Introduction
Osteosarcoma is the most common primary malignant
tumor of the bone predominantly occurring in childhood and
adolescence. Among individuals younger than 20 years of age, the
osteosarcoma incidence rate is 8.7 per million and the risk is
higher in males (1). It usually
occurs in a growing long bone such as the humerus, femur or tibia.
The current treatment for osteosarcoma is neoadjuvant chemotherapy,
surgical resection and chemotherapy again following surgery
(2). Despite the advances in
treatment options, recurrence and chemoresistance have been the
main challenges confronting physicians (3–5). In
patients with localized osteosarcoma, the 5-year survival rates are
~65–75% (6,7). However, the prognosis for patients
with recurrence and metastases is quite poor, with 5-year survival
rates ranging from 15 to 30% (8).
In addition, most neoadjuvant chemotherapy drugs carry the risk of
uncertain effectiveness and severe side effects, and multidrug
resistant cases are common, particularly with cisplatin and
doxorubicin (9). Over the past 35
years, there has been no significant improvement in chemotherapy
for osteosarcoma (10). Therefore,
a better understanding of the molecular mechanisms involved in
osteosarcoma progression should help to explore novel therapeutic
targets or develop new modalities of osteosarcoma therapy.
Recently, research into osteosarcoma treatment has been focused on
novel target therapies including induction of apoptosis and
reduction of cell growth in osteogenic sarcoma (11–13).
Traditional Chinese medicine is an important type of
complementary and alternative medicine, because it has a
standardized system of diagnostics and therapies, and is practised
worldwide (14). Some plant
extracts used in complementary medicine, exert potent anticancer
activity with low toxicity. Celastrol was found to inhibit growth
and accelerate apoptosis in many human cancer cell lines such as
hepatoma, breast, myeloma, pancreas and gastric cancer cell lines
(15–19).
Apoptosis is a strictly controlled mechanism of cell
suicide that is triggered by certain internal or external signals.
It results in cell rounding and shrinkage, chromatin condensation,
DNA fragmentation and shedding of smaller fragments from cells. In
the intrinsic pathway, mitochondria play a central role in the
occurrence of apoptosis induced by many chemotherapeutic agents
(20–22). Mitochondrial membrane
permeabilization, along with the collapse of electrochemical
gradient across the mitochondrial membrane leads to the release of
catabolic hydrolases and activators of some enzymes from the
mitochondria, resulting in cell apoptosis (23,24).
Bcl-2 family proteins serve as crucial regulators of this pathway
through their influence on mitochondrial outer membrane
permeabilization (MOMP) following homo- or hetero-association
(25,26). Among Bcl-2 family proteins,
pro-apoptotic proteins such as Bax, Bad and Bid, increase MOMP
during apoptosis and release apoptogenic proteins into the cytosol,
such as cytochrome c, which can bind to Apaf-1 and further
activate caspase-9. Furthermore, activated caspase-9 activates
downstream caspase-3 and/or -7, which in turn results in the
cleavage or degradation of several key cellular substrates,
including PARP, thus leading to apoptosis (27–30).
However, anti-apoptotic proteins such as Bcl-2 and
Bcl-XL, can bind to activated Bax to decrease membrane permeability
(31). The regulation of activated
anti- and pro-apoptotic Bcl-2 family members is essential for
determining the fate of cells, and disturbance of the normal
apoptotic program due to alteration of the ratio by aberrant
expression of these proteins may cause various apoptosis-related
diseases (32,33). In addition, Bcl-2 and Bcl-XL
overexpression, widely observed in various types of cancers,
inhibits apoptosis and confers resistance to anticancer drugs
(34,35). Therefore, induction of apoptosis
through the mitochondrial-dependent pathway has been one of the
targets of anticancer chemotherapy.
Materials and methods
Materials and reagents
Dulbecco's modified Eagle's medium (DMEM), fetal
bovine serum (FBS), phosphate-buffered saline (PBS) and dimethyl
sulphoxide (DMSO) were provided by Transgen (Beijing, China). A
Hoechst 33258 staining kit was provided by Keygen Biotech (Nanjing,
China). MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide] was obtained from Solarbio (Beijing, China). Antibodies
against Bcl-2, Bax, caspase-3, caspase-8, caspase-9 and β-actin
were purchased from Abcam (Cambridge, UK), and antibodies against
PARP and cytochrome c were purchased from Cell Signaling
Technology (Beverly, MA, USA). Horseradish peroxidase
(HRP)-conjugated secondary antibodies were purchased from Cell
Signaling Technology and Transgen. An Annexin V-PE/7-AAD apoptosis
detection kit was provided by Becton-Dickinson (San Jose, CA, USA).
Celastrol was obtained from Nanjing Zelang Medical Technology Co.,
ltd. (Nanjing, China). Stock solutions of celastrol were prepared
by dissolving the celastrol powder in DMSO to a concentration of 1
M, and stored at −20°C. The working concentrations of celastrol
were made by diluting the stock solution with the culture medium.
The final concentration of DMSO in the medium was <0.5%.
Cell culture
Human osteosarcoma cell lines, MG-63 (wild-type),
U-2OS (wild-type) and HOS (wild-type), were obtained from the
American Type Culture Collection (ATCC; Manassas, VA, USA). Cells
were cultured in DMEM supplemented with 10% (v/v) FBS, 100 U/ml
penicillin and 100 µg/ml streptomycin. They were all placed
in a humidified atmosphere containing 5% CO2 at 37°C.
The cells used were subjected to <20 cell passages and were in
the logarithmic growth phase.
Cell viability by MTT assay
The cells were cultured in 96-well plates at a
concentration of 1×104 cells/well. Cell viability was
determined using an MTT colorimetric assay. The cells were treated
with celastrol at various final concentrations (0.5, 1, 2, 4 and 6
µM, respectively), for 24, 36 and 48 h, and the control
cells were treated with 0.5% DMSO. After the indicated cultivation
time, 50 µl of MTT (5 mg/ml in PBS) was added and the plates
were incubated at 37°C for an additional 4 h. Finally, the formazan
precipitate was dissolved in 100 µl DMSO and the cells were
shaken for 10 min. Absorbance was measured at 490 nm using a
Universal microplate reader (EL800; BioTek Instruments Inc.). ELISA
reader (BioTek, Model EXL800; USA). Cell growth expressed as
percent viability was calculated by comparing the absorbance of
treated vs. untreated cells.
Hoechst 33258 staining of U-2OS
cells
Cells were incubated with 0, 1, 2.5 and 4 µM
of celastrol for 48 h, harvested, fixed with 4% paraformaldehyde
for 30 min at 25°C, washed 3 times with ice-cold PBS and stained
with 10 mg/l Hoechst 33258 (Sigma) for 10 min in the dark at room
temperature. Finally, the stained nuclei were observed under a
fluorescence microscope (Olympus, ×100) with excitation at 350 nm
and emission at 460 nm.
Analysis of cell apoptosis by Annexin
V-PE/7-AAD staining assay
To assess the development of apoptosis induced by
celastrol, U-2OS cells were stained with Annexin V-PE/7-AAD (BD
Biosciences, San Jose, CA, USA). U-2OS cells (1×105)
were cultured in 12-well plates. Following overnight incubation,
these cells were treated with celastrol at various concentrations
for 48 h and collected by trypsinization, not containing EDTA.
After being twice washed with 4°C PBS, the cell pellets were
suspended again in 400 µl ice-cold 1X binding buffer at a
density of nearly 1×106 cells/ml, and then incubated
with 10 µl Annexin V-PE/7-AAD for 10 min in the dark at room
temperature. Samples were analyzed by a flow cytometer within 1 h
of staining.
Western blot analysis
U-2OS cells were cultured in 6-well plates at a
concentration of 2×105 cells/well. After treatment with
celastrol at various concentrations for 48 h, the cells were
collected and lysed in RIPA buffer containing a protease inhibitor
cocktail (Sigma Chemical, USA). The homogenates were centrifuged at
12,000 rpm for 10 min at 4°C and the supernatant fraction was
collected for immunoblotting. Furthermore, protein concentrations
were calculated by a BCA assay using bovine serum albumin as the
standard. The same amounts of proteins were loaded and separated by
electrophoresis on 12% SDS-polyacrylamide gels under a reducing
condition using 100 V for 2 h. After electrophoresis, the proteins
were transferred to PVDF membranes in a Tris-glycine transfer
buffer using a semi-dry blotting system, and incubated with
antibodies against Bcl-2, Bax, cytochrome c, PARP,
caspase-3, caspase-8, caspase-9 and β-actin (1:1,000) overnight at
4°C. After PVDF membranes were washed in TBST 3 times, secondary
HRP-conjugated antibodies were added at 1:2,000 dilution for 1 h at
room temperature and the PVDF membranes were washed again in TBST 3
times. Immunoreactive proteins were detected by enhanced
chemiluminescence (ECL kit; Transgen) followed by exposure to X-ray
film.
Statistical analysis
Data were analyzed using the SPSS package for
Windows (version 17.0). Quantitative data are expressed as the mean
± standard deviation (SD). Statistical analysis of the data was
performed using a student's t-test and ANOVA. P-values of <0.05
were considered to be statistically significant.
Results
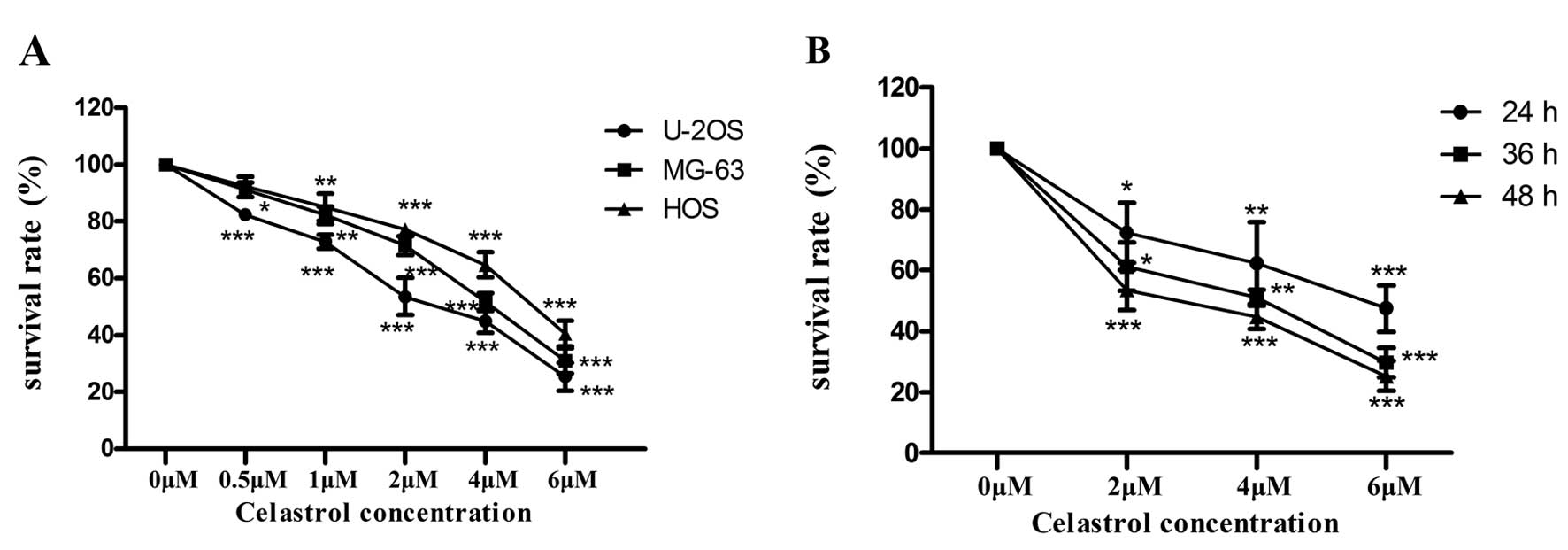
Celastrol reduces the viability of U-2OS
cells
The effect of celastrol on the viability of
osteosarcoma cell lines was determined by MTT assay, and we treated
three human osteosarcoma cell lines (MG-63, U-2OS and HOS) with
celastrol at different concentrations for 24, 36 and 48 h,
respectively. As shown in Fig. 1A,
the inhibitory effects of celastrol on the human osteosarcoma cell
lines were dose-dependent, but each cell line exhibited a different
sensitivity to celastrol. Obviously, the U-2OS cells were the most
sensitive to celastrol. The IC50 value for the U-2OS
cells treated with celastrol was 2.5 µM at 48 h. As shown in
Fig. 1B, the inhibitory effects of
celastrol on the human osteosarcoma cell lines was time-dependent.
Furthermore, U-2OS cells were treated with celastrol at the
concentrations of 0, 1, 2.5 and 4 µM for 48 h in the
following assays. Our findings demonstrated that celastrol
inhibited cellular proliferation in a time- and dose-dependent
manner.
Induction of morphological changes of
U-2OS cells
Untreated U-2OS cells grew well as observed by phase
contrast microscopy. After 48 h of treatment, celastrol produced
broken, necrotic and detached cells in a dose-dependent manner,
which was consistent with the growth inhibition. Celastrol-treated
U-2OS cells stained with the fluorescent DNA-binding dye Hoechst
33258 revealed condensed and fragmented nuclei, which are typical
morphological features of apoptotic cells. In contrast, no
morphological signs of apoptosis were observed in the untreated
cells. The results indicated that cell death occurred through
apoptosis (Fig. 2).
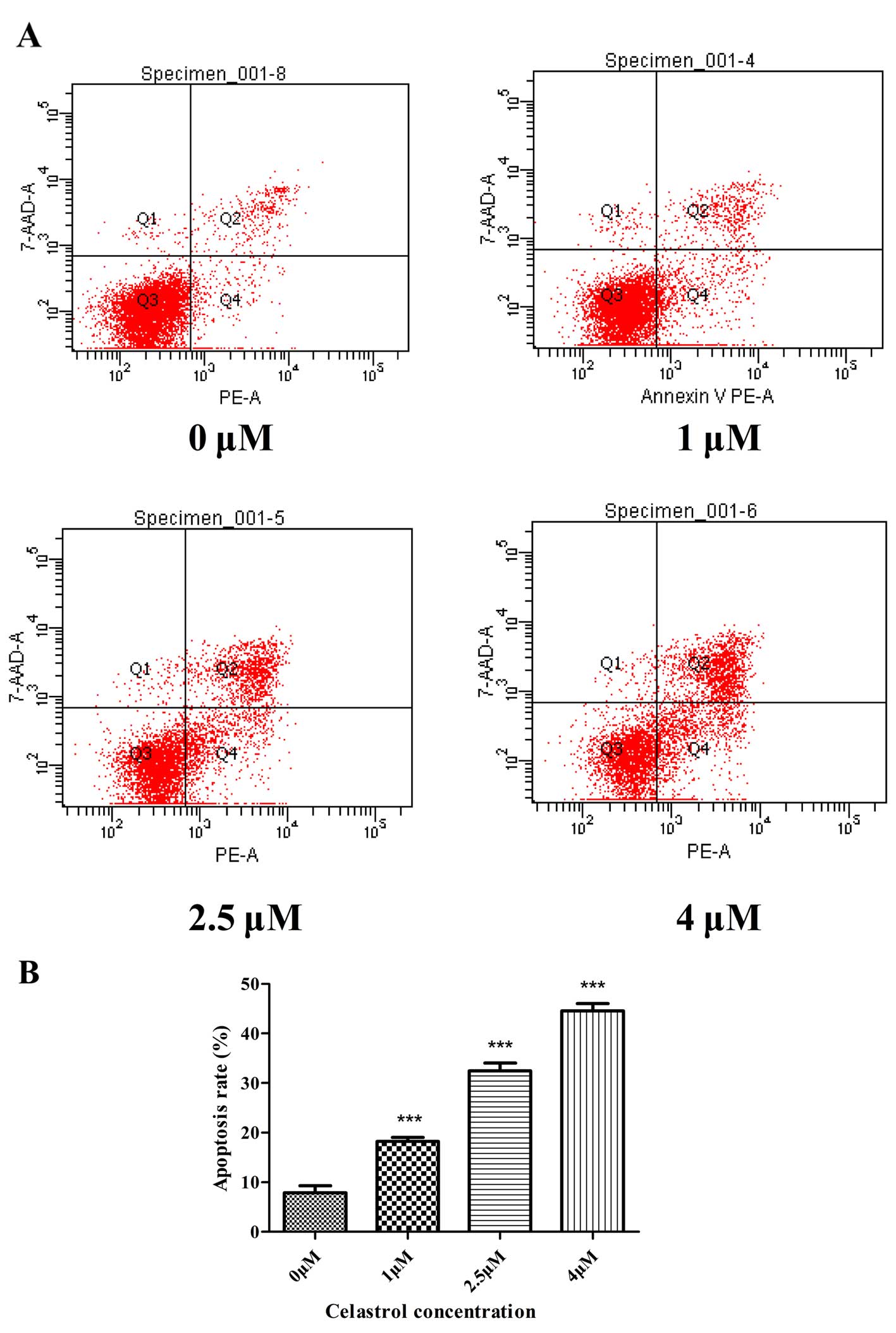
Annexin V-PE/7-AAD staining assay
The rate of cell apoptosis was detected by flow
cytometry following double labeling with Annexin V-PE/7-AAD.
Representative graphs obtained by flow cytometric analysis of the
cells treated with celastrol at different concentrations for 48 h
after double staining with Annexin V-PE and 7-AAD are shown in
Fig. 3A. The apoptosis rate in the
control cells was 7.9±1.4%. There was a dose-dependent increase in
the apoptosis rate of U-2OS cells treated with celastrol. The
apoptosis rates in the U-2OS cells were increased to 18.2±0.8,
32.5±1.6 and 44.6±1.4% following treatment with celastrol at 1, 2.5
and 4 µM for 48 h, respectively (Fig. 3B).
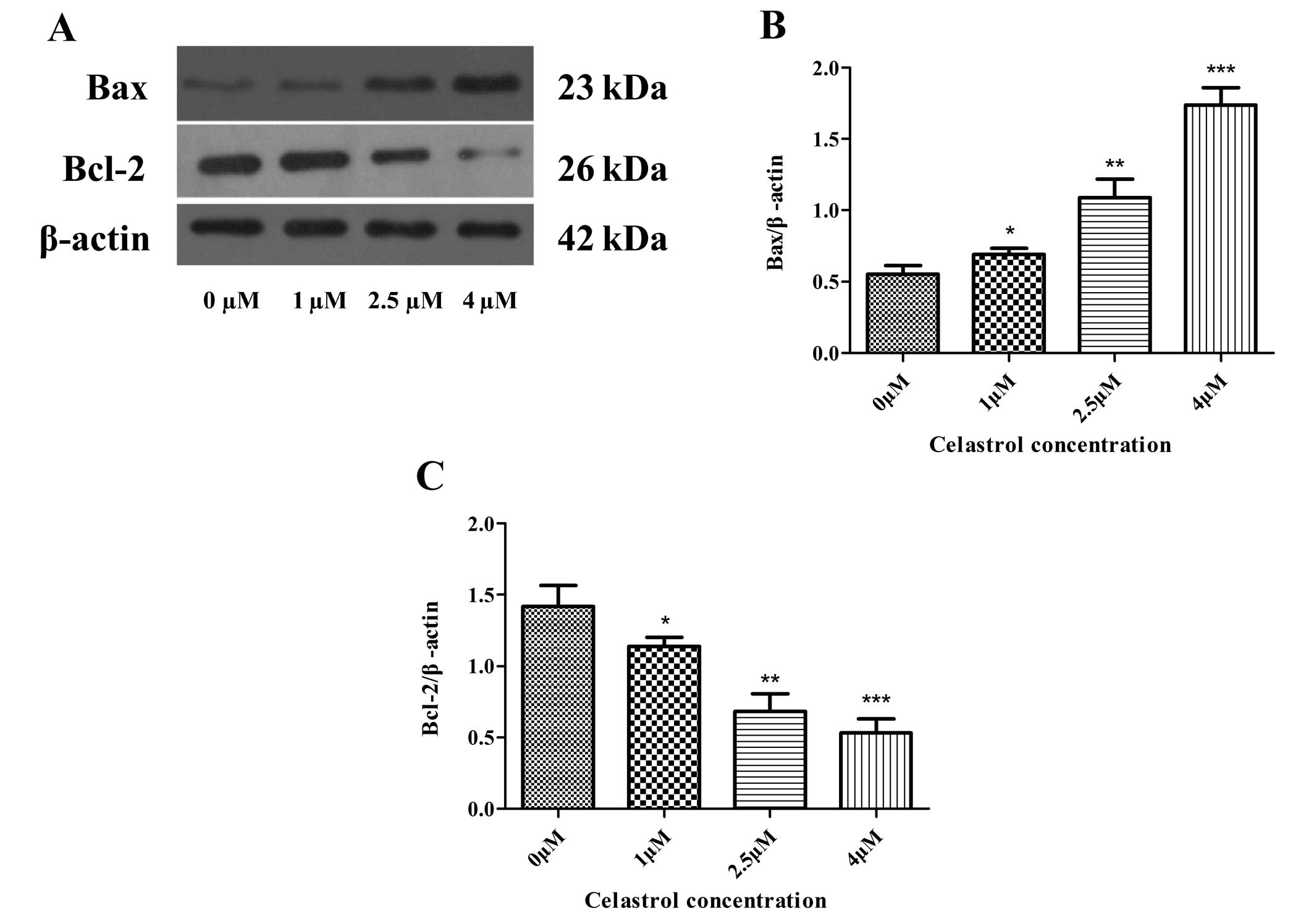
Celastrol decreases the expression of
anti-apoptotic Bcl-2 and increases the expression of pro-apoptotic
Bax and cytochrome c
To determine the molecular mechanism by which
celastrol induces the apoptosis of U-2OS cells, the protein
expression levels of Bcl-2 family proteins, including
anti-apoptotic members such as Bcl-2, and pro-apoptotic members
such as Bax and cytochrome c, were assessed by performing
western blot analysis. The results of the western blot analysis
revealed that celastrol treatment caused a profoundly marked
increase in Bax proteins and the release of cytochrome c,
and a decrease in Bcl-2 protein, when compared to these levels in
the control (Figs. 4A–C and
5A and B). This demonstrates that
celastrol activates the mitochondrial apoptotic pathway in U-2OS
cells via regulating the expression of the Bcl-2 family
proteins.
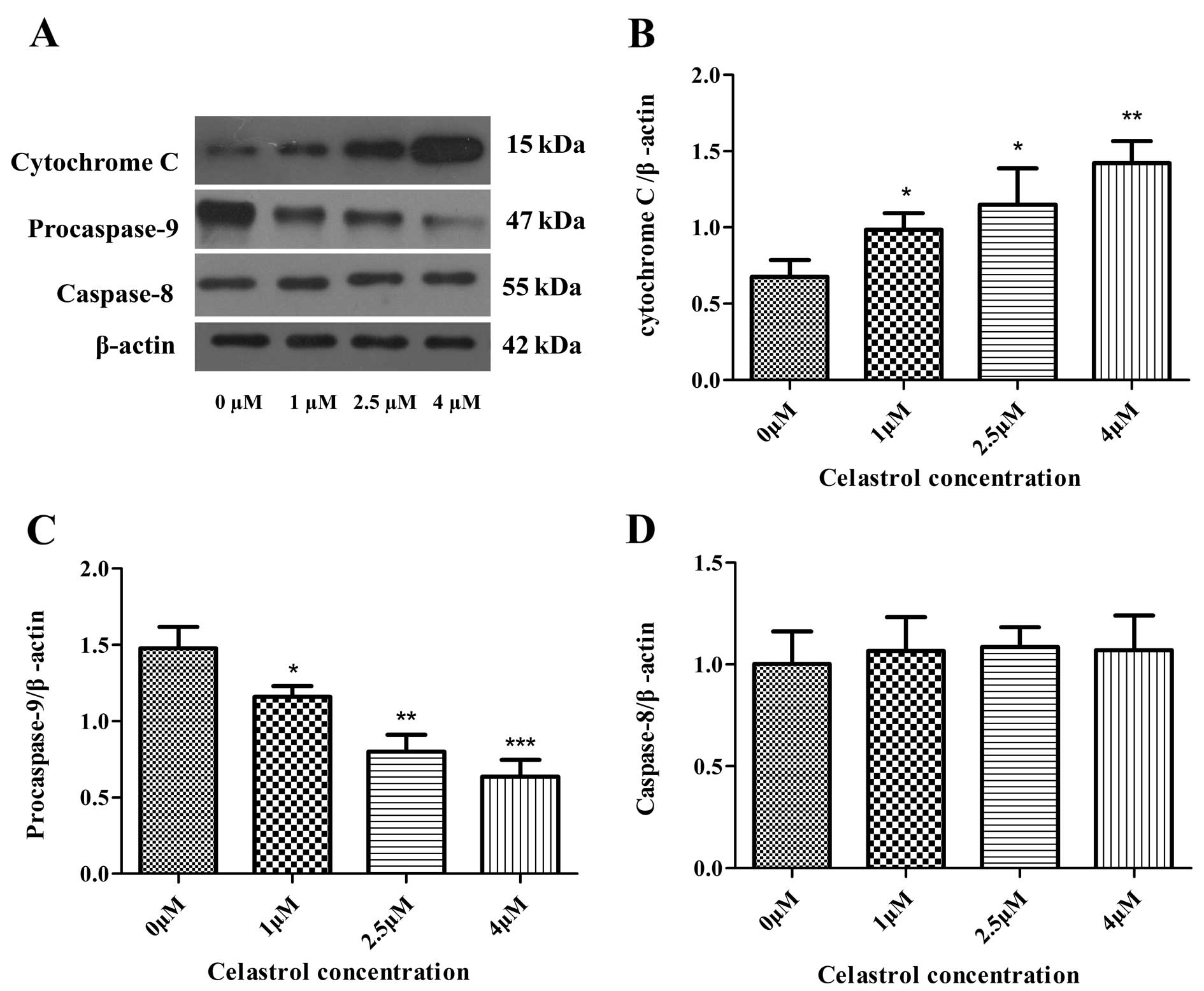
Effects of celastrol on the expression
levels of caspases
The caspase cascade reaction is one of the most
important events in the process of apoptosis through the
mitochondrial pathway. Therefore, the protein expression levels of
caspase-3, -8 and procaspase-9 were assessed by performing western
blot analysis (Figs. 5A and
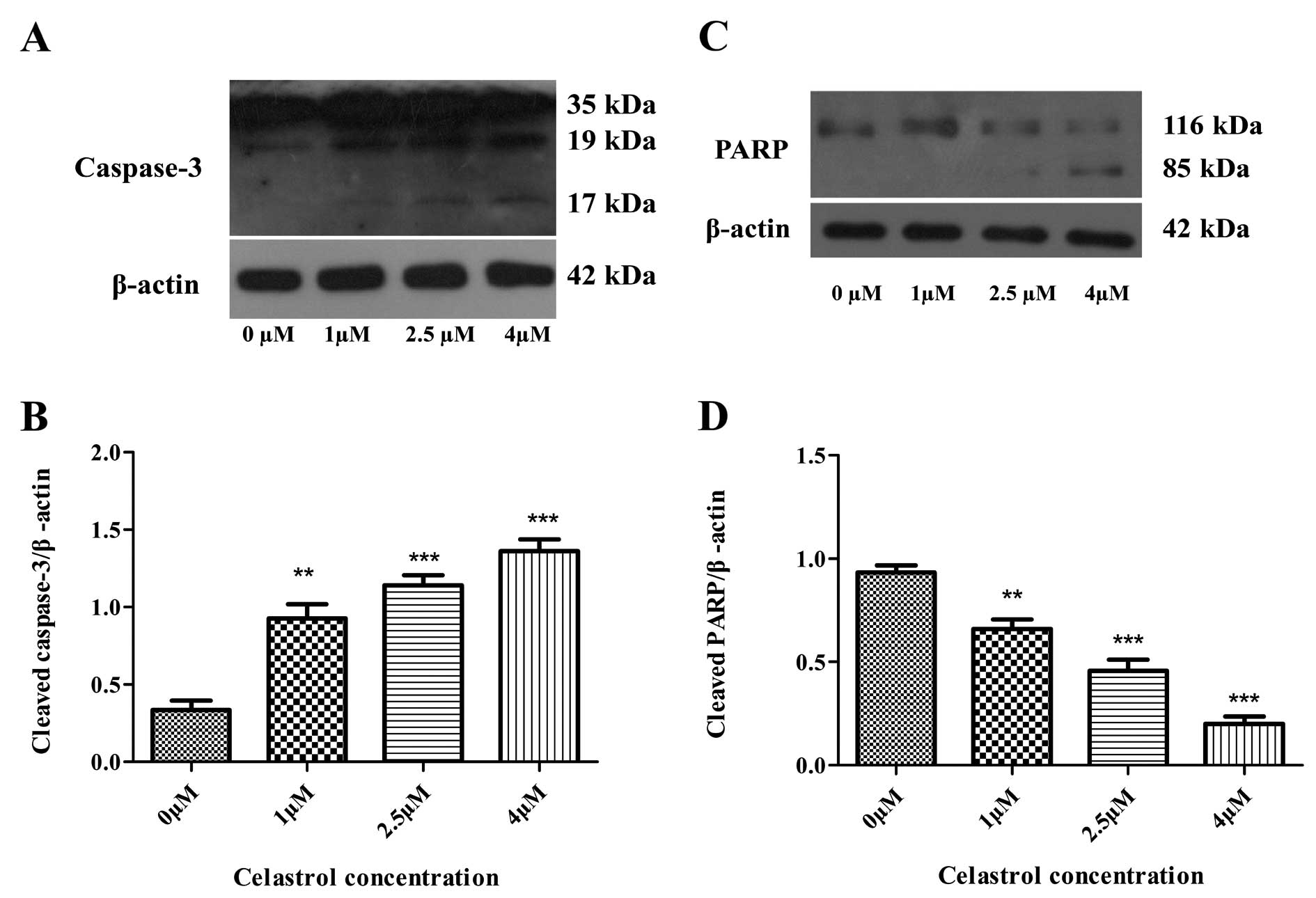
6A). Caspase-3 cleavage was
observed (Fig. 6A and B), and
expression levels of procaspase-9 were downregulated, both in a
concentration-dependent manner as the concentration of celastrol
increased (Fig. 5A and C). However,
expression levels of caspase-8 were not changed in the cells
treated with celastrol (Fig. 5A and
D). Cleavage of PARP, a key cellular substrate, was observed
(Fig. 6C and D). The results
indicated that the apoptosis induced by celastrol involved the
caspase cascade and was triggered through the mitochondrial
pathway.
Discussion
Apoptosis, a program of cell suicide, is an innate
cellular response to eliminate abnormal or redundant cells in
mammals and hence is considered an important mechanism in the
action of many anticancer drugs (22). There is accumulating evidence that a
wide variety of herbal medicines and compounds extracted from
natural products with antitumor effects can trigger apoptosis in
various tumor cells (20–22). Previous studies have demonstrated
that celastrol, a triterpene extracted from the root bark of
Triptergium wilfordii Hook F., also known as 'Thunder of God
Vine, can inhibit tumor promotion (15–19).
In the present study, we determined the anticancer effect and
associated mechanisms of celastrol on human osteosarcoma cells
lines in vitro. MTT results revealed that celastrol
effectively suppressed the proliferation of three human
osteosarcoma cell lines (MG-63, U-2OS and HOS) in a dose- and
time-dependent manner. FACS analysis showed that celastrol
effectively induced apoptosis in the osteosarcoma cells. Thus, we
next investigated the apoptotic mechanism of celastrol on
osteosarcoma cells.
Apoptosis is triggered by two different signals: the
mitochondrial pathway and the cell death receptor pathway,
regulated via caspase-9 and -8, respectively (36). Accumulated evidence has shown that
caspases play critical roles in the apoptotic cascade. In the
mitochondrial pathway (the intrinsic pathway), downstream of
caspase activation is regulated by members of the Bcl-2 family.
Apoptosis-associated MOMP is known to require pro-apoptotic
Bax-like proteins, in the regulation of pore formation in
mitochondria. Anti-apoptotic Bcl-2-like proteins in mitochondrial
morphogenesis are functionally distinct from their role in
apoptosis. Therefore, the ratio of Bax to Bcl-2 is vital for
determining the release of many apoptogenic proteins from the
mitochondrial intermembrane space, such as cytochrome c
which can further activate caspase-9. Activated caspase-9 then
activates downstream caspase-3, which causes the cleavage or
degradation of various key cellular substrates, including PARP,
thus resulting in apoptosis (27–30,37–39).
The cell death receptor pathway (the extrinsic pathway) activates
the death receptor on the cell surface (Fas/FasL) and then promotes
caspase-8 activation (40–42). With this in mind, to demonstrate
which signaling pathway is involved in apoptosis by celastrol,
expression of Bcl-2 family proteins, caspase-3, -8 and -9 and PARP
were assessed in U-2OS cells. The present data showed that
celastrol-induced apoptosis was accompanied by alteration of the
Bax/Bcl-2 ratio and activation of caspase-3 and -9, but not of
caspase-8. Furthermore, cleavage of PARP was also observed. These
findings indicated that celastrol-induced apoptosis in U-2OS cells
was triggered by an intrinsic pathway.
In conclusion, we demonstrated that celastrol
dose-dependently upregulates Bax expression and downregulates Bcl-2
expression in U-2OS cells. This results in the release of
cytochrome c into the cytosol, which further activates
caspase-9. Furthermore, activated caspase-9 activates downstream
caspase-3 which in turn, results in the cleavage or degradation of
several key cellular substrates, including PARP, and leads to the
subsequent apoptosis. These results indicated that celastrol could
be a potential novel therapeutic agent for the treatment of
osteosarcoma. Further studies are required in order to ascertain
whether celastrol can synergize with other chemotherapy drugs. In
addition, studies on the in vivo effect of celastrol on
U-2OS xenograft tumors in nude mice are in progress.
Acknowledgments
This study was supported by the Natural Science
Foundation of Jiangxi Province (20132BAB205081), the Foundation of
the Health Department of Jiangxi Province on Traditional Chinese
Medicine (2012A136) and the Engineering Technology Research Center
Construction Project of Jiangxi Province (20132BCD40026).
References
|
1
|
Mirabello L, Troisi RJ and Savage SA:
International osteosarcoma incidence patterns in children and
adolescents, middle ages and elderly persons. Int J Cancer.
125:229–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Longhi A, Errani C, De Paolis M, Mercuri M
and Bacci G: Primary bone osteosarcoma in the pediatric age: State
of the art. Cancer Treat Rev. 32:423–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chou AJ and Gorlick R: Chemotherapy
resistance in osteosarcoma: Current challenges and future
directions. Expert Rev Anticancer Ther. 6:1075–1085. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schwartz CL, Gorlick R, Teot L, Krailo M,
Chen Z, Goorin A, Grier HE, Bernstein ML and Meyers P; Children's
Oncology Group: Multiple drug resistance in osteogenic sarcoma:
INT0133 from the Children's Oncology Group. J Clin Oncol.
25:2057–2062. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wafa H and Grimer RJ: Surgical options and
outcomes in bone sarcoma. Expert Rev Anticancer Ther. 6:239–248.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oertel S, Blattmann C, Rieken S, Jensen A,
Combs SE, Huber PE, Bischof M, Kulozik A, Debus J and Schulz-Ertner
D: Radiotherapy in the treatment of primary osteosarcoma - a single
center experience. Tumori. 96:582–588. 2010.PubMed/NCBI
|
|
7
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the Surveillance, Epidemiology, and End Results Program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Picci P, Mercuri M, Ferrari S, Alberghini
M, Briccoli A, Ferrari C, Pignotti E and Bacci G: Survival in
high-grade osteosarcoma: Improvement over 21 years at a single
institution. Ann Oncol. 21:1366–1373. 2010. View Article : Google Scholar
|
|
9
|
Arai K, Sakamoto R, Kubota D and Kondo T:
Proteomic approach toward molecular backgrounds of drug resistance
of osteosarcoma cells in spheroid culture system. Proteomics.
13:2351–2360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Robert RS, Ottaviani G, Huh WW, Palla S
and Jaffe N: Psychosocial and functional outcomes in long-term
survivors of osteosarcoma: A comparison of limb-salvage surgery and
amputation. Pediatr Blood Cancer. 54:990–999. 2010.PubMed/NCBI
|
|
11
|
Shangguan WJ, Li H and Zhang YH: Induction
of G2/M phase cell cycle arrest and apoptosis by ginsenoside Rf in
human osteosarcoma MG 63 cells through the mitochondrial pathway.
Oncol Rep. 31:3051–313. 2014.
|
|
12
|
Li Z, Yu Y, Sun S, Qi B, Wang W and Yu A:
Niclosamide inhibits the proliferation of human osteosarcoma cell
lines by inducing apoptosis and cell cycle arrest. Oncol Rep.
33:1763–1768. 2015.PubMed/NCBI
|
|
13
|
Wei J, Zhu Y, Xu G, Yang F, Guan Z, Wang M
and Fang Y: Oxymatrine extracted from Sophora flavescens inhibited
cell growth and induced apoptosis in human osteosarcoma MG-63 cells
in vitro. Cell Biochem Biophys. 70:1439–1444. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ahmed S, Anuntiyo J, Malemud CJ and Haqqi
TM: Biological basis for the use of botanicals in osteoarthritis
and rheumatoid arthritis: A review. Evid Based Complement Alternat
Med. 2:301–308. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li PP, He W, Yuan PF, Song SS, Lu JT and
Wei W: Celastrol induces mitochondria-mediated apoptosis in
hepatocellular carcinoma Bel-7402 cells. Am J Chin Med. 43:137–148.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mi C, Shi H, Ma J, Han LZ, Lee JJ and Jin
X: Celastrol induces the apoptosis of breast cancer cells and
inhibits their invasion via downregulation of MMP-9. Oncol Rep.
32:2527–2532. 2014.PubMed/NCBI
|
|
17
|
Ni H, Zhao W, Kong X, Li H and Ouyang J:
NF-kappa B modulation is involved in celastrol induced human
multiple myeloma cell apoptosis. PLoS One. 9:e958462014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao X, Gao S, Ren H, Huang H, Ji W and
Hao J: Inhibition of autophagy strengthens celastrol-induced
apoptosis in human pancreatic cancer in vitro and in vivo models.
Curr Mol Med. 14:555–563. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee HW, Jang KS, Choi HJ, Jo A, Cheong JH
and Chun KH: Celastrol inhibits gastric cancer growth by induction
of apoptosis and autophagy. BMB Rep. 47:697–702. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang J, Song J, Wu D, Wang J and Dong W:
Hesperetin induces the apoptosis of hepatocellular carcinoma cells
via mitochondrial pathway mediated by the increased intracellular
reactive oxygen species, ATP and calcium. Med Oncol. 32:1012015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pieme C, Santosh G, Tekwu E, Askun T,
Aydeniz H, Ngogang J, Bhushan S and Saxena A: Fruits and barks
extracts of Zanthozyllum heitzii a spice from Cameroon induce
mitochondrial dependent apoptosis and G0/G1 phase arrest in human
leukemia HL-60 cells. Biol Res. 47:542014. View Article : Google Scholar
|
|
22
|
Zhang K, Wang X, Wang C, Zheng H, Li T,
Xiao S, Wang M, Fei C, Zhang L and Xue F: Investigation of
quinocetone-induced mitochondrial damage and apoptosis in HepG2
cells and compared with its metabolites. Environ Toxicol Pharmacol.
39:555–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Green DR and Reed JC: Mitochondria and
apoptosis. Science. 281:1309–1312. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Landriscina M, Laudiero G, Maddalena F,
Amoroso MR, Piscazzi A, Cozzolino F, Monti M, Garbi C, Fersini A,
Pucci P, et al: Mitochondrial chaperone Trap1 and the calcium
binding protein Sorcin interact and protect cells against apoptosis
induced by antiblastic agents. Cancer Res. 70:6577–6586. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Manfredi G, Kwong JQ, Oca-Cossio JA,
Woischnik M, Gajewski CD, Martushova K, D'Aurelio M, Friedlich AL
and Moraes CT: BCL-2 improves oxidative phosphorylation and
modulates adenine nucleotide translocation in mitochondria of cells
harboring mutant mtDNA. J Biol Chem. 278:5639–5645. 2003.
View Article : Google Scholar
|
|
27
|
Chang HY and Yang X: Proteases for cell
suicide: Functions and regulation of caspases. Microbiol Mol Biol
Rev. 64:821–846. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao X, Bennett RL and May WS: c-Myc and
caspase-2 are involved in activating Bax during cytotoxic
drug-induced apoptosis. J Biol Chem. 283:14490–14496. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stennicke HR and Salvesen GS: Properties
of the caspases. Biochim Biophys Acta. 1387:17–31. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hui KK, Kanungo AK, Elia AJ and Henderson
JT: Caspase-3 deficiency reveals a physiologic role for Smac/DIABLO
in regulating programmed cell death. Cell Death Differ.
18:1780–1790. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Volkmann N, Marassi FM, Newmeyer DD and
Hanein D: The rheostat in the membrane: BCL-2 family proteins and
apoptosis. Cell Death Differ. 21:206–215. 2014. View Article : Google Scholar :
|
|
32
|
Renault TT and Chipuk JE: Death upon a
kiss: Mitochondrial outer membrane composition and organelle
communication govern sensitivity to BAK/BAX-dependent apoptosis.
Chem Biol. 21:114–123. 2014. View Article : Google Scholar :
|
|
33
|
Westphal D, Kluck RM and Dewson G:
Building blocks of the apoptotic pore: How Bax and Bak are
activated and oligomerize during apoptosis. Cell Death Differ.
21:196–205. 2014. View Article : Google Scholar :
|
|
34
|
Chen PM, Cheng YW, Wu TC, Chen CY and Lee
H: MnSOD overexpression confers cisplatin resistance in lung
adenocarcinoma via the NF-κB/Snail/Bcl-2 pathway. Free Radic Biol
Med. 79:127–137. 2015. View Article : Google Scholar
|
|
35
|
Donskow-Łysoniewska K, Brodaczewska K and
Doligalska M: Heligmosomoides polygyrus antigens inhibit the
intrinsic pathway of apoptosis by overexpression of survivin and
Bcl-2 protein in CD4 T cells. Prion. 7:319–327. 2013. View Article : Google Scholar
|
|
36
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gillies LA and Kuwana T: Apoptosis
regulation at the mitochondrial outer membrane. J Cell Biochem.
115:632–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jourdain A and Martinou JC: Mitochondrial
outer-membrane permeabilization and remodelling in apoptosis. Int J
Biochem Cell Biol. 41:1884–1889. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wood WG, Igbavboa U, Muller WE and Eckert
GP: Statins, Bcl-2, and apoptosis: Cell death or cell protection?
Mol Neurobiol. 48:308–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Villa-Morales M and Fernández-Piqueras J:
Targeting the Fas/FasL signaling pathway in cancer therapy. Expert
Opin Ther Targets. 16:85–101. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gordon N and Kleinerman ES: Aerosol
therapy for the treatment of osteosarcoma lung metastases:
Targeting the Fas/FasL pathway and rationale for the use of
gemcitabine. J Aerosol Med Pulm Drug Deliv. 23:189–196. 2010.
View Article : Google Scholar : PubMed/NCBI
|