Introduction
Squamous cell carcinoma of the head and neck
(SCCHN), a malignant tumor of epithelial origin, accounts for more
than 90% of all head and neck cancers, with a 5-year survival rate
of only ~30–40% (1). The main
reasons for the poor diagnosis of this disease are tumor recurrence
and metastasis to lymph nodes. Therefore, a better understanding of
the processes involved in its recurrence and metastasis is
necessary to enable the development of therapies designed to
prevent tumor dissemination.
Chemokines are a group of small structurally related
molecules that constitute a superfamily of inducible secreted
proinflammatory proteins that are involved in a variety of immune
responses (2–5). In has been reported that the CC
chemokine CCL19 and its receptor chemokine receptor 7 (CCR7)
participate in the metastasis of various types of tumors, such as
uveal melanoma, pancreatic ductal adenocarcinoma and gastric
cancer, in a manner such as 'lymphocyte homing' (6–8). Our
previous studies demonstrated that CCR7 can regulate cell migration
and invasion of SCCHN, and this process involves several signaling
pathways including PI3K/cdc42, MAPK and JAK2/STAT3 (9–11).
In a previous study, we also found that praline-rich
tyrosine kinase-2 (Pyk2) is activated by CCL19-CCR7 interaction in
SCCHN cells, and the Pyk2 inhibitor inhibits tumor cell mobility
(12). The results suggest that
Pyk2 may be involved in the CCR7-mediated regulation of SCCHN cell
biological activity. The aim of the present study was to further
demonstrate whether Pyk2 participates in the CCR7 downstream
signaling network, and to determine whether this molecule plays a
role in the CCR7-mediated regulation of SCCHN viability and
metastasis in vivo and in vivo and the involved
mechanisms.
Materials and methods
Cell lines
PCI-4B and PCI-37B, well-characterized SCCHN cell
lines derived from the metastatic lymph node of SCCHN patients,
were kindly donated by the University of Pittsburgh Cancer
Institute (13,14). The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA, USA)
containing 10% fetal bovine serum (FBS) (Gibco, Carlsbad, CA, USA),
100 U/ml penicillin G and 100 U/ml streptomycin.
Reagents and antibodies
CCL19- and CCR7-specific monoclonal antibodies
(mouse anti-human CCR7 antibody) were purchased from R&D
Systems (Minneapolis, MN, USA). Anti-Pyk2 and anti-phospho-Pyk2
were purchased from Santa Cruz Biotechnology (Dallas, TX, USA).
Anti-vimentin and anti-E-cadherin were purchased from Cell
Signaling Technology (Danvers, MA, USA). The treatment conditions,
such as concentration and time, were drawn from
pre-experiments.
CCR7 siRNA and PRNK transfection
The Pyk2 related non-kinase (PRNK) is a recently
described isoform of Pyk2 that encodes part of the COOH-terminal
domain of Pyk2 and has been proposed to selectively downregulate
Pyk2 function (15). Lentiviruses
containing the PRNK gene (LV-PRNK-EGFP) or a negative control
(Lenti-EGFP) were obtained from Shanghai GeneChem, China. The
construction of PRNK recombinant lentiviral vectors was as follows.
The GV218 vector (Lenti-EGFP; purchased from Shanghai GeneChem) was
linearized with BamHI and AgeI. The PRNK gene cDNA
sequence was as described by Xiong et al (15), and synthesized and purchased from
Shanghai GeneChem. The cDNA was also digested with BamHI and
AgeI, and ligated into the GV218 vector. The constructs were
verified by sequencing. The Lenti-EGFP was lentiviruses ligated
with EGFP by T4 DNA ligase, 5′ exonuclease and Taq
polymerase. Viruses were propagated in 293 cells, clonally isolated
and titered. SCCHN cells were infected at a matched multiplicity of
infection (MOI) of 10. CCR7 siRNA was purchased from Santa Cruz
Biotechnology. Transfection of the SCCHN cells with 20 nM of siRNA
was carried out using Lipofectamine SiRNAMAX (Invitrogen) for 32
h.
Western blot analysis
Cells were gathered and lysed. Lysates were
sonicated for 3 sec and centrifuged at 4°C and 14,000 rpm for 30
min. The supernatant was collected for protein quantification using
the Bio-Rad protein assay dye reagent (Bio-Rad Laboratories,
Richmond, CA, USA). Fifty micrograms of protein were
size-fractionated through a 10% SDS-PAGE gel and transferred onto
nitrocellulose filters. The filters were blocked [5% non-fat dry
milk, 0.1% Triton X-100, 150 mM NaCl, 50 mM Tris
[tris(hydroxymethyl)aminomethane] (pH 7.5)] and incubated with the
primary antibody, which was diluted to a ratio of 1:1,000.
Nitrocellulose filters were incubated with horseradish
peroxidase-conjugated secondary antibodies. Bands were visualized
using the enhanced chemiluminescence system (Amersham Pharmacia
Biotech, Piscataway, NJ, USA) and quantified by scanning
densitometry using FluorChem v2.0 software (Alpha Innotech
Corporation, San Leandro, CA, USA).
Migration assay
Disposable 24-well Transwell inserts with a
80-μm pore size were run in triplicate in DMEM with 0.5%
(w/v) BSA. CCL19 was added to the lower chamber at a concentration
of 500 ng/ml. Cell suspensions (2x105) were placed in
the top of the inserts. After 24 h of incubation, the cells on the
upper surface of the inserts were removed with a cell harvester,
and the membrane was washed with medium. Cells that penetrated the
membrane were fixed with ice-cold methanol, stained with 0.5%
crystal violet, photographed and counted under a microscope.
Migration index was calculated based on the control involving
random migration.
Matrigel invasion assay
Cell invasion was quantified in vitro using
Matrigel-coated semi-permeable, modified inserts with a pore size
of 8-μm. The analysis of the Matrigel invasion assay was
performed as described in the migration assay incubated with CCL19
for 36 h. Invasion index was calculated based on the control
involving random invasion.
Viability assay
The viability assay is a colorimeter assay that
relies on the ability of viable cells to convert a soluble
tetrazolium salt, MTT, into a quantifiable insoluble formazan
precipitate (yellow to purple color change). Cell death correlates
with reduced purple precipitate formation. In brief, SCCHN cells
were plated into a 96-well plate at an initial density of
5×104 cells/well. Confluent cells (70–80%) were serum
starved for 24 h, and then treated with CCL19. Media were
discarded, and 20 μl of MTT (5 μg/ml in PBS) was
added for 4 h. DMSO (200 μl) was then added to each well,
and the plate was incubated at 37°C in a 5% CO2
atmosphere for 1 h to lyse all of the cells, and the media were
collected for measurement. The optical density (OD) was read by a
spectrometer (Tecan, Männedorf, Switzerland) at 490 nm. The
percentage of cell viability was calculated as follows: Viability
percentage (%) = (OD of treatment group)/(OD of control group) ×
100%.
Adhesion assay
For the assay, 96-well plates (Corning Costar,
Cambridge, MA, USA) were coated with fibro-nectin or left uncoated,
and incubated at room temperature overnight. SCCHN cells were
plated in an initial density of 1×104 cells/well, and
inbubated at 37°C for 1.5 h, and then washed with
phosphate-buffered saline (PBS) three times. MTT (20 μl, 5
μg/ml) was added for 4 h, and then dimethyl-sulphoxide
(DMSO; Sigma, USA) was added to each well to lyse all cells for 1 h
at 37°C at 5% CO2. The OD was read on a spectrometer
(Tecan) through a 490-nm filter. For each experimental condition 8
wells were analyzed in parallel. Adhesion index (%) = [(fibronectin
conditions cell OD/control conditions cell OD) − 1] × 100%.
Apoptosis assay
To determine the role of the Pyk2 pathway in
apoptosis, Annexin V and propidium iodide (PI) staining was
performed. Briefly, the transfected cells were pretreated and
harvested at the indicated time points. Cells were stained with
fluorescein isothiocyanate (FITC)-conjugated Annexin V and PI
according to the manufacturer's recommendation. In these
experiments, the cells that were apoptotic were those that were
Annexin V-positive/PI-negative. Samples were analyzed using a
FACSCalibur flow cytometer and CellQuest software (BD, Sparks, MD,
USA).
Xenograft studies of nude mice
BALB/c athymic nude mice (4 weeks of age) were
maintained under standard laboratory conditions on a 12-h
light-dark cycle and given access to sterilized food and water
ad libitum under a specific pathogen-free environment. For
the subcutaneous injection, the cells (1×106) were
suspended in 200 μl PBS and then inoculated subcutaneously
into the right breast region of the BALB/c nude mice. Each group
consisted of six nude mice. Two-dimensional measurements were taken
with an electronic caliper, and the tumor volume in mm3
was calculated using the formula: Volume = a × b2 ×
0.52, where a is the longest diameter; b is the shortest diameter.
At the terminal point when the mice were sacrificed, the tumors
were fixed in 10% formalin overnight and subjected to routine
histological examination. All of the animals were treated according
to the protocols approved by the Institutional Animal Care and Use
Committee of China Medical University.
Statistical analysis
Data are expressed as the mean ± standard deviation
(SD) of repeated assays. Significant differences between the two
groups were evaluated using an unpaired Student's t-test. A p-value
<0.05 was considered to indicate a statistically significant
result. All statistical analyses were performed with software SPSS
11.0 (SPSS, Inc., Chicago, IL, USA).
Results
PRNK transfection and its role in
Pyk2
PRNK is the C-terminal non-kinase region of Pyk2. It
is characterized as a splice variant form of Pyk2. PRNK will
interact with several of the Pyk2-binding partners due to sequence
similarity with Pyk2. Therefore, it has been reported as a
dominant-negative regulator to prevent Pyk2 binding with its
partners for signaling pathway activation (15). In the present study, we constructed
the lentivirus shuttle vector merged with EGFP for PRNK
transfection, and examined the EGFP expression which represented
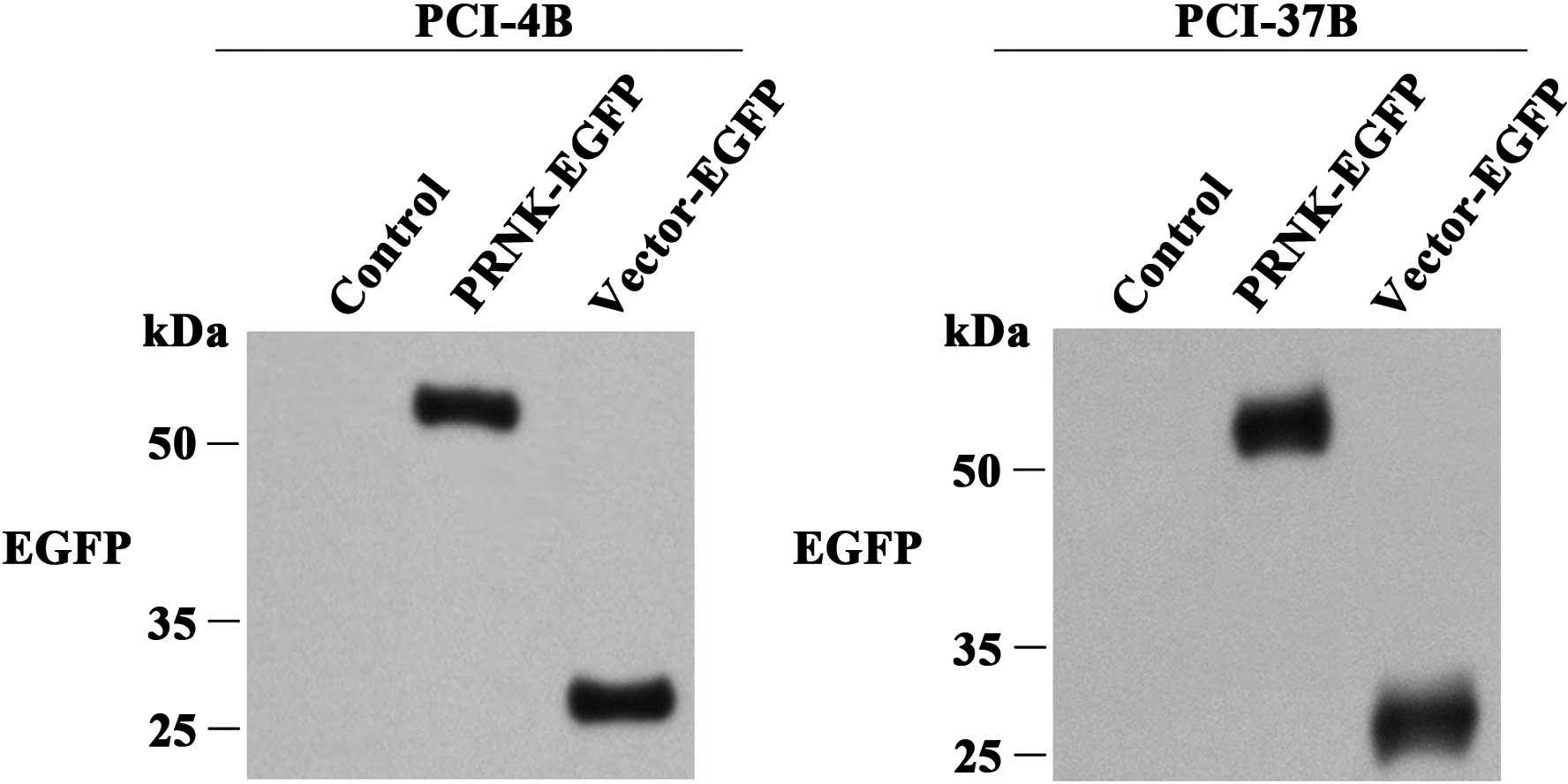
the efficacy of the PRNK transfection. As shown in Fig. 1, in the control group, EGFP
expression was not observed. In the LV (vector)-EGFP control group,
EGFP expression was observed at 27 kDa, indicating that the
transfection was effective. In the LV-PRNK-EGFP group, EGFP
expression was also observed, but it was located at ~52–54 kDa,
suggesting it is a compound of PRNK and EGFP.
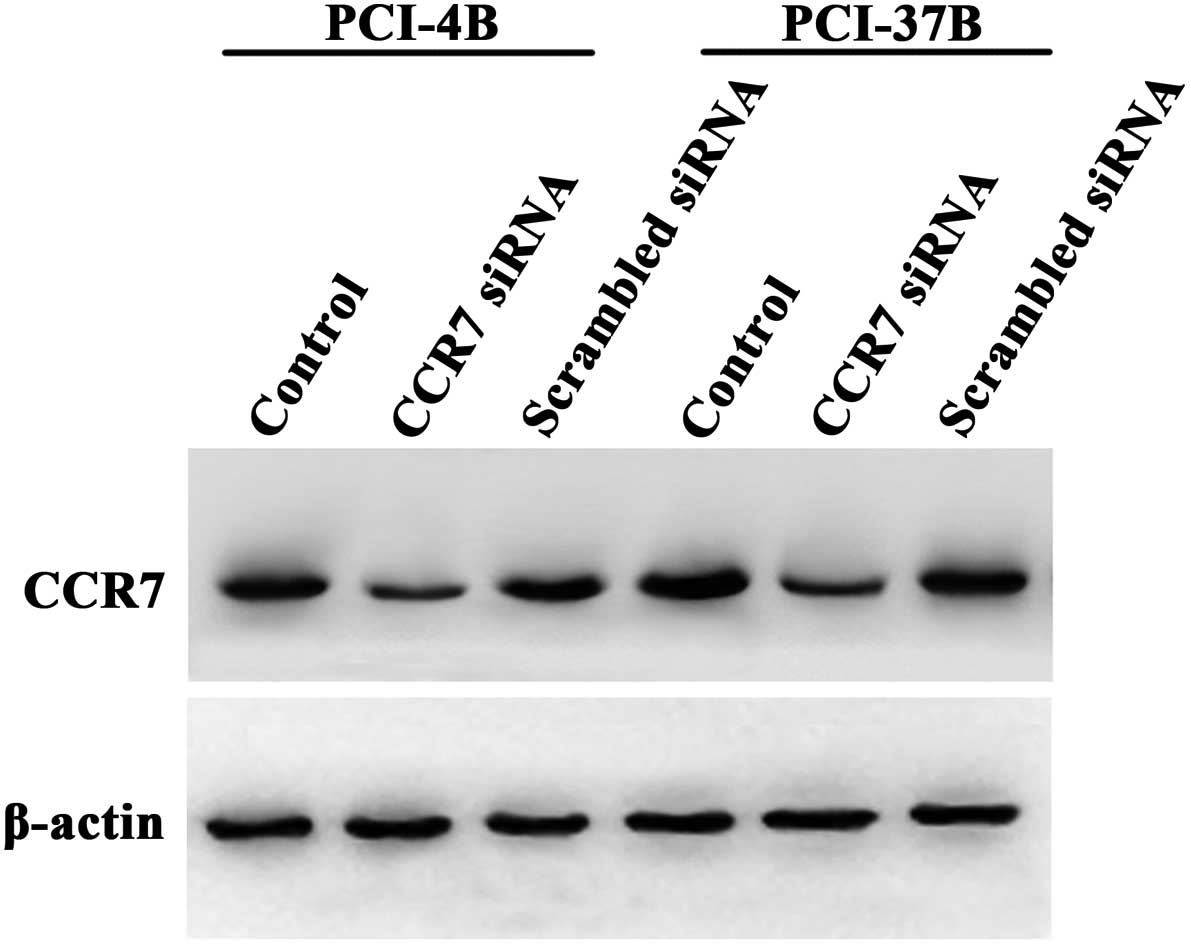
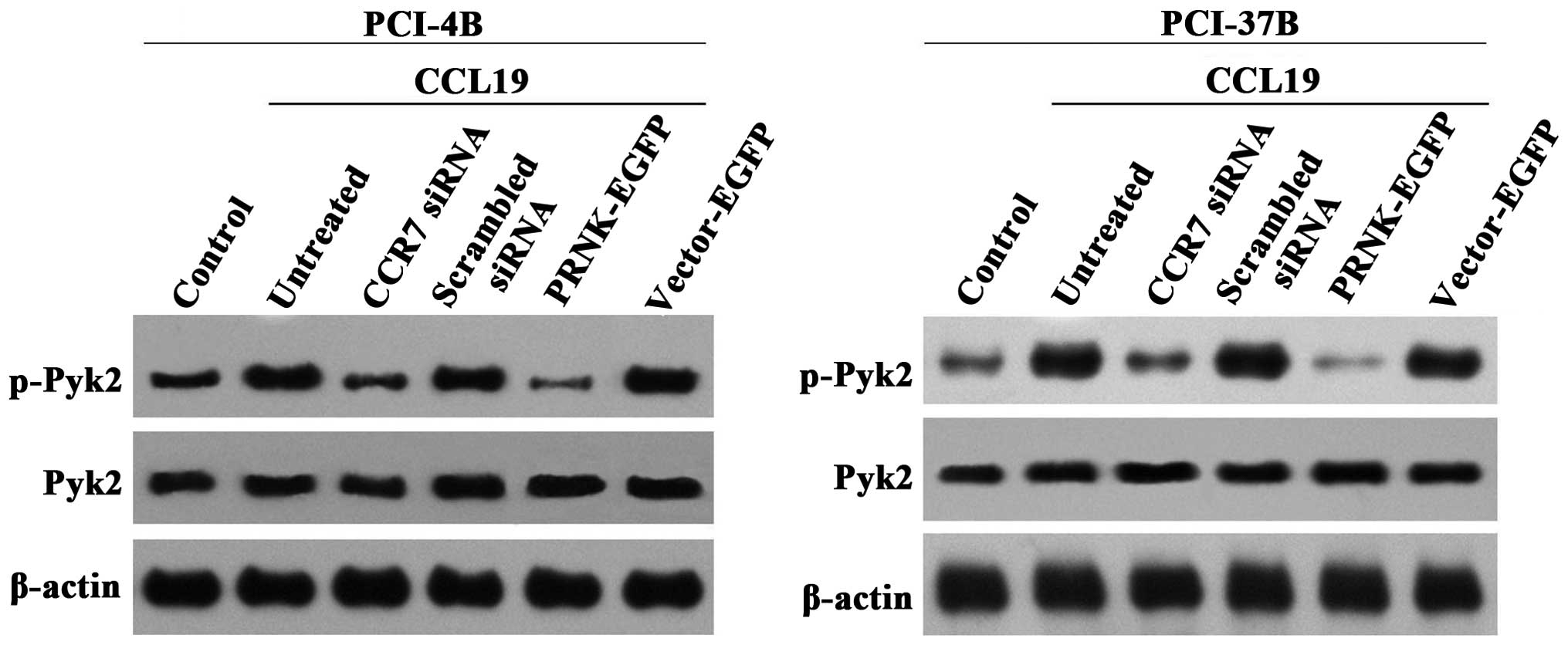
In a previous study, we observed that the
phosphorylation of Pyk2 was inhibited by the CCR7 monoclonal
antibody (12). In the present
study, we transiently transfected CCR7 siRNA into SCCHN cells to
silence CCR7 (Fig. 2) (16), and used CCL19 to induce the CCR7
downstream signaling pathway. The results showed that CCL19 induced
the phosphorylation of Pyk2, and CCR7 siRNA significantly reduced
this phosphorylation, which had a significant difference compare to
the CCL19 and scramble siRNA groups (Fig. 3). These results demonstrated that
Pyk2 is a downstream signaling molecule of CCR7. In the LV-EGFP
group, the Pyk2 phosphorylation was similar to the CCL19 group, and
in the LV-PRNK-EGFP group, the Pyk2 phosphorylation was
significantly reduced. In contrast, the total Pyk2 protein
expression had no change in these groups. The results demonstrated
that PRNK is a competitive inhibitor of Pyk2 constitutional
activation, which will not interfere with Pyk2 expression.
Pyk2 regulates CCL19-induced SCCHN cell
viability, migration, invasion and adhesion
Our previous study used cisplatin to induce cell
death and demonstrated that CCL19 can promote PCI-4B and PCI-37B
cell viability, migration and invasion, and this function can be
blocked by CCR7 monoclonal antibody (10,11,17–20).
In the present study, we examined whether Pyk2 is involved in
regulating these bioactivities.
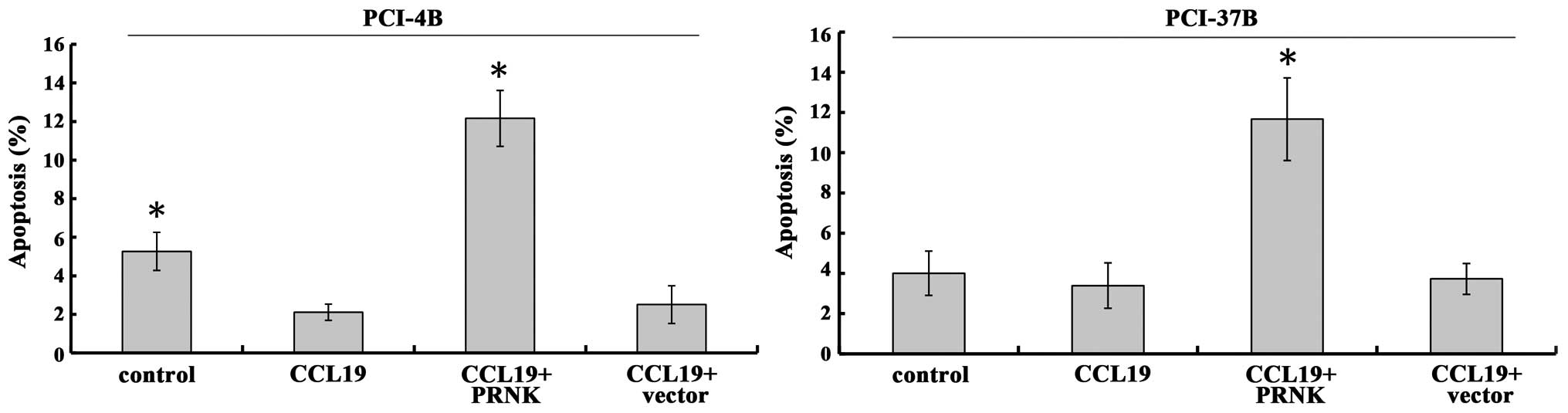
In the PCI-4B and PCI-37B cell lines, treatment of
CCL19 protected the cells from apoptosis (Fig. 4) (19). Yet, in the PRNK-transfected PCI-4B
and PCI-37B cells, the role of CCL19 was blocked, and the apoptosis
was significantly increased ~2–3 times in the PCI-4B and PCI-37B
cell lines. As a control, the vector transfection group had no
significant change. This indicates that PRNK promotes SCCHN cell
apoptosis.
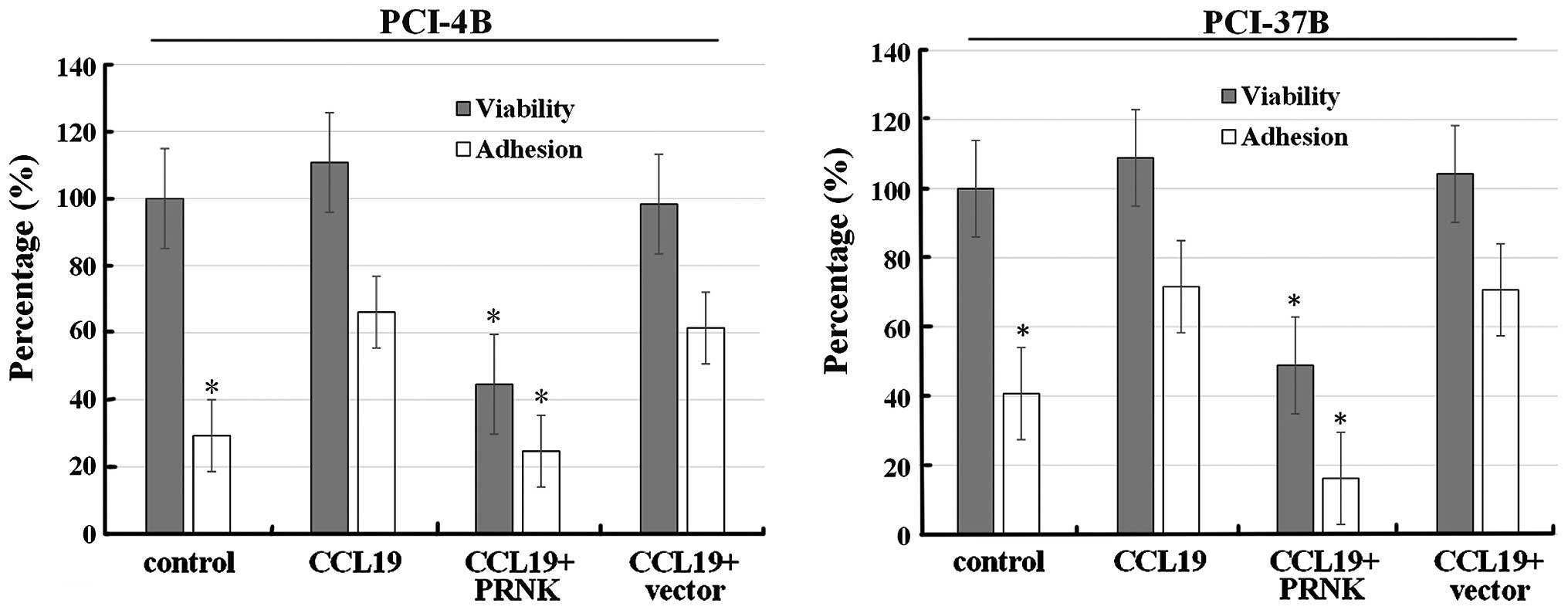
As shown in Fig. 5,
following CCL19 treatment, the viability of the PCI-4B and the
PCI-37B cells was unaffected (~100–110%), while the viability of
the PRNK-transfected PCI-4B and PCI-37B cells was reduced to less
than 50%; the difference was significant. As a control, the vector
transfection group had no significant change. The results indicated
that PRNK can induce cell death.
We also examined the adhesive ability of the SCCHN
cells (Fig. 5). Following CCL19
pretreatment, the adhesion index of the PCI-4B and the PCI-37B
cells increased from 30–40% of control condition to ~70%,
suggesting that CCL19 promotes SCCHN cell adhesion to fibronectin.
In the stable PRNK-expressing PCI-4B and PCI-37B cells, CCL19
played no role in this adhesion induction, and the adhesion index
decreased to the control level. This indicates that the interaction
between CCL19 and CCR7-induced adhesion was Pyk2-dependent.
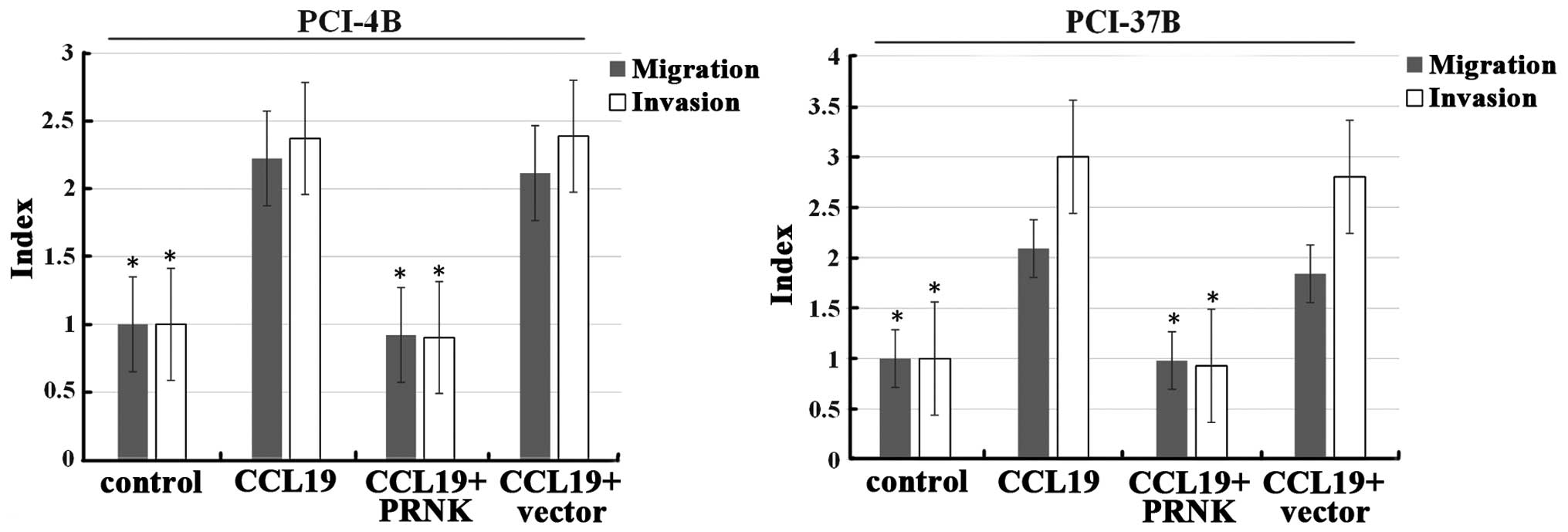
A migration assay was next carried out. Following
CCL19 induction, the cell migration index of the PCI-4B and PCI-37B
cells was >2 times that of the control group (Fig. 6). PRNK transfection significantly
blocked the effect of CCL19 in both cell lines, leading to
decreased cell migration to almost baseline level. The invasion
index was the same as the migration index, and CCL19-induced
invasion was significantly blocked by PRNK transfection. As a
control, the vector transfection group had no significant change in
migration and invasion.
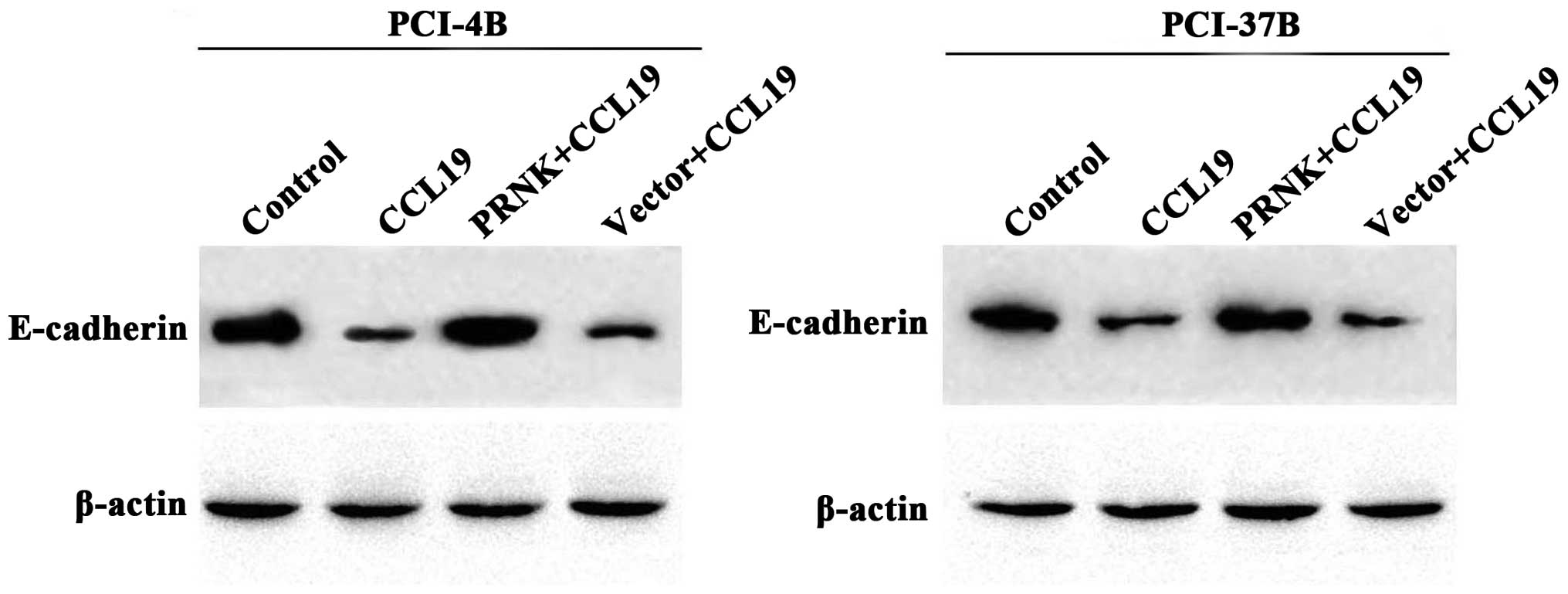
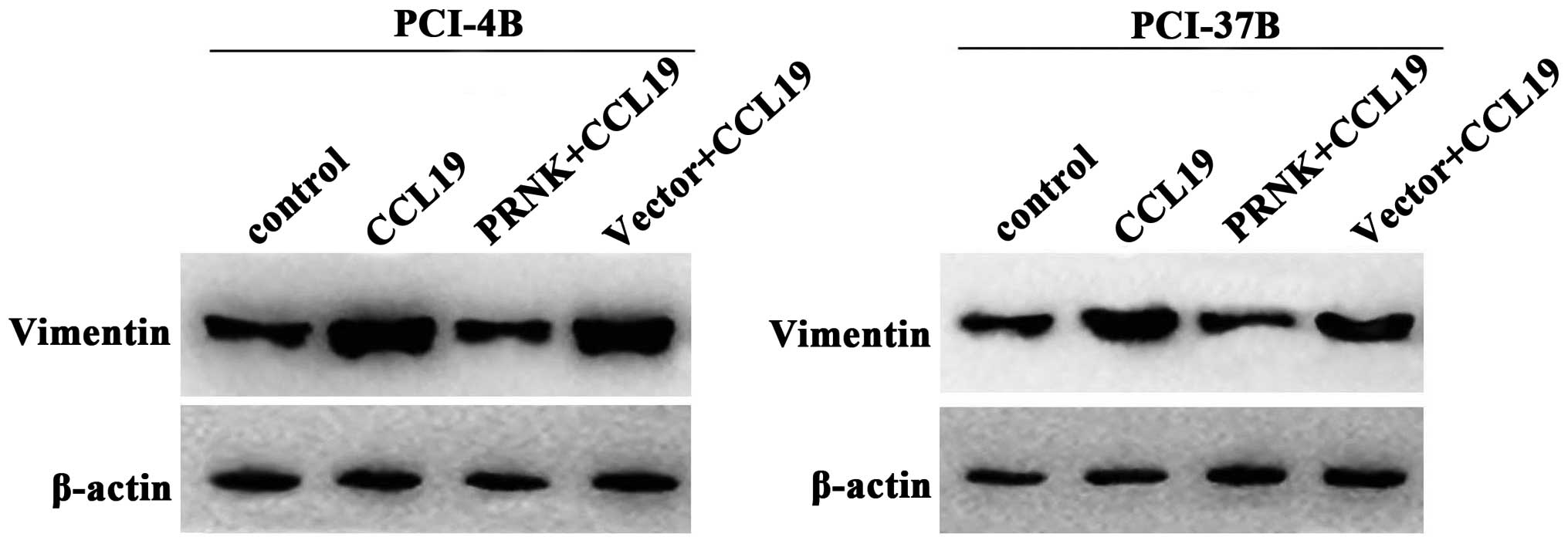
Pyk2 mediates the expression of
E-cadherin and vimentin induced by CCR7
E-cadherin and vimentin are known to be associated
with epithelial-mesenchymal transition (EMT), which is a key point
in tumor progression and generally participates in cell migration
and invasion. Our previous results demonstrated that CCL19-treated
PCI-4B and PCI-37B cells exhibited a significant increase in the
level of vimentin protein and a significant decrease in the level
of E-cadherin (10). In the present
study, we examined the PRNK-transfected PCI-4B and PCI-37B cells,
and found that, with the Pyk2 inaction the CCL19-induced low
E-cadherin expression was significantly increased, and the
CCL19-induced high vimentin expression was also significantly
decreased (Figs. 7 and 8). The results supported that the
CCR7-mediated E-cadherin and vimentin expression levels in the
SCCHN cells were Pyk2-dependent.
Pyk2 inactivation inhibits SCCHN
tumorigenesis in vivo
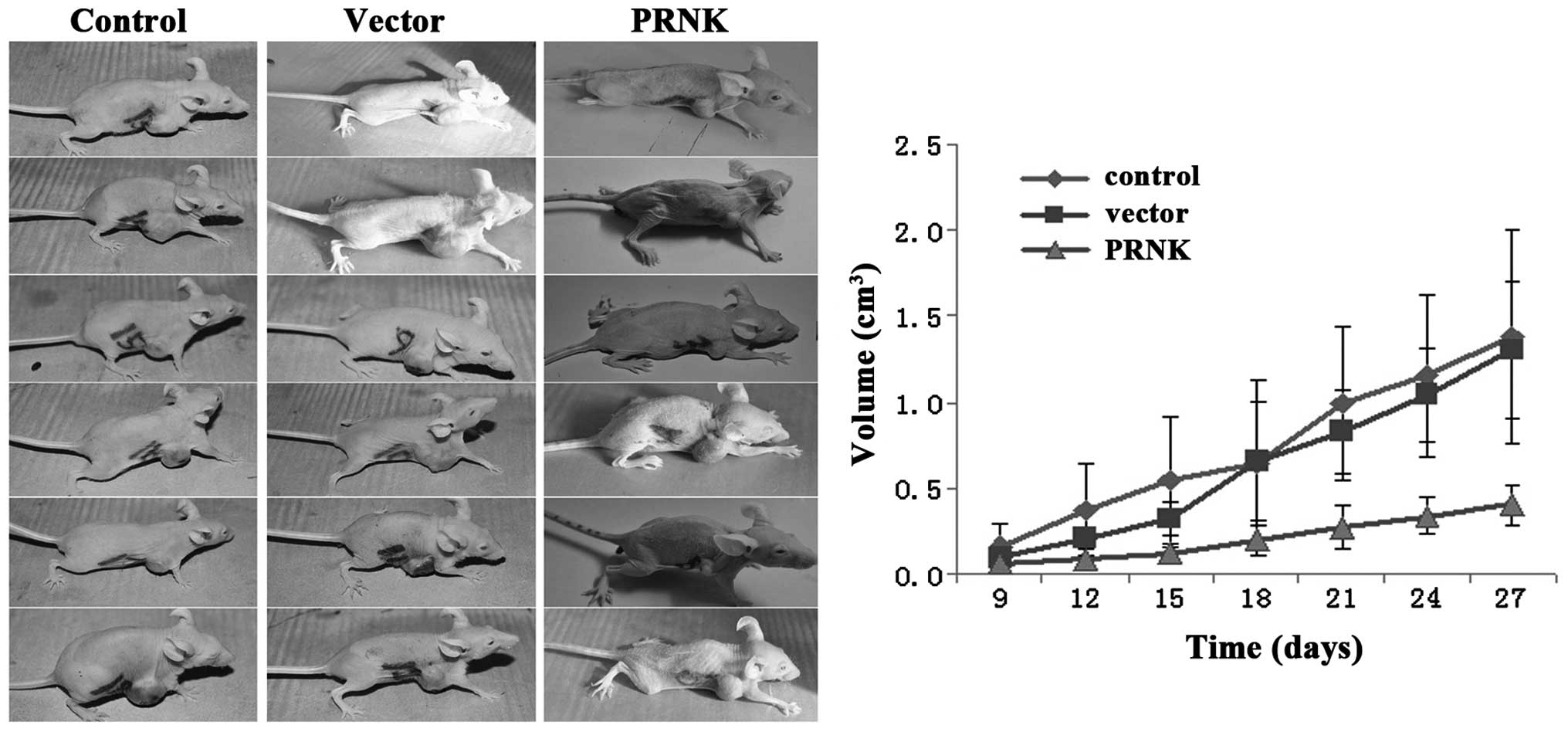
The subcutaneous model of human SCCHN in nude mice
was used to assess the role of Pyk2 in tumorigenesis in
vivo. The stable PRNK-expressing PCI-37B and control
vector-transfected PCI-37B cells were subcutaneously injected into
the breast of male nude mice. Tumor growth curves were obtained. As
shown in Fig. 9, after injection,
the PCI-37B cells developed into a tumor mass on day 9 and grew for
27 days. The tumors formed by the control vector-transfected
PCI-37B cells exhibited the same growth rate as the control PCI-37B
cell group, and reached a volume of ~1.5 cm3 on day 27.
However, the stable PRNK-expressing PCI-37B cells grew slowly in
the bodies of the nude mice, and on day 27, only formed small
tumors, which were no more than 0.5 cm3. There was a
significant difference in tumor size between the cells expressing
PRNK or the cells that did not. This finding indicates that Pyk2
activity promotes SCCHN tumorigenesis in vivo.
Discussion
Pyk2 is a non-receptor tyrosine kinase which is
believed to play a role in transducing extracellular matrix
(ECM)-derived survival signals into cells. The functions of Pyk2
are linked to autophosphorylation of their specific tyrosine
residues, and then association with different signaling proteins
which mediate activation of downstream targets. Thus, modulation of
Pyk2 autophosphorylation may affect several intracellular pathways
and may participate in a variety of pathological settings (21). The Pyk2 related non-kinase (PRNK) is
a recently described isoform of Pyk2 that encodes part of the
COOH-terminal domain of Pyk2 (22).
As a splice variant form of Pyk2, PRNK will interact with some of
the Pyk2-binding partners due to sequence similarity with Pyk2.
Therefore, it has been reported as a dominant-negative regulator to
prevent Pyk2 binding with its partners for signaling pathway
activation and PRNK has been used in numerous studies (23).
In the present study, we constructed stable
PRNK-expressing SCCHN cells, and demonstrated the transfection
efficacy by examining EGFP. CCR7 siRNA, which induces CCR7
silencing (16), blocked the Pyk2
phosphorylation induced by CCL19 as well as PRNK, and had no effect
on Pyk2 expression. This indicated that Pyk2 phosphorylation was
CCR7 activation-dependent. This finding was similar to the results
of a previous study (12).
Squamous cell carcinoma of the head and neck
exhibits metastatic potential. Metastasis involves the separation
of cells from the primary tumor, migration into the extracellular
matrix, blood vessel invasion, adhesion to endothelium and
extravasation and growth in a secondary organ (24). Pyk2 has been reported to widely
participate in this process. In hepatocellular carcinoma,
overexpression of Pyk2 resulted in an upregulation of colony
formation, adhesion toward laminin, cell motility and wound
recovery (25). In malignant
glioma, overexpression of Pyk2 stimulated cell migration (22). Doxorubicin-resistant MCF-7
(MCF-7/Dox) breast cancer cells expressed constitutively active
forms of Pyk2 compared with the expression in parental MCF-7 cells.
α-naphthoflavone (ANF) is a well-known chemopreventive agent; its
anticancer properties were found to function by inhibition of
clonogenic cell survival via de-phosphorylation of FAK, Pyk2 and
EGF-induced Akt in MCF-7/Dox cells and tumor xenografts (26). In a previous study, following
induction with CCL19, SCCHN cells exhibited high viability, a low
rate of apoptosis, high migratory ability, high invasive ability
and adhesion capability (9–11,27,28).
In the present study, we found that the cellular functions mediated
by CCL19 were abrogated by PRNK, which competitively inhibited Pyk2
activity, suggesting that Pyk2 widely participates in the
CCR7-mediated SCCHN cell survival and metastatic ability. In order
to demonstrate this result in vivo, a nude mouse
tumor-bearing model was performed. We found that stable PRNK
expression in SCCHN cells significantly inhibited tumor growth in
nude mice, which supported the in vitro results.
The activity of Pyk2 regulates physiological
processes by many target proteins. Our previous study demonstrated
that the Pyk2 inhibitor Tyrphostin A9 blocked CCL19-induced STAT3
phosphorylation, and this STAT3 phosphorylation was involved in the
migration and invasion of SCCHN cells (11). CCL19 was also found to decrease
E-cadherin expression and increase vimentin expression in SCCHN
cells (10). The constitutive
expression of E-cadherin in both normal and cancer cells may help
to maintain adherence junctions and subsequently decrease the
capacity of cells to invade or migrate through the extracellular
matrix (29,30). The important role that EMT plays in
tumor progression is known to partly depend on dismantling
cadherin-mediated cell-cell junctions (31). Vimentin, the major intermediate
filament (IF) protein of mesenchymal cells, is also associated with
EMT. In the present study, when Pyk2 was inactivated, low
E-cadherin expression was increased and high vimentin expression
was decreased, although with CCL19 induction. Therefore, we
presumed that E-cadherin and vimentin are downstream target
molecules of CCR7-Pyk2, and this signaling pathway may participate
in carcinoma cell growth, separation from the primary tumor,
migration into the extracellular matrix, invasion into the
lymphatic system, adherence to the endothelium and thus achieve
metastasis.
Taken together, based on our in vivo and
in vitro study, our results support the conclusion that Pyk2
plays a key role in CCR7-mediated regulation of SCCHN cell
metastasis and viability. The present study will aid in the
understanding of the mechanism involved in the regulation of CCR7
in SCCHN tumorigenesis and progression. Our ultimate aim is to
develop therapeutic methods according to our basic research in the
near future.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (no. 81372877), the National
Young Scholars Science Foundation of China (no. 81102058), the
Foundation of Education Bureau of Liaoning Province (nos. 2009A755
and L2014317), the Public Welfare Fund Project for Science of
Liaoning Province (no. 2011002001), the Natural Science Foundation
of Liaoning Province (no. 2014021096), and the Excellent Talent
Fund Project of Higher Education of Liaoning Province
(LJQ2014087).
Abbreviations:
|
Pyk2
|
praline-rich tyrosine kinase-2
|
|
CCR7
|
chemokine receptor 7
|
|
SCCHN
|
squamous cell carcinoma of the head
and neck
|
|
PRNK
|
praline-rich tyrosine kinase-2 related
non-kinase
|
|
EGFP
|
enhanced green fluorescent protein
|
|
CCL19
|
c-c motif chemokine 19
|
|
ANF
|
α-naphthoflavone
|
|
EMT
|
epithelial-mesenchymal transition
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
OD
|
optical density
|
|
FITC
|
fluorescein isothiocyanate
|
References
|
1
|
Greenlee RT, Hill-Harmon MB, Murray T and
Thun M: Cancer statistics, 2001. CA Cancer J Clin. 51:15–36. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Butcher EC, Williams M, Youngman K, Rott L
and Briskin M: Lymphocyte trafficking and regional immunity. Adv
Immunol. 72:209–253. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Campbell JJ and Butcher EC: Chemokines in
tissue-specific and microenvironment-specific lymphocyte homing.
Curr Opin Immunol. 12:336–341. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morales J, Homey B, Vicari AP, Hudak S,
Oldham E, Hedrick J, Orozco R, Copeland NG, Jenkins NA, McEvoy LM,
et al: CTACK, a skin-associated chemokine that preferentially
attracts skin-homing memory T cells. Proc Natl Acad Sci USA.
96:14470–14475. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zlotnik A and Yoshie O: Chemokines: A new
classification system and their role in immunity. Immunity.
12:121–127. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
van den Bosch T, Koopmans AE, Vaarwater J,
van den Berg M, de Klein A and Verdijk RM: Chemokine receptor CCR7
expression predicts poor outcome in uveal melanoma and relates to
liver metastasis whereas expression of CXCR4 is not of clinical
relevance. Invest Ophthalmol Vis Sci. 54:7354–7361. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo J, Lou W, Ji Y and Zhang S: Effect of
CCR7, CXCR4 and VEGF-C on the lymph node metastasis of human
pancreatic ductal adenocarcinoma. Oncol Lett. 5:1572–1578.
2013.PubMed/NCBI
|
|
8
|
Wang WN, Chen Y, Zhang YD and Hu TH: The
regulatory mechanism of CCR7 gene expression and its involvement in
the metastasis and progression of gastric cancer. Tumour Biol.
34:1865–1871. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao ZJ, Liu FY, Li P, Ding X, Zong ZH and
Sun CF: CCL19-induced chemokine receptor 7 activates the
phosphoinositide-3 kinase-mediated invasive pathway through Cdc42
in metastatic squamous cell carcinoma of the head and neck. Oncol
Rep. 25:729–737. 2011.
|
|
10
|
Liu FY, Safdar J, Li ZN, Fang QG, Zhang X,
Xu ZF and Sun CF: CCR7 regulates cell migration and invasion
through MAPKs in metastatic squamous cell carcinoma of head and
neck. Int J Oncol. 45:2502–2510. 2014.PubMed/NCBI
|
|
11
|
Liu FY, Safdar J, Li ZN, Fang QG, Zhang X,
Xu ZF and Sun CF: CCR7 regulates cell migration and invasion
through JAK2/STAT3 in metastatic squamous cell carcinoma of the
head and neck. Biomed Res Int. 2014:4153752014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang L, Liu F, Xu Z, Guo N, Zheng X and
Sun C: Chemokine receptor 7 via proline-rich tyrosine kinase-2
upregulates the chemotaxis and migration ability of squamous cell
carcinoma of the head and neck. Oncol Rep. 28:1659–1664.
2012.PubMed/NCBI
|
|
13
|
Wang J, Zhang X, Thomas SM, Grandis JR,
Wells A, Chen ZG and Ferris RL: Chemokine receptor 7 activates
phosphoinositide-3 kinase-mediated invasive and prosurvival
pathways in head and neck cancer cells independent of EGFR.
Oncogene. 24:5897–5904. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mburu YK, Wang J, Wood MA, Walker WH and
Ferris RL: CCR7 mediates inflammation-associated tumor progression.
Immunol Res. 36:61–72. 2006. View Article : Google Scholar
|
|
15
|
Xiong WC, Macklem M and Parsons JT:
Expression and characterization of splice variants of PYK2, a focal
adhesion kinase-related protein. J Cell Sci. 111:1981–1991.
1998.PubMed/NCBI
|
|
16
|
Zhao ZJ, Liu FY and Sun CF: Effect of
chemokine receptor 7 small interfering RNA on proliferation and
invasion of squamous cell carcinoma of head and neck. Zhonghua Kou
Qiang Yi Xue Za Zhi. 44:5–10. 2009.In Chinese. PubMed/NCBI
|
|
17
|
Liu FY, Zhao ZJ, Li P, Ding X, Guo N, Yang
LL, Zong ZH and Sun CF: NF-κB participates in chemokine receptor
7-mediated cell survival in metastatic squamous cell carcinoma of
the head and neck. Oncol Rep. 25:383–391. 2011.
|
|
18
|
Liu FY, Zhao ZJ, Li P, Ding X and Sun CF:
The effect of CCL19 on the viability of head and neck squamous
cancer cells. Shanghai Kou Qiang Yi Xue. 19:158–161. 2010.In
Chinese. PubMed/NCBI
|
|
19
|
Liu FY, Zhao ZJ, Li P, Ding X, Zong ZH and
Sun CF: Mammalian target of rapamycin (mTOR) is involved in the
survival of cells mediated by chemokine receptor 7 through PI3K/Akt
in metastatic squamous cell carcinoma of the head and neck. Br J
Oral Maxillofac Surg. 48:291–296. 2010. View Article : Google Scholar
|
|
20
|
Guo N, Liu F, Yang L, Huang J, Ding X and
Sun C: Chemokine receptor 7 enhances cell chemotaxis and migration
of metastatic squamous cell carcinoma of head and neck through
activation of matrix metalloproteinase-9. Oncol Rep. 32:794–800.
2014.PubMed/NCBI
|
|
21
|
Ziemka-Nalecz M, Jaworska J, Sypecka J and
Zalewska T: OGD induced modification of FAK-and PYK2-coupled
pathways in organotypic hippocampal slice cultures. Brain Res.
1606:21–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lipinski CA, Tran NL, Bay C, Kloss J,
McDonough WS, Beaudry C, Berens ME and Loftus JC: Differential role
of proline-rich tyrosine kinase 2 and focal adhesion kinase in
determining glioblastoma migration and proliferation. Mol Cancer
Res. 1:323–332. 2003.PubMed/NCBI
|
|
23
|
Sun CK, Ng KT, Lim ZX, Cheng Q, Lo CM,
Poon RT, Man K, Wong N and Fan ST: Proline-rich tyrosine kinase 2
(Pyk2) promotes cell motility of hepatocellular carcinoma through
induction of epithelial to mesenchymal transition. PLoS One.
6:e188782011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liotta LA, Steeg PS and Stetler-Stevenson
WG: Cancer metastasis and angiogenesis: An imbalance of positive
and negative regulation. Cell. 64:327–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun CK, Man K, Ng KT, Ho JW, Lim ZX, Cheng
Q, Lo CM, Poon RT and Fan ST: Proline-rich tyrosine kinase 2 (Pyk2)
promotes proliferation and invasiveness of hepatocellular carcinoma
cells through c-Src/ERK activation. Carcinogenesis. 29:2096–2105.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Datta A, Bhasin N, Kim H, Ranjan M, Rider
B, Abd Elmageed ZY, Mondal D, Agrawal KC and Abdel-Mageed AB:
Selective targeting of FAK-Pyk2 axis by alpha-naphthoflavone
abrogates doxorubicin resistance in breast cancer cells. Cancer
Lett. 362:25–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li P, Liu F, Sun L, Zhao Z, Ding X, Shang
D, Xu Z and Sun C: Chemokine receptor 7 promotes cell migration and
adhesion in metastatic squamous cell carcinoma of the head and neck
by activating integrin αvβ3. Int J Mol Med. 27:679–687.
2011.PubMed/NCBI
|
|
28
|
Li P, Zhao ZJ, Liu FY, Sun LY, Ding X,
Zhang WZ, Shang DH and Sun CF: The chemokine receptor 7 regulates
cell adhesion and migration via β1 integrin in metastatic squamous
cell carcinoma of the head and neck. Oncol Rep. 24:989–995.
2010.PubMed/NCBI
|
|
29
|
Vleminckx K, Vakaet L Jr, Mareel M, Fiers
W and van Roy F: Genetic manipulation of E-cadherin expression by
epithelial tumor cells reveals an invasion suppressor role. Cell.
66:107–119. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Frixen UH, Behrens J, Sachs M, Eberle G,
Voss B, Warda A, Löchner D and Birchmeier W: E-cadherin-mediated
cell-cell adhesion prevents invasiveness of human carcinoma cells.
J Cell Biol. 113:173–185. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Savagner P: Leaving the neighborhood:
Molecular mechanisms involved during epithelial-mesenchymal
transition. Bioessays. 23:912–923. 2001. View Article : Google Scholar : PubMed/NCBI
|