Introduction
Long non-coding RNAs (lncRNAs) are non-protein
coding transcripts containing >200 nucleic acids (1). LncRNA functions to regulate gene
transcription via post-transcriptional modification and interacts
with microRNA (miRNA). LncRNA plays a crucial role in the
regulation of human carcino genesis (2–5). In
hepatocellular carcinoma (HCC), an altered lncRNA expression is
associated with tumor progression. Therefore, lncRNA is a potential
target for HCC treatment and should be investigated in HCC
development and progression (6–10).
The metastasis-associated lung adenocarcinoma
transcript 1 (MALAT1) is highly conserved among mammals and
primarily expressed in the nucleus (11–14).
MALAT1 overexpression is associated with elevated levels of
α-fetoprotein (AFP) and a number of tumor lesions, and has been
identified in host reaction after liver transplantation (15). MALAT1 interacts with various target
genes, such as serine/arginine-rich splicing factor 1 (SRSF1) and
potential gene signaling pathways associated with human
carcinogenesis (11–14). Nevertheless, the transcriptional
regulation of MALAT1, per se, remains unknown.
The sequence database, ChIPBase, was established
using the result of 543 ChIP-Seq experiments. Analysis of this
database revealed the potential regulation between transcription
factors and non-coding RNA (ncRNA), indicating a possible
connection of a well-known transcription factor specificity protein
1 (Sp1) with MALAT1 expression (16). Moreover, Sp3 recognizes similar
amino acid sequences of Sp1 DNA-binding domains (17–19).
In the present study, we investigated the role of Sp1/3 in the
regulation of MALAT1 transcription in HCC cells. Our findings
provide mechanistic information for future investigations of the
regulation of HCC progression mediated by Sp1/3 transcription of
MALAT1 expression.
Materials and methods
Patient samples
Thirty-two fresh HCC and paired distant non-tumor
liver tissues were randomly selected from histologically confirmed
HCC patients from the First Affiliated Hospital of Guangxi Medical
University (Guangxi, China) between March and November 2014. The
paired non-tumor tissues were at least 2 cm away from the tumor
lesion. The current study was approved by the Ethics Committee of
the First Affiliated Hospital of Guangxi Medical University.
Written informed consent was obtained from all the patients.
Clinicopathological characteristics were collected and are provided
in Table I. The mean of mRNA
expression was set as the cut-off value to define low or high
expression (>mean, high; <mean, low) (Table I). The relationship between Sp
expression and MALAT1 expression are shown in Table II.
 | Table IAssociation of Sp1, Sp3 and MALAT1
expression with clinicopathological characteristics of HCC
patients. |
Table I
Association of Sp1, Sp3 and MALAT1
expression with clinicopathological characteristics of HCC
patients.
|
Characteristics | N | Sp1 expression
| Sp3 expression
| MALAT1 expression
|
|---|
| Low | High | P-valuea | Low | High | P-valuea | Low | High | P-valuea |
|---|
| Age (years) |
| ≤50 | 18 | 9 | 9 | 0.73 | 10 | 8 | 0.30 | 10 | 8 | 0.29 |
| >50 | 14 | 8 | 6 | | 5 | 9 | | 4 | 10 | |
| Gender |
| Female | 2 | 0 | 2 | 0.21 | 0 | 2 | 0.48 | 0 | 2 | 0.49 |
| Male | 30 | 17 | 13 | | 15 | 15 | | 14 | 16 | |
| AFP (ng/ml) |
| ≤400 | 11 | 10 | 1 | 0.0027 | 5 | 6 | 1 | 10 | 1 | 1.42E-04 |
| >400 | 21 | 7 | 14 | | 10 | 11 | | 4 | 17 | |
| HBV |
| − | 3 | 0 | 3 | 0.091 | 1 | 2 | 1 | 0 | 3 | 0.23 |
| + | 29 | 17 | 12 | | 14 | 15 | | 14 | 15 | |
| Tumor size
(cm) |
| ≤5 | 12 | 5 | 7 | 0.46 | 5 | 7 | 0.72 | 4 | 8 | 0.47 |
| >5 | 20 | 12 | 8 | | 10 | 10 | | 10 | 10 | |
| No. of tumor |
| Single | 19 | 7 | 12 | 0.035 | 12 | 7 | 0.035 | 10 | 9 | 0.28 |
| Multiple | 13 | 10 | 3 | | 3 | 10 | | 4 | 9 | |
| Histological
grade |
| Well +
moderate | 25 | 13 | 12 | 1 | 13 | 12 | 0.40 | 11 | 14 | 1 |
| Poor | 7 | 4 | 3 | | 2 | 5 | | 3 | 4 | |
| PVTT |
| Absent | 24 | 13 | 11 | 1 | 12 | 12 | 0.69 | 10 | 14 | 0.70 |
| Present | 8 | 4 | 4 | | 3 | 5 | | 4 | 4 | |
| Cirrhosis |
| Absent | 10 | 7 | 3 | 0.26 | 4 | 6 | 0.71 | 4 | 6 | 1 |
| Present | 22 | 10 | 12 | | 11 | 11 | | 10 | 12 | |
| Table IIAssoication between Sp and MALAT1
expression. |
Table II
Assoication between Sp and MALAT1
expression.
| Factors | MALAT1 expression
| P-valuea | r | P-valuea |
|---|
| Low | High |
|---|
| Sp1 |
| Low | 10 | 7 | 0.5488 | 0.32 | 0.0709 |
| High | 4 | 11 | | | |
| Sp3 |
| Low | 9 | 6 | 1.0000 | 0.31 | 0.0867 |
| High | 5 | 12 | | | |
RNA isolation and RT-qPCR
Total cell RNA was isolated from fresh tissues or
cells using a TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and
reverse transcribed into cDNA using a Thermo Scientific RevertAid
First Strand cDNA Synthesis kit (Thermo Scientific, Waltham, MA,
USA) according to the manufacturer's instructions. qPCR was
performed in a Light Cycler 480 (Roche, Basel, Switzerland) with a
SYBR-Green Premix Ex Taq (Roche). β-actin mRNA was used as an
internal control. The primers used were: Sp1,
5′-TCCAGACCATTAACCTCAGTGC-3′ and 5′-TGTATTCCATCACCACCAGCC-3′; Sp3,
5′-GCTTGCACCTGTCCCAACTGTA3′ and 5′-CTCCAGAATGCCAACGCAGA-3′; MALAT1,
5′-CGCATTTACTAAACGCAGAC-3′ and 5′-TCTCTATTCTTTTCTTCGCC-3′; β-actin,
5′-GCACCACACCTTCTACAATGAGC-3′ and
5′-GGATAGCACAGCCTGGATAGCAAC-3′.
Cell lines and culture
Human embryonic kidney cells HEK293T, human normal
liver cells L-02, and HCC cell lines Bel-7402, Huh7, and HepG2 were
obtained from the Shanghai Institute of Cell Biology (Shanghai,
China) and cultured in Dulbecco's modified Eagle's medium (DMEM)
with high glucose (Wisent, Nanjing, China) supplemented with 10%
fetal bovine serum (Sijiqing, Hangzhou, China) in a humidified
incubator with 5% CO2 at 37°C.
Protein extraction and western
blotting
The total cell protein was extracted and western
blotting was performed as previously described (20). The primary antibodies anti-Sp1 and
anti-Sp3 IgG were obtained from Biolegend (San Diego, CA, USA). The
horseradish peroxidase-conjugated goat anti-rabbit IgG secondary
antibody was purchased from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA), and the Pierce ECL Western Blotting Substrate was
purchased from Thermo Scientific. The western blot analysis results
were quantified using Quantity One 4.62 (Bio-Rad, Hercules, CA,
USA).
Small interfering RNA (siRNA) and gene
silencing
SiRNAs targeting Sp1 and Sp3 were obtained from
GeneGeme (Shanghai, China). HCC cell lines Bel-7402, Huh7, and
HepG2 were seeded in 6-well plates at a density of 1×105
cells/well, cultured overnight and transfected with lentivirus
(siSp1 and siSp3) adding 5 ug/ml polybrene according to the
manufacturer's instructions. The final MOI value of Sp1 and Sp3
lentivirus was 50 nM. After 72 h transfection, the cells were
harvested and subjected to RT-qPCR and western blot analysis.
Cell viability assay
Cell viability was assayed using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
cell proliferation/viability assay kit (R&D Systems,
Minneapolis, MN, USA) according to the manufacturer's instructions.
Briefly, the cells were treated with different concentrations
(50–250 nm) of mithramycin A (MIT), (Sigma-Aldrich, St. Louis, MO,
USA) and control medium (dimethyl sulfoxide). The 50% inhibition
concentration (IC50) at 96 h was calculated using
Graphpad Prism 5.0. (GraphPad Software, La Jolla, CA, USA).
Chromatin immunoprecipitation (ChIP)
assay
Approximately 5×106 cells were harvested
and cross-linked with 1% formaldehyde for 10 min, washed in cold
PBS and resuspended in a lysis buffer. The cells were sonicated to
obtain 200 and 1,000 bp chromatin fragments using the Sonics
Sonication Instrument (Bioruptor, Diagenode, Liege, Belgium). The
sonicated chromatin samples were then resuspended in an
immunoprecipitation buffer and incubated overnight at 4°C with
magnetic beads conjugated to antibodies for Sp1 (17–601; Millipore)
or Sp3 D-20 (SC-644; Santa Cruz Biotechnology, Inc.). On the
following day, the samples were washed with a lysis buffer, LiCl
buffer and TE buffer, respectively, and then eluted in an elution
buffer. The DNA from these samples was recovered by reversing the
crosslinks and purified by a Qiagen purification kit (Hilden,
Germany). The immunoprecipitated DNA samples were amplified using
qPCR with primers, specifically for MALAT1 promoter sequences
(Table III). An unenriched DNA
sample treated in a similar manner was used as the input
control.
| Table IIIPrimers for the ChIP assay. |
Table III
Primers for the ChIP assay.
| Region name | Region (bp) | Primer |
|---|
| MALAT1-1 | −894 to −811 |
5′-TTACAGGAGCCAAAGGAGTTT-3′
5′-CTGTATAGGTTAGGATGGCAAAA-3′ |
| MALAT1-2 | −1011 to −741 |
5′-TCAACAGGCCCTGCTTTATG-3′
5′-CCGGGTCTTTGGAACCTGT-3′ |
| MALAT1-3 | −772 to −543 |
5′-CCTCGGAGTTGACTGCCTA-3′
5′-AATATCTTCGTCGTTTGTATGTCA-3′ |
| MALAT1-4 | −285 to −4 |
5′-CAGGCACAGGCGTTAGGG-3′
5′-AGTCTCGGGCTGCAGGCT-3′ |
| MALAT1-5 | −85 to −4 |
5′-CGTTTGTCCCTGACGCAG-3′
5′-AGTCTCGGGCTGCAGGC-3′ |
Electrophoretic mobility shift assay
(EMSA)
EMSA was performed using a ProteoJET Cytoplasmic and
Nuclear Protein Extraction kit (Fermentas, Beijing, China)
according to the manufacturer's instructions. The EMSA for Sp1 and
Sp3 was performed using a non-radioactive gel shift assay system
(Viagene Biotech, Beijing, China). Briefly, nuclear extracts were
incubated with biotin-labeled or unlabeled Sp1 and Sp1/3
oligonucleotides (Sp1, 5′-ccagtggcgcccgcccacgagccag-3′; Sp1/3,
5′-tccctccccgcccccgctctcccct-3′, produced by Viagene Biotech).
Nuclear extracts and 80-fold excess of each competing probe were
mixed together and incubated at room temperature for 20 min
followed by the addition of 15 µl of each labeled probe for
20 min. The control sample was also incubated with unlabeled Sp1/3
probe. Protein/DNA complexes were then separated in a 6%
non-denaturing poly-acrylamide gel by electrophoresis. The gel was
exposed to an X-ray film and digitized using a ChemiDoc XRS+ with
Image Lab software (Bio-Rad).
Luciferase assay
LncRNA MALAT1 promoter region covering -400
to -1 bp was amplified from the pcDNA3.1 (GV141) plasmid
(Invitrogen) and then inserted into pGL3 plasmid (Promega, Madison,
WI, USA). After DNA sequencing, the confirmed plasmid was
transiently transfected into HEK293T cells using Lipofectamine 2000
(Invitrogen) according to the manufacturer's instructions. To
detect luciferase reporter activities, HEK293T cells were harvested
48 h after transfection. Luciferase activity was measured using the
Dual Luciferase reporter assay system (Promega) according to the
manufacturer's instructions.
Statistical analysis
The experiments were performed independently at
least three times and the data were statistically analyzed using
SPSS 20.0 software (SPSS, Inc., Chicago, IL, USA). Data were
presented as means ± standard deviations (SD) and analyzed by a
Mann-Whitney-Wilcoxon test, Fisher's exact test, McNemar's test,
Student's t-test, or one-way ANOVA. Spearman's correlation was
applied to assess the relationship of gene expression with
clinicopathological parameters. P≤0.05 was considered statistically
significant.
Results
Upregulation of Sp1, Sp3 and MALAT1
expression in HCC tissues and cells
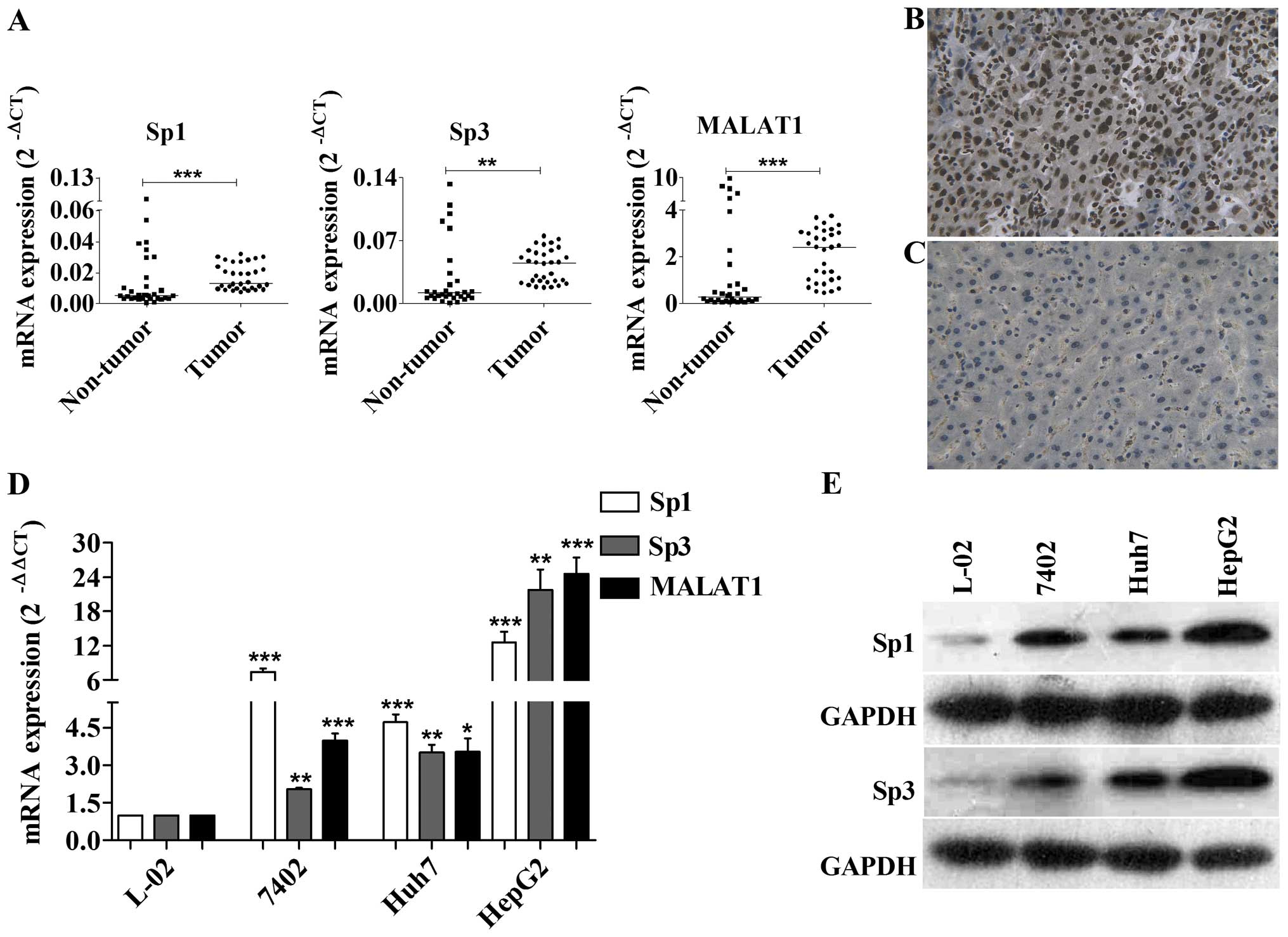
We first measured the expression level of Sp1, Sp3
and MALAT1 in 32 paired HCC and non-tumor liver tissues and found
that the expression of Sp1, Sp3 and MALAT1 was significantly
upregulated in cancer tissues compared to the non-tumor tissues
(Fig. 1A). Expression of Sp1 and
MALAT1 was significantly associated with the preoperative AFP level
(Sp1, r=7.44, P=0.0064; MALAT1, r=12.37, P=0.0004) (Table I). The relationship between Sp and
MALAT1 expression had no obvious statistical significance (Sp1 +
MALAT1, r= 0.32, P=0.0709; Sp3 + MALAT1, r=0.31, P=0.0867)
(Table II). Our results also
showed that a high Sp3 positive immunocytochemical reaction and
nuclear staining were observed (Fig. 1B
and C). Their expression was also much higher in the three
tested HCC cell lines compared to the normal hepatocellular
epithelium (Fig. 1D and E).
Silencing Sp1 and Sp3 expression inhibits
MALAT1 transcription in HCC cells
To investigate whether Sp1 and Sp3 silencing reduced
the endogenous expression of the MALAT1 transcript, we transfected
Sp1 and Sp3 siRNA into Bel-7402, Huh7 and HepG2 cells and found
that after 72 h, Sp1 and Sp3 mRNA and protein levels were
significantly decreased. Notably, silencing of Sp1 or Sp3 alone did
not significantly affect the level of MALAT1 in HCC cells, whereas
silencing Sp1 and Sp3 significantly inhibited MALAT1 expression by
70–85% (Fig. 2A and B). Taken
together, these data indicated that Sp1 cooperated with Sp3 to
influence the basal endogenous expression of MALAT1.
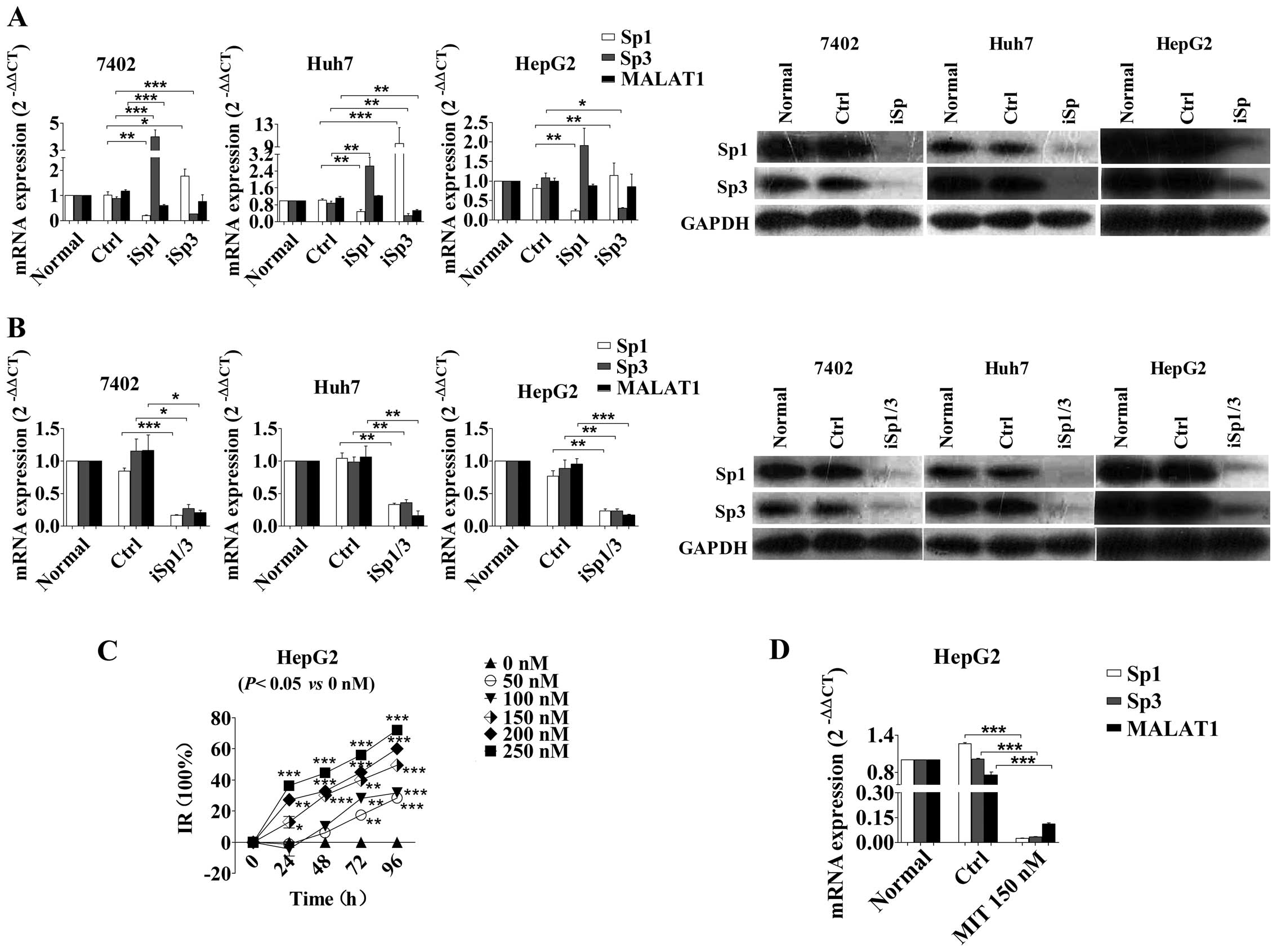 | Figure 2Effects of Sp1 and Sp3 knockdown on
the regulation of MALAT1 level. (A) Bel-7402, Huh7 and HepG2 cells
were transfected with Sp1- or Sp3-specific siRNA and subjected to
RT-qPCR and western blot analysis. *P<0.05,
**P<0.01, ***P<0.001. (B) Effect of
combined Sp1 and Sp3 knockdown on the regulation of MALAT1
expression. *P<0.05, **P<0.01,
***P<0.001. (C) HepG2 cells were treated with
different doses of MIT for up to 96 h and then subjected to cell
viability assay. *P<0.05, **P<0.01,
***P<0.001. (D) HepG2 cells were treated with 150 nM
MIT for 72 h and subjected to RT-qPCR analysis of Sp1, Sp3 and
MALAT1 mRNA levels. *P<0.05, **P<0.01,
***P<0.001. MALAT1, metastasis-associated lung
adenocarcinoma transcript 1; Sp, specificity protein; MIT,
mithramycin A. |
Furthermore, we treated HepG2 HCC cells with
different concentrations (0, 50, 100, 150, 200, and 250 nM) of MIT
(21), an FDA-approved
chemotherapeutic anticancer drug that inhibits the transcriptional
activity of Sp1 by competitively binding to the Sp1-binding sites.
Our data showed that cell viability was significantly reduced
(Fig. 2C) and the IC50
at 96 h was 146.7 nM. The transcription of MALAT1 decreased in
HepG2 cells incubated with 150 nM MIT for 96 h (Fig. 2D).
Sp1/3 binding sites in the MALAT1
proximal promoter region
Online tools were used to predict Sp1/3 binding
sites in the MALAT1 proximal promoter region. These tools
included the TFSEARCH (http://www.cbrc.jp/research/db/TFSEARCH.html),
MatInspector (http://www.genomatix.de/en/produkte/genomatix-sofware-suite.html),
Tfsitescan (http://www.ifti.org/cgi-bin/ifti/Tfsitescan.pl) and
CONSITE (http://asp.ii.uib.no:8090/cgibin/CONSITE/consite).
We identified potential Sp1/3 binding sites upstream of MALAT1
between positions -1,000 bp and -1 bp, especially in the region
-400 to -1 bp. This region contained four similarly high-scored
sites from different online tools (Fig.
3A). From ChIPBase (http://deepbase.sysu.edu.cn/chipbase/), the first
website for decoding the transcriptional regulation of lncRNA, we
found the binding regions of MALAT1 to Sp1. The major predicted
results are shown in Fig. 3B.
Identification of Sp1/3 binding site in
the MALAT1 proximal promoter region using ChIP and EMSA
To confirm the Sp1 and Sp3 binding sites in the
MALAT1 promoter region, we first performed a ChIP assay to
detect the interaction of Sp1/3 with endogenous MALAT1
promoter. Chromatin fragments bound with transcription factor were
prepared from HepG2 cells. Sp1 and Sp3 antibodies were separately
used to immunoprecipitate Sp1- or Sp3-bound chromatin fragments
followed by qPCR with primers corresponding to five different
regions of the MALAT1 promoter. Promoter regions from MALAT1-1 to
MALAT1-5 showed an extremely high affinity, especially the region
-400 to -1 bp (MALAT1-4 and MALAT1-5) (Fig. 4B), which contained the predicted
binding sites nos. 2, 3 and 4. The MALAT1-5 covered no. 4 site
presented a high ratio of the input compared to the adjacent
fragments (Sp1, F=144.9, P<0.0001; Sp3, F=68.54, P<0.0001).
Additionally, Sp3 showed stronger binding than Sp1, which may be
due to the expression disparity of endogenous Sp1 and Sp3 in HepG2
cells (Fig. 4A and B).
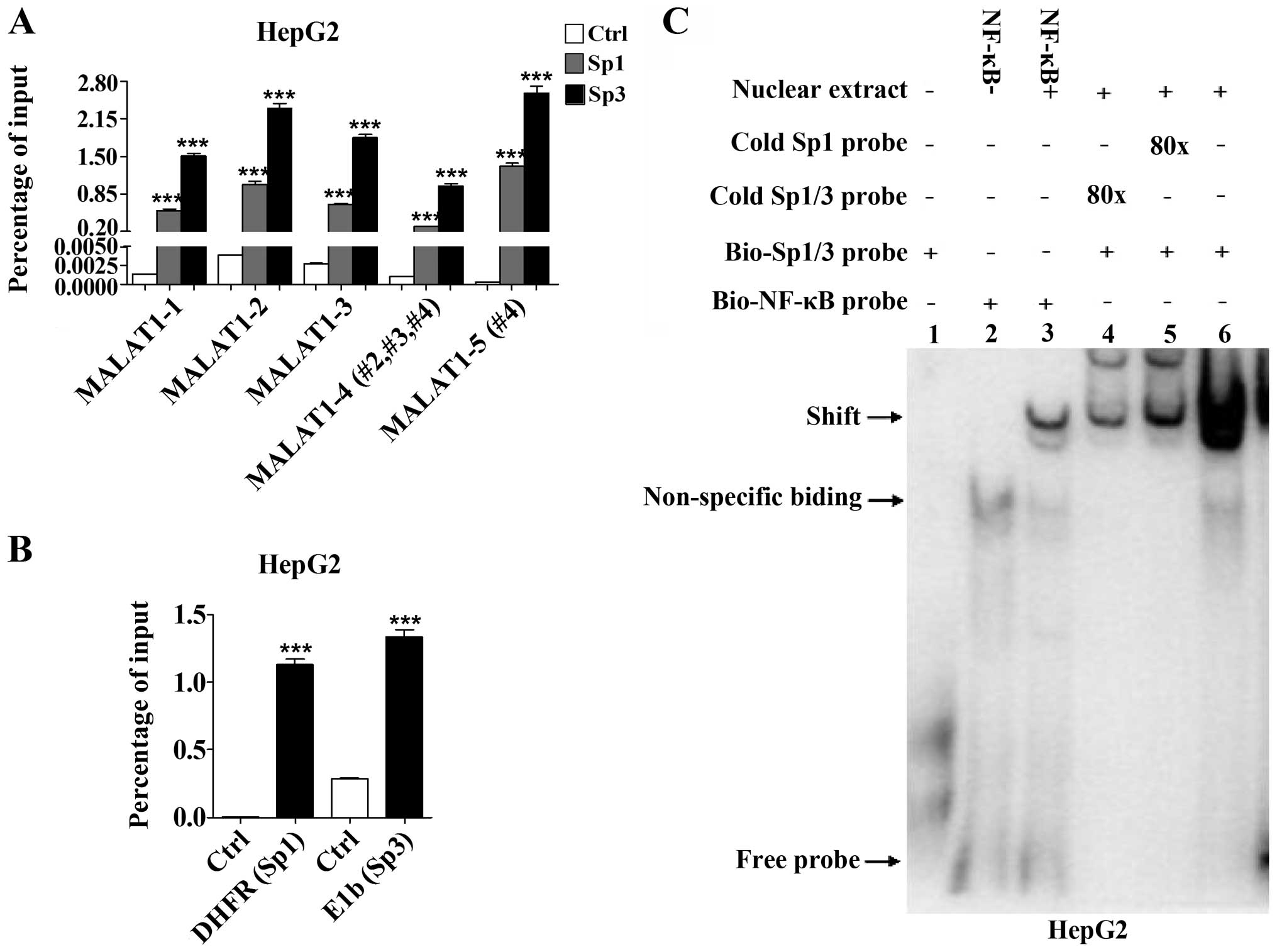 | Figure 4Detection of the binding activity
between Sp1/3 and MALAT1 in HepG2 cells. (A) ChIP assay
demonstrates the special binding activity between Sp1/3 and MALAT1.
Anti-Sp1 and anti-Sp3 successfully enriched five DNA sequence from
-1,000 to -1 bp of the MALAT1 promoter. The normal rabbit
IgG was used as a negative control. Pulled-down chromatin fragments
were then analyzed by RT-qPCR. Percentage of the input suggested
significant combination between transcription factors and the
MALAT1 promoter. The fragment from -85 to -1 bp of the
MALAT1 promoter, which covered predicted site no. 4,
presented the highest ratio of input (Sp1, F=144.9, P<0.0001;
Sp3, F=68.54, P<0.0001). *P<0.05,
**P<0.01, ***P<0.001. (B) The qPCR
primers included the Sp1 binding site in human DHFR promoter and
the Sp3 binding site in human E1b promoter. (C) Two special probes
were designed to determine DNA-binding activity using an
electrophoretic mobility shift assay. Two biotin-labeled probes
bound to the nuclear extract and were most competitively blocked by
an unlabeled one (shift). MALAT1, metastasis-associated lung
adenocarcinoma transcript 1; Sp, specificity protein. |
Furthermore, we performed EMSA and according to the
consensus binding sequence of Sp1 (5′-(G/T)GGGCGG(G/A)(G/A)(C/T)-3′
(22), we designed Sp1 probes with
or without biotin label, which also covers Sp3, according to a
previous study (23). The
biotin-labeled and unlabeled Sp1/3 oligonucleotides were prepared
for EMSA. The NF-κB probes with or without related nuclear extract
were used as a positive and negative control, respectively. The
data showed that 80 excess cold probes successfully blocked the Sp1
and Sp1/3 binding activities (Fig.
4C). Thus, our data indicated that the MALAT1 promoter sites
may not only bind to Sp1, but also to Sp3.
Sp1 and Sp3 enhance MALAT1
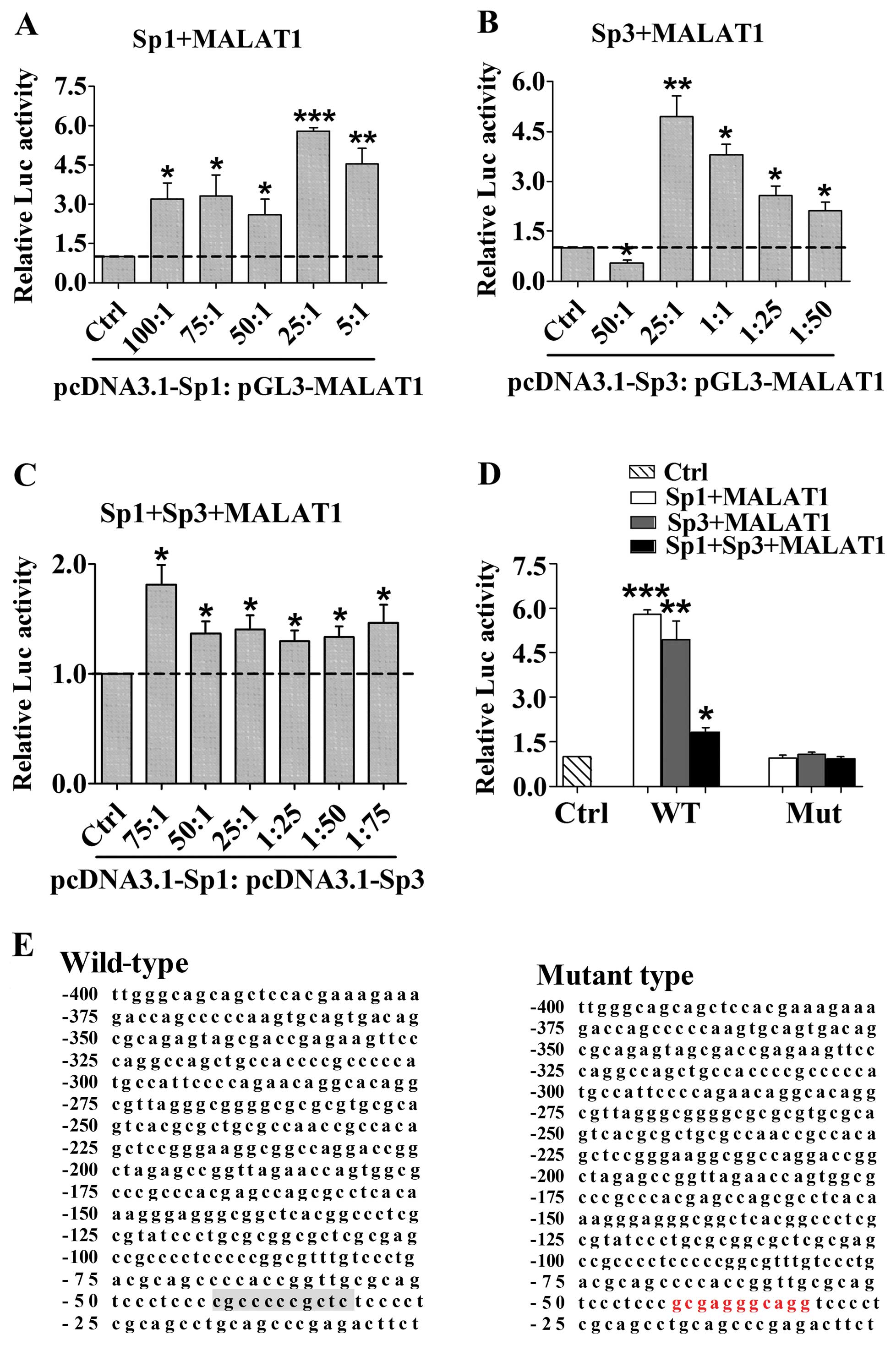
transcriptional activity
To confirm Sp1 and Sp3 MALAT1 transcriptional
activity, we designed and subcloned the MALAT1 -400 to -1 promoter
constructs with mutations in the potential binding sites according
to data from ChIPBase. We co-transfected the clones with Sp1 or Sp3
expression plasmid into 293T cells and measured the transcriptional
activity. As shown in Fig. 5A and
B, the ratio between Sp1/3 plasmid and promoter construct
indicated their positive role in regulating MALAT1 transcriptional
activity. Sp1 and Sp3 wild-type constructs coincidentally showed
the greatest activity in regulating MALAT1 transcription at a ratio
of 25:1 (Sp1, F=3.683, P=0.0172; Sp3, F=21.40, P<0.0001).
Separate transfection of Sp1 or Sp3 also enhanced MALAT1 promoter
activity (Fig. 5A and B), whereas a
combination of Sp1 and Sp3 showed much weaker action (Fig. 5C). Conversely, compared with the
wild-type, mutation at no. 4 significantly abolished Sp1 and Sp3
transcriptional activity (Fig. 5D and
E).
Discussion
MALAT1 is considered to be an oncogene as it is
frequently upregulated in various human cancers (15,24–27).
Previous findings demonstrated that an altered MALAT1 expression
was an independent predictor for postoperative HCC recurrence
(15). Our unpublished data also
showed that knockdown of MALAT1 expression inhibited HCC cell
proliferation and motility, and induced caspase-3/7 activity and
HCC cell apoptosis. MALAT1 may prevent HCC cell apoptosis via
inhibition of caspase-3/7 activity. However, the mechanism of
prediction of postoperative HCC recurrence remains unclear. Thus,
in the present study, we investigated the potential mechanism of
MALAT1 transcriptional regulation. We primarily assessed the levels
of MALAT1, Sp1 and Sp3 in HCC vs. non-tumor liver tissues and found
a significant upregulation of their levels in HCC tissues. We
performed a series of in vitro experiments to determine Sp1
and Sp3 regulation of MALAT1 expression in HCC cells. The present
data provided information that may lead to the therapeutic
targeting of Sp1 and Sp3 in MALAT1-ovexpressed HCCs.
MALAT1 is a large infrequently spliced non-coding
RNA that is highly conserved among mammals (28). MALAT1 regulates alternative splicing
by modulating SRSF1 phosphorylation to influence cancer-associated
aberrant splicing (14,29) and positively regulates cell motility
(30). Wang et al revealed a
novel mechanism underlying the balance between SRSF1,
Yes-associated protein (YAP) and MALAT1 and identified a new role
of YAP in upregulating MALAT1. SRSF1 inhibited YAP activity by
preventing its co-occupation with TCF/β-catenin on the MALAT1
promoter. By contrast, the overexpression of YAP interacted with
Angiomotin (AMOT) to impair the nuclear retention of SRSF1 and
itself. This effect resulted in removing the inhibitory role of
SRSF1 on MALAT1 in the nucleus (29). Recent findings have shown that
MALAT1 positively regulated latent transforming growth factor
β-binding protein-3 (LTBP3) transcription in mesenchymal stem cells
by recruiting Sp1 to the LTBP3 promoter. LTBP3 regulated the
bioavailability of tumor growth factor-β (TGF-β) (31). However, the mechanism of MALAT1
over-expression in human cancers remains to be elucidated (32).
Sp family members have been reported to be
associated with carcinogenesis (23,33).
In the present study, Sp1 and Sp3 were upregulated in HCC. Similar
results were obtained for pancreatic carcinoma (PC) (34). However, the factors that induced the
high expression of Sp1 or Sp3 remain to be determined. More studies
have concentrated on the target genes that are regulated by the two
transcriptional factors and influence carcinogenesis. Sp1, a
ubiquitous nuclear factor, was demonstrated to play a key role in
the growth and metastasis of many human cancers (21,23,34–36).
Previous microarray data showed that Sp1 was a potential regulator
of MALAT1 transcription (16–19).
In addition, Sp3 belongs to the family of Sp1-related genes that
encode transcription factors to regulate transcription of GC-and
GT-box containing genes (37).
Previous results showed that Sp1 was responsible for many
cancer-related genes including those associated with sustained
proliferation (hTERT/hTERC, p53/MDM2, p16, and p21), apoptosis
(survivin, Trail-R2, and Bcl-2), angiogenesis (VEGF, TSP-1, PDGF
and uPA), DNA damage/stress response (Brca1, ATM, and MDC1),
invasion and metastasis (MMP9, MT1-MMP, RECK, E-cadherin, Integrin
α5 and MMP2) (23,33). Similarly, Sp3 may bind to the same
sites of cancer-related genes such as Sp1 and have different
regulatory effects (23,33). For example, Sp1 and Sp3 physically
interacted and co-operated with GABP to activate the utrophin
promoter (37). Thus, although
Sp1/Sp3 is upregulated in cancers such as HCC and PC, if Sp1/Sp3
were inhibited the cancer-related genes transcriptions would be
affected. Thus, malignant behaviors, including proliferation,
apoptosis, angiogenesis, DNA damage/stress response, invasion and
metastasis, would be altered.
In the present study, we combined the two factors
and analyzed their transcriptional regulation of MALAT1. The levels
of Sp1, Sp3 and MALAT1 were markedly increased in the HCC tissues.
The expression of Sp1 and MALAT1 was associated with AFP level.
This result suggests that the co-analysis of serum AFP and MALAT1
levels may become a useful biomarker for HCC detection. In
addition, Sp1 or Sp3 expression was not obviously associated with
MALAT1 expression. Thus, MALAT1 may not be regulated by a single
factor. Subsequent data have verified that Sp1 and Sp3 were
involved in the transcriptional regulation of MALAT1. In addition,
Li et al (38) identified
that only Sp1 can independently upregulate the expression of MALAT1
in A549 cells by binding to two specific transcriptional binding
sites. Findings of the present study on HCC show that level of
MALAT1 only decreased with the silencing of both Sp1 and Sp3, and
not only of Sp1, which suggests a complementary effect between Sp1
and Sp3. The expression of MALAT1 is under the associated
regulation of Sp1 and Sp3. The Sp1 inhibitor, MIT, showed a similar
effect on MALAT1 inhibition. MIT, as a GC-rich region inhibitor,
can suppress protein biosynthesis via transcription inhibition
(39). It has been shown to inhibit
cancer growth by blocking the Sp-family transcription factors from
binding to GC-rich regions of gene regulatory elements and is used
for the treatment of leukemia and testicular cancer in the US
(21,40–45).
By contrast, MIT has also been shown to exert neuroprotective
effects in normal cells and potential as an agent for
neurodegenerative diseases (41).
Although MIT may produce some side effects, the data mentioned
above has revealed that MALAT1 expression was downregulated in
HepG2 cells treated with MIT. This result has demonstrated that
MALAT1 expression is associated with Sp.
Furthermore, MALAT1 promoter contained at
least five biding regions of Sp1/3, particularly in the region -85
to -4 bp. The present results confirmed the prediction of Sp1
binding sites in the MALAT1 promoter by using online
prediction tools. Additionally, using the luciferase assay it was
found that Sp1 contributed more to enhancing transcriptional
activity than Sp3 in HCC cells. Sp1 and Sp3 affected
transcriptional activity at a specific ratio, and alterations to
this ratio induced suppression of MALAT1 transcriptional
activation. However, additional studies are needed to investigate
the mechanism of Sp1 and Sp3 overexpression in HCC tissues. The
present study indicates that the specific Sp1 and Sp3 binding sites
may be developed as novel therapeutic targets for treating HCC.
In summary, we have demonstrated the role of Sp1 and
Sp3 in the regulation of MALAT1 expression in HCC cells. Future
studies are to investigate targeting Sp1/3 and MALAT1 as a
potential therapeutic strategy, including use of the Sp1 inhibitor
MIT, to control HCC progression.
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
lncRNAs
|
long non-coding RNAs
|
|
NEAT2
|
nuclear-enriched transcript 2
|
|
ncRNA
|
non-coding RNA
|
|
miRNA
|
micro-RNA
|
|
AFP
|
α-fetoprotein
|
|
MALAT1
|
metastasis-associated lung
adenocarcinoma transcript 1
|
|
Sp
|
specificity protein
|
|
SRSF1
|
serine/arginine-rich splicing factor
1
|
|
YAP
|
Yes-associated protein
|
|
AMOT
|
angiomotin
|
|
LTBP3
|
latent-transforming growth factor
β-binding protein-3
|
|
TGF-β
|
tumor growth factor-β
|
|
ChIP
|
chromatin immunoprecipitation
|
|
EMSA
|
electrophoretic mobility shift
assay
|
|
MIT
|
mithramycin A
|
Acknowledgments
This study was supported in part by grants from the
Guangxi Provincial Science and Technology Development Projects
(nos. 1298003-2-5 and 10124001A-1), the Guangxi Natural Scientific
Research (no. 2014GXNSFBA118167) and the Guangxi Provincial Talent
Projects for Graduate Students (no. YCSZ2014099).
References
|
1
|
Kapranov P, Willingham AT and Gingeras TR:
Genome-wide transcription and the implications for genomic
organization. Nat Rev Genet. 8:413–423. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lau E: Non-coding RNA: Zooming in on
lncRNA functions. Nat Rev Genet. 15:574–575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ponjavic J, Ponting CP and Lunter G:
Functionality or transcriptional noise? Evidence for selection
within long noncoding RNAs. Genome Res. 17:556–565. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang G, Lu X and Yuan L: LncRNA: A link
between RNA and cancer. Biochim Biophys Acta. 1839:1097–1109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang L, Yang F, Yuan JH, Yuan SX, Zhou
WP, Huo XS, Xu D, Bi HS, Wang F and Sun SH: Epigenetic activation
of the MiR-200 family contributes to H19-mediated metastasis
suppression in hepatocellular carcinoma. Carcinogenesis.
34:577–586. 2013. View Article : Google Scholar
|
|
7
|
Huang JF, Guo YJ, Zhao CX, Yuan SX, Wang
Y, Tang GN, Zhou WP and Sun SH: Hepatitis B virus X protein
(HBx)-related long noncoding RNA (lncRNA) down-regulated expression
by HBx (Dreh) inhibits hepatocellular carcinoma metastasis by
targeting the intermediate filament protein vimentin. Hepatology.
57:1882–1892. 2013. View Article : Google Scholar
|
|
8
|
Du Y, Kong G, You X, Zhang S, Zhang T, Gao
Y, Ye L and Zhang X: Elevation of highly up-regulated in liver
cancer (HULC) by hepatitis B virus X protein promotes hepatoma cell
proliferation via down-regulating p18. J Biol Chem.
287:26302–26311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Z, Zhou L, Wu LM, Lai MC, Xie HY,
Zhang F and Zheng SS: Overexpression of long non-coding RNA HOTAIR
predicts tumor recurrence in hepatocellular carcinoma patients
following liver transplantation. Ann Surg Oncol. 18:1243–1250.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takahashi K, Yan I, Haga H and Patel T:
Long noncoding RNA in liver diseases. Hepatology. 60:744–753. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mohamadkhani A: Long Noncoding RNAs in
interaction with RNA binding proteins in hepatocellular carcinoma.
Hepat Mon. 14:e187942014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ji Q, Zhang L, Liu X, Zhou L, Wang W, Han
Z, Sui H, Tang Y, Wang Y, Liu N, et al: Long non-coding RNA MALAT1
promotes tumour growth and metastasis in colorectal cancer through
binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2
complex. Br J Cancer. 111:736–748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong Y, Liang G, Yuan B, Yang C, Gao R and
Zhou X: MALAT1 promotes the proliferation and metastasis of
osteosarcoma cells by activating the PI3K/Akt pathway. Tumour Biol.
36:1477–1486. 2015. View Article : Google Scholar
|
|
14
|
Tripathi V, Ellis JD, Shen Z, Song DY, Pan
Q, Watt AT, Freier SM, Bennett CF, Sharma A, Bubulya PA, et al: The
nuclear-retained noncoding RNA MALAT1 regulates alternative
splicing by modulating SR splicing factor phosphorylation. Mol
Cell. 39:925–938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lai MC, Yang Z, Zhou L, Zhu QQ, Xie HY,
Zhang F, Wu LM, Chen LM and Zheng SS: Long non-coding RNA MALAT-1
over-expression predicts tumor recurrence of hepatocellular
carcinoma after liver transplantation. Med Oncol. 29:1810–1816.
2012. View Article : Google Scholar
|
|
16
|
Yang JH, Li JH, Jiang S, Zhou H and Qu LH:
ChIPBase: A database for decoding the transcriptional regulation of
long non-coding RNA and microRNA genes from ChIP-Seq data. Nucleic
Acids Res. 41(D1): D177–D187. 2013. View Article : Google Scholar :
|
|
17
|
Wu Z, Liu X, Liu L, Deng H, Zhang J, Xu Q,
Cen B and Ji A: Regulation of lncRNA expression. Cell Mol Biol
Lett. 19:561–575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uesaka M, Nishimura O, Go Y, Nakashima K,
Agata K and Imamura T: Bidirectional promoters are the major source
of gene activation-associated non-coding RNAs in mammals. BMC
Genomics. 15:352014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alam T, Medvedeva YA, Jia H, Brown JB,
Lipovich L and Bajic VB: Promoter analysis reveals globally
differential regulation of human long non-coding RNA and
protein-coding genes. PLoS One. 9:e1094432014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen G, Kronenberger P, Teugels E, Umelo
IA and De Grève J: Targeting the epidermal growth factor receptor
in non-small cell lung cancer cells: The effect of combining RNA
interference with tyrosine kinase inhibitors or cetuximab. BMC Med.
10:282012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yuan P, Wang L, Wei D, Zhang J, Jia Z, Li
Q, Le X, Wang H, Yao J and Xie K: Therapeutic inhibition of Sp1
expression in growing tumors by mithramycin a correlates directly
with potent antiangiogenic effects on human pancreatic cancer.
Cancer. 110:2682–2690. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Narayan VA, Kriwacki RW and Caradonna JP:
Structures of zinc finger domains from transcription factor Sp1.
Insights into sequence-specific protein-DNA recognition. J Biol
Chem. 272:7801–7809. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li L and Davie JR: The role of Sp1 and Sp3
in normal and cancer cell biology. Ann Anat. 192:275–283. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ren S, Wang F, Shen J, Sun Y, Xu W, Lu J,
Wei M, Xu C, Wu C, Zhang Z, et al: Long non-coding RNA metastasis
associated in lung adenocarcinoma transcript 1 derived miniRNA as a
novel plasma-based biomarker for diagnosing prostate cancer. Eur J
Cancer. 49:2949–2959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gutschner T, Hämmerle M, Eissmann M, Hsu
J, Kim Y, Hung G, Revenko A, Arun G, Stentrup M, Gross M, et al:
The noncoding RNA MALAT1 is a critical regulator of the metastasis
phenotype of lung cancer cells. Cancer Res. 73:1180–1189. 2013.
View Article : Google Scholar :
|
|
26
|
Gutschner T, Hämmerle M and Diederichs S:
MALAT1 - a paradigm for long noncoding RNA function in cancer. J
Mol Med Berl. 91:791–801. 2013. View Article : Google Scholar
|
|
27
|
Ji P, Diederichs S, Wang W, Böing S,
Metzger R, Schneider PM, Tidow N, Brandt B, Buerger H, Bulk E, et
al: MALAT-1, a novel noncoding RNA, and thymosin beta4 predict
metastasis and survival in early-stage non-small cell lung cancer.
Oncogene. 22:8031–8041. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hutchinson JN, Ensminger AW, Clemson CM,
Lynch CR, Lawrence JB and Chess A: A screen for nuclear transcripts
identifies two linked noncoding RNAs associated with SC35 splicing
domains. BMC Genomics. 8:392007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang J, Wang H, Zhang Y, Zhen N, Zhang L,
Qiao Y, Weng W, Liu X, Ma L, Xiao W, et al: Mutual inhibition
between YAP and SRSF1 maintains long non-coding RNA, Malat1-induced
tumourigenesis in liver cancer. Cell Signal. 26:1048–1059. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tano K, Mizuno R, Okada T, Rakwal R,
Shibato J, Masuo Y, Ijiri K and Akimitsu N: MALAT-1 enhances cell
motility of lung adenocarcinoma cells by influencing the expression
of motility-related genes. FEBS Lett. 584:4575–4580. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li B, Chen P, Qu J, Shi L and Zhuang W, Fu
J, Li J, Zhang X, Sun Y and Zhuang W: Activation of LTBP3 gene by a
long noncoding RNA (lncRNA) MALAT1 transcript in mesenchymal stem
cells from multiple myeloma. J Biol Chem. 289:29365–29375. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bonasio R and Shiekhattar R: Regulation of
transcription by long noncoding RNAs. Annu Rev Genet. 48:433–455.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Beishline K and Azizkhan-Clifford J: Sp1
and the 'hallmarks of cancer'. FEBS J. 282:224–258. 2015.
View Article : Google Scholar
|
|
34
|
Sankpal UT, Maliakal P, Bose D, Kayaleh O,
Buchholz D and Basha R: Expression of specificity protein
transcription factors in pancreatic cancer and their association in
prognosis and therapy. Curr Med Chem. 19:3779–3786. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tong Y, Tan Y, Zhou C and Melmed S:
Pituitary tumor transforming gene interacts with Sp1 to modulate
G1/S cell phase transition. Oncogene. 26:5596–5605. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang NY, Woda BA, Banner BF, Whalen GF,
Dresser KA and Lu D: Sp1, a new biomarker that identifies a subset
of aggressive pancreatic ductal adenocarcinoma. Cancer Epidemiol
Biomarkers Prev. 17:1648–1652. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Galvagni F, Capo S and Oliviero S: Sp1 and
Sp3 physically interact and co-operate with GABP for the activation
of the utrophin promoter. J Mol Biol. 306:985–996. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li S, Wang Q, Qiang Q, Shan H, Shi M, Chen
B, Zhao S and Yuan L: Sp1-mediated transcriptional regulation of
MALAT1 plays a critical role in tumor. J Cancer Res Clin Oncol.
16–March;2015.(Epub ahead of print). View Article : Google Scholar
|
|
39
|
Mir MA, Majee S, Das S and Dasgupta D:
Association of chromatin with anticancer antibiotics, mithramycin
and chromomycin A3. Bioorg Med Chem. 11:2791–2801. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Choi ES, Nam JS, Jung JY, Cho NP and Cho
SD: Modulation of specificity protein 1 by mithramycin A as a novel
therapeutic strategy for cervical cancer. Sci Rep. 4:71622014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Osada N, Kosuge Y, Ishige K and Ito Y:
Mithramycin, an agent for developing new therapeutic drugs for
neurodegenerative diseases. J Pharmacol Sci. 122:251–256. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kajita Y, Kato T Jr, Tamaki S, Furu M,
Takahashi R, Nagayama S, Aoyama T, Nishiyama H, Nakamura E,
Katagiri T, et al: The transcription factor Sp3 regulates the
expression of a metastasis-related marker of sarcoma, actin
filament-associated protein 1-like 1 (AFAP1L1). PLoS One.
8:e497092013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huong PT, Soung NK, Jang JH, Cha-Molstad
HJ, Sakchaisri K, Kim SO, Jang JM, Kim KE, Lee KS, Kwon YT, et al:
Regulation of CEP131 gene expression by SP1. Gene. 513:75–81. 2013.
View Article : Google Scholar
|
|
44
|
Zhang M, Mathur A, Zhang Y, Xi S, Atay S,
Hong JA, Datrice N, Upham T, Kemp CD, Ripley RT, et al: Mithramycin
represses basal and cigarette smoke-induced expression of ABCG2 and
inhibits stem cell signaling in lung and esophageal cancer cells.
Cancer Res. 72:4178–4192. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hsu TI, Wang MC, Chen SY, Huang ST, Yeh
YM, Su WC, Chang WC and Hung JJ: Betulinic acid decreases
specificity protein 1 (Sp1) level via increasing the sumoylation of
sp1 to inhibit lung cancer growth. Mol Pharmacol. 82:1115–1128.
2012. View Article : Google Scholar : PubMed/NCBI
|