR protein level
AR is the key factor of growth regulation and cell
cycle progression in PCa. Jankovic et al reported that
capsaicin delayed the PSA doubling time in a PCa patient who
suffered biochemical failure after radiation (15). It appears that capsaicin may be
associated with the inactivation of AR signaling. Aiming to explore
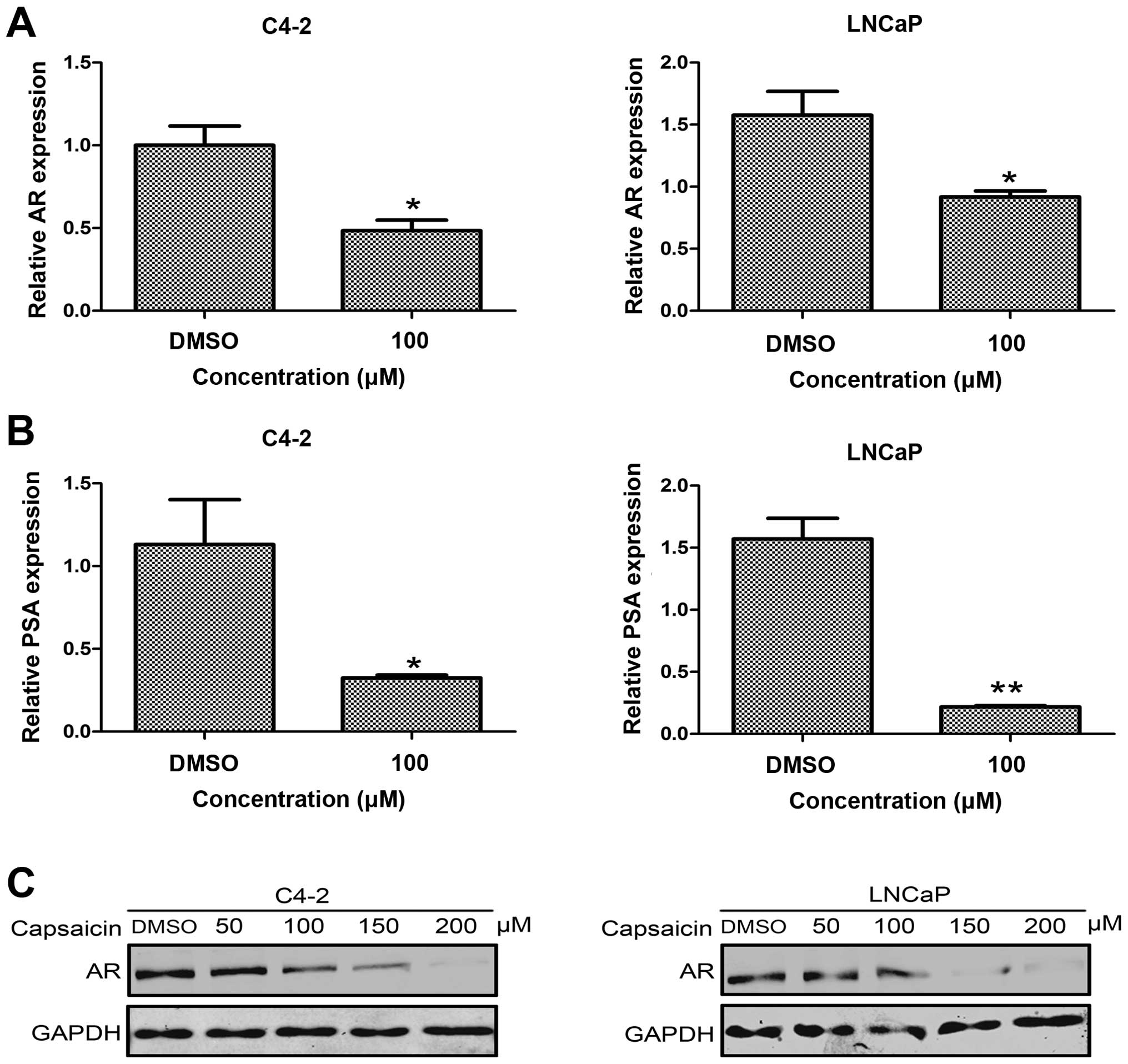
the influence of capsaicin on AR function, we determined the
expression of AR mRNA and protein in the control group and the
capsaicin treatment groups. Compared with the control group,
capsaicin treatment significantly decreased the AR mRNA levels in
the C4-2 and LNCaP cells (Fig. 3A and
C). In addition, the AR protein level was decreased in a
dose-dependent manner following treatment with the indicated
capsaicin concentrations in these two cell lines. In order to
detect the AR function, we measured the expression of PSA, one of
the target genes downstream of AR signaling, by real-time PCR.
After capsaicin treatment for 48 h, PSA expression was
significantly reduced in the C4-2 and LNCaP cells, compared with
the control group (Fig. 3B). Thus,
capsaicin treatment may inhibit AR function and affect the
expression of genes located downstream of AR signaling.
Restoration of miR-449a expression
induced by capsaicin mediates a decrease in AR protein and
inactivation of AR signaling
Previous research has confirmed that capsaicin
regulates the expression of various miRNAs, such as miR-34a
(16) and miR-520-5p (17). Firstly, we detected miR-449a
expression in several AR-positive PCa cell lines. miR-449a was
expressed in all selected AR-positive PCa cells (data not shown).
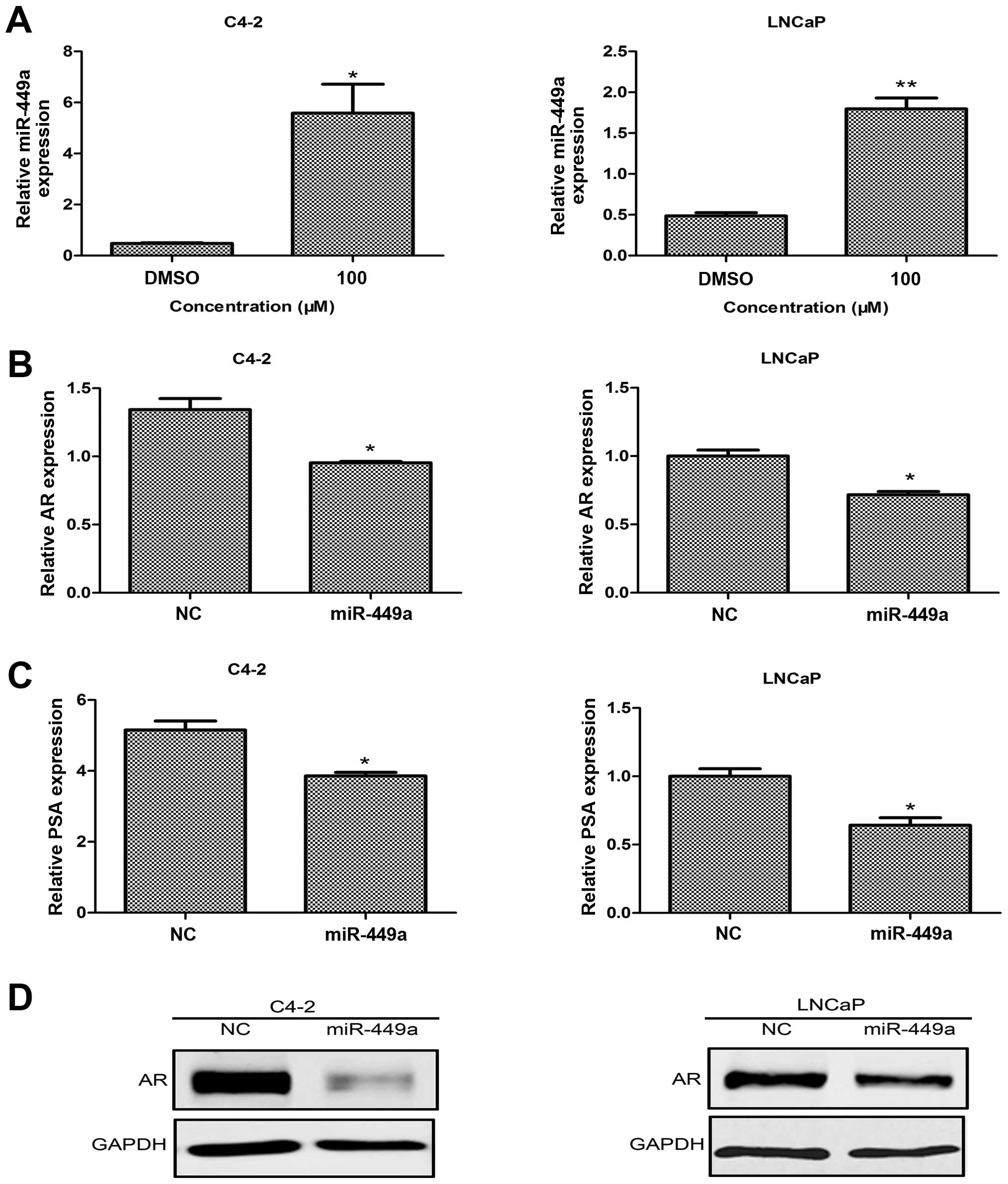
Then, we detected the influence of capsaicin treatment on the
expression of miR-449a. As shown in Fig. 4A, following capsaicin treatment,
miR-449a expression was significantly elevated in both the C4-2 and
LNCaP cells, compared with the control group. As previous studies
have indicated, miR-449a is a potential negative regulator of AR.
Next, we overexpressed miR-449a by a lentivirus to detect whether
or not it functions with AR. Upon overexpression of miR-449a, the
western blot results showed that the level of AR was downregulated
(Fig. 4D), as compared with the
level in the NC group in both cancer cell lines. In addition,
expression of AR and PSA mRNA was inhibited (Fig. 4B and C). All data indicated that
miR-449a mediated a decrease in AR following capsaicin
treatment.
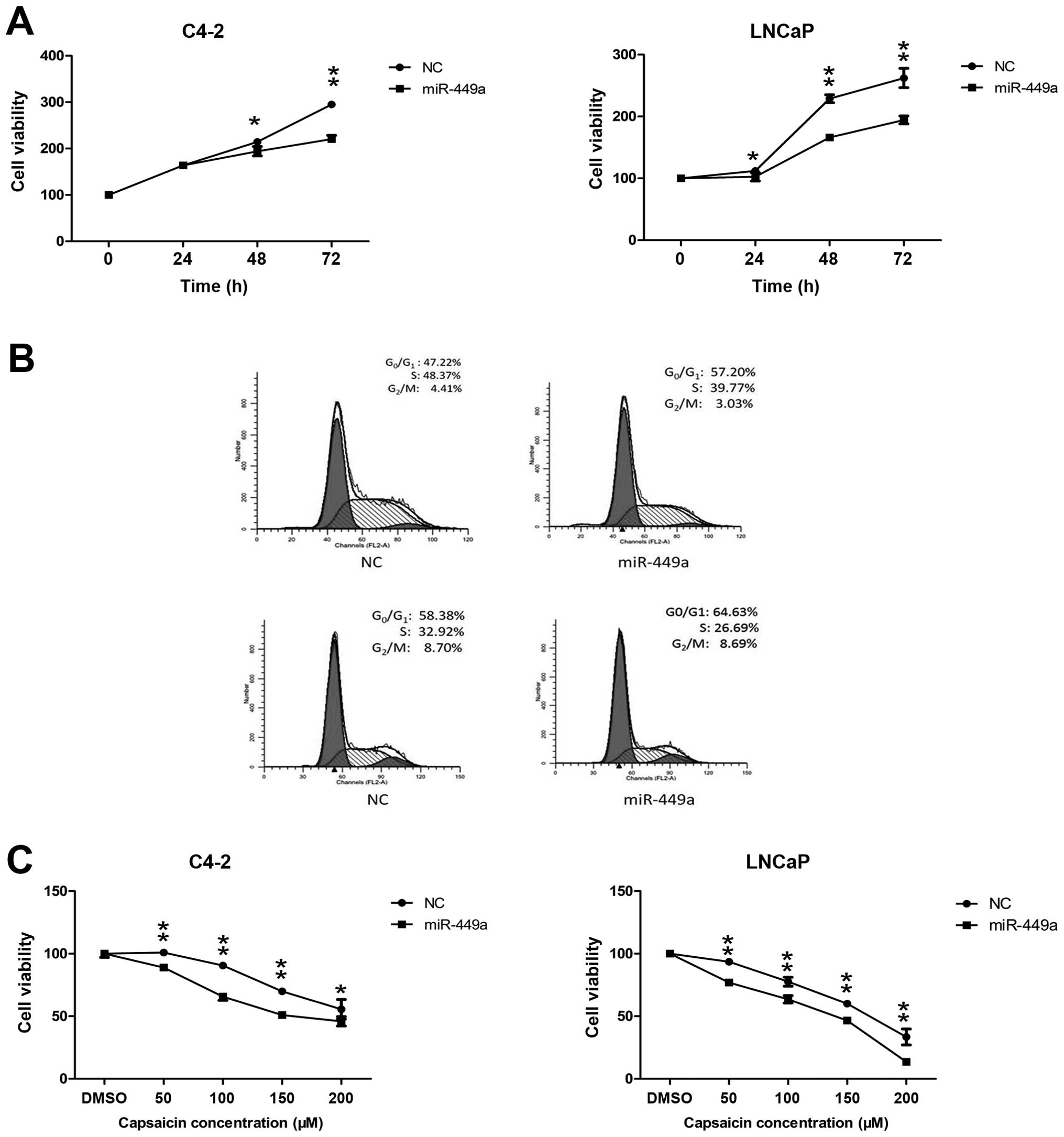
Overexpression of miR-449a inhibits PCa
proliferation and enhances the antitumor effect of capsaicin
Aiming to further verify whether miR-449a has a
growth inhibitory effect on PCa cells, we carried out MTT assay and
cell cycle analysis. Based on the data, we determined that
overexpression of miR-449a inhibited PCa growth in a time-dependent
manner, when compared to the NC group (Fig. 5A). Considering that the cell cycle
may play a role in growth inhibition, we detected cell cycle
progression in the cells transfected with lenti-NC and
lenti-miR449a. miR-449a expression induced
G0/G1 phase arrest in both PCa cell lines
(Fig. 5B). Notably, we found that
miR-449a overexpression enhanced the antitumor effect of capsaicin
treatment (Fig. 5C), indicating
there may be some interactive loop circuits between capsaicin and
miR-449a.
Discussion
Capsaicin, the pungent ingredient of natural
capsicum, has been reported to prevent PCa growth and improve
strategies of PCa therapy in previous studies. Ceramide
accumulation, JNK activation and induction of the endoplasmic
reticulum stress protein gADD153/CHOP were found to be involved in
the mechanisms of apoptosis caused by capsaicin treatment (9,18).
Mori et al found that capsaicin inhibited TNF-α-stimulated
degradation of IκBα and then inactivated NF-κB by preventing its
nuclear migration in PC-3 cells (19). In addition, capsaicin also
functioned as a radio-sensitizing agent through the inhibition of
NF-κB signaling in an athymic mouse model of PCa (20). In the present study, we verified the
antitumor effect of capsaicin on AR-positive PCa cells. Capsaicin
treatment at the indicated concentrations decreased cell viability
in a dose-dependent manner in the C4-2 and LNCaP cells (Fig. 1). The cell cycle is a series of
tightly ordered steps regulated by different checkpoints which
detect extracellular growth signals, protein synthesis and DNA
integrity (21). Aberrant cell
proliferation characteristic of cancer cells is due to the disorder
of the cell cycle. Flow cytometric analysis showed that capsaicin
treatment (100 μM) resulted in G0/G1
phase arrest, but had little impact on cell apoptosis (Fig. 2). Therefore, we confirmed that
capsaicin resulted in growth arrest through arrest of cell cycle
progression, consistent with the findings of other researchers.
Currently, AR remains the most important therapeutic
target for PCa treatment. Anti-androgen drugs, such as casodex,
enzalutamide and flutamide, have been used to cure PCa. However,
clinical therapies that target the androgen-dependent AR signaling
axis eventually fail due to the fact that AR is activated by other
stimuli or signaling pathways. Thus, therapeutic strategies which
promotes AR degradation in castration-resistant PCa have attracted
the research interest of scientists. Researchers have indicated
that promoting the degradation of AR protein shows better antitumor
efficacy. Previous studies demonstrated that ASC-J9, a novel drug
derived from curcumin, enhanced the degradation of AR to suppress
the growth of castration-resistant PCa cells (22–24).
Scolit et al demonstrated that hsp90 function inhibitor,
17-allylamino-17-demethoxygeldanamycin (17-AAG) resulted in a
marked reduction in AR degradation and inhibition of prostate tumor
growth in mice (25). In the
present study, we detected the influence of capsaicin on the AR
protein level and its downstream target gene, PSA (Fig. 3). We found that capsaicin promoted
AR protein degradation and inhibited AR activity in
androgen-independent PCa cells and then inhibited proliferation of
PCa cells. Our data indicated that capsaicin acts as a tumor
suppressor though negative regulation of AR expression and
activation.
Previous research has shown the roles of miRNAs as
novel oncogenes or tumor-suppressor genes and their potential for
PCa diagnosis, prognosis and therapy (10). The function of AR in PCa is related
to expression of miRNAs. Interfering with AR mRNA translation
directly or genes that may regulate AR are main mechanisms of
miRNAs. For example, miR-let-7c was found to suppress AR expression
and activity in human PCa cells via negative regulation of c-Myc
activity (26). Epis et al
demonstrated that miR-331-3p inhibited AR function and blocked AR
signaling via interfering with ERBB-2 expression and AKT activity
in PCa cells (27). In addition, a
similar result was also discovered in PCa with overexpression of
miR-448 (28). In the present
study, we focused on miR-449a. As mentioned previously, miR-449a
plays a role in inducing growth arrest in PCa cells. Östling et
al conducted a systematic screening of 1129 miRnAs in a series
of human PCa cell lines and quantified the changes in AR protein
using a protein lysate microarray. Their data indicated that
miR-449a regulated AR 3′UTR and decreased androgen-induced
proliferation of PCa cells (29).
Our data found that capsaicin treatment significantly increased the
miR-449a level. Further experiments proved that an increased level
of miR-449a downregulated AR protein and decreased the level of PSA
(Fig. 4). Therefore, we
demonstrated that restoration of miR-449a by capsaicin treatment
mediated the degradation of AR and inhibition of AR activity. Then,
we discovered that an increased level of miR-449a prevented
proliferation and arrested the cell cycle in PCa cells. In fact,
capsaicin has been indicated to interact with miRNAs. Triggering
pain signaling by acute noxious stimulation of capsaicin increased
the expression of miR-1 and miR-16 in DRG cells (13). Chakraborty et al found that
increased miR-34a expression by capsaicin decreased the survival
advantage and enhanced Bax-dependent apoptosis in non-small cell
lung cancers (16). Kaymaz et
al proved that capsaicin suppressed the growth of chronic
myelogenous leukemia cells by decreasing expression of miR-520a-5p
which was shown as a probable oncogene (17). They also found that inhibition of
miR520-5p enhanced the antitumor effect of capsaicin. In the
present study, overexpression of miR-449a, a tumor suppressor,
facilitated the growth inhibition of capsaicin treatment (Fig. 5). However, the mechanisms involved
with capsaicin induced restoration of miR-449a needs further
study.
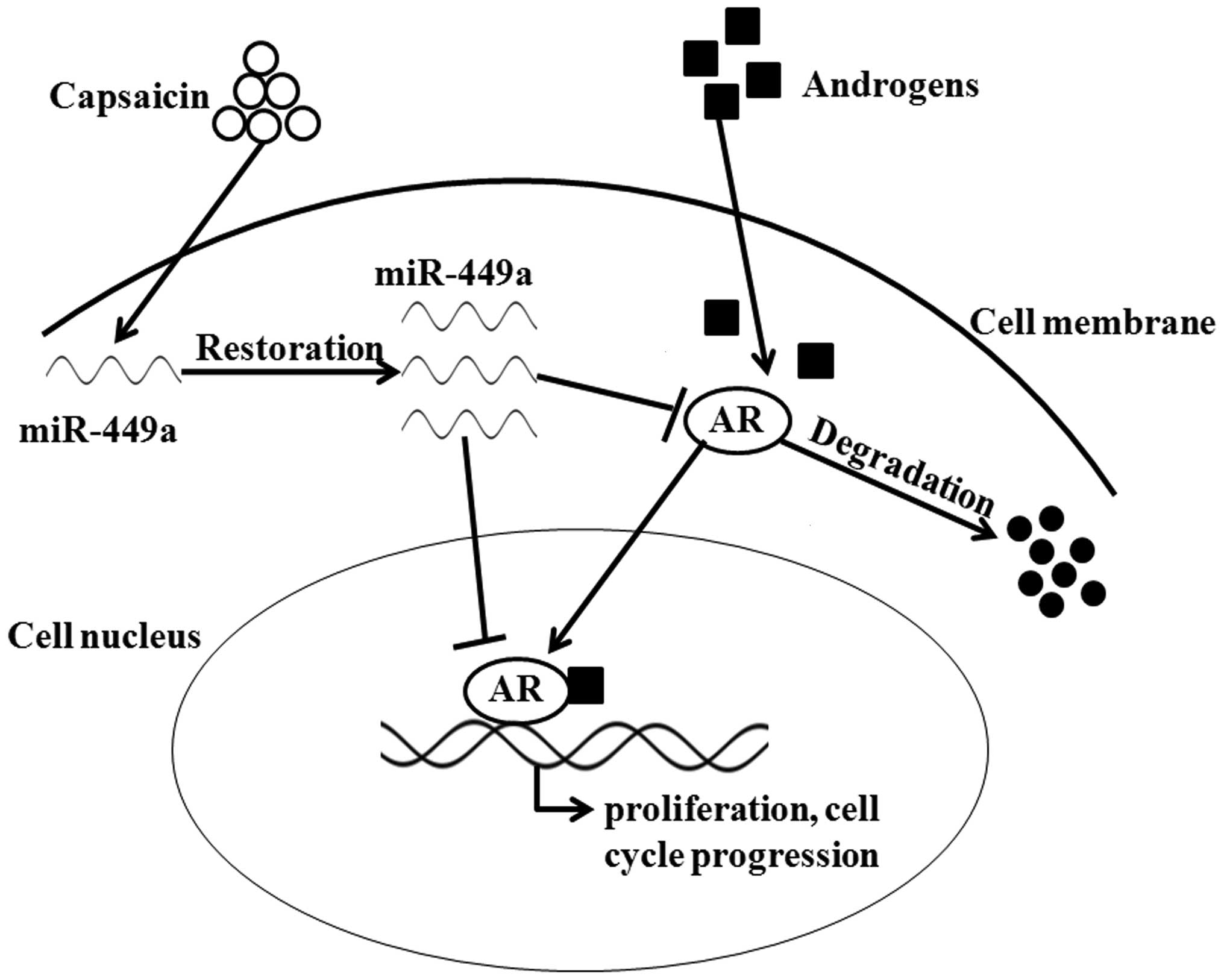
In conclusion, the results from our experiments
clearly demonstrated that capsaicin degrades AR protein and
inactivates AR function leading to decreased growth in AR-positive
PCa cells. Then, we discovered that restoration of miR-449a by
capsaicin mediated the degradation of AR and inactivation of AR
signaling (Fig. 6). The present
study indicated that capsaicin, a biomolecule extracted from
natural capsicum, may be used as a novel drug targeting AR
signaling in PCa treatment.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant no. 81072107, 81472679).
Abbreviations:
|
AR
|
androgen receptor
|
|
PSA
|
prostate-specific antigen
|
|
CRPC
|
castration-resistant prostate
cancer
|
|
miRNA
|
microRNAs
|
|
DMSO
|
dimethyl sulfoxide
|
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Scher HI, Beer TM, Higano CS, Anand A,
Taplin ME, Efstathiou E, Rathkopf D, Shelkey J, Yu EY, Alumkal J,
et al Prostate Cancer Foundation/Department of Defense Prostate
Cancer Clinical Trials Consortium: Antitumour activity of MDV3100
in castration-resistant prostate cancer: A phase 1–2 study. Lancet.
375:1437–1446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Culig Z, Klocker H, Bartsch G and Hobisch
A: Androgen receptors in prostate cancer. Endocr Relat Cancer.
9:155–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heinlein CA and Chang C: Androgen receptor
in prostate cancer. Endocr Rev. 25:276–308. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y, Clegg NJ and Scher HI:
Anti-androgens and androgen-depleting therapies in prostate cancer:
New agents for an established target. Lancet Oncol. 10:981–991.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Montgomery RB, Mostaghel EA, Vessella R,
Hess DL, Kalhorn TF, Higano CS, True LD and Nelson PS: Maintenance
of intratumoral androgens in metastatic prostate cancer: A
mechanism for castration-resistant tumor growth. Cancer Res.
68:4447–4454. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Watson PA, Chen YF, Balbas MD, Wongvipat
J, Socci ND, Viale A, Kim K and Sawyers CL: Constitutively active
androgen receptor splice variants expressed in castration-resistant
prostate cancer require full-length androgen receptor. Proc Natl
Acad Sci USA. 107:16759–16765. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Díaz-Laviada I and Rodriguez-Henche N: The
potential antitumor effects of capsaicin. Progr Drug Res.
68:181–208. 2014.
|
|
9
|
Sánchez AM, Malagarie-Cazenave S, Olea N,
Vara D, Chiloeches A and Díaz-laviada I: Apoptosis induced by
capsaicin in prostate PC-3 cells involves ceramide accumulation,
neutral sphingomyelinase, and JNK activation. Apoptosis.
12:2013–2024. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jansson MD and Lund AH: MicroRnA and
cancer. Mol Oncol. 6:590–610. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Noonan EJ, Place RF, Basak S, Pookot D and
Li LC: miR-449a causes Rb-dependent cell cycle arrest and
senescence in prostate cancer cells. Oncotarget. 1:349–358.
2010.PubMed/NCBI
|
|
12
|
Noonan EJ, Place RF, Pookot D, Basak S,
Whitson JM, Hirata H, Giardina C and Dahiya R: miR-449a targets
HDAC-1 and induces growth arrest in prostate cancer. Oncogene.
28:1714–1724. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kusuda R, Cadetti F, Ravanelli MI, Sousa
TA, Zanon S, De Lucca FL and Lucas G: Differential expression of
microRNAs in mouse pain models. Mol Pain. 7:172011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gill R, Kuriakose R, Gertz ZM, Salloum FN,
Xi L and Kukreja RC: Remote ischemic preconditioning for myocardial
protection: Update on mechanisms and clinical relevance. Mol Cell
Biochem. 402:41–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jankovic B, Loblaw DA and Nam R: Capsaicin
may slow PSA doubling time: Case report and literature review. Can
Urol Assoc J. 4:E9–E11. 2010.PubMed/NCBI
|
|
16
|
Chakraborty S, Mazumdar M, Mukherjee S,
Bhattacharjee P, Adhikary A, Manna A, Chakraborty S, Khan P, Sen A
and Das T: Restoration of p53/miR-34a regulatory axis decreases
survival advantage and ensures Bax-dependent apoptosis of non-small
cell lung carcinoma cells. FEBS Lett. 588:549–559. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaymaz BT, Cetintaş Kaymaz VB, Aktan C and
Kosova B: MicroRNA-520a-5p displays a therapeutic effect upon
chronic myelogenous leukemia cells by targeting STAT3 and enhances
the anticarcinogenic role of capsaicin. Tumour Biol. 35:8733–8742.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sánchez AM, Martínez-Botas J,
Malagarie-Cazenave S, Olea N, Vara D, Lasunción MA and Díaz-Laviada
I: Induction of the endoplasmic reticulum stress protein
gADD153/CHOP by capsaicin in prostate PC-3 cells: A microarray
study. Biochem Biophys Res Commun. 372:785–791. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mori A, Lehmann S, O’Kelly J, Kumagai T,
Desmond JC, Pervan M, McBride WH, Kizaki M and Koeffler HP:
Capsaicin, a component of red peppers, inhibits the growth of
androgen-independent, p53 mutant prostate cancer cells. Cancer Res.
66:3222–3229. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Venier NA, Colquhoun AJ, Sasaki H, Kiss A,
Sugar L, Adomat H, Fleshner NE, Klotz LH and Venkateswaran V:
Capsaicin: A novel radio-sensitizing agent for prostate cancer.
Prostate. 75:113–125. 2015. View Article : Google Scholar
|
|
21
|
Park M-T and Lee S-J: Cell cycle and
cancer. J Biochem Mol Biol. 36:60–65. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamashita S, Lai KP, Chuang KL, Xu D,
Miyamoto H, Tochigi T, Pang ST, Li L, Arai Y, Kung HJ, et al:
ASC-J9 suppresses castration-resistant prostate cancer growth
through degradation of full-length and splice variant androgen
receptors. Neoplasia. 14:74–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin TH, Lee SO, Niu Y, Xu D, Liang L, Li
L, Yeh SD, Fujimoto N, Yeh S and Chang C: Differential androgen
deprivation therapies with anti-androgens casodex/bicalutamide or
MDV3100/Enzalutamide versus anti-androgen receptor ASC-J9(R) lead
to promotion versus suppression of prostate cancer metastasis. J
Biol Chem. 288:19359–19369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin TH, Izumi K, Lee SO, Lin WJ, Yeh S and
Chang C: Anti-androgen receptor ASC-J9 versus anti-androgens
MDV3100 (Enzalutamide) or Casodex (Bicalutamide) leads to opposite
effects on prostate cancer metastasis via differential modulation
of macrophage infiltration and STAT3-CCL2 signaling. Cell Death
Dis. 4:e7642013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Solit DB, Zheng FF, Drobnjak M, Münster
PN, Higgins B, Verbel D, Heller G, Tong W, Cordon-Cardo C, Agus DB,
et al: 17-Allylamino-17-demethoxygeldanamycin induces the
degradation of androgen receptor and HER-2/neu and inhibits the
growth of prostate cancer xenografts. Clin Cancer Res. 8:986–993.
2002.PubMed/NCBI
|
|
26
|
Nadiminty N, Tummala R, Lou W, Zhu Y,
Zhang J, Chen X, eVere White RW, Kung HJ, Evans CP and Gao AC:
MicroRNA let-7c suppresses androgen receptor expression and
activity via regulation of Myc expression in prostate cancer cells.
J Biol Chem. 287:1527–1537. 2012. View Article : Google Scholar :
|
|
27
|
Epis MR, Giles KM, Barker A, Kendrick TS
and Leedman PJ: miR-331–3p regulates ERBB-2 expression and androgen
receptor signaling in prostate cancer. J Biol Chem.
284:24696–24704. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sikand K, Slaibi JE, Singh R, Slane SD and
Shukla GC: miR 488* inhibits androgen receptor
expression in prostate carcinoma cells. Int J Cancer. 129:810–819.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Östling P, Leivonen SK, Aakula A, Kohonen
P, Mäkelä R, Hagman Z, Edsjö A, Kangaspeska S, Edgren H, Nicorici
D, et al: Systematic analysis of microRNAs targeting the androgen
receptor in prostate cancer cells. Cancer Res. 71:1956–1967. 2011.
View Article : Google Scholar : PubMed/NCBI
|