Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors and the fourth primary cause of
tumor-related deaths worldwide with high mortality and poor
prognosis (1). Although many types
of therapeutic measures including surgical resection, transarterial
chemoembolization (TACE), radiation and chemotherapy have been used
for the treatment of HCC, most patients progress to an advanced
stage after the initial therapeutic benefit attributed to high
chemoresistance, particularly due to the multidrug resistance (MDR)
of HCC (2). Sorafenib (SOR), an
oral multikinase inhibitor, which inhibits tumor growth and
angiogenesis by way of inhibiting vascular endothelial growth
factor receptor 2 and other receptor tyrosine kinases has been used
as the standard treatment for advanced stages of HCC based on two
large randomized phase III trials, which led to the Food and Drug
Administration (FDA) approval since it prolongs survival for 2–3
months in advanced and inoperable HCC cases (3–5).
However, clinical results have been disappointing showing that a
large number of advanced HCC patients are unresponsive or acquire
resistance to SOR. Therefore, it is urgent to seek new effective
therapy strategies to combat HCC.
Non-steroidal anti-inflammatory drugs (NSAIDs) have
been reported to reduce the risk of developing cancer (6–8).
Meloxicam (MEL), a selective cyclooxygenase-2 (COX-2) inhibitor,
has been demonstrated to inhibit proliferation and promote
apoptosis in many malignant diseases (9–11). Our
previous experimental results showed that COX-2 inhibitor exhibits
antiproliferative and proapoptotic effects in HCC cell lines
(12,13). However, the detailed effects and
mechanisms of MEL combined with SOR for treating HCC cells have not
been fully cleared. Recently, a number of studies have revealed
that certain chemotherapeutics lead to cell death by the way of the
ER stress-related apoptosis (14,15).
Therefore, we hypothesized that ER stress promoting proapoptotic
effects or inhibiting its proliferative function may be a potential
target for the treatment of HCC. The endoplasmic reticulum (ER), a
central cellular organelle, plays a crucial role in protein folding
and maturation as well as accumulation of intracellular calcium.
Small errors in these processes could disturb normal ER processes
and lead to ER stress known as the unfolded protein response (UPR).
GRP78, as an ER molecular chaperone, is upregulated when ER stress
is induced and functions as a sensory hub and inhibitor of three ER
transmembrane receptors: eukaryotic translation initiation factor
2α kinase 3 (EIF2AK3/PERK), inositol-requiring enzyme 1 (IRE1) and
activating transcription factor-6 (ATF6) (16). The UPR initially targets proteins
for degradation and restores the proper ER homeostasis. However, it
eventually induces cell death during intense ER stress (17). Our purpose in the present study was
to explore the combined effects of MEL and SOR on apoptosis and
evaluate the probable mechanisms of action in HCC cell lines.
Materials and methods
Cell culture and animals
Human hepatocellular cancer (HCC) SMMC-7721 cells
were obtained from the American Type Culture Collection (ATCC;
Rockville, MD, USA). The cells were cultured in RPMI-1640 medium
(Gibco)/Dulbecco's modified Eagle's medium (DMEM) (HyClone)
containing 10% fetal bovine serum (FBS; Gibco), 100 U/ml penicillin
and 100 μg/ml streptomycin (Gibco) at 37°C in 95% air and 5%
CO2. BALB/c male athymic mice (5–6 weeks old, 18–22 g)
were purchased from the Animal Supplier Center of Shandong
University. All the animal studies were approved by the Ethics
Committee of Shandong University. All surgical procedures were
performed under anesthesia with sodium pentobarbital.
Reagents and antibodies
The MEL was purchased from Merck Millipore
(Darmstadt, Germany), dissolved in dimethylsulfoxide (DMSO;
Sigma-Aldrich, St. Louis, MO, USA) at 10 or 50 mM stock and diluted
immediately before each experiment. SOR tosylate was obtained from
Bayer Health Care (Berlin, Germany) and dissolved in DMSO to a 10
mM stock. MG132 was obtained from Sigma-Aldrich (San Diego, CA,
USA). Primary antibodies to GRP78, caspase-12, PARP and caspase-3
were purchased from Abcam (Cambridge, UK). Antibodies to IRE1 and
phos-eIF2α were obtained from Cell Signaling Technology (Danvers,
MA, USA) and the anti-GAPDH antibody was obtained from Abcam.
Measurement of cell viability
Cell viability assays were performed using the Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Japan).
Cells (5×103/well) were seeded with culture medium onto
96-well plates and incubated at 37°C for 24 h. After adaptation,
cells were treated with either MEL or SOR, or in combination for 48
h. Then the culture medium was replaced with fresh medium
containing 10 ml of CCK-8 solution. The optical density (OD) at 450
nm was assayed following cell incubation at 37°C for 2 h. The
viability inhibition rate was calculated as: (control OD value -
experiment OD value)/control group OD value × 100%. The coefficient
of drug interaction (CDI) analysis for evaluating effects of drug
combinations was calculated according to Cao et al (18) using the equation: CDI = AB/(A × B).
A or B is the ratio of the single agent group to the control group
and AB is the ratio of the combination groups to the control group.
A CDI of ≤ or >1 indicates synergy, additivity or antagonism,
respectively. A CDI <0.7 shows that the drugs are significantly
synergistic.
Apoptosis assay
Cells were stained with Annexin V-FITC apoptosis
detection kit (BD Biosciences, San Jose, CA, USA). According to the
manufacturer's instructions, the cells were incubated with 5 ml of
Annexin V and 5 ml of propidium iodide (PI) for 15 min at room
temperature, and then the stained cells were analyzed on a FACS
flow cytometer.
Cell cycle analysis by flow
cytometry
SMMC-7721 cells were treated with MEL or SOR or in
combination for 24 h and then the cells were performed by cell
cycle analysis. In brief, 5×104 cells were suspended in
0.5 ml of PI solution, and incubated 30 min in the dark according
to the manufacturer's instructions. Cell cycle distribution was
analyzed by FACS flow cytometry.
Cell migration and invasion assays
Cells (1×105) in 300 ml of RPMI-1640
medium/DMEM (with 1% FBS) containing MEL or SOR alone, or in
combination were seeded into the upper chamber of a Transwell
chamber (Corning, New York, NY, USA). The bottom wells of the
chambers were filled with 500 ml RPMI-1640 medium/DMEM containing
10% FBS. After 48 h of incubation, the chambers were fixed with 95%
ethanol and then stained with 1% crystal violet. Similarly, the
cell invasion assay was performed by adding Matrigel Basement
Matrix to the upper chamber.
Western blot analysis
Western blotting was used to evaluate apoptosis and
ER stress-related signaling. After different treatments, protein
concentrations in cell extracts were determined (Bio-Rad, Richmond,
CA, USA), equal amounts of each sample were resolved in SDS-PAGE
gels, then transferred to a polyvinylidene fluoride (PVDF) membrane
(Millipore, Billerica, MA, USA), and probed with specific
antibodies. Blots were developed using applicable horseradish
peroxidase (HRP)-conjugated secondary antibodies and visualized by
enhanced chemiluminescence with ECL (Millipore). Protein band
intensities were quantified by densitometric analysis using ImageJ
software (National Institutes of Health, USA).
Gene transfection and RNAi
For knockdown of GRP78, a small interfering RNA
(si-RNA) targeting human GRP78 and a control siRNA were obtained
from Santa Cruz Biotechnology (Santa Cruz, CA, USA). SMMC-7721
cells were seeded onto 6-well plates and after 24 h were
transfected using Lipofectamine 2000 transfection reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer's
instructions. The transfected cells were treated with MEL or SOR or
in combination in complete medium for 24 h.
Immunofluorescence assay
Human SMMC-7721 cells seeded onto coverslips in
6-well plates, were fixed with 4% paraformaldehyde (PFA) and were
permeabilized in 0.1% Triton X-100. Incubation with primary
antibodies for 2 h at room temperature was followed by incubation
with fluorescein isothiocyanate (FITC)-labeled IgG secondary
antibodies, and then cells were mounted onto microscope slides with
a DAPI mounting solution (Abcam). Fluorescent images of the HCC
cells were photographed and analyzed with a light microscope
(magnification, ×200; Olympus, Tokyo, Japan).
Immunohistochemistry assay
Briefly, tissue sections (4 mm) were deparaffinized
in graded xylene and rehydrated in graded ethanol, followed by
three washes with phosphate-buffered saline (PBS) for 3 min each
and 1% H2O2 for 30 min in the dark to inhibit
endogenous peroxidase activity. Primary anti-caspase-12 (1:500
dilution) antibodies were applied overnight at 4°C. After washing,
the sections were incubated with a biotinylated secondary antibody
(Zhongshan, Beijing, China) for 30 min at 37°C. Negative control
sections were incubated with PBS instead of the primary antibody.
The slides were examined under a light microscope (magnification,
×400; Olympus).
Measurement of in vivo activity
Under sterile conditions, SMMC-7721 cells
(5×106 cells/animal) were subcutaneously inoculated into
the nude mice. The inoculated mice were randomly divided into four
groups, with five mice in each group; the body weight difference
between groups was not significant. In the control group, an
intraperitoneal injection of saline (10 ml/kg) was performed every
other day. In group 2 and 3, an intraperitoneal injection of MEL
(30 mg/kg) or SOR (30 mg/kg) was administered every other day,
respectively. In experimental group 4 (the combination group), MEL
(30 mg/kg) and SOR (30 mg/kg) were administered by injection every
other day. The mice were sacrificed five weeks after tumor
implantation. The weights and tumor volumes of the nude mice were
recorded every second day until the animals were sacrificed. The
tumor volume (V) was monitored by measuring its length (L) and
width (W) with calipers and calculated using the following formula:
V = (L×W2) × 0.5. The animal care and experimental
protocols were in accordance with the institutional guidelines.
Statistical analysis
Data are presented as the mean ± standard deviation
(SD) and were analyzed by one-way ANOVA followed by the Dunnett's
test with SPSS software (version 17.0; SPSS China, Shanghai,
China), with values of P<0.05 considered to indicate a
statistically significant result.
Results
Combination of MEL and SOR significantly
inhibits cell viability in SMMC-7721 cells
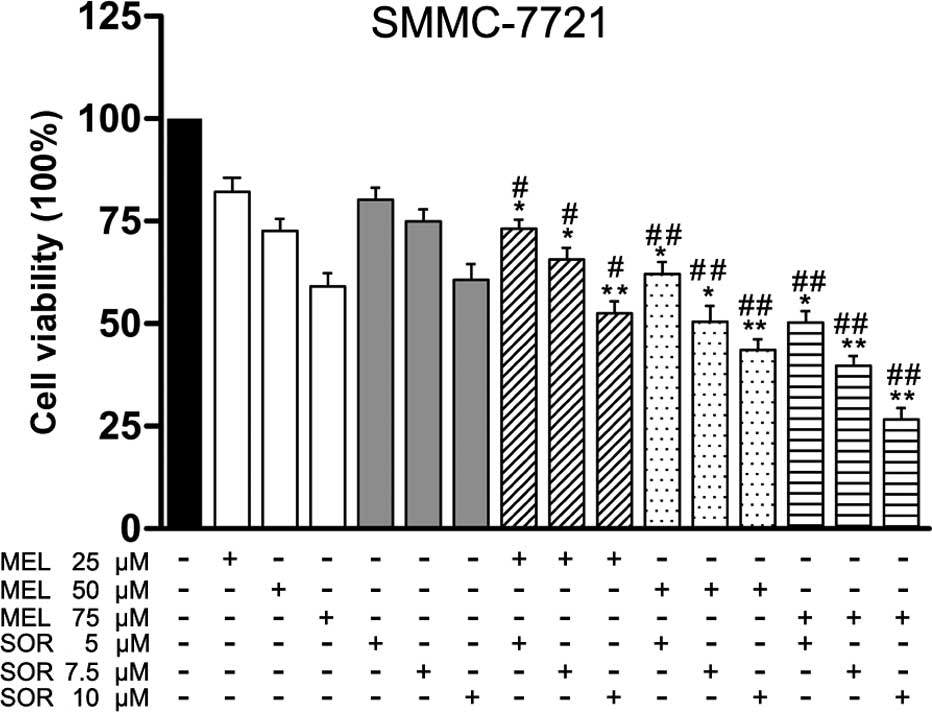
In order to investigate the effect of MEL, SOR and
their combination on the cell viability of HCC cells in
vitro, SMMC-7721 cells were treated with different
concentrations of MEL (0–75 μM) or SOR (0–10 μM). As
depicted in Fig. 1, treatment with
MEL or SOR for 48 h significantly inhibited cell viability in
SMMC-7721 cells with an IC50 value of 75.6±0.8 μM
of MEL alone or an IC50 value of 10.2±1.5 μM of
SOR alone. Next, we investigated whether MEL enhanced sensitivity
of SMMC-7721 cells to SOR treatment. The CDI was utilized to
display the effects of interaction between these two drugs. Our
results showed that combination with MEL significantly enhanced SOR
lethality and exhibited strong synergistic effects for SMMC-7721
cells (Table I).
 | Table ICDI of the combination of meloxicam
and sorafenib in SMMC-7721 cells. |
Table I
CDI of the combination of meloxicam
and sorafenib in SMMC-7721 cells.
| SMMC-7721 Sorafenib
(μM)
|
|---|
| 5 | 7.5 | 10 |
|---|
| Meloxicam
(μM) | 25 | 0.893 | 0.838 | 0.734 |
| 50 | 0.726 | 0.672 | 0.613 |
| 75 | 0.683 | 0.607 | 0.558 |
Combination of MEL and SOR inhibits
migration and invasion in SMMC-7721 cells
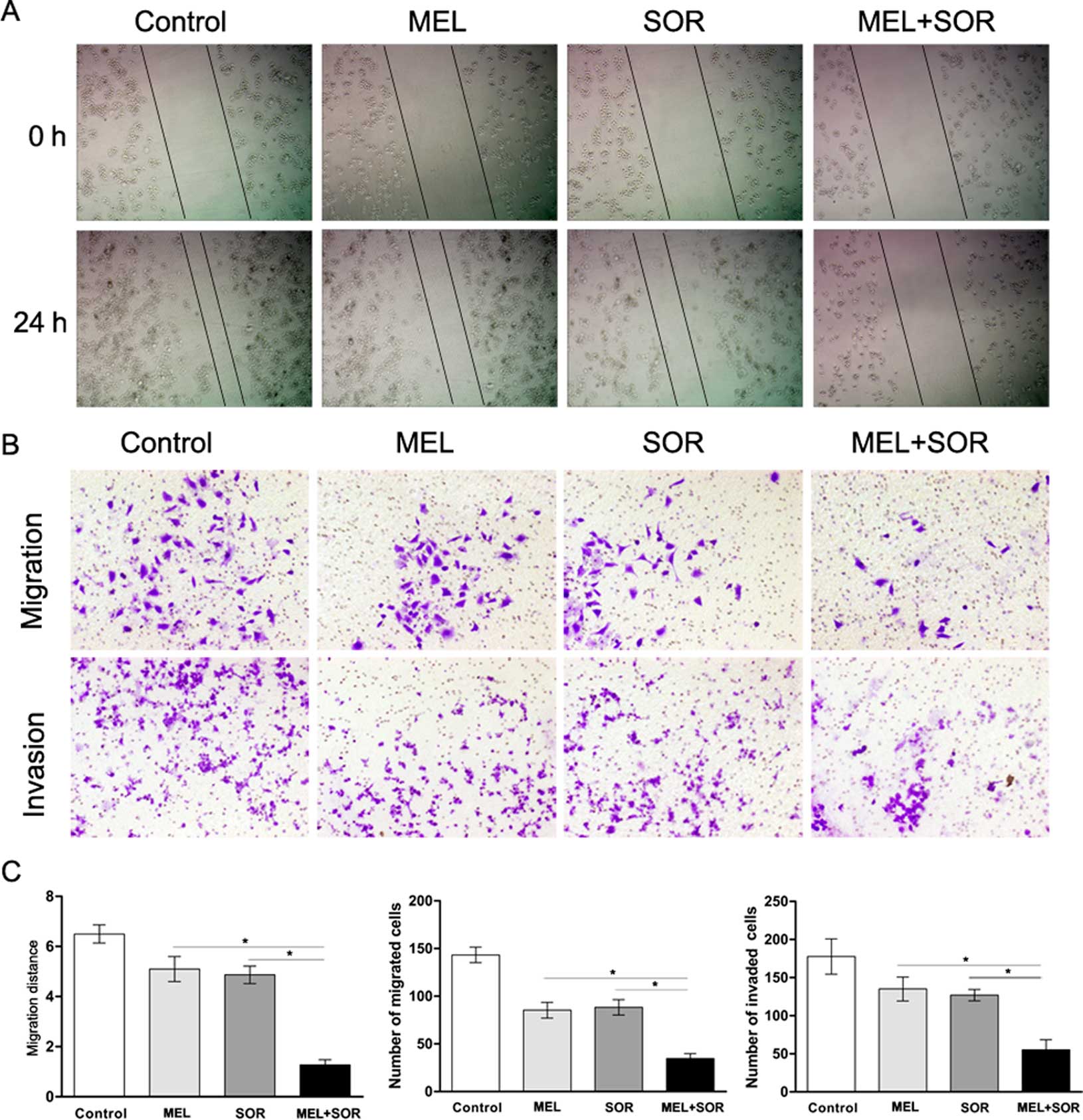
Given the association of HCC with a high degree of
invasion and metastasis, we investigated whether the migratory
potential of SMMC-7721 cells could be affected by exposure to MEL
or SOR alone or in combination. As determined by scratch motility
assay, MEL or SOR treatment alone induced a partial inhibition of
migration whereas the combined treatment with MEL and SOR notably
inhibited the migratory potential of SMMC-7721 cells (Fig. 2A and C). Furthermore, we applied
SMMC-7721 cells to a migration and invasion assay and the results
were consistent with those of the scratch assay (Fig. 2B and C).
Combination of MEL with SOR induces cell
cycle arrest and apoptosis in SMMC-7721 cells
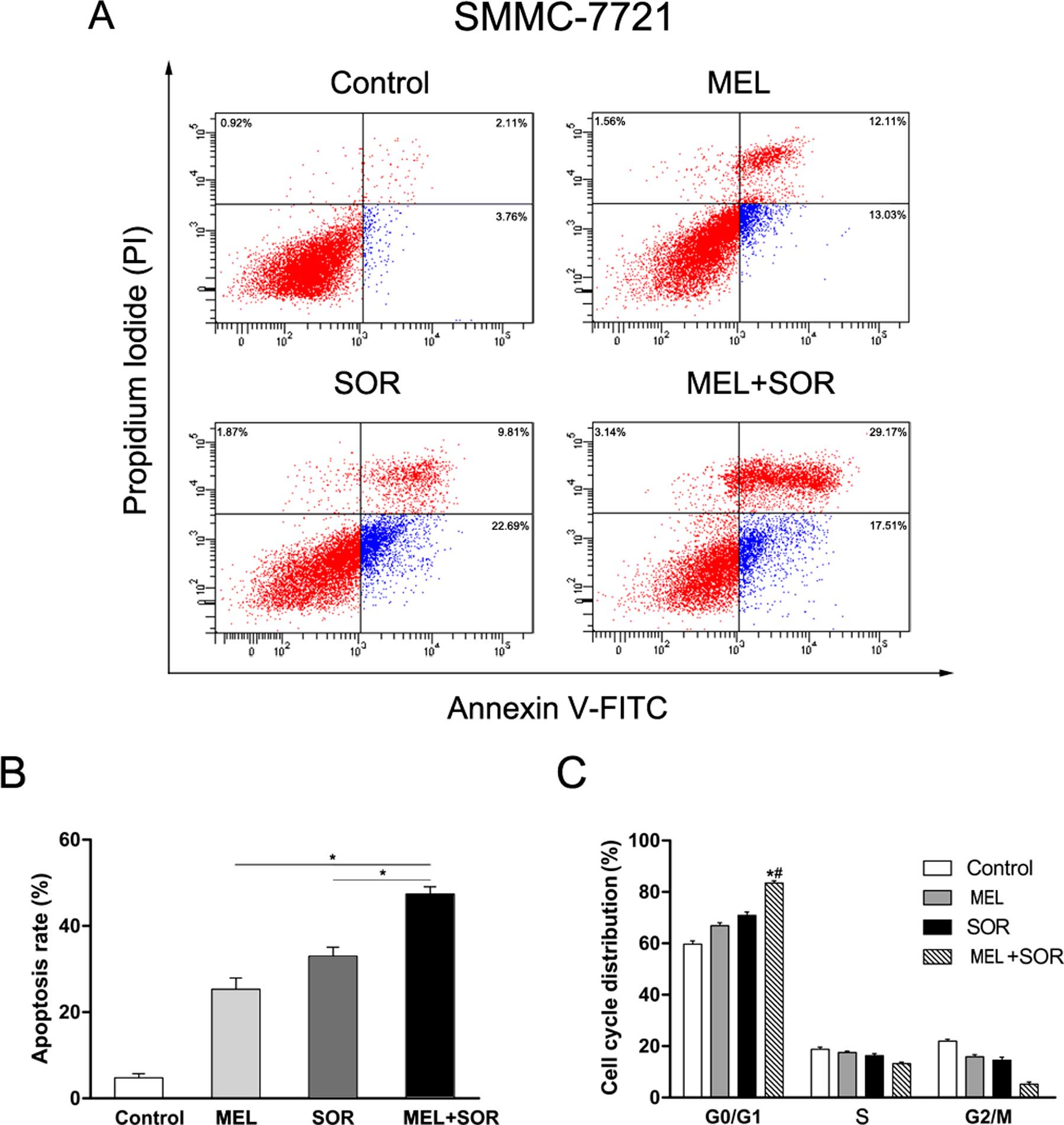
Given the superior synergistic interactions observed
between MEL and SOR, we investigated the potential effects on cell
apoptosis mediated by these combinations. Apoptotic cell death
induction was analyzed by flow cytometry at 24 h after SMMC-7721
cells were treated-with either MEL or SOR alone or in combination.
As shown in Fig. 3A and B, MEL and
SOR as a single agent led to apoptosis in SMMC-7721 cells. We also
observed the MEL+SOR combination significantly increased apoptotic
cell death compared with MEL or SOR as a single agent. Next, we
used flow cytometry to evaluate the potential effects of MEL and
SOR on the cell cycle distribution of SMMC-7721 cells. We found
that cell cycle arrest at the G0/G1 phase was increased with
treatment of MEL or SOR compared with the control group (Fig. 3C). In addition, the MEL+SOR
combination led to enhanced accumulation of cells in the G0/G1
phase compared to the single agents. These data revealed an
additive mechanism of the MEL+SOR combination inducing cell death
via apoptosis.
Combination of MEL with SOR induces ER
stress in SMMC-7721 cells
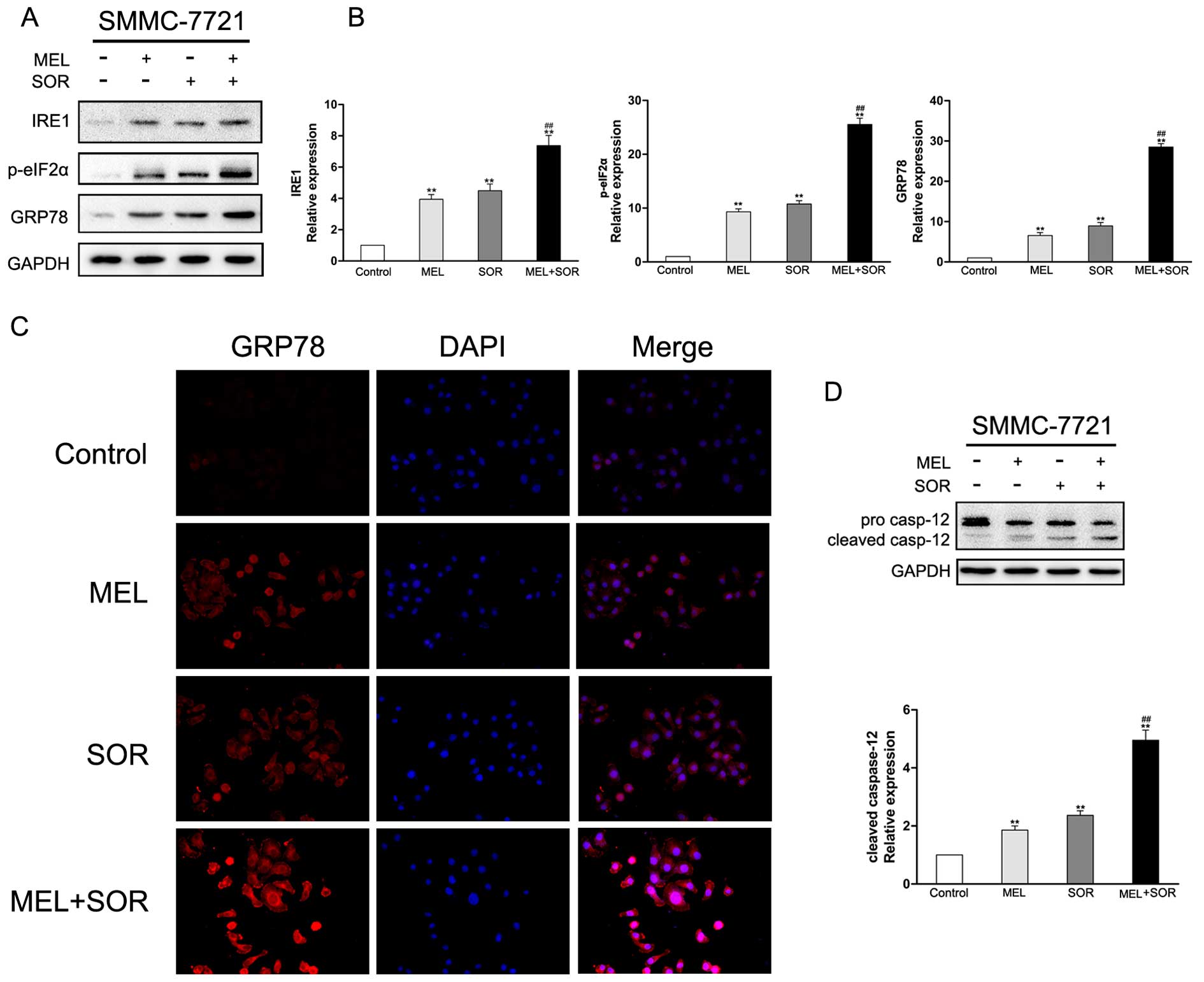
Previous studies have demonstrated that COX-2
inhibitor and SOR as single agent treatments induced ER stress
(19–22). To ascertain whether the MEL+SOR
combination treatment enhanced ER stress in HCC cell lines, certain
ER-specific signals were measured. Immunoblotting analysis results
indicated that the levels of IRE1, p-eIF2α and GRP78 were
upregulated in response to both MEL and SOR alone, and these ER
stress marker were significantly increased by the MEL+SOR
combination treatment (Fig. 4A and
B). To ensure the observations that ER stress-associated
markers were increased in SMMC-7721 cells after exposed to MEL or
SOR alone or in combination, GRP78 were visualized by
immunofluorescence staining. As shown in Fig. 4, immunofluorescence staining of
GRP78 was partially increased after MEL or SOR single treatment.
However, the combined treatment with MEL and SOR markedly increased
GRP78 of SMMC-7721 cells. Several studies have revealed that
caspase-12 is activated by continuous ER stress and plays a key
role in leading to cell death not via the cytochrome
c-dependent pathway (23).
To investigate the involvement of ER stress in the MEL+SOR
combination treatment-induced toxicity and explore potential
mechanisms in the present study, western blotting assay was used to
detect expression and distribution of caspase-12 proteins in
SMMC-7721 cells. As shown in Fig.
4D, the activation of the caspase-12 protein was significantly
increased in SMMC-7721 cells that were treated with the MEL+SOR
combination compared to the single agents, in agreement with cell
death assays. These data indicated that ER impairment targeted the
process of apoptosis.
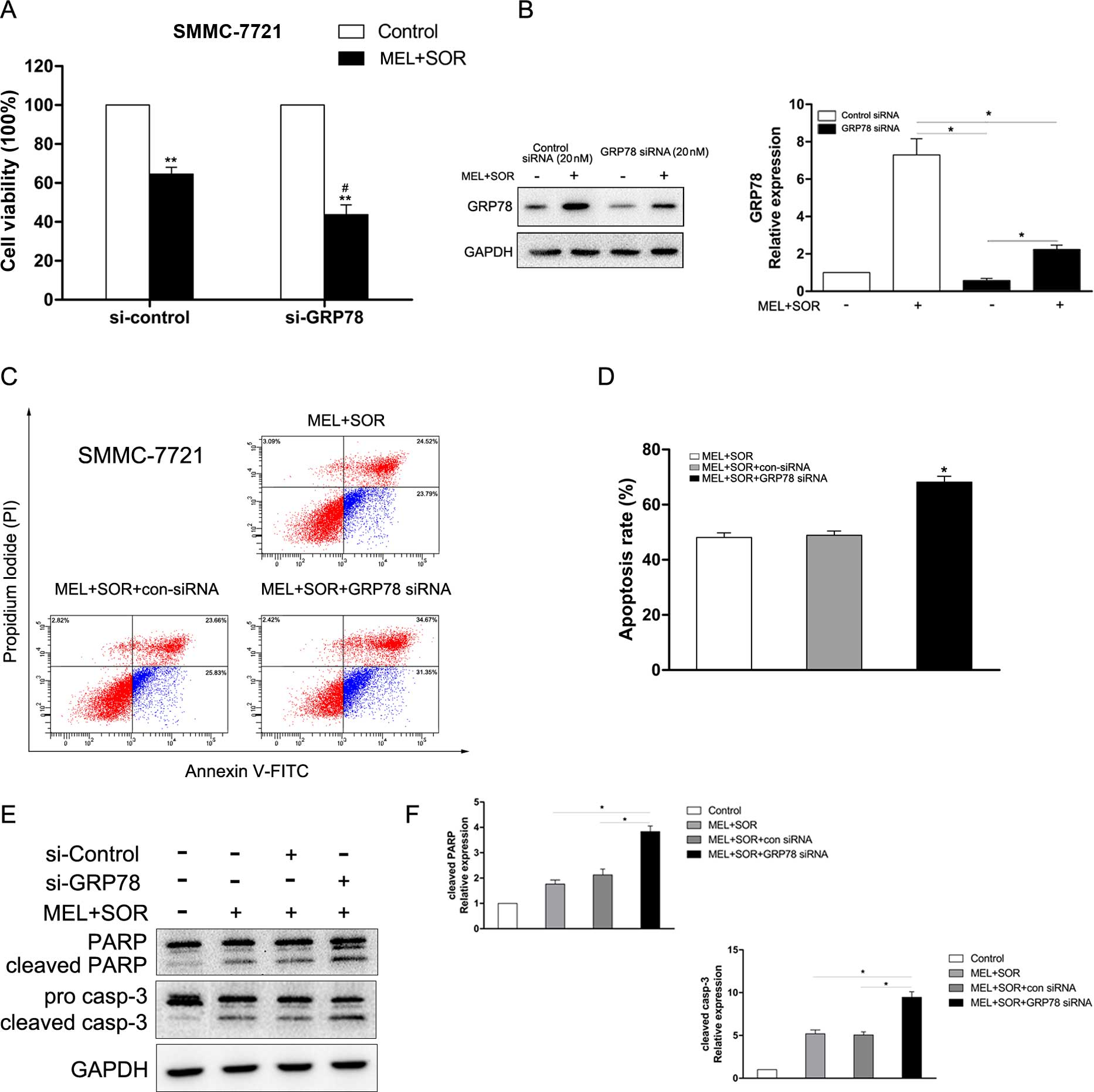
Involvement of GRP78 in combined MEL with
SOR treatment-induced apoptosis
GRP78, one of the most important protective
mechanisms induced by UPR has been demonstrated to be associated
with chemoresistance (24). In the
present study, we explored the role of GRP78 in the MEL+SOR
combination treatment-induced apoptosis. As shown in Fig. 5A, transfection of GRP78 siRNA, the
MEL+SOR combination treatment significantly reduced cell viability,
as expected. Additionally, transfection of GRP78 siRNA which
downregulated the level of GRP78 protein (Fig. 5B), notably strengthened the increase
of cell apoptosis (Fig. 5C and D)
and the cleavage of PARP and caspase-3 (Fig. 5E and F) in the MEL+SOR
combination-treated SMMC-7721 cells. These data revealed that GRP78
exerts a protective function in HCC cells to promote drug
resistance.
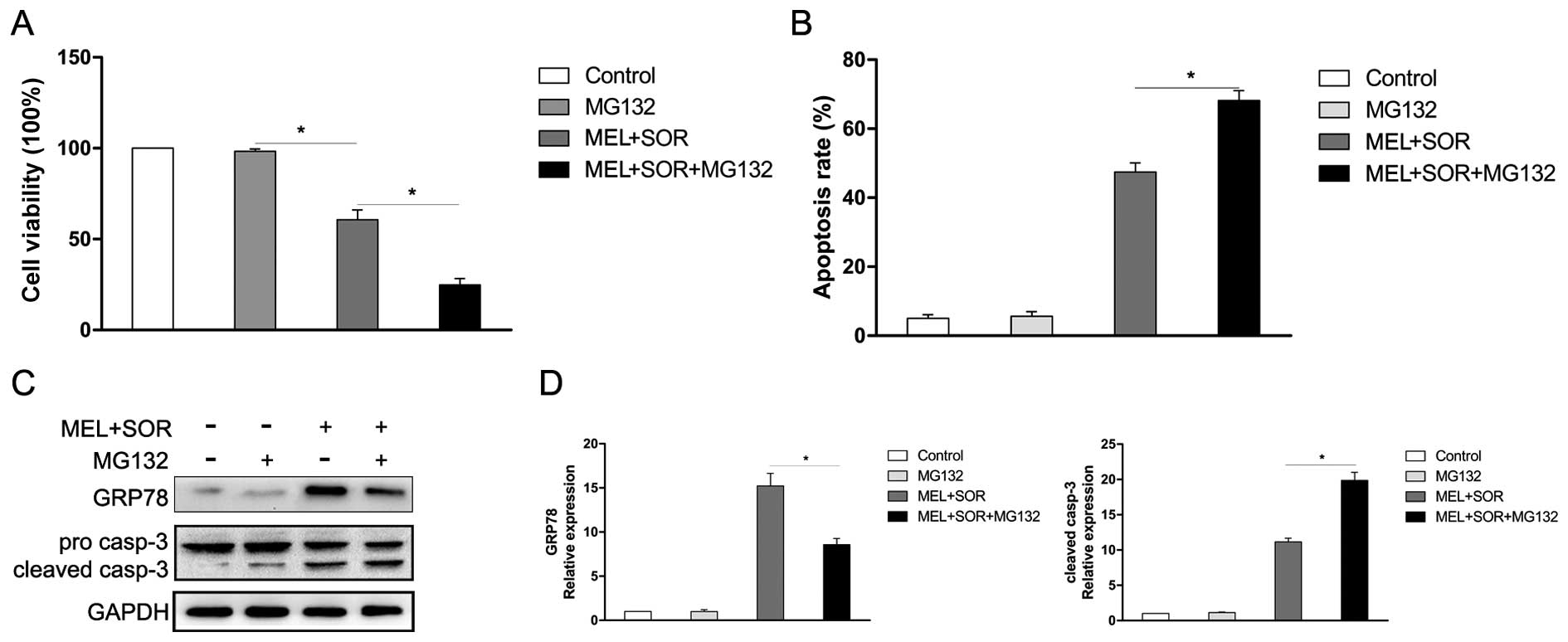
MG132 enhances the MEL and SOR
combination treatment-induced apoptosis in SMMC-7721 cells
Previous studies have revealed that the proteasome
pathway exerts a crucial role in the degradation of unfolded
protein (25,26). In the present study, we hypothesized
that inhibition of proteasome enhances the MEL+SOR combination
treatment-induced SMMC-7721 cells apoptosis attributed to the
accumulation of unfolded protein. To verify our assumption, the
proteasome inhibitor MG132, was used to evaluate the combination
effect of the MEL+SOR on human SMMC-7721 cells. Our results showed
that when exposed to low-dose (1 μM), MG132 mildly affected
cell viability. However, the MEL+SOR+MG132 combination treatment
significantly enhanced the cell toxicity (Fig. 6A) and apoptosis (Fig. 6B). Furthermore, MG132 significantly
suppressed the MEL+SOR combination treatment-induced GRP78
expression and enhanced the cleavage of caspase-3 (Fig. 6C and D). These data revealed that
proteosome inhibitor MG132 enhanced the MEL+SOR combination
treatment-induced apoptosis.
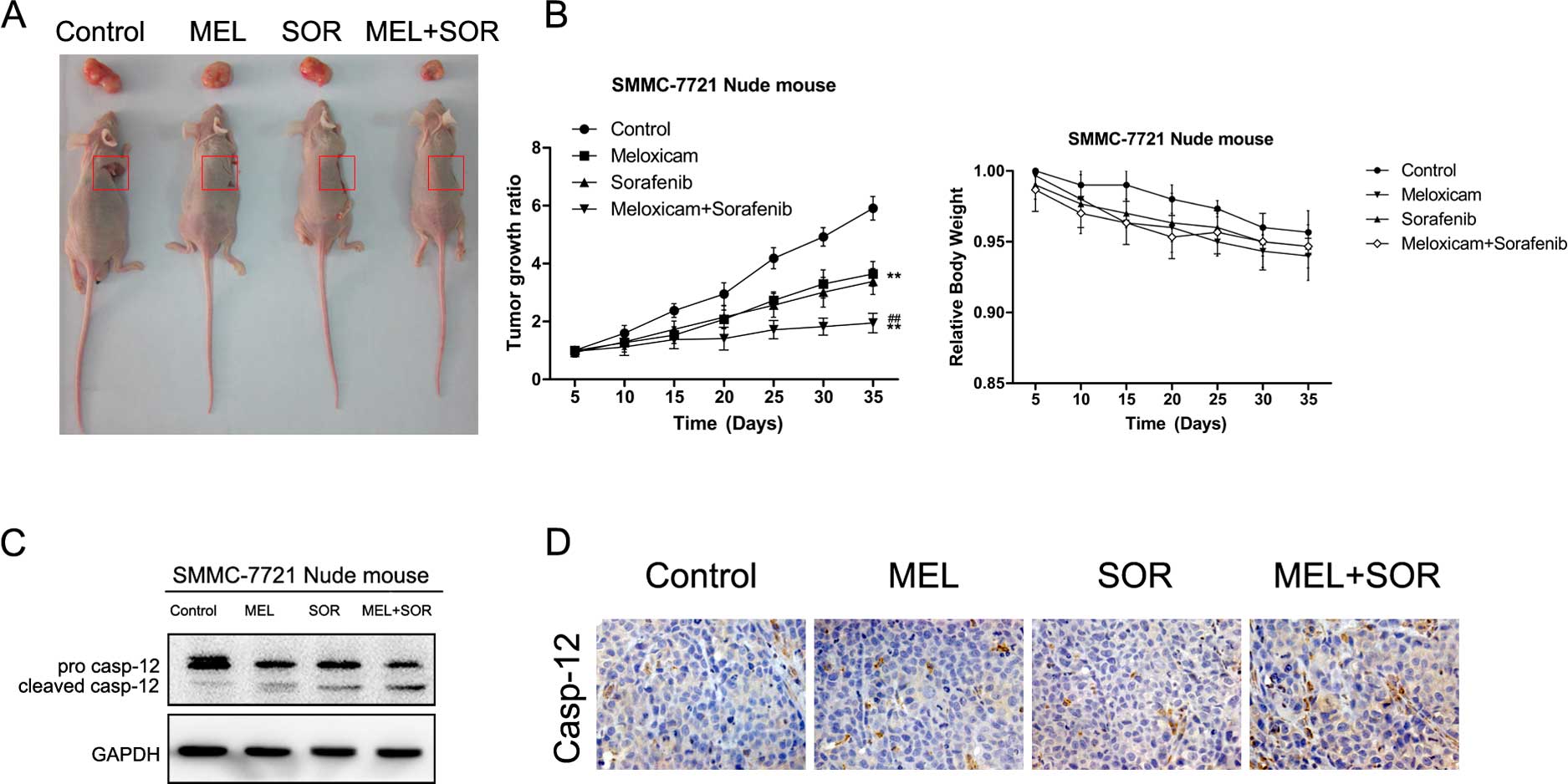
Combination of MEL with SOR arrests tumor
growth in vivo
Due to the superior antitumor effects of the MEL+SOR
combination treatment in vitro, we explored whether the
MEL+SOR combination treatment inhibited tumor growth in
vivo. As shown in Fig. 7A and
B, the MEL+SOR combination treatment exerted marked antitumor
activity in SMMC-7721 xenograft tumors compared to the single
agents. However, we found that the combination treatment caused
only mild weight change in the in vivo models. Furthermore,
we used western blotting and immunohistochemistry to analyze tumor
xenografts. The results suggested that the MEL+SOR combination
treatment notably activated the ER stress-related apoptosis in
SMMC-7721 cell-derived tumors (Fig. 7C
and D). In conclusion, our data revealed that the MEL+SOR
combination treatment significantly arrests tumor growth in
vivo via ER stress-associated regulatory mechanisms.
Discussion
Hepatocellular carcinoma (HCC), a hypervascular
tumor type with characteristic of high levels of neovascularization
and angiogenesis, exerts effects in the growth and progression
which needs interacting approaches for effective therapy (27,28).
Due to the association of single agents with treatment resistance,
we considered that the combination therapy increased the lethality
in HCC. Sorafenib (SOR) has been applied as the standard
therapeutic strategy for advanced HCC patients. In contrast, the
selective COX-2 inhibitor has been demonstrated to exert antitumor
effects in various types of tumors including HCC (12,13,21,29,30).
Thus, in the present study, we investigated whether the
combinations of meloxicam (MEL)+SOR led to more superior antitumor
effects than MEL or SOR alone in human SMMC-7721 cells. Our results
revealed that either MEL or SOR alone reduce cell viability and
colony formation and induce cell cycle arrest and apoptosis.
However, the MEL+SOR combination exhibited more potent antitumor
effects in terms of cytotoxicity and apoptotic induction via ER
stress in human SMMC-7721 cells. GRP78 knockdown by siRNA or
proteasome inhibitor significantly enhanced the MEL+SOR combination
treatment-induced apoptosis.
ER exerts a key role in regulating protein
synthesis, folding and trafficking. A large number of signal
pathways have been demonstrated to disrupt the ER function and
induce dysfunction of UPR, resulting in ER stress. The initial aim
of UPR is to restore ER homeostasis, however, when pro-survival
responses failed, these signaling pathways ultimately led to cell
apoptosis (14,31–33).
GRP78, the ER molecular chaperone, exerts a crucial role in protein
folding and assembly (34).
Perturbation of ER homeostasis leads to activation of ER stress
which results in GRP78 dissociation (35,36).
Furthermore, several studies have reported that GRP78 is associated
with chemoresistance in cancer therapy (24,37,38).
In the present study, the MEL+SOR combination treatment led to ER
stress in human SMMC-7721 cells and is associated with the increase
of IRE1, p-eIF2α, GRP78 and activation of caspase-12. Silencing
GRP78 enhanced the cytotoxic and apoptotic effect of MEL+SOR
combination treatment in SMMC-7721 cells. Therefore, it is
concluded that GRP78 plays a protective function in HCC cells to
promote drug resistance. GRP78 knockdown by siRNA notably increased
the susceptibility to MEL+SOR in SMMC-7721 cells.
Certain studies have revealed that the ubiquitin
proteosome pathway exerts a crucial role in intracellular protein
degradation by maintaining ER homeostasis when cells encounter the
UPR (25). In the present study,
our results showed that combined treatment with proteosome
inhibitor MG132 significantly enhanced the MEL+SOR-induced
cytotoxicity and apoptosis with concomitant downregulation of GRP78
and activation of caspase-3.
In conclusion, these data demonstrated that the
MEL+SOR combination treatment notably reduced cell viability and
induced apoptosis in human SMMC-7721 cells. GRP78 knockdown or by
proteasome inhibitor MG132 significantly enhances the MEL+SOR
combination treatment-induced SMMC-7721 cell apoptosis. These
findings provide a basis for and warrant future study to
investigate the combination of MEL+SOR therapy for the treatment of
drug resistant tumors with targeted therapy.
Acknowledgments
The present study was supported from the National
Natural Scientific Foundation of China (nos. 30972890 and
81172331), the Shandong Provincial Science and Technology
Development Planning, China (2010GSF10230), and the Medicine and
Health Science Technology of Shandong Province, China (2013WS0145).
Thanks to Dr Edward C. Mignot, Shandong University for linguistic
advice.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Asghar U and Meyer T: Are there
opportunities for chemotherapy in the treatment of hepatocellular
cancer? J Hepatol. 56:686–695. 2012. View Article : Google Scholar
|
|
3
|
Wilhelm SM, Carter C, Tang L, Wilkie D,
McNabola A, Rong H, Chen C, Zhang X, Vincent P, McHugh M, et al:
BAY 43-9006 exhibits broad spectrum oral antitumor activity and
targets the RAF/MEK/ERK pathway and receptor tyrosine kinases
involved in tumor progression and angiogenesis. Cancer Res.
64:7099–7109. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu L, Cao Y, Chen C, Zhang X, McNabola A,
Wilkie D, Wilhelm S, Lynch M and Carter C: Sorafenib blocks the
RAF/MEK/ERK pathway, inhibits tumor angiogenesis, and induces tumor
cell apoptosis in hepatocellular carcinoma model PLC/PRF/5. Cancer
Res. 66:11851–11858. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Llovet JM, Ricci S, Mazzaferro V, Hilgard
P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A,
et al SHARP Investigators Study Group: Sorafenib in advanced
hepatocellular carcinoma. N Engl J Med. 359:378–390. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thun MJ, Henley SJ and Patrono C:
Nonsteroidal anti-inflammatory drugs as anticancer agents:
Mechanistic, pharmacologic, and clinical issues. J Natl Cancer
Inst. 94:252–266. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lönnroth C, Andersson M, Asting AG,
Nordgren S and Lundholm K: Preoperative low dose NSAID treatment
influences the genes for stemness, growth, invasion and metastasis
in colorectal cancer. Int J Oncol. 45:2208–2220. 2014.PubMed/NCBI
|
|
8
|
Ding N, Cui XX, Gao Z, Huang H, Wei X, Du
Z, Lin Y, Shih WJ, Rabson AB, Conney AH, et al: A triple
combination of atorvastatin, celecoxib and tipifarnib strongly
inhibits pancreatic cancer cells and xenograft pancreatic tumors.
Int J Oncol. 44:2139–2145. 2014.PubMed/NCBI
|
|
9
|
Zhivkova T, Alexandrova R, Dyakova L,
Georgieva M, Miloshev G, Culita DC, Marinescu G, Patron L and
Alexandrov M: P7.05. Metal complexes of meloxicam and isoxicam
decrease viability and proliferation of virus-transformed cancer
cells. Ann Oncol. 26(Suppl 2): ii322015. View Article : Google Scholar
|
|
10
|
Hassan MH, El-Beshbishy HA, Aly H, Attia
SM, Bahashwan SA and Ghobara MM: Modulatory effects of meloxicam on
cardiotoxicity and antitumor activity of doxorubicin in mice.
Cancer Chemother Pharmacol. 74:559–569. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Arantes-Rodrigues R, Pinto-Leite R,
Ferreira R, Neuparth MJ, Pires MJ, Gaivão I, Palmeira C, Santos L,
Colaço A and Oliveira P: Meloxicam in the treatment of in vitro and
in vivo models of urinary bladder cancer. Biomed Pharmacother.
67:277–284. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li J, Chen X, Dong X, Xu Z, Jiang H and
Sun X: Specific COX-2 inhibitor, meloxicam, suppresses
proliferation and induces apoptosis in human HepG2 hepatocellular
carcinoma cells. J Gastroenterol Hepatol. 21:1814–1820. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong X, Li R, Xiu P, Dong X, Xu Z, Zhai B,
Liu F, Jiang H, Sun X, Li J, et al: Meloxicam executes its
antitumor effects against hepatocellular carcinoma in
COX-2-dependent and -independent pathways. PLoS One. 9:e928642014.
View Article : Google Scholar
|
|
14
|
Tabas I and Ron D: Integrating the
mechanisms of apoptosis induced by endoplasmic reticulum stress.
Nat Cell Biol. 13:184–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu J, Chen M, Chen N, Ma A, Zhu C, Zhao
R, Jiang M, Zhou J, Ye L, Fu H, et al: Glycyrrhetinic acid induces
G1-phase cell cycle arrest in human non-small cell lung cancer
cells through endoplasmic reticulum stress pathway. Int J Oncol.
46:981–988. 2015.PubMed/NCBI
|
|
16
|
Xu C, Bailly-Maitre B and Reed JC:
Endoplasmic reticulum stress: Cell life and death decisions. J Clin
Invest. 115:2656–2664. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meusser B, Hirsch C, Jarosch E and Sommer
T: ERAD: The long road to destruction. Nat Cell Biol. 7:766–772.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cao SS and Zhen YS: Potentiation of
antimetabolite antitumor activity in vivo by dipyridamole and
amphotericin B. Cancer Chemother Pharmacol. 24:181–186. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Inamoto T and Azuma H: Sorafenib increases
endoplasmic reticulum (ER) stress in concert with vorinostat.
Cancer Biol Ther. 12:10182011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi YH, Ding ZB, Zhou J, Hui B, Shi GM, Ke
AW, Wang XY, Dai Z, Peng YF, Gu CY, et al: Targeting autophagy
enhances sorafenib lethality for hepatocellular carcinoma via ER
stress-related apoptosis. Autophagy. 7:1159–1172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suzuki K, Gerelchuluun A, Hong Z, Sun L,
Zenkoh J, Moritake T and Tsuboi K: Celecoxib enhances
radiosensitivity of hypoxic glioblastoma cells through endoplasmic
reticulum stress. Neurooncol. 15:1186–1199. 2013.
|
|
22
|
Kim SJ, Ha GH, Bae JH, Kim GR, Son CH,
Park YS, Yang K, Oh SO, Kim SH and Kang CD: COX-2- and endoplasmic
reticulum stress-independent induction of ULBP-1 and enhancement of
sensitivity to NK cell-mediated cytotoxicity by celecoxib in colon
cancer cells. Exp Cell Res. 330:451–459. 2015. View Article : Google Scholar
|
|
23
|
Momoi T: Caspases involved in ER
stress-mediated cell death. J Chem Neuroanat. 28:101–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Virrey JJ, Dong D, Stiles C, Patterson JB,
Pen L, Ni M, Schönthal AH, Chen TC, Hofman FM and Lee AS: Stress
chaperone GRP78/BiP confers chemoresistance to tumor-associated
endothelial cells. Mol Cancer Res. 6:1268–1275. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Egger L, Madden DT, Rhême C, Rao RV and
Bredesen DE: Endoplasmic reticulum stress-induced cell death
mediated by the proteasome. Cell Death Differ. 14:1172–1180. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Komatsu S, Miyazawa K, Moriya S, Takase A,
Naito M, Inazu M, Kohno N, Itoh M and Tomoda A: Clarithromycin
enhances bortezomib-induced cytotoxicity via endoplasmic reticulum
stress-mediated CHOP (GADD153) induction and autophagy in breast
cancer cells. Int J Oncol. 40:1029–1039. 2012.
|
|
27
|
Dupuy E, Hainaud P, Villemain A,
Bodevin-Phèdre E, Brouland JP, Briand P and Tobelem G: Tumoral
angiogenesis and tissue factor expression during hepatocellular
carcinoma progression in a transgenic mouse model. J Hepatol.
38:793–802. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Geis T, Döring C, Popp R, Grossmann N,
Fleming I, Hansmann ML, Dehne N and Brüne B: HIF-2alpha-dependent
PAI-1 induction contributes to angiogenesis in hepatocellular
carcinoma. Exp Cell Res. 331:46–57. 2015. View Article : Google Scholar
|
|
29
|
Yusup G, Akutsu Y, Mutallip M, Qin W, Hu
X, Komatsu-Akimoto A, Hoshino I, Hanari N, Mori M, Akanuma N, et
al: A COX-2 inhibitor enhances the antitumor effects of
chemotherapy and radiotherapy for esophageal squamous cell
carcinoma. Int J Oncol. 44:1146–1152. 2014.PubMed/NCBI
|
|
30
|
Agarwal V, Hodgkinson VC, Eagle GL, Scaife
L, Lind MJ and Cawkwell L: Proteomic (antibody microarray)
exploration of the molecular mechanism of action of the specific
COX-2 inhibitor DuP 697. Int J Oncol. 42:1088–1092. 2013.PubMed/NCBI
|
|
31
|
Rutkowski DT and Kaufman RJ: A trip to the
ER: Coping with stress. Trends Cell Biol. 14:20–28. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dolai S, Pal S, Yadav RK and Adak S:
Endoplasmic reticulum stress-induced apoptosis in Leishmania
through Ca2+-dependent and caspase-independent
mechanism. J Biol Chem. 286:13638–13646. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fribley AM, Miller JR, Brownell AL,
Garshott DM, Zeng Q, Reist TE, Narula N, Cai P, Xi Y, Callaghan MU,
et al: Celastrol induces unfolded protein response-dependent cell
death in head and neck cancer. Exp Cell Res. 330:412–422. 2015.
View Article : Google Scholar
|
|
34
|
Li J and Lee AS: Stress induction of
GRP78/BiP and its role in cancer. Curr Mol Med. 6:45–54. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kulkarni P, Rajagopalan K, Yeater D and
Getzenberg RH: Protein folding and the order/disorder paradox. J
Cell Biochem. 112:1949–1952. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu J and Kaufman RJ: From acute ER stress
to physiological roles of the Unfolded Protein Response. Cell Death
Differ. 13:374–384. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee AS: GRP78 induction in cancer:
Therapeutic and prognostic implications. Cancer Res. 67:3496–3499.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ermakova SP, Kang BS, Choi BY, Choi HS,
Schuster TF, Ma WY, Bode AM and Dong Z: (−)-Epigallocatechin
gallate overcomes resistance to etoposide-induced cell death by
targeting the molecular chaperone glucose-regulated protein 78.
Cancer Res. 66:9260–9269. 2006. View Article : Google Scholar : PubMed/NCBI
|