Introduction
Lipocalin 2 (LCN2) is a member of lipocalin family
that binds and transports a small lipophilic ligand, sharing a
highly conserved tertiary structure (1). The binding ligands of lipocalin
include retinoic acid, progesterone and prostaglandin. The function
of LCN2 is known to be associated with cellular iron uptake,
antibacterial activity and epithelial cell differentiation
(2). Alpízar-Alpízar et al
reported that LCN2 is upregulated in gastric mucosa infected with
Helicobacter pylori known to be associated with gastric
cancer. They proposed that the investigation of the connection of
LCN2 and gastric cancer progression should be carried out (3).
Matrix metalloproteinase 9 (MMP9) is a member of
MMPs that break down the basement membranes through the degradation
of type IV collagen, exposing cryptic sites within matrix and
allowing cancer cell invasion. Degradation of extracellular matrix
(ECM) in tissue of the tumor is a principal process of cancer
invasion and metastasis (4,5). Particularly, MMP2 and MMP9 have been
in focus recently in experimental models (6,7). Some
in vitro and in vivo studies showed that MMP level
was possibly associated with cancer invasion and metastasis acting
in colon cancer. Sier et al reported that the levels of MMP2
and MMP9 were enhanced in gastric cancer tissue compared to
adjacent normal tissue (6), and the
fact that polymorphism of alle of MMP promotor is significantly
associated with gastric cancer invasion and metastasis, was
reported (8).
Hepatocyte growth factor (HGF), produced primarily
by mesenchymal cells, was known to have a role with a different
activity of inducing epithelial cell dissociation. HGF has been
shown to be an important factor of acting cancer cell invasion with
interaction by tumor stromal tissue. We studied HGF-induced
expressing genes in gastric cancer, and found that LCN2 also is
expressed in NUGC3 and MKN28 cells treated with HGF using 17K human
cDNA microarrays.
LCN2 performs epithelial-to-mesenchymal transition
via MMP9 dependent pathway. MMP9 is able to facilitate the
extracellular matrix remodeling and bind the LCN2 with high
affinity. LCN2 can be found as a monomer, homodimer, heterodimer
with MMP9 and the binding of LCN2 to MMP9 promotes MMP9 activation
and blocks the MMP9 autodegradation (9). This action restructures ECM persisting
the action of invading the tissue, which facilitates metastasis
(10).
High molecule of LCN2 was detected in urine of
breast cancer patients (11). In
vitro data proved that LCN2 is principal factor of
tumorigenesis and metastasis in breast cancer (12). The fact that in gastric cancer high
level of LCN2 was detected compared to adjacent control tissue, and
complexes of LCN2 and MMP9 are increased (13). However, the mechanisms of regulation
of LCN2 in MMP9 activity or stabilization is not known.
In the present study, we examined the effect LCN2
knockdown on in vitro proliferation and invasiveness, and
MMP9 regulation to identify the definite role of LCN2 in cancer
proliferation and invasion in gastric cancer cell line. Also, we
performed experiments to confirm the pathway facilitating the
regulation MMP9 by LCN2.
Materials and methods
Cell culture
We used two human gastric cell lines in our
experiments: NUGC3 and MKN28, which were obtained from the Korea
Cell Line Bank. These cells were maintained on plastic in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS), 1 mM sodium pyruvate, 0.1 mM nonessential
amino acids, 2 mM L-glutamine, two-fold vitamin solution and 50
U/ml penicillin/streptomycin (Life Technologies, Inc.,
Gaithersburg, MD, USA). Unless otherwise noted, the cells were
passaged and removed at 70–80% confluency.
Reagents and antibodies
Horseradish peroxidase-conjugated anti-mouse and
anti-rabbit antibodies were purchased from Bio-Rad Laboratories
(Philadelphia, PA, USA). Recombinant human HGF and pyrrolidine
dithiocarbamate (PDTC) were purchased from R&D Systems, Inc.
(Minneapolis, MN, USA). LCN2 was purchased from Abnova (Taipei,
Taiwan). MMP9 was purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). PD098059 was purchased from BIOMOL Research
Laboratories, Inc. (Butler Pike, PA, USA). SB203580 and LY294002
were purchased from Calbiochem Inc. (San Diego, CA, USA). NFκB
antibodies were purchased from Cell Signaling Technology (Beverly,
MA, USA).
Semi-quantitative reverse
transcription-polymerase chain reaction (RT-PCR)
Complementary DNA (cDNA) was synthe-sized from total
RNA using M-MLV Reverse Transcriptase (Promega Corp., Madison, WI,
USA) by the oligo(dT) priming method in a 10 μl reaction
mixture. PCR was performed in 10 μl reaction volume
containing 10 mM Tris-HCl pH 8.5, 50 mM KCl, 1 μl cDNA, 200
μM dNTPs, 1 mM MgSO4, 1 U Platinum Pfx Taq
polymerase and 2 μM primers. The reactions were the initial
denaturation at 95°C for 4 min; 27 cycles at 94°C for 15 sec, and
60°C for 15 sec, and 72°C for 30 sec; and the final extension at
72°C for 10 min. The PCR products were separated on a 1.5% agarose
gel containing ethidium bromide and visualized on a UV
transilluminator.
cDNA microarray analysis
The cDNA microarray, containing a set of 17,448
sequence-verified human cDNA clones, was provided by GenomicTree,
Inc. (Daejeon, Korea). cDNA microarray experiments were performed
as described by Yang et al (14). Briefly, total RNA (100 μg)
was reverse transcribed in the presence of Cy3-dUTP or Cy5-dUTP (25
mM stock; NEN Life Science Products, Boston, MA, USA) at 42°C for 2
h. The labeled cDNA was then hybridized with the cDNA microarray at
65°C for 16 h. The hybridized slides were washed, scanned with an
Axon 4000B scanner, and analyzed using GenePix Pro 4.0 (both from
Axon Instruments). Raw data were normalized and analyzed using
GeneSpring 6.0 (Silicon Genetics). Genes were filtered according to
their intensities in the control channel. When control channel
values were below 80 the samples were considered as unreliable.
Intensity-dependent normalization (LOWESS) was performed, where the
ratio was reduced to the residual of LOWESS fit of the intensity
vs. ratio curve. Average normalized ratios were calculated by
dividing the averaged normalized signal channel intensity by the
averaged normalized control channel intensity. The Welch ANOVA test
was performed for P-values ≤0.1 of 0.05 to identify genes
differentially expressed samples. Correlation analysis was
performed using Pearson's correlation (−1 to 1). Spots showing
changes of 2-fold or more were considered significant.
Western blot analysis
Cells were harvested and incubated with a lysis
buffer [50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1 mM EDTA, 1% Triton
X-100, 10% glycerol, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1
mM sodium vanadate and 5 mM NaF] with protease inhibitors and
centrifuged at 15,000 rpm and 4°C for 10 min. Proteins (50
μg) were separated on 10% SDS-polyacrylamide gels and
transferred to nitrocellulose membranes. The membranes were soaked
with 5% non-fat dried milk in TTBS [10 mM Tris-HCl (pH 7.5), 150 mM
NaCl and 0.05% Tween-20] for 30 min and then incubated overnight
with a primary antibody at 4°C. After washing 6 times with TTBS for
5 min, the membranes were incubated with a horseradish
peroxidase-conjugated secondary antibody for 90 min at 4°C. The
membranes were rinsed 3 times with TTBS for 30 min and
antigen-antibody complexes was detected using the enhanced
chemiluminescence detection system.
Zymography for MMP9
Culture supernatants were denatured in the absence
of reducing agent and were electrophoresed in 10% polyacrylamide
gel containing 0.1% (W/V) gelatin for MMP9. The gel was incubated
at room temperature for 2 h in the presence of 2.5% Triton X-100
and subsequently at 37°C overnight in a buffer containing 10 mM
CaCl2, 0.15 M NaCl and 50 mM Tris (pH 7.5). The gel was
then stained for protein with 0.25% Coomassie brilliant blue
solution in methanol:acetic acid:water (4:1:5) and destained in the
same solution without the dye; the enzyme activity was detected as
the negatively-stained regions. Zymographic analyses were performed
in at least 3 independent experiments.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
Cells (1,500/well) were seeded in 96-well plates in
DMEM supplemented with 5% FBS and incubated for 24 h. The cells
were then serum-starved for 24 h and treated for 72 h with or
without HGF (10 ng/ml). At the end of this incubation period, 50
μl of 2 mg/ml MTT solution was added and the cells were
allowed to incubate for 3 h at 37°C. The supernatant was carefully
removed by aspiration, and the converted dye was dissolved with 100
μl DMSO. The plates were placed in a microplate shaker for 5
min, and the absorbance was measured at 570 nm using a Bio-Rad
multiscan plate reader.
Standard two chamber invasion assay
Control and transfected cells (1×104)
were placed in the upper chamber of a Matrigel migration chamber
with 0.8-μ pores (Fisher Scientific, Houston, TX, USA) in
media containing 5% FBS with or without HGF (10 ng/ml). After
incubation for 48 h, the cells were fixed and stained using the
HEMA 3 stain set (Curtin Matheson Scientific, Inc., Houston, TX,
USA) according to the manufacturer's instructions. The stained
filter membrane was cut and placed on a glass slide. The migrated
cells were counted under light microscopy (10 fields at x200
magnification).
LCN2 knockdown with short hairpin
RNA
The human LCN2-specific short hairpin RNA (shRNA)
expression vector (LCN2-shRNA, RHS3979-201778075) containing
LCN2-targeted shRNA sequence (AAACCCAGGGCTGCCTTGGA AAAG) was
purchased from Open Biosystems (Huntsville, AL, USA). NUGC3 and
MKN28 cells were transfected with LCN2-shRNA using Lipofectamine
(Life Technologies, Inc.). Clonal selection was conducted by
culturing with puromycin (25 μg/ml) followed by serial
dilution of the cells. Stable transfectant clones with low
expression of the target genes were identified by western blot
analysis.
Chromatin immunoprecipitation assay
The chromatin immunoprecipitation (ChIP) assay was
carried out using the ChIP assay kit (Upstate Biotechnology,
Waltham, MA, USA) following the manufacturer's directions. Briefly,
cells were fixed with 1% formaldehyde at 37°C for 10 min. Cells
were washed twice with ice-cold PBS with protease inhibitors (1 mM
PMSF, 1 mg/ml aprotinin and 1 mg/ml pepstatin A), scraped and
pelleted by centrifugation at 4°C. Cells were resuspended in a
lysis buffer (1% SDS, 10 mM EDTA and 50 mM Tris-HCl, pH 8.1),
incubated for 10 min on ice, and sonicated to shear DNA. After
sonication, lysate was centrifuged for 10 min at 13,000 rpm at 4°C.
The supernatant was diluted in ChIP dilution buffer (0.01% SDS, 1%
Triton X-100, 2 mM EDTA, 16.7 mM Tris-HCl, pH 8.1, 167 mM NaCl and
protease inhibitors). Primary antibodies were added and incubated
overnight at 48°C with rotation. The immunocomplex was collected by
protein A/G agarose beads and washed with low salt washing buffer
(0.1% SDS, 1% Triton X-100, 2 mM EDTA, 200 mM Tris-HCl, pH 8.1 and
150 mM NaCl), high-salt buffer (0.1% SDS, 1% Triton X-100, 2 mM
EDTA, 200 mM Tris-HCl, pH 8.1 and 500 mM NaCl), LiCl washing buffer
(0.25 M LiCl, 1% NP40, 1% deoxycolate, 1 mM EDTA and 10 mM
Tris-HCl, pH 8.1), and finally 1 TE buffer (10 mM Tris-HCl and 1 mM
EDTA, pH 8.0). Then, the immunocomplex was eluted by the elution
buffer (1% SDS, 0.1 M NaHCO3 and 200 mM NaCl) and the
crosslinks were reversed by heating at 65°C for 4 h. After
reaction, the samples were adjusted to 10 mM EDTA, 20 mM Tris-HCl,
pH 6.5 and 40 mg/ml proteinase K, and incubated at 45°C for 1 h.
DNA was recovered and was subjected to PCR amplification of the
LCN2 promoter region, the primers were: 5′-gacagctcttccggctcacag-3′
(forward) and 5′-cgctgtggt ggctgctgggcc-3′ (reverse).
Statistical analysis
Values are expressed as means ± SD. The Student's
t-test was employed for the analyses. A P-value of <0.05 was
considered to indicate a statistically significant result.
Results
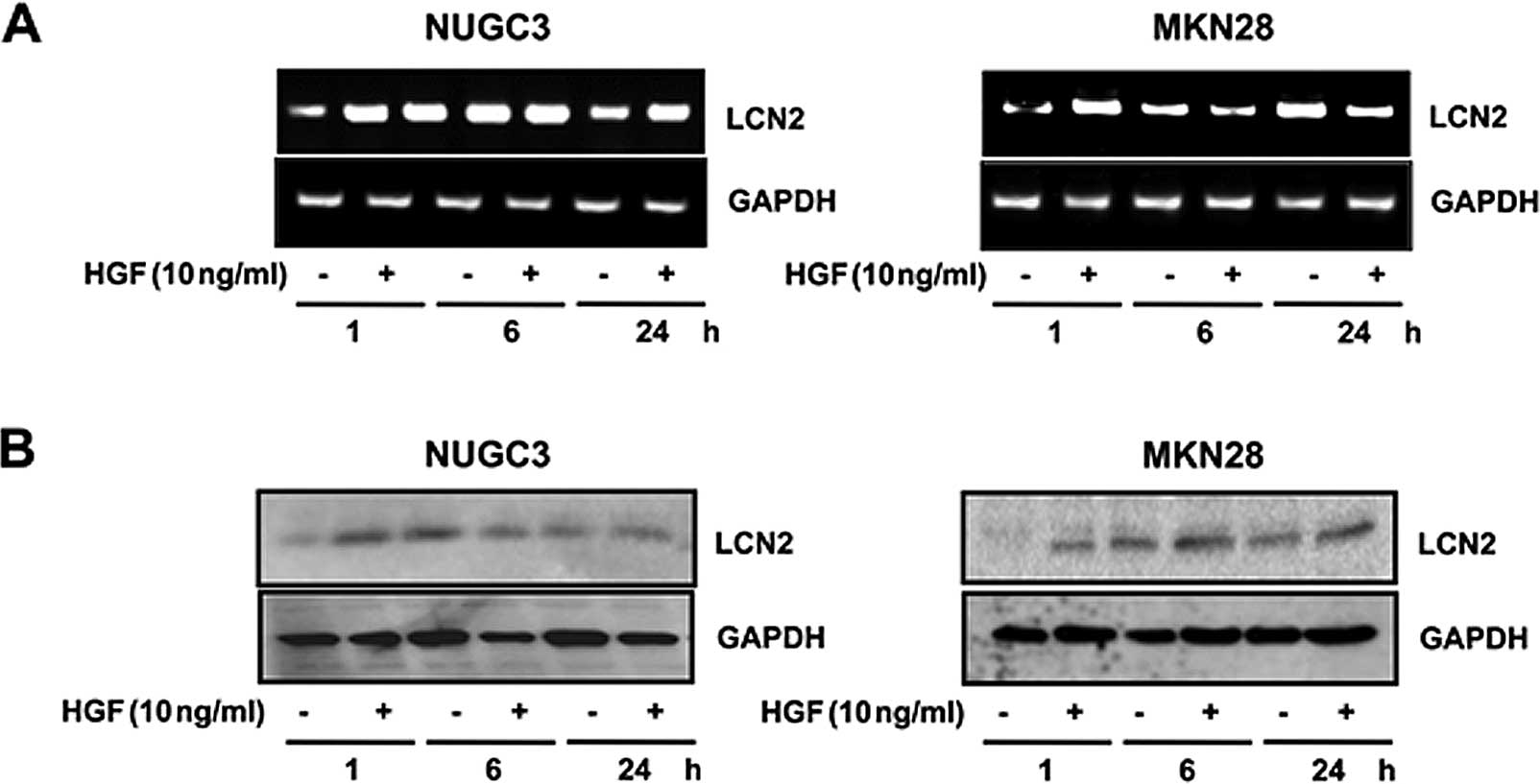
Upregulation of LCN2 level after
treatment with HGF
To investigate the upregulation of the LCN2 gene,
western blotting and RT-PCR analysis were performed. RT-PCR showed
that the expression level of LCN2 mRNA was increased by treatment
with HGF (Fig. 1A). The LCN2
protein level was increased after treatment with HGF, confirmed by
western blot analysis (Fig.
1B).
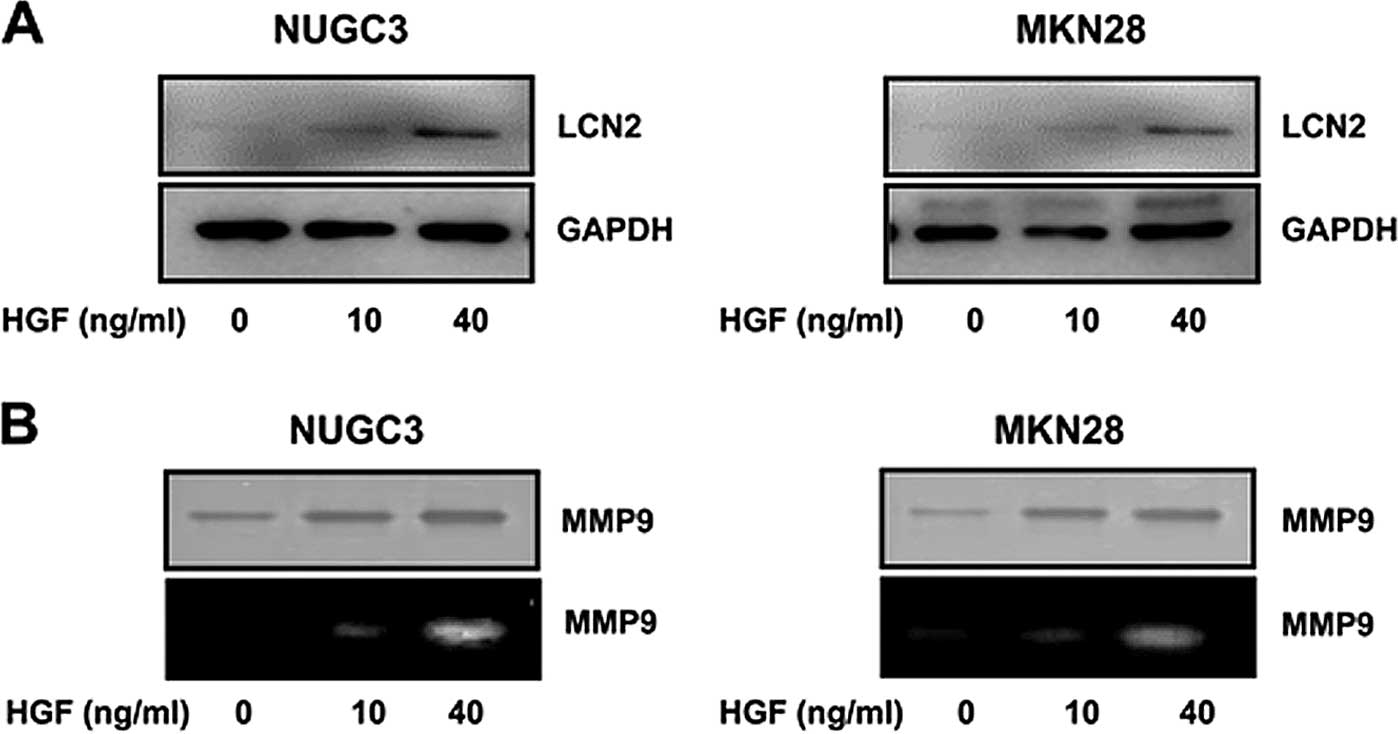
Upregulation of LCN2 and MMP9 level after
treatment with HGF
We also tested the LCN2 protein level in HGF
treatment in a dose-dependent manner confirmed by western blot
analysis. The expression level of LCN2 protein was increased with
increasing concentration of HGF (0, 10 and 40 ng/ml) (Fig. 2A).
It is well known that MMP9 is associated with cancer
invasion induced by HGF. To validate the MMP9 protein level in HGF
treatment, western blotting and zymogram analysis were performed.
The expression level of MMP9 protein was increased dose-dependently
by HGF (0, 10 and 40 ng/ml) (Fig.
2B).
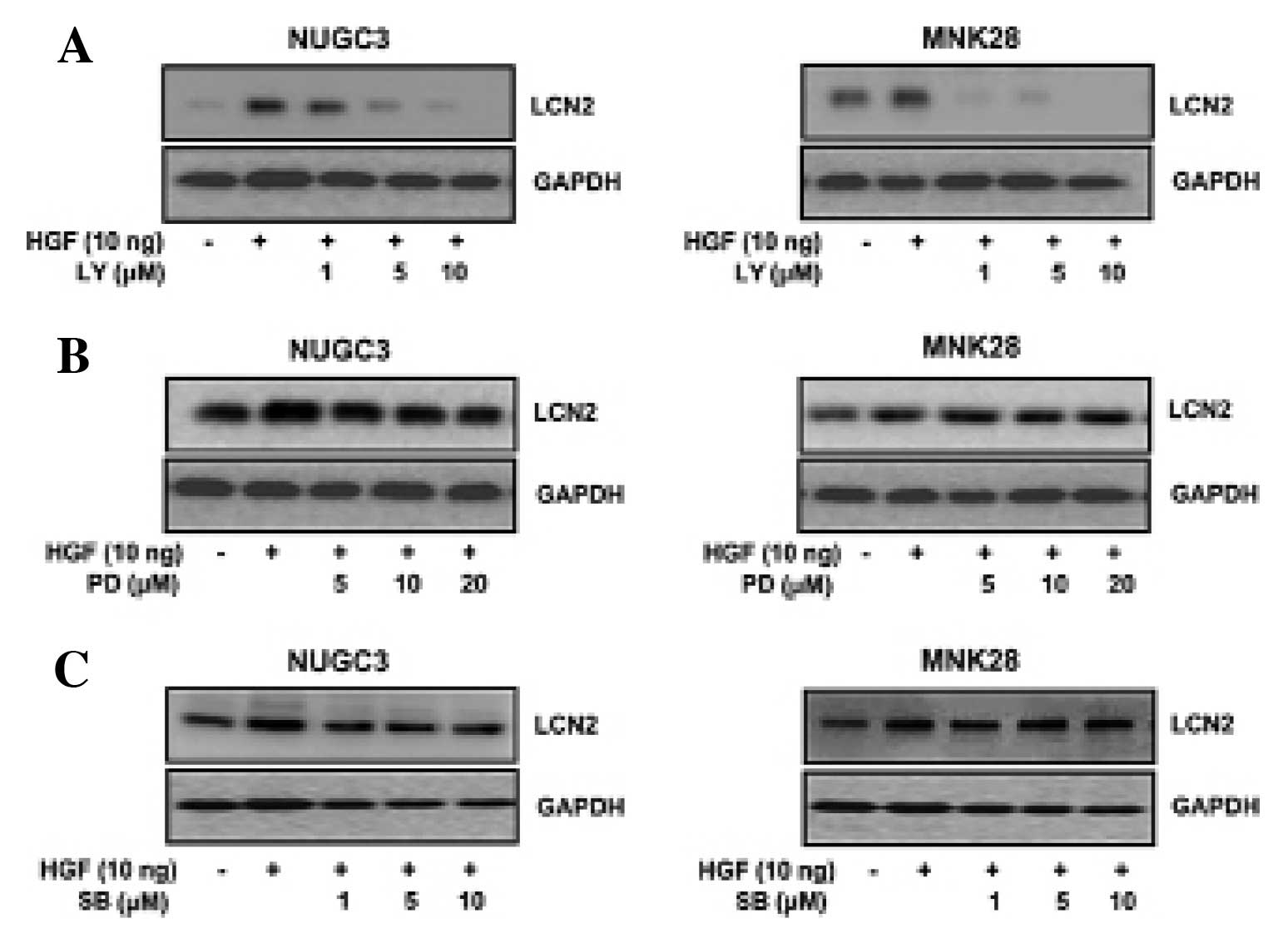
Effects of LY294002, PD098059 and
SB203580 on LCN2 expression
To identify whether or not PI3-kinase, ERK and MAPK
activation was associated with HGF-induced LCN2, the cells were
treated with PI3-kinase inhibitor (LY294002), or MEK inhibitor
(PD098059), or p38 inhibitor (SB203580) and then analyzed by
western blotting. The HGF-mediated LCN2 protein level was decreased
with LY294002 (Fig. 3A). Yet,
treatment with PD098059 (Fig. 3B)
and SB203580 (Fig. 3C) showed no
change in LCN2 expression in either NUGC3 and MNK28 cell lines
(Fig. 3). These results suggested
that HGF-mediated LCN2 is regulated by PI3-kinase, not by ERK or
p38.
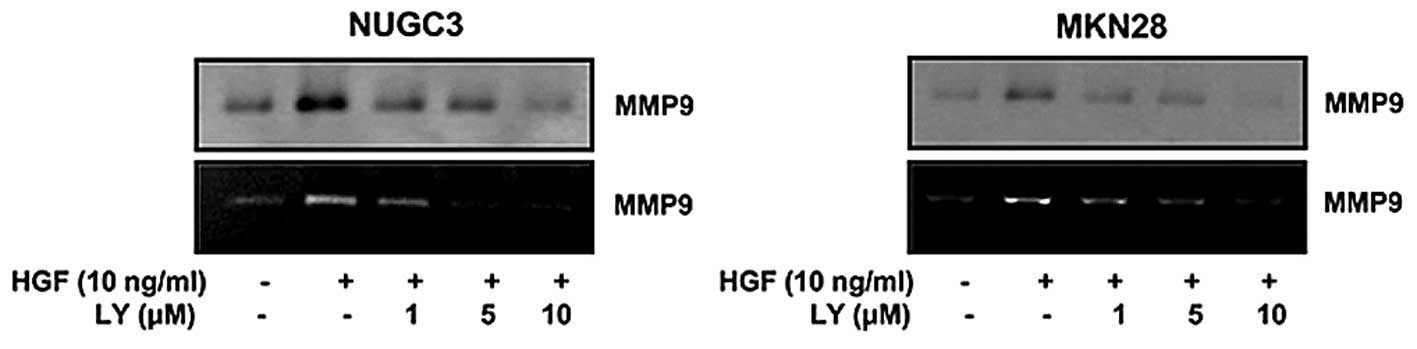
Effect of LY294002 on MMP9
expression
Some studies suggested that LCN2 was observed to
stabilize MMP9 associated with tumor growth and metastasis. We
tested whether MMP9 is also regulated by PI3-kinase-like LCN2. The
cells were treated with PI3-kinase inhibitor (LY294002) and
measured by western blotting and zymography. The results showed
that HGF-mediated MMP9 was decreased with LY294002 (Fig. 4). We thusidentified that MMP9 is
regulated by a PI3-kinase.
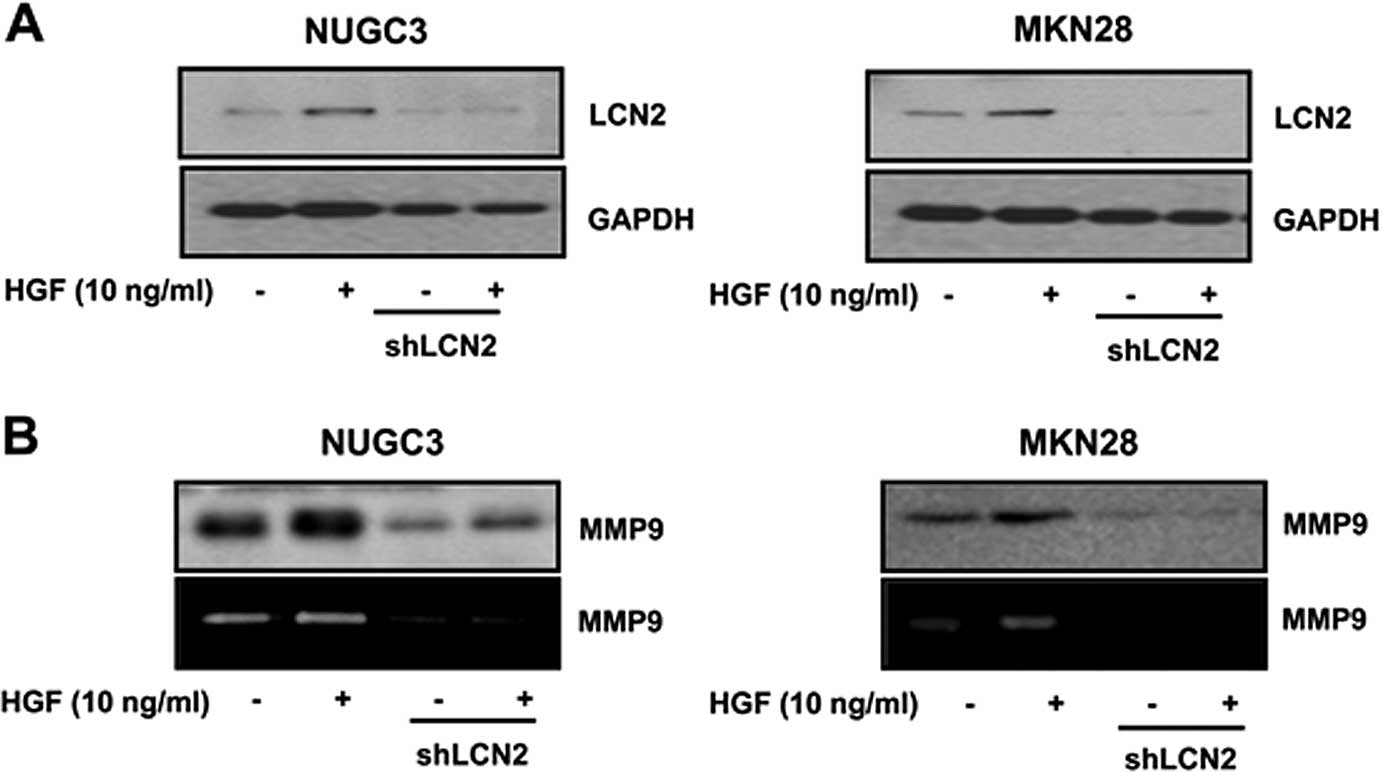
Effect of the LCN2 knockdown on
HGF-mediated MMP9
To confirm that MMP9 is regulated by LCN2, the
effect of LCN2 knockdown on HGF-mediated MMP9 was measured. We
showed that LCN2 level was decreased in both LCN2 knockdown cell
lines to identify the appropriate function of knockdown cells
(Fig. 5A). We measured the effect
of LCN2 knockdown of HGF-mediated MMP9 by western blotting.
LCN2-shRNA cells showed a decreased level of HGF-mediated MMP9
(Fig. 5B).
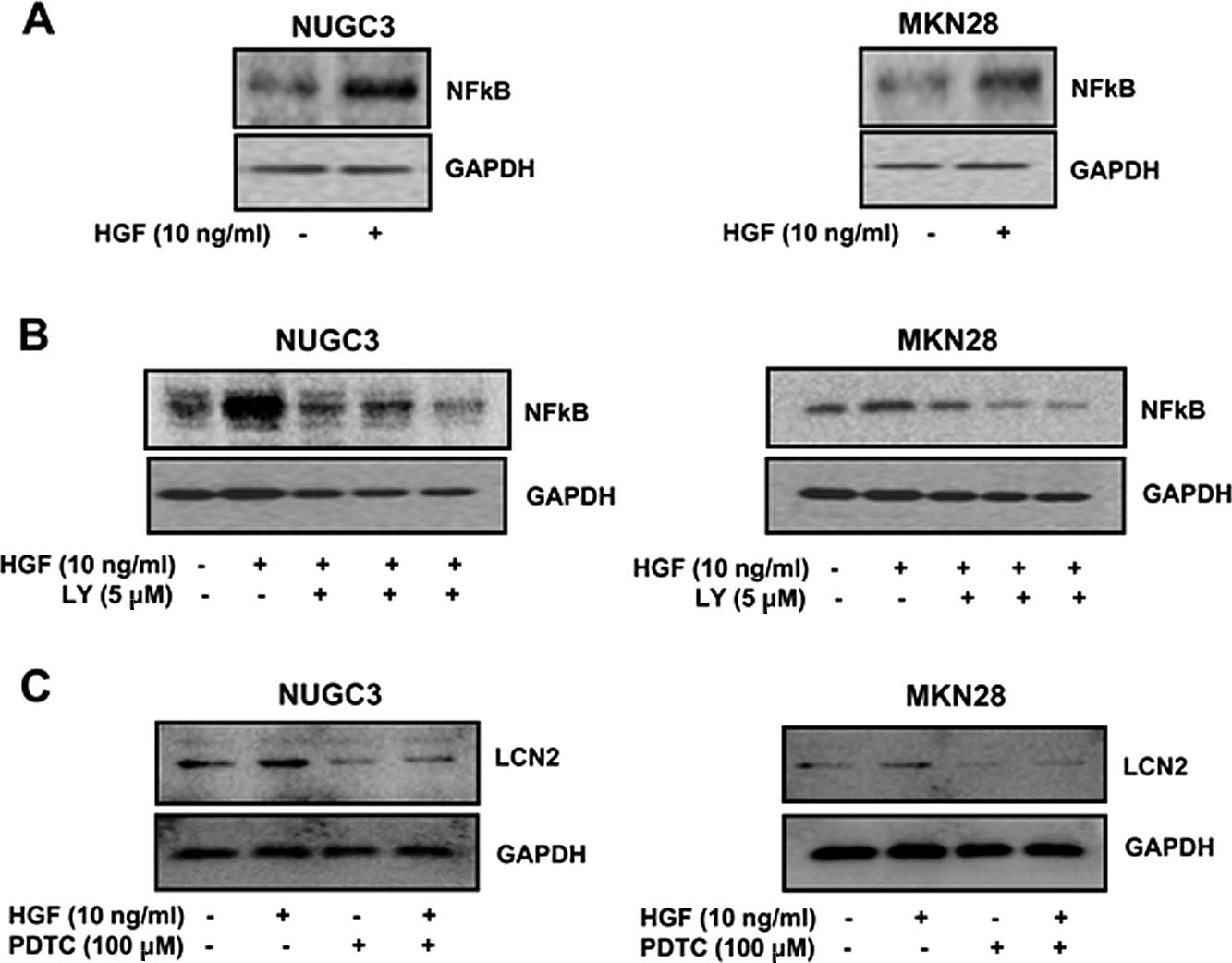
Effect of NFκB on LCN2 expression
First we tested whether NFκB is mediated by HGF and
regulated by a PI3-kinase. NFκB level was increased by HGF
(Fig. 6A). Also, HGF-mediated NFκB
protein level was decreased with the PI3-kinase inhibitor LY294002
(Fig. 6B). To explore the
regulation of NFκB on LCN2, the cells were treated with NFκB
inhibitor (PDTC) and then the level of LCN2 was evaluated by
western blotting. The HGF-mediated LCN2 protein level was decreased
with treatment of PDTC in both NUGC3 and MNK28 cell lines (Fig. 6C). These results suggested that
HGF-mediated LCN2 is regulated by NFκB.
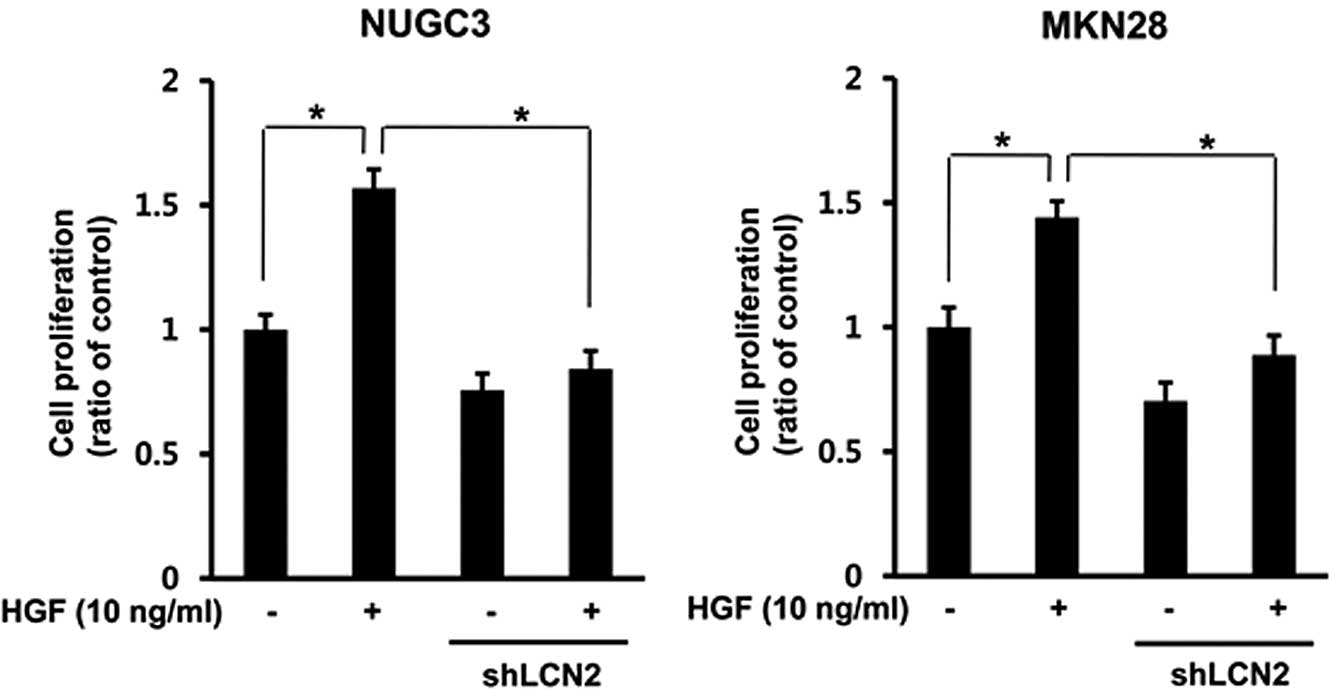
Effect of LCN2 knockdown on HGF-mediated
cell proliferation
To explore the effects of the LCN2 knockdown on
HGF-mediated cell proliferation in gastric cancer cells, stable
LCN2-shRNA cells were prepared by transfection of shRNA into NUGC-3
and MKN-28 cells. Knockdown of the LCN2-shRNA stable cell was
confirmed by RT-PCR. The control and LCN2-sh RNA cells were treated
with HGF. After 72 h, we measured the cell proliferation by MTT
assay. The data show that HGF-mediated cell proliferation was
decreased in LCN2-sh RNA cells compared to control cells in both
gastric cell lines (P<0.05) (Fig.
7).
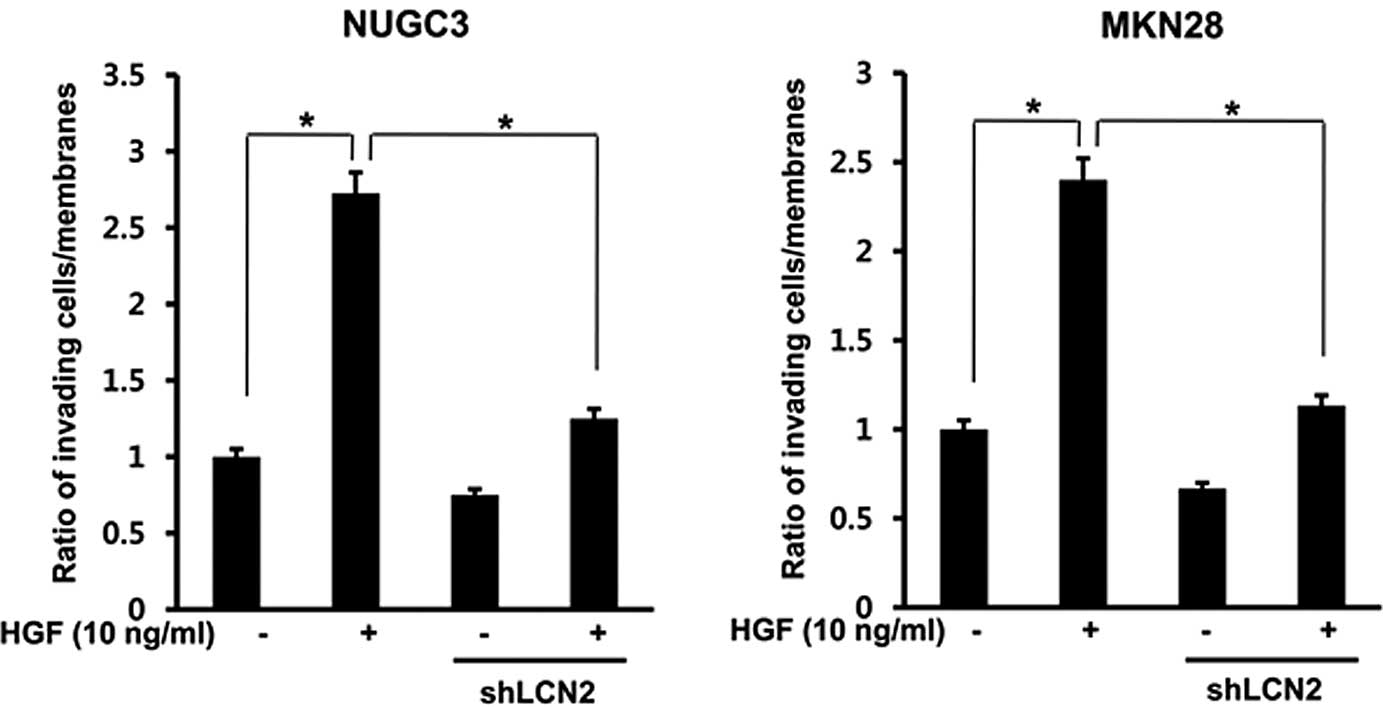
Effect of the LCN2 knockdown on
HGF-mediated cell invasion
To determine whether LCN2 has a role in cell
invasion, an in vitro invasion assay was performed using
Matrigel migration chamber. The generated LCN2-shRNA was treated
with HGF in NUGC3 and MKN28 cell lines. After 72 h, HGF-mediated
cell invasion was decreased in LCN2-shRNA in both cell lines
(P<0.05) (Fig. 8).
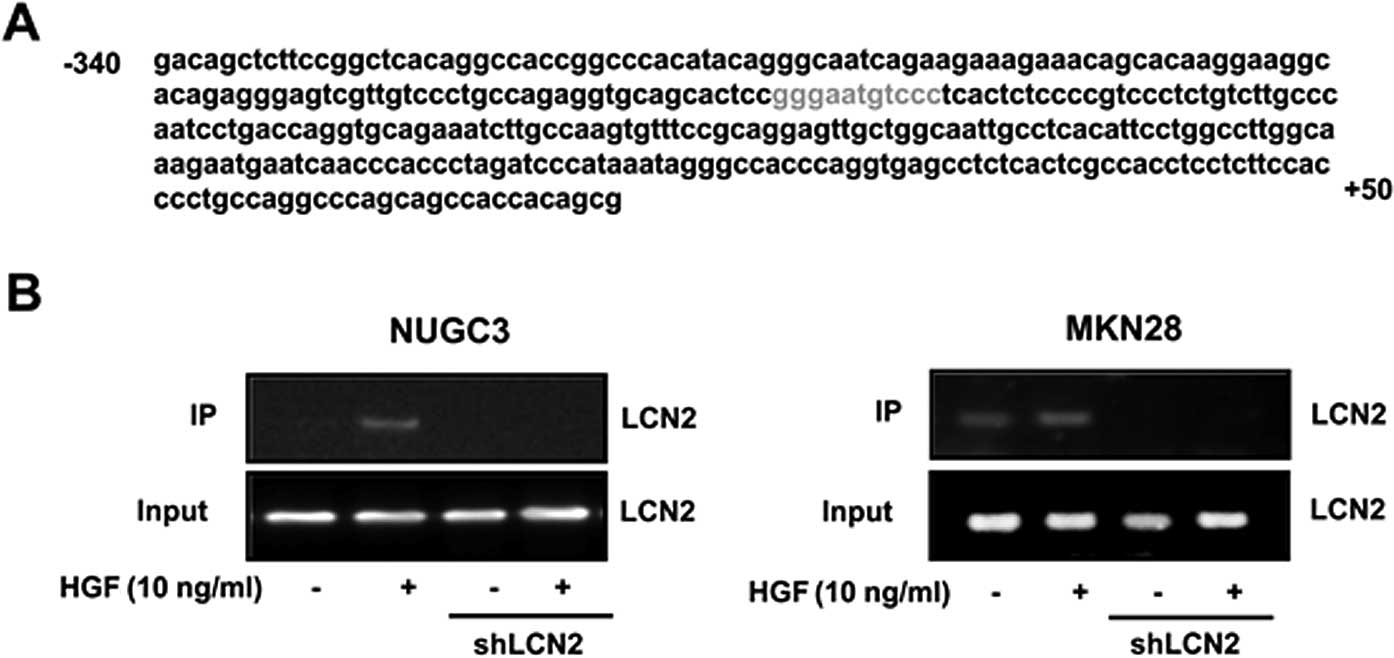
Binding NFκB to the LCN2 promoter
To confirm that NFκB regulate transcriptional
activity of LCN2 mRNA by binding to the LCN promoter, we
investigated the promoter sequence of LCN2 genes to establish the
putative NFκB binding sequence using computer program for sequence
analysis. One putative binding site was found within LCN2 promoter.
NFκB binding site of LCN2 promoter sequence was located in the
proximal promoter region of the transcriptional start site
(Fig. 9A). To examine the function
of NFκB binding site in the LCN2 promoter, LCN2 shRNA and control
cells were treated with HGF and binding activity of NFκB to
putative NFκB binding sites was measured by the ChIP assay. HGF
enhanced NFκB binding activity to the LCN2 promoter with strong
activity in control cells, yet not in LCN2-shRNA cells (Fig. 9B).
Discussion
Gastric cancer is the most common malignant tumor
and the second most common cause of cancer death worldwide
(15,16). The small reduction of gastric cancer
mortality showed the need for further identification of targets or
pathways that facilitate the proliferation and invasion of
cancer.
MMP9 has been associated with cancer invasion by
degradation of matrix in various tumor types. Recent studies
reported that the high molecule LCN2/MMP9 complex which protect the
MMP9 proteolytic degradation, was found in several types of cancer
including breast, colon and gastric cancer (12,13,17).
This complex could perform remodeling of the ECM by degradation of
an anchor protein of cell-to-cell adhesion (4,18).
Yet, there is little information concerning exact correlation
between LCN2 and MMP9 and function of LCN2 in tumorigenesis. Li
et al suggested that LCN2 expression is upregulated by ErbB2
in the NFκB dependent pathway in breast cancer cells. They studied
the association between ErbB2 and LCN2, not the association between
LCN2 and MMP9 (19).
LCN2 is expressed in various epithelial and
endothelial cells, fibroblasts and hepatocytes (20,21).
The level of LCN2 is enhanced in various types of cancer including
stomach cancer (13),
cholangiocarcinoma (10), breast
(22), colon (17) and pancreatic cancer (23). It has been suggested that stimuli in
the tumor microenvironment including hypoxia and inflammation
induce LCN2 (24–27). Furthermore, LCN2 is associated with
tumorigenesis and metastasis by enhancing gelatinase activity with
LCN2 expression, which facilitates tumor growth by enhancing cell
invasion of surrounding tissues in breast cancer (28).
It was shown that the complex of LCN2/MMP9 was
enhanced in human gastric tumors and suggested that enhancing LCN2
level stimulates the formation of the complex with MMP9, which
results in the maintenance of powerful proteinase action by
protection of MMP9 autodegradation, and that high level of
LCN2/MMP9 appears obviously correlated with overall survival of
gastric cancer patients (13). On
the contrary, the in vitro experiments suggested a
protective role against cancer by showing that LCN2 induces the
expression of E-cadherin, to diminish the invasiveness and
metastasis in cancer cells (29).
Another study reported that overexpression of LCN2 decreased cell
invasion of colon and pancreatic cancer cells, suggesting the role
of LCN2 as a suppressor of cancer metastasis (30).
We investigated whether HGF is associated with LCN2
and MMP9 in gastric cancer and identified the role of LCN2
upregulated by HGF associated with MMP9 in gastric cancer invasion
and metastasis at molecular level. To identify whether LCN2 is
associated with regulation of cell proliferation and invasion of
metastatic characteristics, we knocked down the expression of LCN2
in NUGC3 and NKN28 cells using shRNA and evaluate in vitro
cell proliferation, invasiveness and MMP9 activity. The present
study showed that HGF upregulates LCN2 and MMP9, and MMP9 activity
is upregulated by LCN2. Also, we identified that LCN2 is associated
with cell proliferation and invasion by MTT and in vitro
invasion assays in gastric cancer cell lines. Our data support the
promotion of cancer metastasis by showing the suppression of in
vitro invasiveness and proliferation of gastric cancer cell
lines. These results are similar to the report of Nuntagowat et
al, where LCN2 knockdown in cholangiocarcinoma is association
with invasion and MMP activity. Yet, they reported the lack of
effect in cell proliferation by LCN2 knockdown (10).
Furthermore, we found out the regulator pathway of
LCN2 by identifying putative NFκB binding site in LCN2 promoter and
confirmed that LCN2 in gastric cancer cells is upregulated directly
by rhe NFκB pathway. MMP9 was inhibited with the PI3-kinase
inhibitor LY294002 treatment, HGF-mediated MMP9 is also regulated
by PI3-kinase. Moreover, the present study suggested that both LCN2
and MMP9 are regulated by the NFκB pathway. Pathway experiments on
the regulation of LCN2 and MMP9 are rare. Only one study reported
that LCN2 is upregulated by ErbB2 through NFκB activation (19). The present study provide evidence
concerning the regulator pathway in invasion and proliferation of
gastric cancer cell lines by LCN2 and MMP9.
Based on these results, we confirmed the definite
relationship between LCN2 and MM9 and the function of cell
proliferation and invasion of LCN2 in gastric cancer cell lines.
Therefore, LCN2 appear to have a possible role in tumorigenesis and
tumor invasion associated with MMP9 through NFκB pathway in gastric
cancer. Our findings support a previous study which suggested that
LCN2 level stimulates the formation of the complex with MMP9 with
human gastric tumor tissue (13).
Nuntagowat et al showed that silencing of
LCN2 expression induced a significant suppression of in
vitro invasion which paralleled a reduction of the LCN2/MMP9
complex (10). Zhang et al
reported that the activity of LCN2/MMP9 complex correlated with the
depth of esophageal cancer using gelatin zymography (31). The present study does not provide
evidence whether LCN2 directly regulates MMP9 and which mechanism
acts in making the complex LCN2/MMP9 and we do not have evidence
whether the complex LCN2/MMP9 also acts in cancer invasion and
proliferation in gastric cancer. We only confirmed that LCN2
regulates MMP9, that expression of LCN2 increased in vitro
invasiveness and proliferation of gastric cancer cell lines and
that LCN2 and MMP9 is separately regulated by NFκB, which means
that NFκB pathway does not have direct connection between LCN2 and
MMP9.
To clarify our findings, further experiments are
warranted using in vivo a knockout mouse model with both
LCN2 and MMP9 and it is necessary to study the relation of the
levels LCN2/MMP9 complex in samples of gastric cancer patients and
clinical parameters and survival of the patients. It would be
useful to examine the role of the LCN2/MMP9 complex.
In conclusion, the present study demonstrated that
the MMP9 activity was upregulated by LCN2, and both LCN2 and MMP9
are controlled by the NFκB pathway which results in progression and
invasion of two gastric cancer cell lines. This pathway may serve
as possible therapeutic target option and provide information for
further identification of other targets in gastric cancer.
Acknowledgments
This study was supported by the 2014 Yeungnam
University Research Grant.
References
|
1
|
Yang J and Moses MA: Lipocalin 2: A
multifaceted modulator of human cancer. Cell Cycle. 8:2347–2352.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu S and Venge P: Lipocalins as
biochemical markers of disease. Biochim Biophys Acta. 1482:298–307.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alpízar-Alpízar W, Laerum OD, Illemann M,
Ramírez JA, Arias A, Malespín-Bendaña W, Ramírez V, Lund LR,
Borregaard N and Nielsen BS: Neutrophil gelatinase-associated
lipocalin (NGAL/Lcn2) is upregulated in gastric mucosa infected
with Helicobacter pylori. Virchows Arch. 455:225–233. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan L, Borregaard N, Kjeldsen L and Moses
MA: The high molecular weight urinary matrix metalloproteinase
(MMP) activity is a complex of gelatinase B/MMP-9 and neutrophil
gelatinase-associated lipocalin (NGAL). Modulation of MMP-9
activity by NGAL. J Biol Chem. 276:37258–37265. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tschesche H, Zölzer V, Triebel S and
Bartsch S: The human neutrophil lipocalin supports the allosteric
activation of matrix metalloproteinases. Eur J Biochem.
268:1918–1928. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sier CF, Kubben FJ, Ganesh S, Heerding MM,
Griffioen G, Hanemaaijer R, van Krieken JH, Lamers CB and Verspaget
HW: Tissue levels of matrix metalloproteinases MMP-2 and MMP-9 are
related to the overall survival of patients with gastric carcinoma.
Br J Cancer. 74:413–417. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kubben FJ, Sier CF, van Duijn W, Griffioen
G, Hanemaaijer R, van de Velde CJ, van Krieken JH, Lamers CB and
Verspaget HW: Matrix metalloproteinase-2 is a consistent prognostic
factor in gastric cancer. Br J Cancer. 94:1035–1040. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsumura S, Oue N, Nakayama H, Kitadai Y,
Yoshida K, Yamaguchi Y, Imai K, Nakachi K, Matsusaki K, Chayama K,
et al: A single nucleotide polymorphism in the MMP-9 promoter
affects tumor progression and invasive phenotype of gastric cancer.
J Cancer Res Clin Oncol. 131:19–25. 2005. View Article : Google Scholar
|
|
9
|
Kjeldsen L, Johnsen AH, Sengeløv H and
Borregaard N: Isolation and primary structure of NGAL, a novel
protein associated with human neutrophil gelatinase. J Biol Chem.
268:10425–10432. 1993.PubMed/NCBI
|
|
10
|
Nuntagowat C, Leelawat K and Tohtong R:
NGAL knockdown by siRNA in human cholangiocarcinoma cells
suppressed invasion by reducing NGAL/MMP-9 complex formation. Clin
Exp Metastasis. 27:295–305. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fernández CA, Yan L, Louis G, Yang J,
Kutok JL and Moses MA: The matrix metalloproteinase-9/neutrophil
gelatinase-associated lipocalin complex plays a role in breast
tumor growth and is present in the urine of breast cancer patients.
Clin Cancer Res. 11:5390–5395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leng X, Ding T, Lin H, Wang Y, Hu L, Hu J,
Feig B, Zhang W, Pusztai L, Symmans WF, et al: Inhibition of
lipocalin 2 impairs breast tumorigenesis and metastasis. Cancer
Res. 69:8579–8584. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kubben FJ, Sier CF, Hawinkels LJ,
Tschesche H, van Duijn W, Zuidwijk K, van der Reijden JJ,
Hanemaaijer R, Griffioen G, Lamers CB, et al: Clinical evidence for
a protective role of lipocalin-2 against MMP-9 autodegradation and
the impact for gastric cancer. Eur J Cancer. 43:1869–1876. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang SH, Kim JS, Oh TJ, Kim MS, Lee SW,
Woo SK, Cho HS, Choi YH, Kim YH, Rha SY, et al: Genome-scale
analysis of resveratrol-induced gene expression profile in human
ovarian cancer cells using a cDNA microarray. Int J Oncol.
22:741–750. 2003.PubMed/NCBI
|
|
15
|
Parkin DM, Bray FI and Devesa SS: Cancer
burden in the year 2000. The global picture. Eur J Cancer. 37(Suppl
8): S4–S66. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nielsen BS, Borregaard N, Bundgaard JR,
Timshel S, Sehested M and Kjeldsen L: Induction of NGAL synthesis
in epithelial cells of human colorectal neoplasia and inflammatory
bowel diseases. Gut. 38:414–420. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rudd PM, Mattu TS, Masure S, Bratt T, Van
den Steen PE, Wormald MR, Küster B, Harvey DJ, Borregaard N, Van
Damme J, et al: Glycosylation of natural human neutrophil
gelatinase B and neutrophil gelatinase B-associated lipocalin.
Biochemistry. 38:13937–13950. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li SH, Hawthorne VS, Neal CL, Sanghera S,
Xu J, Yang J, Guo H, Steeg PS and Yu D: Upregulation of neutrophil
gelatinase-associated lipocalin by ErbB2 through nuclear
factor-kappaB activation. Cancer Res. 69:9163–9168. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Q and Nilsen-Hamilton M:
Identification of a new acute phase protein. J Biol Chem.
270:22565–22570. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Q, Nilsen-Hamilton M and Xiong S:
Synergistic regulation of the acute phase protein SIP24/24p3 by
glucocorticoid and pro-inflammatory cytokines. Sheng Li Xue Bao.
55:525–529. 2003.PubMed/NCBI
|
|
22
|
Bauer M, Eickhoff JC, Gould MN, Mundhenke
C, Maass N and Friedl A: Neutrophil gelatinase-associated lipocalin
(NGAL) is a predictor of poor prognosis in human primary breast
cancer. Breast Cancer Res Treat. 108:389–397. 2008. View Article : Google Scholar
|
|
23
|
Tong Z, Kunnumakkara AB, Wang H, Matsuo Y,
Diagaradjane P, Harikumar KB, Ramachandran V, Sung B, Chakraborty
A, Bresalier RS, et al: Neutrophil gelatinase-associated lipocalin:
A novel suppressor of invasion and angiogenesis in pancreatic
cancer. Cancer Res. 68:6100–6108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Viau A, El Karoui K, Laouari D, Burtin M,
Nguyen C, Mori K, Pillebout E, Berger T, Mak TW, Knebelmann B, et
al: Lipocalin 2 is essential for chronic kidney disease progression
in mice and humans. J Clin Invest. 120:4065–4076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang W, Constante M and Santos MM: Anemia
upregulates lipocalin 2 in the liver and serum. Blood Cells Mol
Dis. 41:169–174. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Greten FR, Eckmann L, Greten TF, Park JM,
Li ZW, Egan LJ, Kagnoff MF and Karin M: IKKbeta links inflammation
and tumorigenesis in a mouse model of colitis-associated cancer.
Cell. 118:285–296. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li C and Chan YR: Lipocalin 2 regulation
and its complex role in inflammation and cancer. Cytokine.
56:435–441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Leng X, Wu Y and Arlinghaus RB:
Relationships of lipocalin 2 with breast tumorigenesis and
metastasis. J Cell Physiol. 226:309–314. 2011. View Article : Google Scholar
|
|
29
|
Hanai J, Mammoto T, Seth P, Mori K,
Karumanchi SA, Barasch J and Sukhatme VP: Lipocalin 2 diminishes
invasiveness and metastasis of Ras-transformed cells. J Biol Chem.
280:13641–13647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee HJ, Lee EK, Lee KJ, Hong SW, Yoon Y
and Kim JS: Ectopic expression of neutrophil gelatinase-associated
lipocalin suppresses the invasion and liver metastasis of colon
cancer cells. Int J Cancer. 118:2490–2497. 2006. View Article : Google Scholar
|
|
31
|
Zhang H, Xu L, Xiao D, Xie J, Zeng H, Wang
Z, Zhang X, Niu Y, Shen Z, Shen J, et al: Upregulation of
neutrophil gelatinase-associated lipocalin in oesophageal squamous
cell carcinoma: Significant correlation with cell differentiation
and tumour invasion. J Clin Pathol. 60:555–561. 2007. View Article : Google Scholar : PubMed/NCBI
|