Introduction
MicroRNAs (miRNAs) are a group of endogenous small
non-coding RNA molecules, which regulate protein-coding gene
expression by interacting with complementary sites within the
3′-untranslated region (UTR) of target mRNAs and targeting mRNAs
for cleavage or translational repression (1). Mounting evidence has indicated that
miRNAs are involved in tissue morphogenesis, cell processes such as
proliferation, cell cycle and apoptosis and major signaling
pathways (2–4). Previous studies have identified the
critical role of miRNAs in human cancers and suggest that
deregulation of miRNAs is involved in tumor development and
progression by modulating the expression of oncogenes or tumor
suppressors (5).
miR-92a, which belongs to the miR-17-92 cluster,
plays a critical role in the progression of lung cancer (6), esophageal squamous cell carcinoma
(7), colorectal (8), breast (9), ovarian (10) and cervical cancer (11). Previous findings show that miR-92a
functions as a regulator of cell proliferation, apoptosis and
invasion in human cancers (11–13).
miR-92a promotes lung cancer cell invasion by targeting
reversion-inducing-cysteine-rich protein with Kazal motifs (RECK)
and a high expression of miR-92a is associated with the poor
survival rate of lung cancer patients (6,13).
Plasma miR-92a is a promising novel biomarker for the early
detection of colorectal cancer and its overexpression is also a
prognostic marker for predicting the poor overall and disease-free
survival rate of patients (8,14).
However, the expression of miR-92a is downregulated in ovarian and
breast cancer (9,10). Upregulation of miR-92a inhibits
ovarian cancer cell adhesion, invasion and proliferation by
suppressing integrin α5 expression (10). Downregulation of miR-92a is
associated with aggressive breast cancer features and increased
tumor macrophage infiltration (9).
Thus, the functional significance of miR-92a in cancer initiation
and development seems to be cancer-type specific. Previously,
miR-92a was found to be significantly upregulated in hepatocellular
carcinoma (HCC) samples and was also identified as hepatitis B
virus (HBV)-specific (15,16). The proliferation of HCC-derived cell
lines was enhanced by the upregulation of miR-92a and inhibited by
the downregulation of miR-92a (16). However, the clinical significance of
miR-92a and the underlying mechanisms involved in the development
of HCC remain to be investigated.
The present study aim was to determine the role of
miR-92a in HCC. The results showed that the expression of miR-92a
was upregulated in HCC tissues. miR-92a promoted proliferation,
cell cycle and apoptosis resistance in vitro. Moreover, the
downregulation of miR-92a inhibited the tumor growth of HCC in
vivo. Notably, F-box and WD repeat domain-containing 7 (FBXW7)
was identified as a direct target of miR-92a. A high expression of
miR-92a was associated with poor clinicopathological
characteristics and the reduced survival of HCC patients. The
results showed a new role for miR-92a in prediction of prognosis
and promoting tumor growth of human HCC.
Materials and methods
Clinical samples
HCC and matched normal tumor-adjacent tissues were
obtained from 106 patients including 94 males and 12 females, who
underwent resection of their primary HCC in the Department of
Hepatobiliary Surgery at the First Affiliated Hospital of Xi'an
Jiaotong University (Shaanxi, China) from January, 2006 to
December, 2009, with a median follow-up period of 38,7 months. None
of the patients received preoperative chemo- or radiotherapy. The
stage of cancer was determined according to the cancer staging
system published in 2010 by the Union for International Cancer
Control (UICC). The demographic features and clinicopathological
data are shown in Table I. Samples
were used after informed consent was obtained. The Xi'an Jiaotong
University Ethics Committee approved all the protocols according to
the Declaration of Helsinki (as revised in Tokyo 2004).
 | Table IClinicopathological correlation of
miR-92a expression in HCC. |
Table I
Clinicopathological correlation of
miR-92a expression in HCC.
| Clinicopathological
characteristics | miR-92a
| P-value |
|---|
| High expression
(n=53) | Low expression
(n=53) |
|---|
| Age (years) |
| ≤50 | 20 | 22 | 0.691 |
| >50 | 33 | 31 | |
| Gender |
| Male | 46 | 48 | 0.540 |
| Female | 7 | 5 | |
| HBsAg |
| No | 3 | 11 | 0.022a |
| Yes | 50 | 42 | |
| Serum AFP level
(ng/ml) |
| ≤20 | 14 | 18 | 0.397 |
| >20 | 39 | 35 | |
| Tumor size
(cm) |
| ≤5 | 18 | 30 | 0.019a |
| >5 | 35 | 23 | |
| No. of tumor
nodules |
| 1 | 43 | 45 | 0.605 |
| ≥2 | 10 | 8 | |
| Cirrhosis |
| Absent | 7 | 17 | 0.020a |
| Present | 46 | 36 | |
| Venous
infiltration |
| Absent | 24 | 27 | 0.560 |
| Present | 29 | 26 | |
| Edmondson-Steiner
grading |
| I+II | 33 | 47 | 0.002a |
| III+IV | 20 | 6 | |
| TNM tumor
stage |
| I+II | 34 | 45 | 0.014a |
| III+IV | 19 | 8 | |
Reverse transcription-quantitative PCR
(RT-qPCR)
qPCR primer against mature miRNA hsa-miR-92a-3p
(HmiRQP0832) and Homo sapiens snRNA U6 qPCR Primer (HmiRQP9001)
were purchased from Genecopoeia (Guangzhou, China). The PCR
amplification for the quantification of the miR-92a and U6 was
performed using the TaqMan miRNA Reverse Transcription kit (Applied
Biosystems, Foster City, CA, USA) and TaqMan Human miRNA Assay kit
(Applied Biosystems). The relative expression of miR-92a was shown
as the fold difference relative to U6.
The PCR amplification for the quantification of the
FBXW7 and GAPDH mRNAs was performed using an ABI PRISM 7300
Sequence Detection system (Applied Biosystems) and a
SYBR® Premix Ex Taq™ II (Perfect Real Time) kit (Takara
Bio, Shiga, Japan), as previously reported (17).
Cell lines and transfection
The human immortalized normal hepatic cell line LO2
and five HCC cell lines (HepG2, Hep3B, Huh7, SMMC-7721 and
Bel-7402) were purchased from the Institute of Biochemistry and
Cell Biology, Chinese Academy of Sciences, Shanghai, China. The
cells were maintained in Dulbecco's modified Eagle's medium (DMEM;
Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine
serum (Gibco) with 100 U/ml penicillin and 100 µg/ml
streptomycin (Sigma, St. Louis, MO, USA) at 37°C with 5%
CO2.
miRNA vectors, including miR-92a expression vector
(HmiR0204-MR04), the control vector for miR-92a (CmiR0001-MR04 and
miR-control), miR-92a inhibitor (HmiR-AN0832-AM04 and anti-miR-92a)
and the negative control for the miR-92a inhibitor
(CmiR-AN0001-AM04 and anti-miR-NC), were purchased from
Genecopoeia. Retroviral vectors pMMP-FBXW7 were generated by
inserting the FBXW7 cDNA into pMMP. Retrovirus packaging and
transduction were performed as previously described (18). The targeted sequences for FBXW7
siRNA sense, 5′-GGA GUA UGG UCA UCA CAA Att-3′ and antisense,
5′-UUU GUG AUG ACC AUA CUC Cac-3′ or a non-specific duplex
oligonucleotide as a negative control were produced by Sangon
Biotech Co., Ltd. (Shanghai, China). The cells were transfected
with the vectors mentioned above using Lipofectamine 2000 according
to the manufacturer's instructions (Invitrogen, Carlsbad, CA,
USA).
Cell cycle, proliferation and detection
of apoptosis
Flow cytometry was performed using the
fluorescence-activated cell sorting (FACS) Calibur and Cell Quest
software (both from Becton-Dickinson, San Jose, CA, USA). For the
proliferation assay, HCC cells transfected with different vectors
were seeded in 96-well plates at a density of 5×103
cells/well for 24 h and assessed using Cell Proliferation ELISA,
BrdU kit (5-bromodeoxyuridine) (chemiluminescent) (Roche,
Indianapolis, IN, USA). For cell cycle analysis, the cells were
seeded in 6-well plates at 2×105/well. Forty-eight hours
after transfection, the cells were fixed in 70% ethanol at 4°C for
24 h and stained with 50 µg/ml propidium iodide (Keygen,
Nanjing, China). An Annexin-V-Fluos Staining kit (Roche) was used
to analyze apoptosis levels, as previously described (17).
Western blot analysis
The primary antibodies used in the immunoblotting
assays were: FBXW7 (WH0055294M2; Sigma) and GAPDH (G8140; US
Biological, Swampscott, MA, USA). Horseradish peroxidase
(HRP)-conjugated sheep anti-mouse secondary antibodies (Bio-Rad,
Hercules, CA, USA) were used at a 1:1,000–1:5,000 dilution and
detected using a Western Blotting Luminol Reagent (sc-2048; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA), as described in a
previous study (18).
Immunohistochemical staining
Immunohistochemistry was performed on
paraformaldehyde-fixed paraffin sections. FBXW7 (WH0055294M2;
Sigma) antibody was used in immunohistochemistry using a
streptavidin peroxidase-conjugated (SP-IHC) method.
Immunohistochemistry was performed as previously reported (19). The percentage of positive tumor
cells was graded as: 0, <10%; 1, 10–30%; 2, 31–50%; 3,
>50%.
Luciferase reporter assay
The predicted 3′-UTR sequence of FBXW7 that
interacted with miR-92a, together with a corresponding mutated
sequence within the predicted target sites, were created and
inserted into the pRL-TK control vector (Promega, Madison, WI,
USA). SMMC-7721 cells that were seeded in a 96-well plate were
transfected with 120 ng miR-92a expression vector, miR-92a
inhibitor, control vector or negative control. Cells were
co-transfected with 30 ng of the wild-type or mutant 3′-UTR of
FBXW7 mRNA. Transfections were performed using 0.45 µl of
Fugene (Promega). Fourty eight hours after transfection, the cells
were collected and measured according to the manufacturer's
instructions (Dual-Luciferase Assay System; Promega). The pRL-TK
expressing Renilla luciferase was cotransfected as an
internal control to correct the differences in transfection and
harvest efficiencies (20).
In vivo experiments
Four-to-six week-old female BALB/c nude mice (Centre
of Laboratory Animals, The Medical College of Xi'an Jiaotong
University, Xi'an, China) were used to establish the nude mouse
xenograft model. SMMC-7721 (5×106) cells that were
transfected with anti-miR-92a or -miR-NC vectors were mixed in 150
µl of Matrigel and were inoculated subcutaneously into the
flank of nude mice. Tumor growth curves were generated as
previously described (18). Animal
protocols were approved by the Institutional Animal Care and Use
Committee of Xi'an Jiaotong University.
Statistical analysis
Data are presented as the mean ± SEM from at least
three independent replicates. Where appropriate, a Pearson's
Chi-squared test, a Kaplan-Meier plot, a log-rank test, the
multi-variant Cox regression analysis, a Spearman's rank
correlation coefficient and a two-tailed Student's t-test were used
with the SPSS statistical package for Windows Version 13 (SPSS,
Chicago, IL, USA) or GraphPad Prism 5 software (GraphPad Software,
Inc, San Diego, CA, USA). P≤0.05 was considered significant.
Results
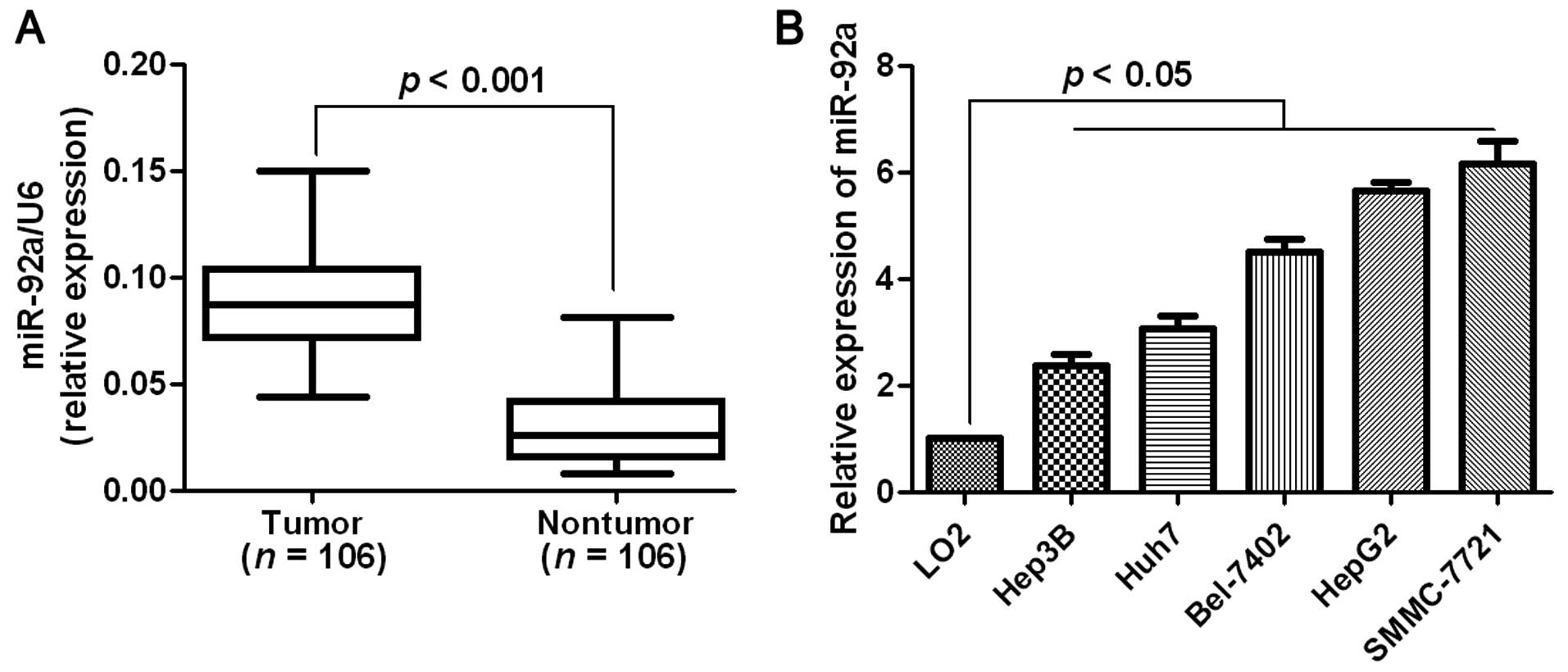
Elevated expression of miR-92a is
observed in HCC tissues and cells
One hundred and six pairs of HCC tissues and matched
tumor-adjacent tissues were tested for miR-92a using RT-qPCR. The
results showed that the mean level of miR-92a expression in HCC
tissues was significantly higher than that in the non-tumor tissues
(P<0.001, Fig. 1A). We then
analyzed miR-92a expression in a non-transformed hepatic cell line
(LO2) and a panel of HCC cell lines (HepG2, Huh7, Hep3B, SMCC-7721
and Bel-7402). As expected, the relative expression of miR-92a was
obviously upregulated in the HCC cell lines as compared with that
in the non-transformed LO2 hepatic cell line (P<0.05, Fig. 1B). The results suggested that an
elevated expression of miR-92a may facilitate
hepatocarcinogenesis.
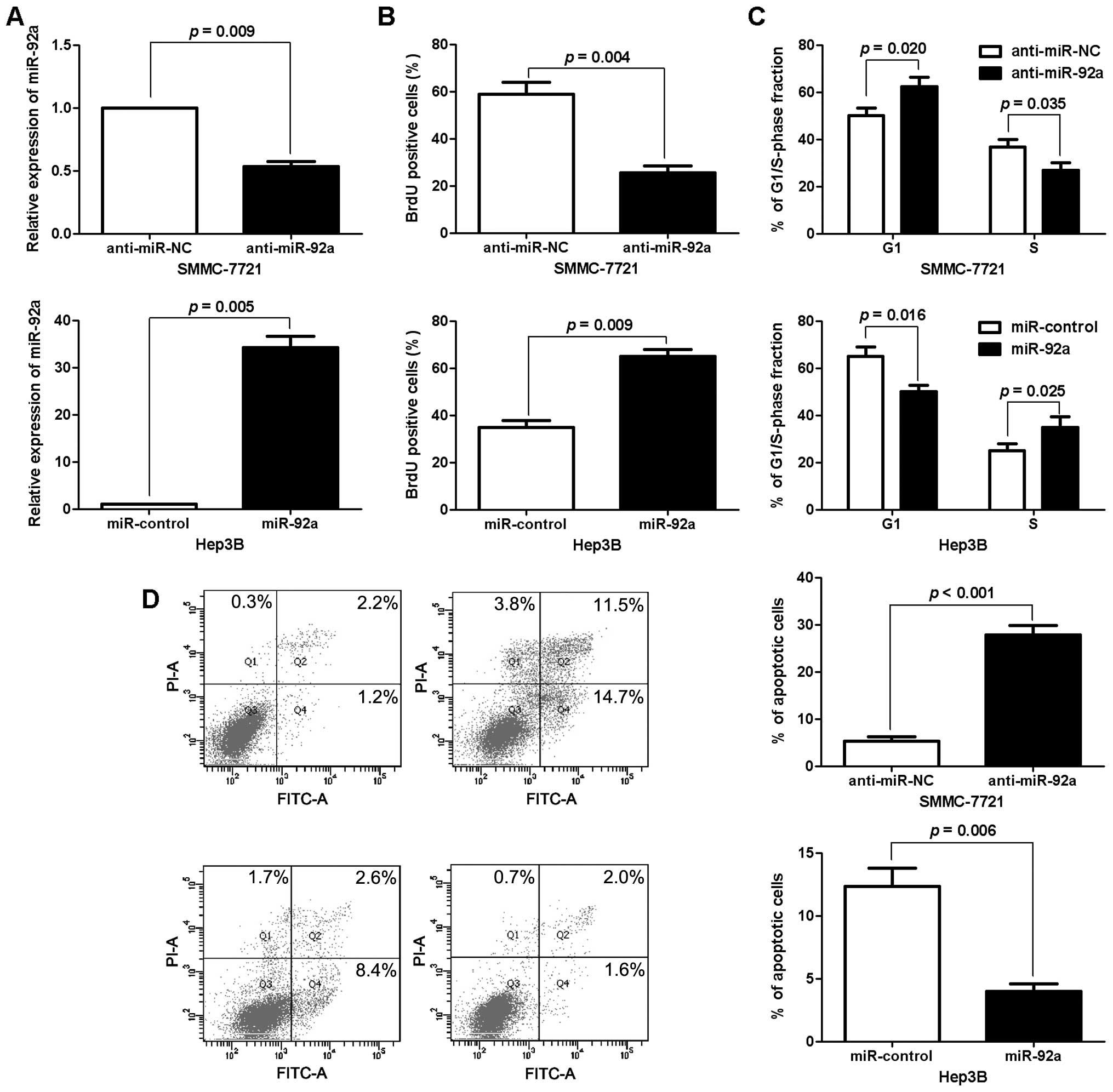
Promotive effects of miR-92a on
proliferation, cell cycle and apoptosis of HCC cells
To identify the biological function of miR-92a in
HCC, we transduced an miR-92a expression plasmid or anti-miR-92a
vector into human HCC cell lines with different endogenous
expression levels of miR-92a. As measured by RT-qPCR, the
expression of miR-92a was significantly altered by the
corresponding vector in HCC cells (P<0.05, respectively,
Fig. 2A). BrdU incorporation assays
were performed to determine the effect of altering miR-92a levels
on HCC cell proliferation. We found that the downregulation of
miR-92a led to a significant reduction of cell proliferation in
SMMC-7721 cells (P<0.05, Fig.
2B). Furthermore, as determined by flow cytometric analysis,
the downregulation of miR-92a resulted in G1 phase arrest in
SMMC-7721 cells (P<0.05, Fig.
2C). Otherwise, the percentage of apoptotic SMMC-7721 cells was
significantly increased following the reduction of miR-92a
(P<0.001, Fig. 2D). By contrast,
the upregulation of miR-92a promoted the proliferation, cell cycle
transition from G1 to S phase and apoptosis resistance in Hep3B
cells (P<0.05, respectively, Fig.
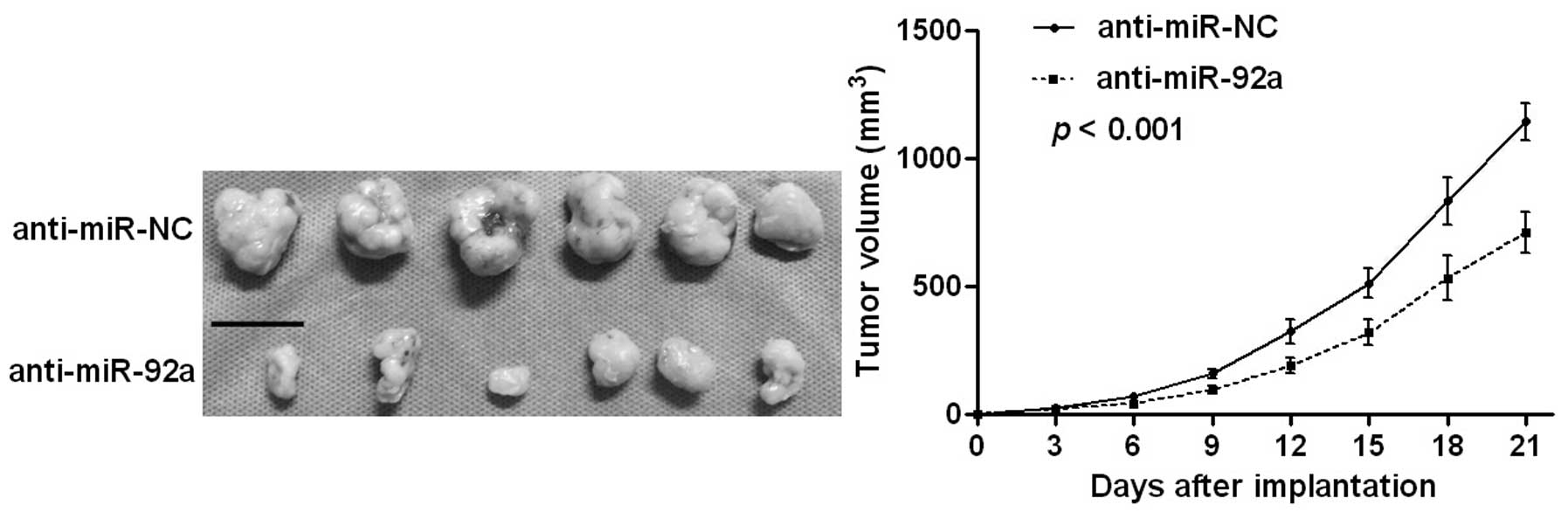
2B–D). Notably, the tumor growth curve revealed that knockdown
of miR-92a significantly retarded the tumor growth of human HCC in
subcutaneous nude mouse models (P<0.001, Fig. 3). These results demonstrated that
miR-92a regulates the proliferation, cell cycle and apoptosis of
HCC cells.
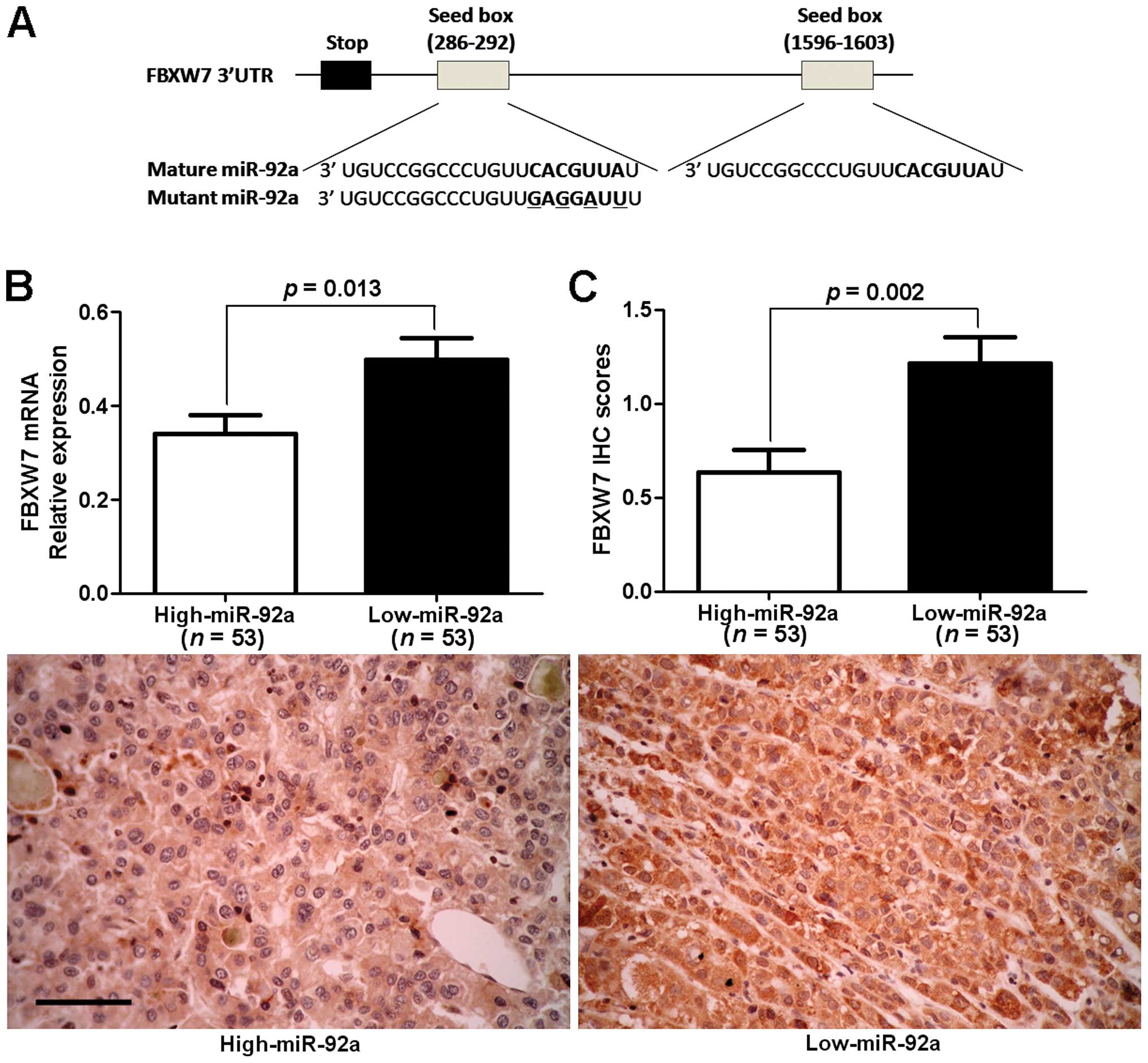
FBXW7 is a direct downstream target of
miR-92a
A search for the candidate target genes of miR-92a
was conducted using publically available databases TargetScan 6.2
(http://www.targetscan.org/) and miRanda
(microrna.org and miRbase). Of note, the
complementary sequence of miR-92a was identified in the 3′-UTR of
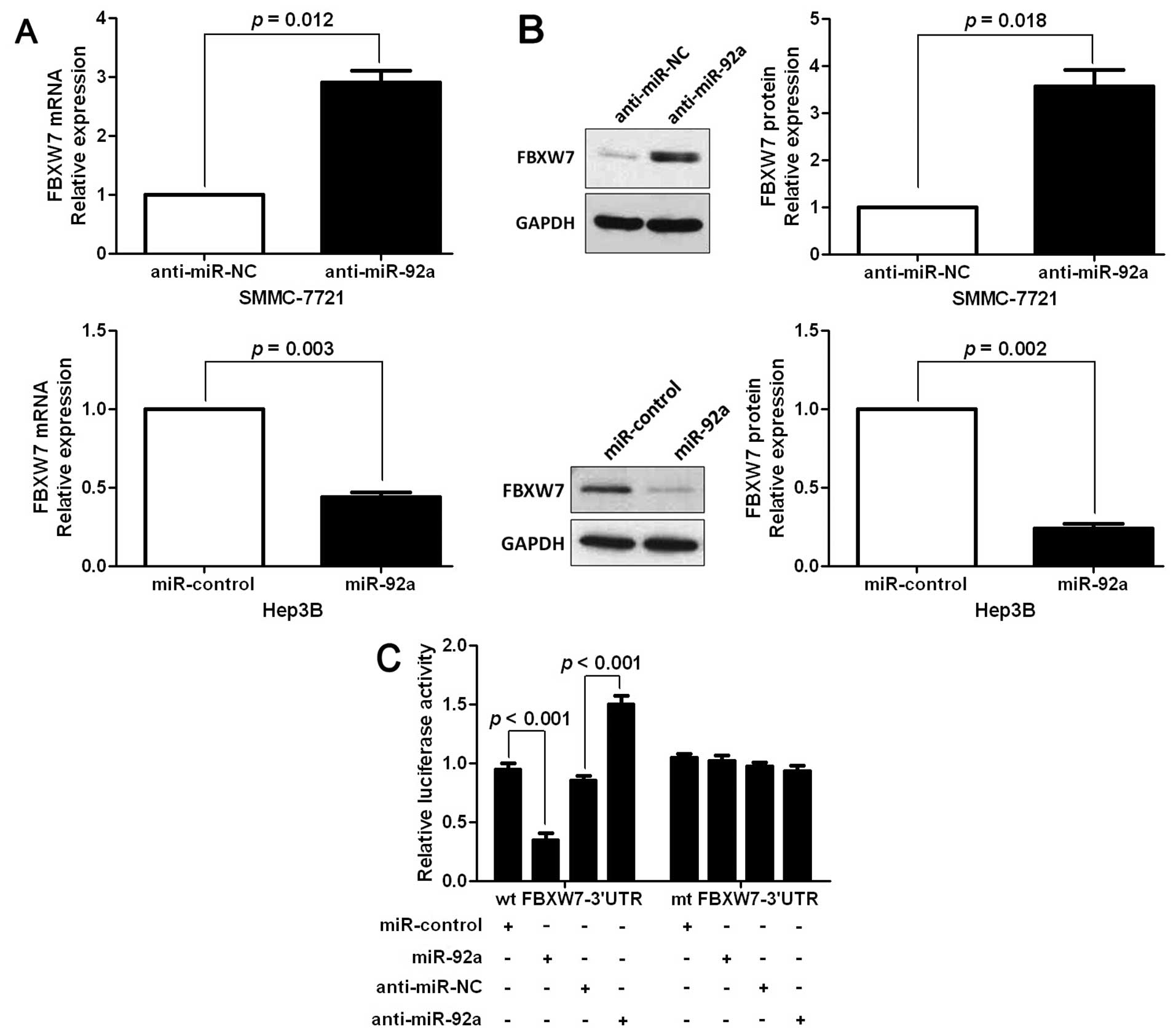
FBXW7 mRNA, which was selected for subsequent studies (Fig. 4A). One hundred and six samples of
HCC tissues were subjected to RT-qPCR and immunostaining for FBXW7
expression. The expression levels of FBXW7 mRNA and protein in the
miR-92a high-expressing tumors were significantly lower than those
in the miR-92a low-expressing tumors (P<0.05, respectively,
Fig. 4B and C). Notably, an obvious
inverse correlation between mRNA levels of miR-92a and FBXW7 (r=
0.745, P<0.001) and between miR-92a mRNA and FBXW7 protein
(r=−0.632, P<0.001) was revealed by Spearman's correlation
analysis in HCC tissues. Furthermore, the expression levels of
FBXW7 mRNA and protein were significantly increased by the
downregulation of miR-92a in SMMC-7721 cells (P<0.05,
respectively, Fig. 5A and B). By
contrast, the overexpression of miR-92a markedly reduced the mRNA
and protein levels of FBXW7 in Hep3B cells (P<0.05,
respectively, Fig. 5A and B). In
addition, the upregulation of miR-92a prominently inhibited the
luciferase activity of FBXW7 containing a wild-type (wt) 3′-UTR but
did not suppress the activity of FBXW7 with a mutant (mt) 3′-UTR
(P<0.001, Fig. 5C). Suppression
of miR-92a by anti-miR-92a increased the luciferase activity of wt
FBXW7 3′-UTR (P<0.001, Fig. 5C).
However, with the mt FBXW7 3′-UTR constructs, there was no relative
increase in activity. Collectively, these results strongly
suggested that FBXW7 is a downstream target of miR-92a in HCC.
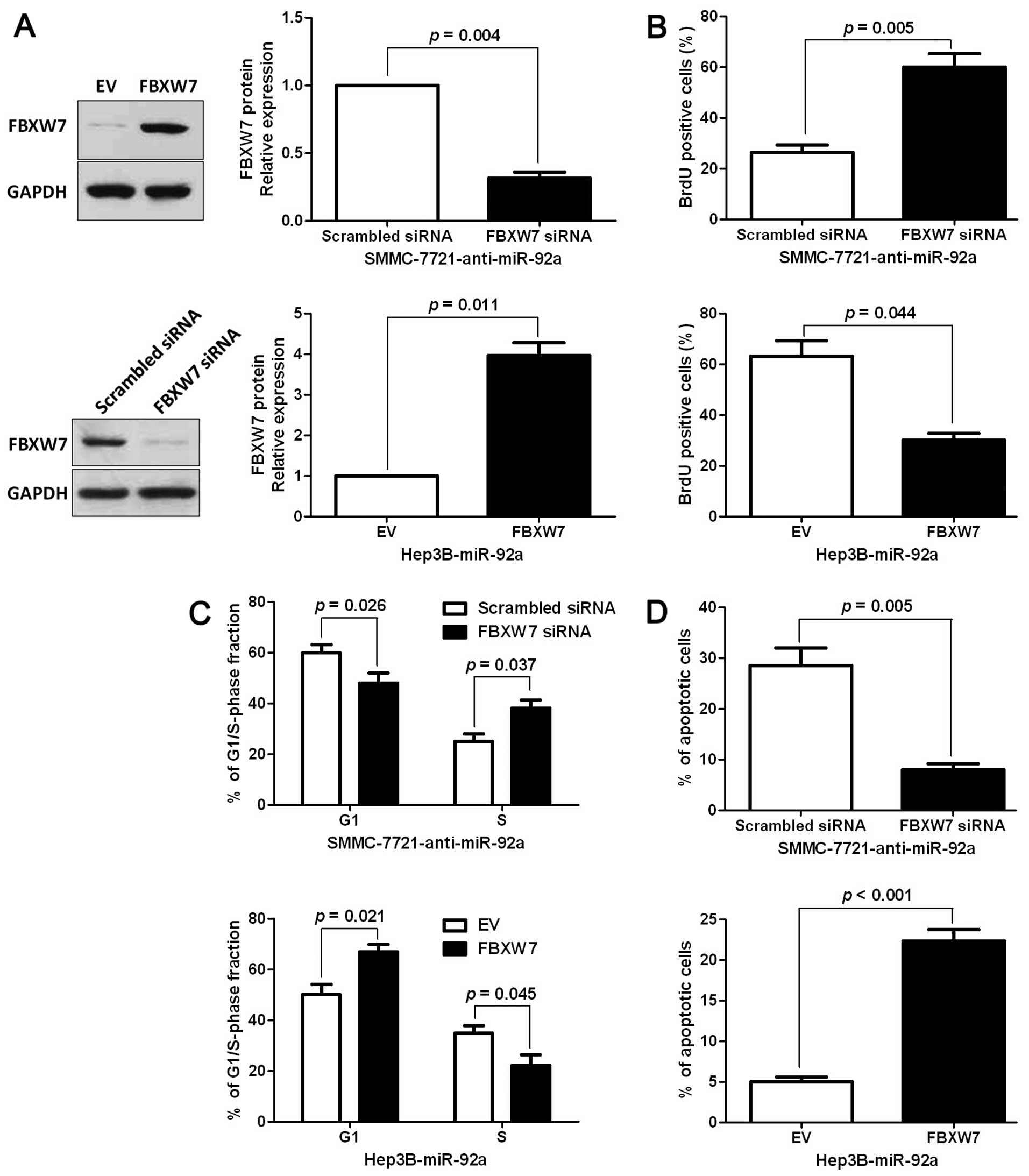
Alterations of FBXW7 levels influence the
effects of miR-92a on HCC cells
To confirm that FBXW7 is a functional target of
miR-92a, FBXW7 was knocked down by a specific siRNA in
miR-92a-suppressive SMMC-7721 cells (P<0.05, Fig. 6A). We found that cell proliferation
was significantly increased by FBXW7 knockdown (P<0.05, Fig. 6B). Furthermore, FBXW7 knockdown
markedly rescued the downregulation of miR-92a-induced cell cycle
arrest and apoptosis (P<0.05, respectively, Fig. 6C and D). Similarly, FBXW7
overexpression inhibited cell proliferation and promoted G1 phase
arrest and apoptosis in miR-92a-overexpressing Hep3B cells
(P<0.05, respectively, Fig.
6A–D). The results showed that FBXW7 is a downstream mediator
of miR-92a in HCC.
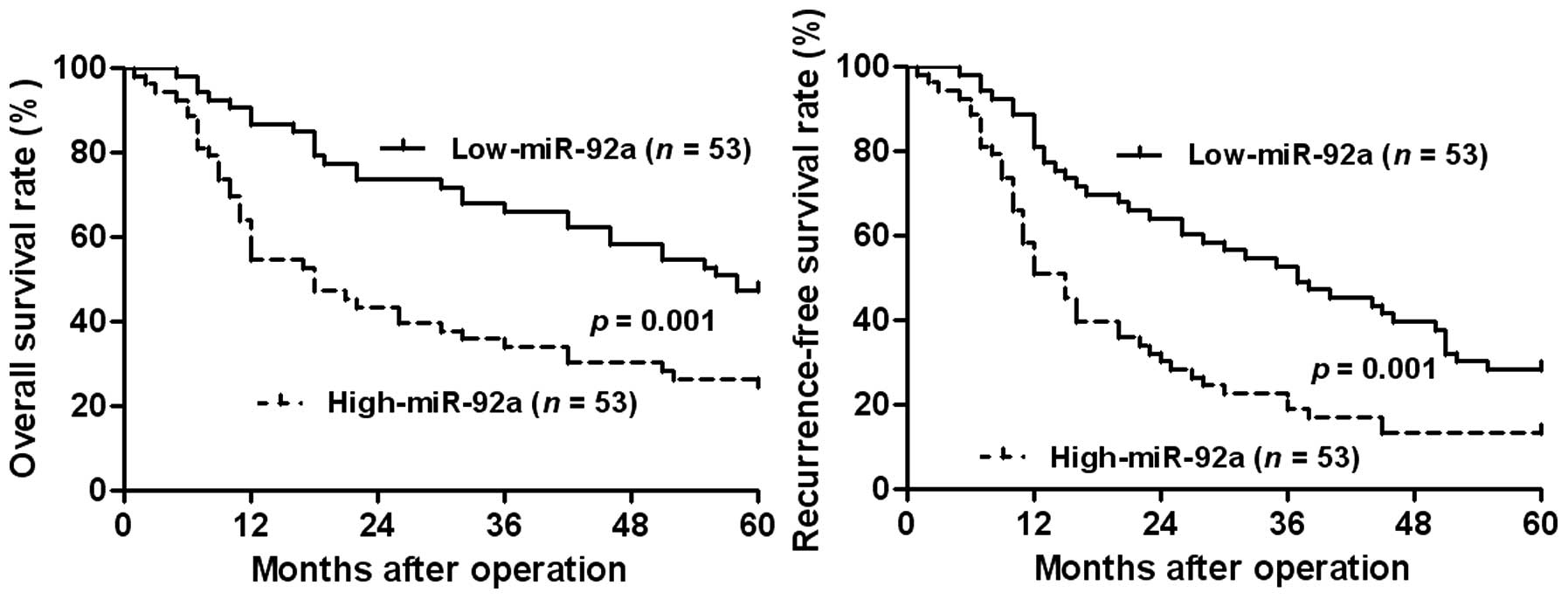
Clinical significance of miR-92a
expression in HCC cases
The expression of miR-92a was considered as low
(n=53) or high (n=53) in accordance with the cut-off value, which
was defined as the median of the cohort. The association between
clinicopathological characteristics and miR-92a expression was
subsequently analyzed. As shown in Table I, a high expression of miR-92a was
evidently associated with HBV infection (P=0.022), large tumor size
(P=0.019), cirrhosis (P=0.020), high Edmondson-Steiner grading
(P=0.002) and advanced tumor-node-metastasis (TNM) tumor stage
(P=0.014). Furthermore, the prognostic value of miR-92a was
analyzed by Kaplan-Meier estimation. A high expression of miR-92a
conferred a poor overall survival (OS) and recurrence-free survival
(RFS) of HCC patients (P=0.001, respectively, Fig. 7). The multi-variant Cox regression
analysis revealed that miR-92a expression was an independent
prognostic marker for predicting 5-year OS and RFS in HCC patients
(P=0.026 and P=0.042, respectively, Table II). These results showed that
miR-92a is a potent biomarker for predicting prognosis of HCC
patients.
| Table IIMultivariate Cox regression analysis
of 5-year OS and RFS of 106 HCC patients. |
Table II
Multivariate Cox regression analysis
of 5-year OS and RFS of 106 HCC patients.
| Variables | OS
| RFS
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age | 0.853 | 0.512–1.44 | 0.523 | 0.869 | 0.558–1.362 | 0.552 |
| Gender | 1.373 | 0.587–3.192 | 0.454 | 0.781 | 0.424–1.479 | 0.455 |
| HBV | 0.934 | 0.396–2.213 | 0.869 | 0.978 | 0.463–2.156 | 0.996 |
| No. of tumor
nodules | 1.761 | 0.966–3.192 | 0.063 | 1.684 | 1.002–2.849 | 0.051 |
| Tumor size | 3.598 | 2.041–6.352 | <0.001a | 2.571 | 1.613–4.076 | <0.001a |
| Venous
infiltration | 4.042 | 2.362–6.931 | <0.001a | 3.531 | 2.159–5.782 | <0.001a |
| Serum AFP
level | 1.554 | 0.872–2.755 | 0.137 | 1.463 | 0.883–2.420 | 0.140 |
| Cirrhosis | 1.008 | 0.561–1.798 | 0.964 | 1.053 | 0.631–1.753 | 0.851 |
| Edmondson-Steiner
grading | 1.241 | 0.702–2.186 | 0.450 | 1.132 | 0.678–1.888 | 0.639 |
| TNM tumor
stage | 2.801 | 1.348–5.814 | 0.006a | 2.198 | 1.030–4.673 | 0.032a |
| miR-92a
expression | 2.283 | 1.104–4.717 | 0.026a | 3.706 | 1.079–5.155 | 0.042a |
Discussion
In previous studies, the deregulation of miR-92a has
been studied in various human types of cancer and it is a
prognostic marker for predicting survival of patients (6,7,9,11,14,16).
miR-92a has been found to regulate HCC cell proliferation (16), facilitate G1/S transition (11,21),
mediate apoptosis (21) and promote
in vivo tumor growth (22).
The clinical significance of miR-92a and the molecular mechanisms
by which miR-92a promotes HCC development remain unclear. In the
present study, we identified that the mean level of miR-92a
expression was obviously increased in HCC tissues vs. matched
tumor-adjacent tissues. Elevated expression of miR-92a was observed
in the HCC cell lines as compared with the normal hepatic cell
line. The role of miR-92a deregulation in tumor growth was examined
via in vitro and in vivo experiments. We found that
the downregulation of miR-92a inhibited HCC cell proliferation and
induced G1 phase arrest and apoptosis in vitro and
suppressed in vivo tumor growth. By contrast, the
overexpression of miR-92a promoted proliferation and cell cycle
transition to S phase and inhibited apoptosis in vitro. The
results suggested that miR-92a is a novel tumor-suppressive miRNA
that plays a critical role in the regulation of tumor growth in
HCC.
Results of the present study suggest that miR-92a
promoted tumor growth, at least in part, by targeting FBXW7.
Firstly, miR-92a was inversely correlated with the levels of both
FBXW7 mRNA and protein in HCC tissues. Secondly, miR-92a negatively
regulated FBXW7 abundance in HCC cells. Thirdly, the complementary
sequence of miR-92a was identified in the 3′-UTR of FBXW7 mRNA.
Knockdown of miR-92a increased the luciferase reporter activity of
wt 3′-UTR but not mt 3′-UTR of FBXW7. Conversely, the
overexpression of miR-92a decreased the luciferase activity of wt
3′-UTR but not mt 3′-UTR of FBXW7. The effects of miR-92a
alteration on proliferation, cell cycle and apoptosis of HCC cells
were also abolished by FBXW7 modulation. In addition, FBXW7 has
been reported to be a direct target of miR-92a in cervical cancer
(11). Collectively, our results
support FBXW7 as a downstream mediator of miR-92a function in
HCC.
Our previous findings have shown that FBXW7 is a
potent tumor suppressor and inhibits tumor growth of HCC by
inducing growth arrest and apoptosis in vitro and in
vivo (17–19). FBXW7 is a well-known E3 ligase and
recognizes target proteins for ubiquitination and degradation.
Several oncoproteins have been identified as substrates of FBXW7
including c-Myc (23) and Cyclin E
(24). Impaired expression of FBXW7
leads to the accumulation of these oncoproteins, which promote
cancer cell proliferation, cell cycle progression and apoptosis
resistance. c-Myc protein plays crucial roles in mitogenic
signaling and cell growth responses (25) and Cyclin E is a key component of the
cell cycle machinery (26). Active
Cyclin E-Cdk2 complex is required for G1 and S-phase transition,
modulating pRb and thereby activating E2F transcription factors
that enable DNA replication (27).
It has been reported that c-Myc can induce apoptosis by
Caspase-3-dependent and caspase-independent signaling (28). Upregulation of c-Myc and Cyclin E
have been reported in HCC and lead to rapid tumor growth in
vitro and in vivo (29,30).
Thus, upregulation of c-Myc and Cyclin E may be a mechanism by
which miR-92a promotes the proliferation and cell cycle transition
from G1 to S phase and inhibits apoptosis of HCC. Alternatively,
our previous results showed that Yes-associated protein (YAP) is a
potential target of FBXW7 and is involved in FBXW7-induced growth
arrest and apoptosis of HCC (18).
It has been shown that YAP plays critical role in
hepatocarcinogenesis (31). This
mechanism may account for the effects of miR-92a in HCC.
The role of miR-92a in cancer cell proliferation,
cell cycle and apoptosis suggests its potential application as a
prognostic biomarker of HCC patients. Furthermore, our previous
findings showed that a low expression of FBXW7 was an independent
prognostic marker for predicting the survival of HCC patients
(18). Thus, we demonstrated, for
the first time, that high expression of miR-92a was associated with
poor prognostic characteristics including HBV infection, large
tumor size, cirrhosis, high Edmondson-Steiner grading and advanced
TNM tumor stage. We also confirmed that a high expression of
miR-92a was an independent prognostic marker for predicting 5-year
OS and RFS of HCC patients.
In conclusion, the results show that the expression
of miR-92a is reduced in HCC tissues and cell lines. We confirm
that miR-92a is an independent prognostic marker for HCC. In
vitro and in vivo studies revealed that miR-92a
functions as a novel oncomiRNA by promoting proliferation, cell
cycle transition and apoptosis resistance of HCC. Its multiple
tumor-promotive effects are mediated by FBXW7. Collectively, the
deregulation of miR-92a may play an important role in tumor growth
and may be a novel prognostic factor and potential therapeutic
target for HCC.
Acknowledgments
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 81402039).
References
|
1
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Szabo G and Bala S: MicroRNAs in liver
disease. Nat Rev Gastroenterol Hepatol. 10:542–552. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mendell JT: MicroRNAs: Critical regulators
of development, cellular physiology and malignancy. Cell Cycle.
4:1179–1184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tu K, Zheng X, Dou C, Li C, Yang W, Yao Y
and Liu Q: MicroRNA-130b promotes cell aggressiveness by inhibiting
peroxisome proliferator-activated receptor gamma in human
hepatocellular carcinoma. Int J Mol Sci. 15:20486–20499. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ranade AR, Cherba D, Sridhar S, Richardson
P, Webb C, Paripati A, Bowles B and Weiss GJ: MicroRNA
92a-2*: A biomarker predictive for chemoresistance and
prognostic for survival in patients with small cell lung cancer. J
Thorac Oncol. 5:1273–1278. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen ZL, Zhao XH, Wang JW, Li BZ, Wang Z,
Sun J, Tan FW, Ding DP, Xu XH, Zhou F, et al: microRNA-92a promotes
lymph node metastasis of human esophageal squamous cell carcinoma
via E-cadherin. J Biol Chem. 286:10725–10734. 2011. View Article : Google Scholar :
|
|
8
|
Wu CW, Ng SS, Dong YJ, Ng SC, Leung WW,
Lee CW, Wong YN, Chan FK, Yu J and Sung JJ: Detection of miR-92a
and miR-21 in stool samples as potential screening biomarkers for
colorectal cancer and polyps. Gut. 61:739–745. 2012. View Article : Google Scholar
|
|
9
|
Nilsson S, Möller C, Jirström K, Lee A,
Busch S, Lamb R and Landberg G: Downregulation of miR-92a is
associated with aggressive breast cancer features and increased
tumour macrophage infiltration. PLoS One. 7:e360512012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ohyagi-Hara C, Sawada K, Kamiura S, Tomita
Y, Isobe A, Hashimoto K, Kinose Y, Mabuchi S, Hisamatsu T,
Takahashi T, et al: miR-92a inhibits peritoneal dissemination of
ovarian cancer cells by inhibiting integrin α5 expression. Am J
Pathol. 182:1876–1889. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou C, Shen L, Mao L, Wang B, Li Y and Yu
H: miR-92a is upregulated in cervical cancer and promotes cell
proliferation and invasion by targeting FBXW7. Biochem Biophys Res
Commun. 458:63–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ranji N, Sadeghizadeh M, Shokrgozar MA,
Bakhshandeh B, Karimipour M, Amanzadeh A and Azadmanesh K:
MiR-17-92 cluster: An apoptosis inducer or proliferation enhancer.
Mol Cell Biochem. 380:229–238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin HY, Chiang CH and Hung WC: STAT3
upregulates miR-92a to inhibit RECK expression and to promote
invasiveness of lung cancer cells. Br J Cancer. 109:731–738. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ke TW, Wei PL, Yeh KT, Chen WT and Cheng
YW: MiR-92a Promotes Cell Metastasis of Colorectal Cancer Through
PTEN-Mediated PI3K/AKT Pathway. Ann Surg Oncol. 22:2649–2655. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li LM, Hu ZB, Zhou ZX, Chen X, Liu FY,
Zhang JF, Shen HB, Zhang CY and Zen K: Serum microRNA profiles
serve as novel biomarkers for HBV infection and diagnosis of
HBV-positive hepatocarcinoma. Cancer Res. 70:9798–9807. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shigoka M, Tsuchida A, Matsudo T, Nagakawa
Y, Saito H, Suzuki Y, Aoki T, Murakami Y, Toyoda H, Kumada T, et
al: Deregulation of miR-92a expression is implicated in
hepatocellular carcinoma development. Pathol Int. 60:351–357. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tu K, Zheng X, Zhou Z, Li C, Zhang J, Gao
J, Yao Y and Liu Q: Recombinant human adenovirus-p53 injection
induced apoptosis in hepatocellular carcinoma cell lines mediated
by p53-Fbxw7 pathway, which controls c-Myc and cyclin E. PLoS One.
8:e685742013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tu K, Yang W, Li C, Zheng X, Lu Z, Guo C,
Yao Y and Liu Q: Fbxw7 is an independent prognostic marker and
induces apoptosis and growth arrest by regulating YAP abundance in
hepatocellular carcinoma. Mol Cancer. 13:1102014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tu K, Zheng X, Zan X, Han S, Yao Y and Liu
Q: Evaluation of Fbxw7 expression and its correlation with the
expression of c-Myc, cyclin E and p53 in human hepatocellular
carcinoma. Hepatol Res. 42:904–910. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Z, Tu K and Liu Q: Effects of
microRNA-30a on migration, invasion and prognosis of hepatocellular
carcinoma. FEBS Lett. 588:3089–3097. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lv XB, Zhang X, Deng L, Jiang L, Meng W,
Lu Z and Wang X: MiR-92a mediates AZD6244 induced apoptosis and
G1-phase arrest of lymphoma cells by targeting Bim. Cell Biol Int.
38:435–443. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li M, Guan X, Sun Y, Mi J, Shu X, Liu F
and Li C: miR-92a family and their target genes in tumorigenesis
and metastasis. Exp Cell Res. 323:1–6. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Welcker M, Orian A, Jin J, Grim JE, Harper
JW, Eisenman RN and Clurman BE: The Fbw7 tumor suppressor regulates
glycogen synthase kinase 3 phosphorylation-dependent c-Myc protein
degradation. Proc Natl Acad Sci USA. 101:9085–9090. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Koepp DM, Schaefer LK, Ye X, Keyomarsi K,
Chu C, Harper JW and Elledge SJ: Phosphorylation-dependent
ubiquitination of cyclin E by the SCFFbw7 ubiquitin ligase.
Science. 294:173–177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Grandori C, Cowley SM, James LP and
Eisenman RN: The Myc/Max/Mad network and the transcriptional
control of cell behavior. Annu Rev Cell Dev Biol. 16:653–699. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hwang HC and Clurman BE: Cyclin E in
normal and neoplastic cell cycles. Oncogene. 24:2776–2786. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ohtsubo M, Theodoras AM, Schumacher J,
Roberts JM and Pagano M: Human cyclin E, a nuclear protein
essential for the G1-to-S phase transition. Mol Cell Biol.
15:2612–2624. 1995.PubMed/NCBI
|
|
28
|
Prendergast GC: Mechanisms of apoptosis by
c-Myc. Oncogene. 18:2967–2987. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sargent LM, Zhou X, Keck CL, Sanderson ND,
Zimonjic DB, Popescu NC and Thorgeirsson SS: Nonrandom cytogenetic
alterations in hepatocellular carcinoma from transgenic mice
overexpressing c-Myc and transforming growth factor-alpha in the
liver. Am J Pathol. 154:1047–1055. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li K, Lin SY, Brunicardi FC and Seu P: Use
of RNA interference to target cyclin E-overexpressing
hepatocellular carcinoma. Cancer Res. 63:3593–3597. 2003.PubMed/NCBI
|
|
31
|
Xu MZ, Chan SW, Liu AM, Wong KF, Fan ST,
Chen J, Poon RT, Zender L, Lowe SW, Hong W, et al: AXL receptor
kinase is a mediator of YAP-dependent oncogenic functions in
hepatocellular carcinoma. Oncogene. 30:1229–1240. 2011. View Article : Google Scholar
|