Introduction
Gastrointestinal stromal tumor (GIST) is the most
common mesenchymal tumor of the gastrointestinal tract derived from
the interstitial cells of Cajal (1). GIST develops in the stomach most
frequently, followed by the small intestine, colon and esophagus.
Expression of receptor tyrosine kinase KIT seems to be closely
associated with GIST development, and 75–90% of GISTs elicit
functional mutations of the KIT gene at exon 9, 11, 13 and 17
(2–4). Although tyrosine kinase inhibitors
(TKIs) such as imatinib mesylate or sunitinib malate have been
applied for the treatment of inoperable GISTs, tolerance to such
agents has often been induced and the antitumor effects by such
agents on GISTs is restricted (5,6).
Accordingly, new therapeutic modalities for GISTs which are based
on new biological mechanisms are urgently required.
Studies indicate that GIST has immunogenic
properties which attract the antitumor immune response.
Infiltration of immune cells into GIST tissues has been shown as
possible immune responses between the tumor and immune cells
(7,8). It has also been reported that NK cells
and the interferon-γ status of GIST patients predict prognosis
(9), and imatinib mesylate induced
NK cell activation and promoted antitumor immunity to GIST
(10). Of note, GISTs express
various types of tumor-associated antigens recognized by specific
cytotoxic T lymphocytes (11–14).
However, there is no evidence indicating that host antitumor
immunity inhibits the development of GISTs and no therapeutic
modalities which could suppress GISTs by activation of antitumor
immunity have been developed.
Recently, treatment with immune checkpoint
inhibitors has provided monumental progress in cancer treatment
(15,16). Some melanoma patients showed marked
responses to treatment with a monoclonal antibody to cytotoxic T
lymphocyte antigen-4 (CTLA-4) (17). Marked tumor regression, durable
antitumor effect and no autoimmune-associated adverse events of the
therapy indicated that inhibition of tumor-associated
immune-suppression could become a potential therapeutic modality to
suppress tumor development (18).
Recent studies have shown that the neo-antigens generated by
somatic mutations may become targets recognized by specific CTLs
(19). Blockade between programmed
cell death-1 (PD-1) on T cells and programmed cell death ligand-1
(PD-L1) on tumor cells by monoclonal antibody treatment also showed
a significant antitumor effect against melanomas as well as lung
and renal cancers (20,21). Combined treatment with anti-CTLA-4
and anti-PD-1 treatment of melanomas may show high therapeutic
efficacy which has never been observed before and may decrease the
toxicity of anti-CTAL-4 by decreasing its dose (18). Besides these molecules, T cell
immunoglobulin and mucin protein 3 (Tim-3) on immune cells and
galectin-9 on tumor cells play an important role in immune
checkpoint mechanism in malignancies and the blockade of the
Tim-3/galectin-9 pathway is expected for future immune checkpoint
therapy against malignant tumors (22,23).
Exciting results by immune-checkpoint blockade has opened a new era
for cancer therapy, and patients with various malignant tumors
other than melanomas may be able to receive the benefits in the
near future.
Considering that GISTs have immunogenic properties,
it is possible that antitumor immune activity of GISTs may be
inhibited by immune checkpoint mechanisms. Although immune
checkpoint blockade would be expected as a noted method for GIST
treatment, no data concerning the immune checkpoint mechanism of
GISTs have been reported. In the present study, we demonstrated
that the Tim-3/galectin-9 pathway may be involved in the immune
checkpoint mechanism of GISTs.
Materials and methods
Patient characteristics
Human GIST tumors investigated in the present study
were obtained from 2006 to 2014 from 12 male and 7 female patients
ranging from 40 to 82 years of age (median 60 years). All samples
were primary GIST tissues of the stomach, whose diameters were
>30 mm in 16 cases and from 30–100 mm in 3 cases. All of the
GIST tissue samples were obtained by surgical procedure, and no
distant metastatic lesions were noted. All the GIST tissues were
structured with spindle or epithelioid cells with positive c-kit
expression. None of the GIST cases were treated with tyrosine
kinase inhibitors. Analyses of the GIST tissues were approved by
the institutional review board according to the guidelines of the
Jikei University School of Medicine and the Jikei University
Hospital (25–299, 7434).
Antibodies
The primary antibodies for immunohistochemistry were
as follows: anti-human CD69 rabbit polyclonal antibody (pAb),
anti-galectin-9 rabbit pAb and anti-programmed death-1 (PD-1)
rabbit pAb (all from Abcam, Cambridge, UK), anti-Tim-3 rabbit pAb
(BioVision, Milpitas, CA, USA), anti-human CD4 mouse monoclonal
antibody (mAb), 1F6, anti-human CD8 mouse mAb, C8/144B and
anti-human CD56 mouse mAb, 1B6 (all from Nichirei Biosciences,
Tokyo, Japan), anti-FOXP3 mouse mAb, 236A/E7 (Abcam) and anti-human
CD68 mouse mAb, PG-M1 (Dako, Glostrup, Denmark).
Secondary antibodies for immunohistochemistry were
anti-mouse immunoglobulin goat pAb conjugated with peroxidase in
Max-PO(M) and anti-rabbit immunoglobulin goat pAb conjugated with
alkaline-phospatase in Max-PO(R) (both from Nichirei
Biosciences).
Secondary antibodies for immune-fluorescence
microscopy were goat anti-mouse immunoglobulin conjugated with
Alexa Fluor 488 and donkey anti-rabbit immunoglobulin conjugated
with Alexa Fluor 594 (both from Thermo Fischer Scientific, Waltham,
MA, USA).
Immunohistochemistry by immune-peroxidase
or alkaline phosphatase method
All stainings of tissue sections were performed on
4-µm formalin-fixed and paraffin-embedded tumor sections.
Tissue sections were de-waxed in xylene and re-hydrated through
decreasing concentrations of ethanol. Before immunostaining with
the primary antibody, the sections were washed with
phosphatase-buffered saline (PBS) and incubated with 10% bovine
serum in PBS for 60 min. They were then washed extensively with PBS
and incubated with each primary antibody at a dilution of 1:100 at
room temperature for 1 h. The slides were washed three times with
PBS and incubated with secondary antibodies at room temperature
according to the manufacturer's instructions. Each slide was
immersed in hematoxylin bath for 1 min and washed with water.
Tissue sections were re-hydrated in ethanol and cleared in xylene
and mounted with permanent mounting media.
Enumeration of the immune cells in the
GIST tissues
Numbers of CD8+ T, CD4+ T and
CD56+ NK cells, CD68+ macrophages and
FOXP3+ regulatory T cells were counted in 10 randomly
chosen x40 microscopic fields.
Immunofluorescence microscopy
Formalin-fixed and de-waxed GIST tissue sections
were incubated with the primary antibodies at a dilution of
1:100–1:200 at 4°C overnight. After being washed, they were
incubated with goat anti-mouse IgG (H+L) conjugated with Alexa
Fluor 488 or donkey anti-rabbit IgG (H+L) conjugated with Alexa
Fluor 594 (Thermo Fischer Scientific) at room temperature for 1 h.
Immunofluorescence images were detected with a immunofluorescence
microscope (BZ-9000 All-in-One; Keyence, Tokyo, Japan).
Correlation analysis
Correlation analyses were performed using Microsoft
Office Excel 2007 (Microsoft Corporation, Redmond, WA, USA).
Results
Infiltration of the immune cells into
GIST tissues
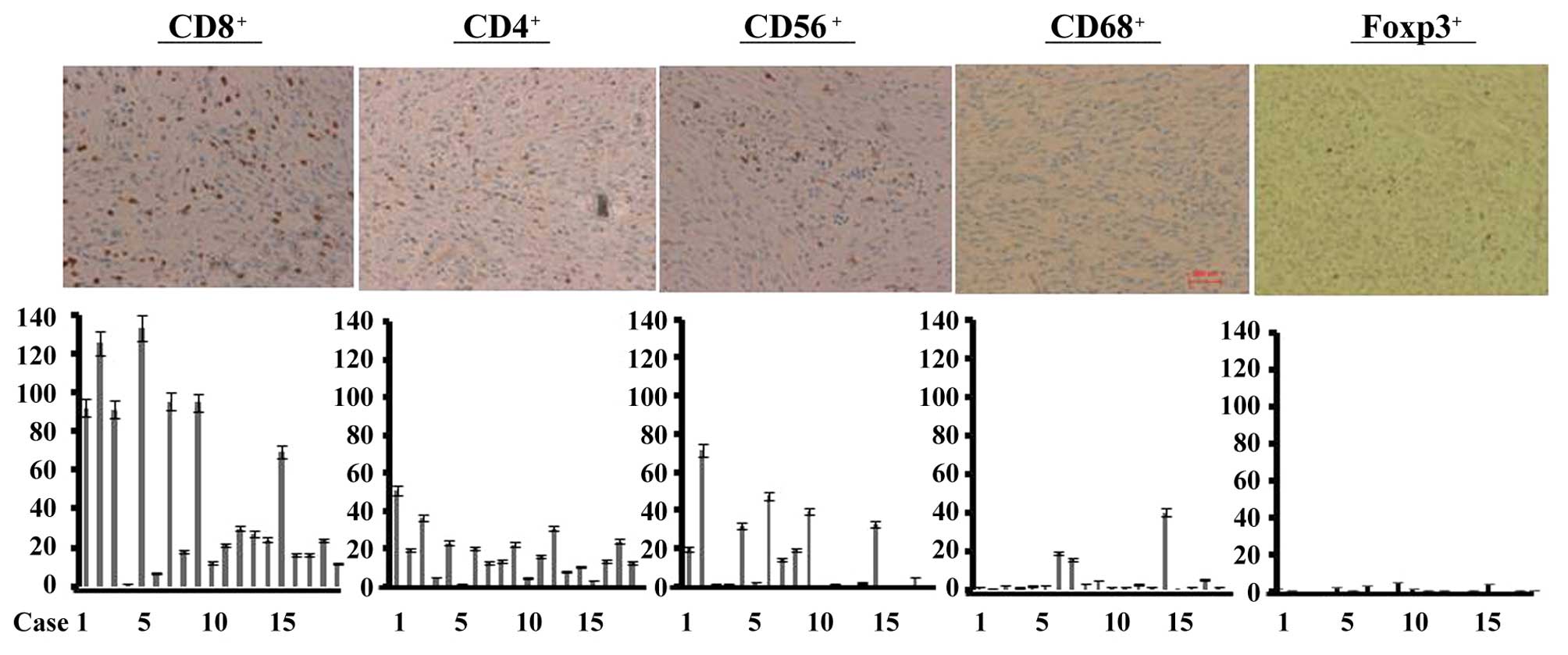
CD8+ T cells were infiltrated into all of
the GIST tissues at various degrees of infiltration (Fig. 1). CD4+ T cells were also
infiltrated into all of the GIST tissues, but fewer than that in
the CD8+ T cells. Significant infiltration of
CD56+ NK cells was observed in 8 GIST tissues (Fig. 1). Several cases showed macrophage
infiltration. The number of Foxp3+ cells was extremely
low in all of the GIST tissues examined (Fig. 1).
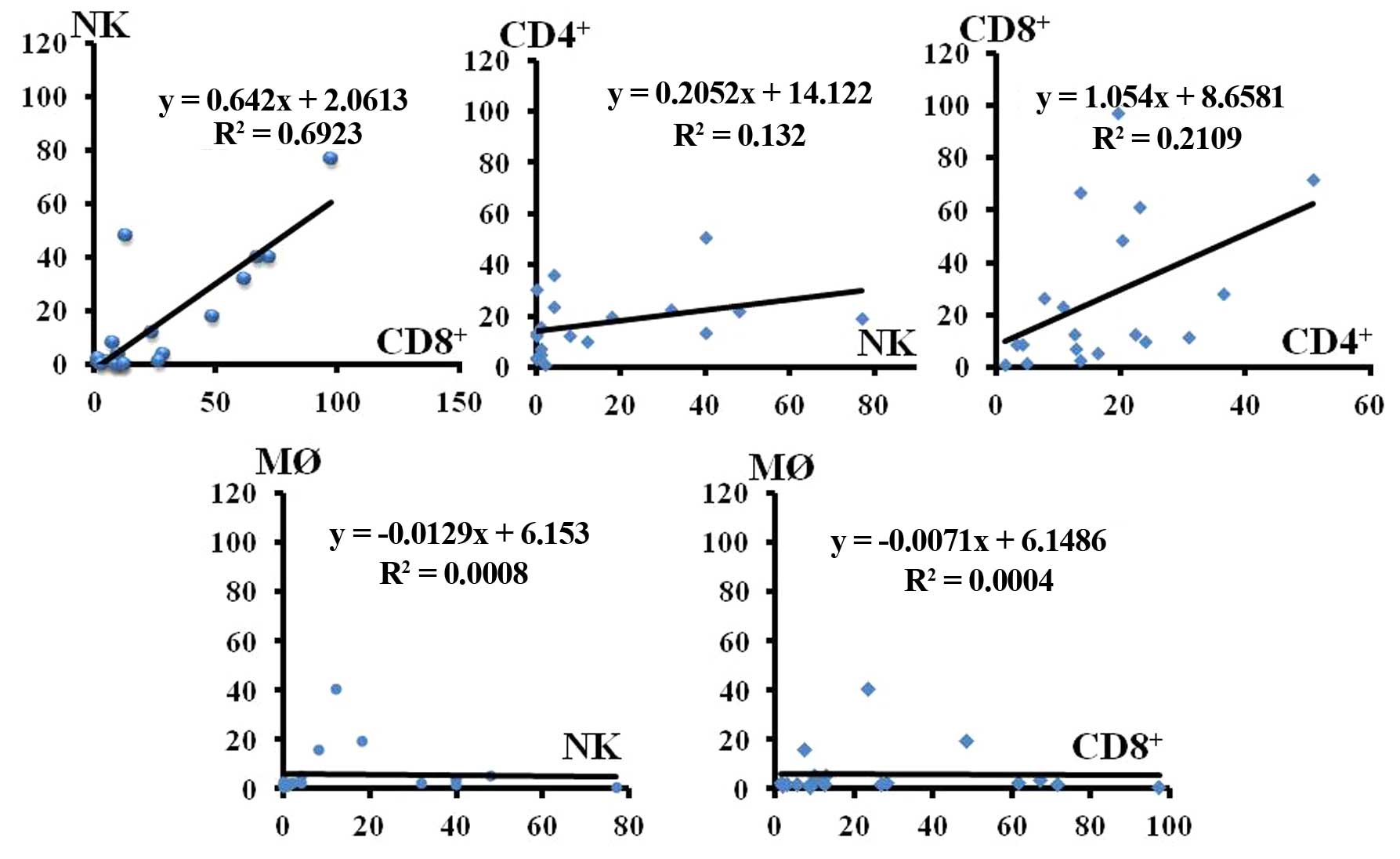
Correlation of the infiltration between each immune
cell type was examined. Infiltration of CD8+ T and NK
cells was found to be strongly correlated (Fig. 2, correlation coefficient 0.6923).
The correlation of infiltration between CD4+ T and NK
cells was low. Although the infiltration of CD8+ T cells
was correlated with that of CD4+ T cells, the
correlation was lower than that between CD8+ T and NK
cells (Fig. 2, correlation
coefficient 0.2109).
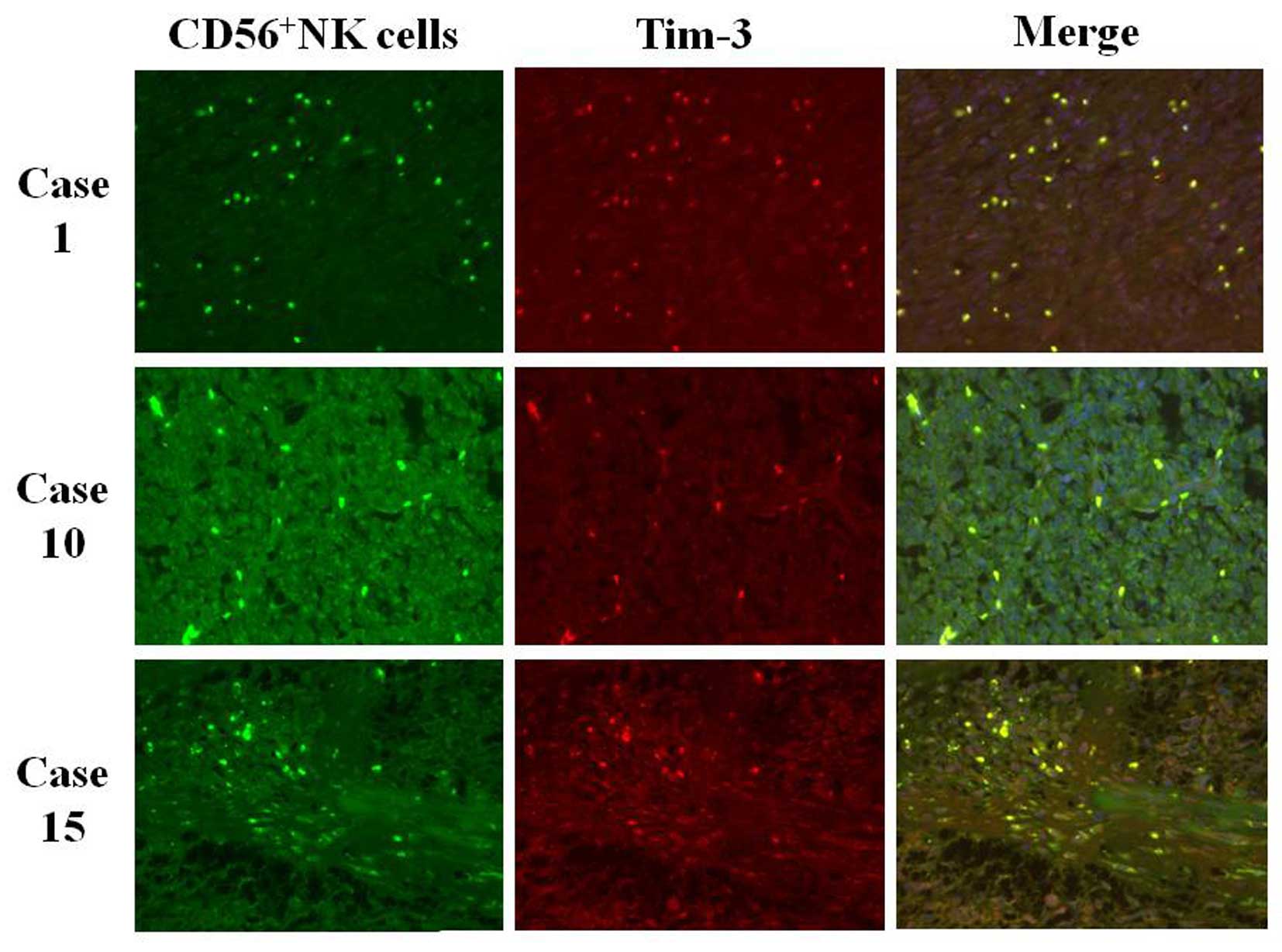
Positive Tim-3 but negative PD-1
expression is noted in the infiltrated NK cells in the GIST
tissues
The characteristics of the NK cells infiltrated into
the GIST tissues were examined. No CD69-positive NK cells were
found in all of the GIST tissues (Table
I). Tim-3 was significantly expressed in the infiltrated NK
cells in 6 out of 8 GIST tissues (Table
I and Fig. 3), but no GIST
tissue showed positive PD-1 staining in the infiltrated NK cells
(Table I).
 | Table IExpression of CD69, Tim-3 and PD-1 in
the CD56+ cells in the GIST tissues. |
Table I
Expression of CD69, Tim-3 and PD-1 in
the CD56+ cells in the GIST tissues.
| CD69
| Tim-3
| PD-1
|
|---|
| Positive | Negative | Positive | Negative | Positive | Negative |
|---|
| CD56+
cells | 0 | 8 | 6 | 2 | 0 | 8 |
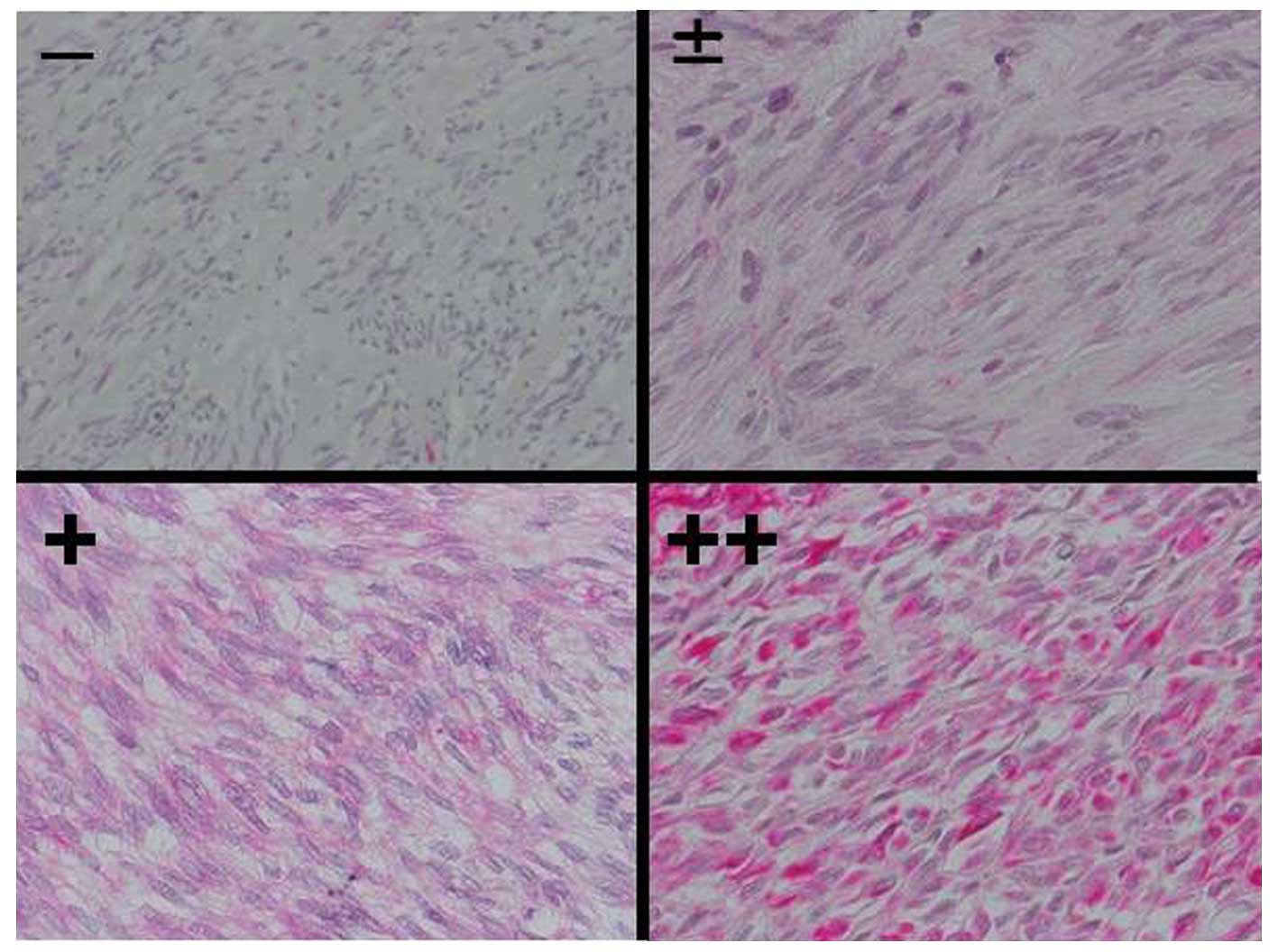
Galectin-9 expression in the GIST tumor
tissues and corresponding expression of Tim-3 in the infiltrated NK
cells
Expression of galectin-9 in the GIST tissues was
classified according to 4 grades: −, +/−, + and ++ (Fig. 4). All of the GIST tissues with
positive galectin-9 expression showed a cytoplasmic staining
pattern. Thirteen out of 19 GIST tissues (68.4%) were significantly
positive (+ to ++) for galectin-9 immunohistochemical staining
(Table II). Six GIST tissues with
Tim-3+ NK cell infiltration exhibited positive
galectin-9 expression (Table
III).
| Table IIExpression of galectin-9 in human
GIST tissues of the stomach. |
Table II
Expression of galectin-9 in human
GIST tissues of the stomach.
| Galectin-9
| Total |
|---|
| − | +/− | + | ++ |
|---|
| No. of tissues | 2 | 4 | 8 | 5 | 19 |
| Table IIIExpression of galectin-9 in GIST
tumor cells and Tim-3 in infiltrated NK cells in 8 GIST tissues
with significant NK cell infiltration. |
Table III
Expression of galectin-9 in GIST
tumor cells and Tim-3 in infiltrated NK cells in 8 GIST tissues
with significant NK cell infiltration.
| Case
|
|---|
| 1 | 2 | 5 | 7 | 8 | 9 | 10 | 15 |
|---|
| Galectin-9 | + | + | ++ | ++ | + | ++ | ++ | + |
| Tim-3 | + | + | − | + | + | − | + | + |
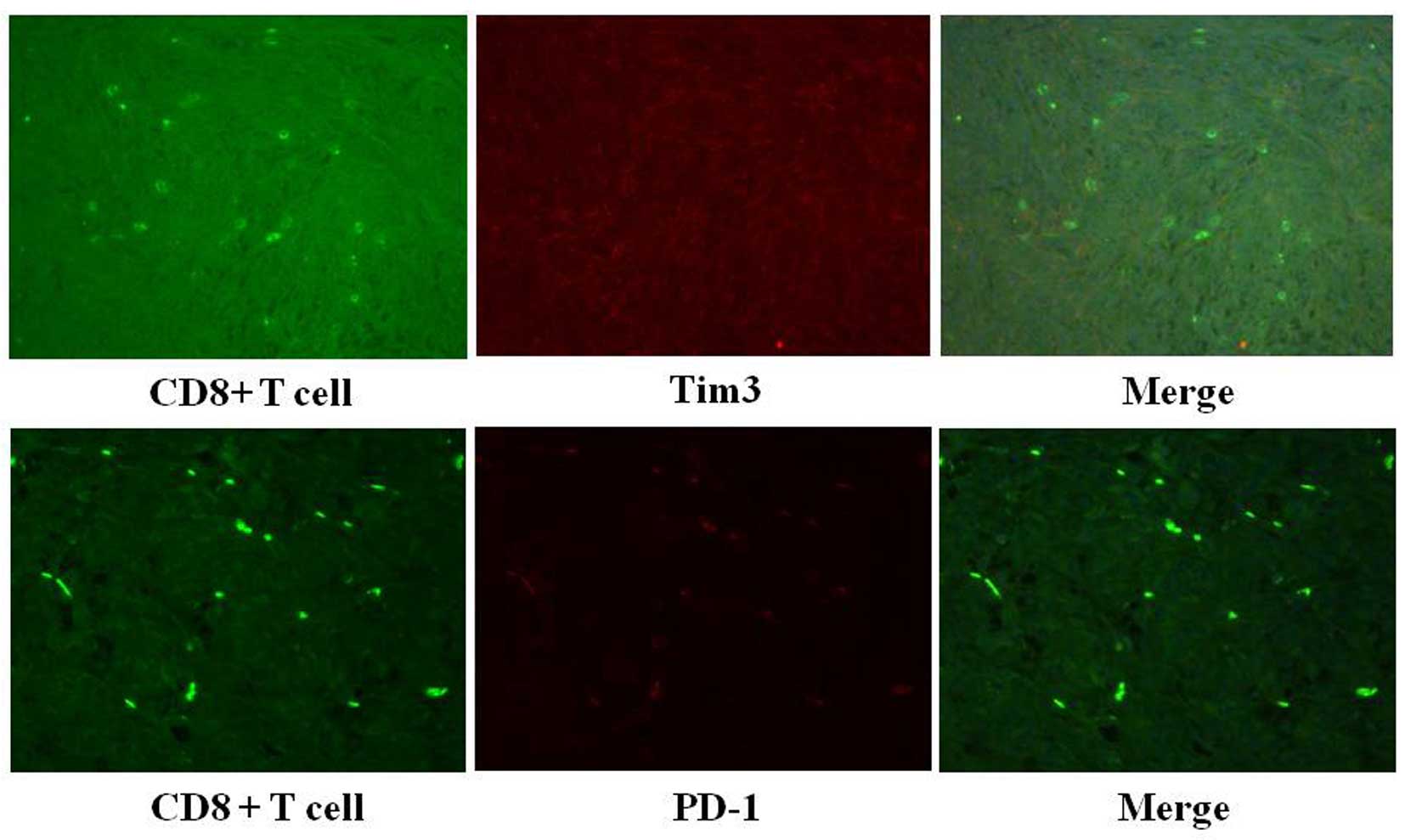
Negative Tim-3 or PD-1 expression in the
infiltrated CD8+ T cells in the GIST tissues
All of the CD8+ T cells in the 19 GIST
tissues were negative for CD69 staining (Table IV and Fig. 5). Furthermore, none of the
infiltrated CD8+ T cells in the GIST tissues showed
positive staining for Tim-3 or PD-1 (Table IV).
| Table IVExpression of CD69, Tim-3 and PD-1 in
CD8+ cells in the GIST tissues. |
Table IV
Expression of CD69, Tim-3 and PD-1 in
CD8+ cells in the GIST tissues.
| CD69
| Tim-3
| PD-1
|
|---|
| Positive | Negative | Positive | Negative | Positive | Negative |
|---|
| CD8+
cells | 0 | 19 | 0 | 19 | 0 | 19 |
Discussion
In the present study, we found that galectin-9 was
expressed in 68.4% of the human GISTs. Galectins elicit
β-galactoside binding capacity with the conserved carbohydrate
recognition domains (CRDs). Fifteen mammalian galectins have been
identified to date with three subtypes among which galectin-9 shows
tandem-repeat with two CRDs joined by a linker peptide (24). The multi-faceted and occasionally
conflicting roles of galectin-9 in tumor biology have been
demonstrated, and tumor progression, tumor cell adhesion,
metastasis and immune escape are the most likely involved (25). Galectin-9 expression has been shown
as a prognostic marker of malignancies indicating that
galectin-9-negative malignancies show frequent metastasis and those
with high galectin-9 expression are associated with prolonged
survival (26,27). In fact, GISTs are associated with a
rare incidence of recurrence after surgical resection and
infrequent distant metastases (1,4). It
remains unclear whether this low aggressiveness of GISTs is
associated with galectin-9 expression and whether GISTs showing
distant metastasis lack galectin-9 expression. Another important
biological implication of galectin-9 is its association with cell
cycle control and induction of apoptosis (28,29).
Galectin-9 induces the apoptosis of various types of blood cancer
cells (30,31). Galectin-9 may contribute to the
prevention of tumor metastasis by blocking adhesion to the
endothelium and extracellular matrix (32).
Previous studies have found that GISTs express known
tumor-associated antigens such as Wilms' tumor-1 (11–14),
suggesting that GISTs are immunogenic tumors (7). However, in the present study, all of
the CD8+ T and NK cells infiltrated into the GIST
tissues were CD69-negative inactivated cells. In spite of the
immune cell infiltration, no immune-related tumor cell destructions
were observed in the GIST tissues. These results suggest that
antitumor immunity is strongly suppressed in GIST tissues. Immune
suppression in GIST tissues by a macrophage-mediated mechanism or
others may be involved (33,34).
Galectin-9 induces inactivation and apoptosis of T
cells via Tim-3 receptor signaling (35,36)
and also suppresses NK cell function via Tim-3/galectin-9
interaction (37–40). On the contrary, a promoting effect
of galectin-9 on NK cell-mediated antitumor activity was also
reported (41), Tim-3/galectin-9
interaction seems to be complicated. The Tim-3 receptor has been
implicated as a negative regulator of adaptive immune responses and
has been linked to T-cell dysfunction in chronic viral infections
or cancer (42) and the regulatory
roles of Tim-3 on innate immune responses have also been shown
(38). In the present study, we
found that NK cells that infiltrated into the GIST tissues were
inactive and frequently expressed Tim-3. Considering that
galectin-9, a ligand of Tim-3, was frequently expressed in the GIST
tumor cells and Tim-3 was expressed in the infiltrated NK cells
corresponding to galectin-9 expression in the GIST tumor cells, it
is quite likely that the functions of NK cells in the GIST tissues
were suppressed by an immune checkpoint mechanism mediated by the
Tim-3/galectin-9 pathway. This suppressive effect may be achieved
through a complex mosaic of inhibitory or activating receptors
probably depending on the signal intensity, and Tim-3 signaling
acts as a suppressor for NK-cell response in GISTs (38,43).
CD8+ T cells that infiltrated into the
GIST tissues were all CD69-negative inactivated cells and no
CD8+ T cells expressed Tim-3 or PD-1. Unfortunately,
expression of PD-L1, a ligand of PD-1, in the GIST tissues was
undefined, since the results of PD-L1 staining of the GIST tissues
was quite different depending on the type of anti-PD-L1 antibody
used for immunohistochemistry. Considering the absence of PD-1
expression in the infiltrated CD8+ T and
CD56+ NK cells, the PD-1/PD-L1 pathway is not involved
in the immune checkpoint mechanism in GISTs.
Although tumor-associated antigens recognized by
specific CTLs were expressed in the GISTs, adaptive antitumor
immunity was deceased. We performed primary cultures of resected
GIST tissues in IL-2-containing medium in some cases and observed
vigorously proliferating mononuclear cells migrating from the GIST
tissues. They mainly consisted of CD56+ NK and
CD8+ T cells, and HLA-A24 restricted WT1
tetramer-positive CD8+ T cells were present. These
findings indicate that adaptive immunity targeting known tumor
antigens may have been primed but suppressed in the GIST tissues.
It has been stressed that innate immune responses including NK cell
response are an important process for the induction of antitumor
immune activity (44) and that
activation of NK cell-mediated immune response is closely
associated with the following activation of adaptive immunity
(45,46). Suppression of NK cell activity may
be correlated with the impaired CD8+ T cell-mediated
adaptive antitumor immunity in GIST tissues. Finally, blockade of
the immune checkpoint mediated by Tim-3/Galectin-9 interaction may
potentially re-activate NK cell activity and may be beneficial for
the treatment of advanced GISTs.
Abbreviations:
|
GIST
|
gastrointestinal stromal tumor
|
|
NK cell
|
natural killer cell
|
|
CTL
|
cytotoxic T lymphocyte, PD-1,
programmed death-1
|
|
Tim-3
|
T cell immunoglobulin and mucin
protein 3
|
|
mAb
|
monoclonal antibody
|
|
pAb
|
polyclonal antibody
|
References
|
1
|
Miettinen M and Lasota J: Gastrointestinal
stromal tumors-definition, clinical, histological,
immunohistochemical, and molecular genetic features and
differential diagnosis. Virchows Arch. 438:1–12. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hirota S, Isozaki K, Moriyama Y, Hashimoto
K, Nishida T, Ishiguro S, Kawano K, Hanada M, Kurata A, Takeda M,
et al: Gain-of-function mutations of c-kit in human
gastrointestinal stromal tumors. Science. 279:577–580. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rubin BP, Singer S, Tsao C, Duensing A,
Lux ML, Ruiz R, Hibbard MK, Chen CJ, Xiao S, Tuveson DA, et al: KIT
activation is a ubiquitous feature of gastrointestinal stromal
tumors. Cancer Res. 61:8118–8121. 2001.PubMed/NCBI
|
|
4
|
Corless CL, Fletcher JA and Heinrich MC:
Biology of gastrointestinal stromal tumors. J Clin Oncol.
22:3813–3825. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Demetri GD, von Mehren M, Blanke CD, Van
den Abbeele AD, Eisenberg B, Roberts PJ, Heinrich MC, Tuveson DA,
Singer S, Janicek M, et al: Efficacy and safety of imatinib
mesylate in advanced gastrointestinal stromal tumors. N Engl J Med.
347:472–480. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Le Cesne A, Ray-Coquard I, Bui BN, Adenis
A, Rios M, Bertucci F, Duffaud F, Chevreau C, Cupissol D, Cioffi A,
et al French Sarcoma Group: Discontinuation of imatinib in patients
with advanced gastrointestinal stromal tumours after 3 years of
treatment: An open-label multicentre randomised phase 3 trial.
Lancet Oncol. 11:942–949. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rusakiewicz S, Semeraro M, Sarabi M,
Desbois M, Locher C, Mendez R, Vimond N, Concha A, Garrido F,
Isambert N, et al: Immune infiltrates are prognostic factors in
localized gastrointestinal stromal tumors. Cancer Res.
73:3499–3510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rubin BP, Heinrich MC and Corless CL:
Gastrointestinal stromal tumour. Lancet. 369:1731–1741. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ménard C, Blay JY, Borg C, Michiels S,
Ghiringhelli F, Robert C, Nonn C, Chaput N, Taïeb J, Delahaye NF,
et al: Natural killer cell IFN-gamma levels predict long-term
survival with imatinib mesylate therapy in gastrointestinal stromal
tumor-bearing patients. Cancer Res. 69:3563–3569. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Borg C, Terme M, Taïeb J, Ménard C,
Flament C, Robert C, Maruyama K, Wakasugi H, Angevin E, Thielemans
K, et al: Novel mode of action of c-kit tyrosine kinase inhibitors
leading to NK cell-dependent antitumor effects. J Clin Invest.
114:379–388. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Perez D, Herrmann T, Jungbluth AA,
Samartzis P, Spagnoli G, Demartines N, Clavien PA, Marino S,
Seifert B and Jaeger D: Cancer testis antigen expression in
gastrointestinal stromal tumors: New markers for early recurrence.
Int J Cancer. 123:1551–1555. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Perez D, Hauswirth F, Jäger D, Metzger U,
Samartzis EP, Went P and Jungbluth A: Protein expression of cancer
testis antigens predicts tumor recurrence and treatment response to
imatinib in gastrointestinal stromal tumors. Int J Cancer.
128:2947–2952. 2011. View Article : Google Scholar
|
|
13
|
Ghadban T, Perez DR, Vashist YK, Bockhorn
M, Koenig AM, El Gammal AT, Izbicki JR, Metzger U, Hauswirth F,
Frosina D, et al: Expression of cancer testis antigens CT10
(MAGE-C2) and GAGE in gastrointestinal stromal tumors. Eur J Surg
Oncol. 40:1307–1312. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kang GH, Kim KM, Noh JH, Sohn TS, Kim S,
Park CK, Lee CS and Kang DY: WT-1 expression in gastrointestinal
stromal tumours. Pathology. 42:54–57. 2010. View Article : Google Scholar
|
|
15
|
Pardoll DM: The blockade of immune
checkpoints in cancer immunotherapy. Nat Rev Cancer. 12:252–264.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ramsay AG: Immune checkpoint blockade
immunotherapy to activate anti-tumour T-cell immunity. Br J
Haematol. 162:313–325. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hodi FS, O'Day SJ, McDermott DF, Weber RW,
Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel
JC, et al: Improved survival with ipilimumab in patients with
metastatic melanoma. N Engl J Med. 363:711–723. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wolchok JD, Kluger H, Callahan MK, Postow
MA, Rizvi NA, Lesokhin AM, Segal NH, Ariyan CE, Gordon RA, Reed K,
et al: Nivolumab plus ipilimumab in advanced melanoma. N Engl J
Med. 369:122–133. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Snyder A, Makarov V, Merghoub T, Yuan J,
Zaretsky JM, Desrichard A, Walsh LA, Postow MA, Wong P, Ho TS, et
al: Genetic basis for clinical response to CTLA-4 blockade in
melanoma. N Engl J Med. 371:2189–2199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Topalian SL, Hodi FS, Brahmer JR,
Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD,
Sosman JA, Atkins MB, et al: Safety, activity, and immune
correlates of anti-PD-1 antibody in cancer. N Engl J Med.
366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ,
Topalian SL, Hwu P, Drake CG, Camacho LH, Kauh J, Odunsi K, et al:
Safety and activity of anti-PD-L1 antibody in patients with
advanced cancer. N Engl J Med. 366:2455–2465. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
da Silva IP, Gallois A, Jimenez-Baranda S,
Khan S, Anderson AC, Kuchroo VK, Osman I and Bhardwaj N: Reversal
of NK-cell exhaustion in advanced melanoma by Tim-3 blockade.
Cancer Immunol Res. 2:410–422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fujihara S, Mori H, Kobara H, Rafiq K,
Niki T, Hirashima M and Masaki T: Galectin-9 in cancer therapy.
Recent Pat Endocr Metab Immune Drug Discov. 7:130–137. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Leffler H, Carlsson S, Hedlund M, Qian Y
and Poirier F: Introduction to galectins. Glycoconj J. 19:433–440.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heusschen R, Griffioen AW and Thijssen VL:
Galectin-9 in tumor biology: A jack of multiple trades. Biochim
Biophys Acta. 1836:177–185. 2013.PubMed/NCBI
|
|
26
|
Irie A, Yamauchi A, Kontani K, Kihara M,
Liu D, Shirato Y, Seki M, Nishi N, Nakamura T, Yokomise H, et al:
Galectin-9 as a prognostic factor with antimetastatic potential in
breast cancer. Clin Cancer Res. 11:2962–2968. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van den Brûle F, Califice S and Castronovo
V: Expression of galectins in cancer: A critical review. Glycoconj
J. 19:537–542. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang RY and Liu FT: Galectins in cell
growth and apoptosis. Cell Mol Life Sci. 60:267–276. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu FT, Yang RY, Saegusa J, Chen HY and
Hsu DK: Galectins in regulation of apoptosis. Adv Exp Med Biol.
705:431–442. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kageshita T, Kashio Y, Yamauchi A, Seki M,
Abedin MJ, Nishi N, Shoji H, Nakamura T, Ono T and Hirashima M:
Possible role of galectin-9 in cell aggregation and apoptosis of
human melanoma cell lines and its clinical significance. Int J
Cancer. 99:809–816. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi T, Kuroda J, Ashihara E, Oomizu
S, Terui Y, Taniyama A, Adachi S, Takagi T, Yamamoto M, Sasaki N,
et al: Galectin-9 exhibits anti-myeloma activity through JNK and
p38 MAP kinase pathways. Leukemia. 24:843–850. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nobumoto A, Nagahara K, Oomizu S, Katoh S,
Nishi N, Takeshita K, Niki T, Tominaga A, Yamauchi A and Hirashima
M: Galectin-9 suppresses tumor metastasis by blocking adhesion to
endothelium and extracellular matrices. Glycobiology. 18:735–744.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
van Dongen M, Savage ND, Jordanova ES,
Briaire-de Bruijn IH, Walburg KV, Ottenhoff TH, Hogendoorn PC, van
der Burg SH, Gelderblom H and van Hall T: Anti-inflammatory M2 type
macrophages characterize metastasized and tyrosine kinase
inhibitor-treated gastrointestinal stromal tumors. Int J Cancer.
127:899–909. 2010.
|
|
34
|
Cameron S, Haller F, Dudas J, Moriconi F,
Gunawan B, Armbrust T, Langer C, Füzesi L and Ramadori G: Immune
cells in primary gastrointestinal stromal tumors. Eur J
Gastroenterol Hepatol. 20:327–334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhu C, Anderson AC, Schubart A, Xiong H,
Imitola J, Khoury SJ, Zheng XX, Strom TB and Kuchroo VK: The Tim-3
ligand galectin-9 negatively regulates T helper type 1 immunity.
Nat Immunol. 6:1245–1252. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li H, Wu K, Tao K, Chen L, Zheng Q, Lu X,
Liu J, Shi L, Liu C, Wang G, et al: Tim-3/galectin-9 signaling
pathway mediates T-cell dysfunction and predicts poor prognosis in
patients with hepatitis B virus-associated hepatocellular
carcinoma. Hepatology. 56:1342–1351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ju Y, Hou N, Meng J, Wang X, Zhang X, Zhao
D, Liu Y, Zhu F, Zhang L, Sun W, et al: T cell immunoglobulin - and
mucin-domain-containing molecule-3 (Tim-3) mediates natural killer
cell suppression in chronic hepatitis B. J Hepatol. 52:322–329.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Han G, Chen G, Shen B and Li Y: Tim-3: An
activation marker and activation limiter of innate immune cells.
Front Immunol. 4:4492013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hou H, Liu W, Wu S, Lu Y, Peng J, Zhu Y,
Lu Y, Wang F and Sun Z: Tim-3 negatively mediates natural killer
cell function in LPS-induced endotoxic shock. PLoS One.
9:e1105852014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Golden-Mason L, McMahan RH, Strong M,
Reisdorph R, Mahaffey S, Palmer BE, Cheng L, Kulesza C, Hirashima
M, Niki T, et al: Galectin-9 functionally impairs natural killer
cells in humans and mice. J Virol. 87:4835–4845. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gleason MK, Lenvik TR, McCullar V, Felices
M, O'Brien MS, Cooley SA, Verneris MR, Cichocki F, Holman CJ,
Panoskaltsis- Mortari A, et al: Tim-3 is an inducible human natural
killer cell receptor that enhances interferon gamma production in
response to galectin-9. Blood. 119:3064–3072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ferris RL, Lu B and Kane LP: Too much of a
good thing? Tim-3 and TCR signaling in T cell exhaustion. J
Immunol. 193:1525–1530. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ndhlovu LC, Lopez-Vergès S, Barbour JD,
Jones RB, Jha AR, Long BR, Schoeffler EC, Fujita T, Nixon DF and
Lanier LL: Tim-3 marks human natural killer cell maturation and
suppresses cell-mediated cytotoxicity. Blood. 119:3734–3743. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Campbell KS and Hasegawa J: Natural killer
cell biology: An update and future directions. J Allergy Clin
Immunol. 132:536–544. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee KM, Bhawan S, Majima T, Wei H,
Nishimura MI, Yagita H and Kumar V: Cutting edge: The NK cell
receptor 2B4 augments antigen-specific T cell cytotoxicity through
CD48 ligation on neighboring T cells. J Immunol. 170:4881–4885.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mocikat R, Braumüller H, Gumy A, Egeter O,
Ziegler H, Reusch U, Bubeck A, Louis J, Mailhammer R, Riethmüller
G, et al: Natural killer cells activated by MHC class I(low)
targets prime dendritic cells to induce protective CD8 T cell
responses. Immunity. 19:561–569. 2003. View Article : Google Scholar : PubMed/NCBI
|