Introduction
Small cell lung cancer (SCLC) accounts for ~15% of
all lung cancer cases (1,2); however, increased attention has been
paid to this histotype due to its rapid growth and early
dissemination to regional lymph nodes and remote organs (3). Chemotherapy has dramatically improved
the survival rate of SCLC patients, yet multi-drug resistance
(MDR), emerging after chemotherapy, leads to relapse and mortality
(4). MDR has become the major
clinical obstacle to the treatment of SCLC. New chemotherapeutics
and adjustment of chemotherapy programs have not improved the
long-term survival rates. Research on the mechanisms of MDR in SCLC
has continuously increased since a main treatment for SCLC has not
markedly evolved during the past three decades.
To date, it has not been fully elucidated how
acquired MDR evolves, yet various meaningful theories and ideas
have guided the direction of research efforts. Research has
confirmed that MDR is caused by different mechanisms. What is more,
there could be more than one mechanism involved in one type of
cancer (5,6). Moreover, a theory, known as the
'Warburg effect', concerning energy metabolism reprogramming was
introduced and recognized to be closely related to tumorigenesis
and cancer progression. Warburg first proposed that cancer cells,
different from normal cells, increase their glycolytic activity and
reduce mitochondrial respiration whether there is an abundant
supply of oxygen or not (7,8). Increasing evidence indicates that
numerous molecular mechanisms are involved in the Warburg effect
(9), and mitochondria were first to
be indicated since this organelle has been so closely related to
energy metabolism and cell survival. An abundance of research
indicates that mitochondrial dysfunction resulting from mutation in
mitochondrial DNA (mtDNA) is responsible for tumorigenesis
(10).
Mitochondrial dysfunction is also believed to
provide a survival benefit to cancer cells (11). It has been reported in several
cancer types such as prostate (12), kidney (13), breast (14), liver (15), colon and rectal cancer cells that
mutations and/or depletion of mtDNA are the reasons for the
multi-drug-resistant features (16). More importantly,
chemotherapy-induced low levels of mtDNA have been proven to be
related to acquired drug resistance and anti-apoptotic properties
of cancer cells (17,18). A typical example was that mtDNA
mutations and reduced mtDNA copy number in breast and prostate
cancer patients were associated with enhanced metastasis and poor
patient prognosis (12,17,18).
It appears that mitochondrial dysfunction resulting from mtDNA
mutation is closely associated with cancer occurrence and
progression. However, there is no research clearly illustrating the
possible role of mtDNA mutation in tumorigenesis and MDR of
SCLC.
Thus, we investigated the sequence of mtDNA from
SCLC cell lines, H446 and H446/CDDP, and compared these sequences
with the revised Cambridge reference sequence (rCRS) (19) in order to ascertain whether mtDNA
mutations participate in the occurrence and acquired MDR of SCLC.
After that, the differences in mitochondrial function were
evaluated between H446 and H446/CDDP cells. Finally, we
investigated the activation of the mitochondrial apoptotic pathway
in H446 and H446/CDDP cells challenged by cisplatin in order to
illustrate the possible mechanism of the mitochondrial dysfunction
in the MDR of SCLC.
Materials and methods
Cell lines and culture
Small cell lung cancer cell line H446 and its
multi-drug-resistant cell line H446/CDDP were kindly gifted by Dr
Guisheng Qian (Institute of Human Respiratory Disease, Xinqiao
Hospital, Third Military Medical University). The H446 cell line
was purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and the MDR properties of H446/CDDP cells were
induced by a low dosage of cisplatin. These cells were maintained
in RPMI-1640 medium with 10% fetal calf serum (Gibco-BRL, Grand
Island, NY, USA) in a humidified atmosphere containing 5%
CO2 at 37°C. In order to maintain the MDR properties of
the H446/CDDP cells, cisplatin (with a final concentration of 0.5
µg/ml) was added to the culture medium for the H446/CDDP
cells.
Evaluation of drug resistance
The sensitivity of H446 and H446/CDDP cells to
cisplatin was evaluated by a microculture tetrazolium (MTT) assay.
Briefly, the cells were seeded in 96-well plates at a density of
1×104 cells/well. The cells were treated with 5
µg/ml cisplatin for 24, 48 and 72 h when grown to
confluency. The supernatant was discarded at corresponding
time-points, and 20 µl MTT (5 mg/ml, dissolved in PBS and
filtered through a 0.22-mm membrane) was added into each well and
incubated for 4 h at 37°C. Finally, the absorption values were
determined at 492 nm on an automated Bio-Rad 550 microtiter plate
reader.
Extraction of genomic DNA
Genomic DNAs were extracted from the H446 and
H446/CDDP cells in a logarithmic growth phase according to the
genomic DNA extraction kit instructions (Tiangen Biotechnology Co.,
Beijing, China). Optical density values of A260 and A280 were
determined by using an ultraviolet spectrophotometer in order to
evaluate the purity of the DNAs.
Amplification and sequencing of
mtDNA
mtDNAs were amplified by using 26 primers (sequences
shown in Table I) according to
previous research (12). To amplify
mtDNA in the genomic DNA samples, 1 µg of diluted genomic
DNA was mixed with paired primers (primers were provided by AuGCT
Biotechnology Co., Beijing, China). Along with genomic sequences,
Premix Taq version 2.0 (Takara Biotechnology Co., Dalian, China)
was added into a 20-µl total volume. The mtDNA was amplified
under the following conditions: 35 cycles, and the cycling
conditions were as follows: 95°C for 30 sec; 54°C for 30 sec; and
72°C for 1 min. After that, the PCR products were purified and
sequenced by ABI Prism 3700 sequencing machine.
 | Table ISequences of primers for the mtDNAs
used in the present research. |
Table I
Sequences of primers for the mtDNAs
used in the present research.
| Primer pair no. | Primer no. | Sequence 5′-3′ | 3′ combined
location | Length (bp) | Overlapping length of
two-way sequencing (bp) |
|---|
| 1 | 1F |
CTCCTCAAAGCAATACACTG | 611 | 840 | 202 |
| 1R |
TGCTAAATCCACCTTCGACC | 1411 | | |
| 2 | 2F |
CGATCAACCTCACCACCTCT | 1245 | 802 | 204 |
| 2R |
TGGACAACCAGCTATCACCA | 2007 | | |
| 3 | 3F |
GGACTAACCCCTATACCTTCTGC | 1854 | 860 | 196 |
| 3R |
GGCAGGTCAATTTCACTGGT | 2669 | | |
| 4 | 4F |
AAATCTTACCCCGCCTGTTT | 2499 | 887 | 208 |
| 4R |
AGGAATGCCATTGCGATTAG | 3346 | | |
| 5 | 5F |
TACTTCACAAAGCGCCTTCC | 3169 | 832 | 215 |
| 5R |
ATGAAGAATAGGGCGAAGGG | 3961 | | |
| 6 | 6F |
TGGCTCCTTTAACCTCTCCA | 3796 | 898 | 203 |
| 6R |
AAGGATTATGGATGCGGTTG | 4854 | | |
| 7 | 7F |
ACTAATTAATCCCCTGGCCC | 4485 | 975 | 207 |
| 7R |
CCTGGGGTGGGTTTTGTATG | 5420 | | |
| 8 | 8F |
CTAACCGGCTTTTTGCCC | 5255 | 814 | 201 |
| 8R |
ACCTAGAAGGTTGCCTGGCT | 6031 | | |
| 9 | 9F |
GAGGCCTAACCCCTGTCTTT | 5855 | 827 | 214 |
| 9R |
ATTCCGAAGCCTGGTAGGAT | 6642 | | |
| 10 | 10F |
CTCTTCGTCTGATCCGTCCT | 6469 | 886 | 211 |
| 10R |
AGCGAAGGCTTCTCAAATCA | 7315 | | |
| 11 | 11F |
ACGCCAAAATCCATTTCACT | 7148 | 987 | 205 |
| 11F |
CGGGAATTGCATCTGTTTTT | 8095 | | |
| 12 | 12F |
ACGAGTACACCGACTACGGC | 7937 | 900 | 196 |
| 12R |
TGGGTGGTTGGTGTAAATGA | 8797 | | |
| 13 | 13F |
TTTCCCCCTCTATTGATCCC | 8621 | 816 | 214 |
| 13R |
GTGGCCTTGGTATGTGCTTT | 9397 | | |
| 14 | 14F |
CCCACCAATCACATGCCTAT | 9230 | 940 | 205 |
| 14R |
TGTAGCCGTTGAGTTGTGGT | 10130 | | |
| 15-1 | 15-1F |
CTTCTATTGATGAGGGTCTT | 9991 | 670 | 218 |
| 15-1R |
GGTGTTGAGGGTTATGAGA | 10622 | | |
| 15-2 | 15-2F |
AAGGATTAGACTGAACCGAA | 10404 | 627 | 135 |
| 15-2R |
CTGATTGTGAGGGGTAGGA | 10992 | | |
| 15-3 | 15-3F |
CAACCACCCACAGCCTAA | 10857 | 693 | 198 |
| 15-3R |
TTGAGAATGAGTGTGAGGCG | 11512 | | |
| 17 | 17F |
TCACTCTCACTGCCCAAGAA | 11314 | 802 | 196 |
| 17R |
GGAGAATGGGGGATAGGTGT | 12076 | | |
| 18 | 18F |
TATCACTCTCCTACTTACAG | 11948 | 866 | 166 |
| 18R |
AGAAGGTTATAATTCCTACG | 12772 | | |
| 19 | 19F |
AAACAACCCAGCTCTCCCTAA | 12571 | 977 | 242 |
| 19R |
TCGATGATGTGGTCTTTGGA | 13507 | | |
| 20 | 20F |
ACATCTGTACCCACGCCTTC | 13338 | 970 | 207 |
| 20R |
AGAGGGGTCAGGGTTCATTC | 14268 | | |
| 21 | 21F |
GCATAATTAAACTTTACTTC | 14000 | 938 | 206 |
| 21R |
AGAATATTGAGGCGCCATTG | 14998 | | |
| 22 | 22F |
TGAAACTTCGGCTCACTCCT | 14856 | 1,162 | 180 |
| 22R |
AGCTTTGGGTGCTAATGGTG | 15978 | | |
| 23 | 23F |
TCATTGGACAAGTAGCATCC | 15811 | 765 | 190 |
| 23R |
GAGTGGTTAATAGGGTGATAG | 5 | | |
| 24-1 | 24-1F |
CATTATCCCGCACAAGAGTG | 16419 | 420 | 160 |
| 24-1R |
TGGAAAGTGGCTGTGCAGACAT | 250 | | |
| 24-2 | 24-2F |
CTTTGATTCCTGCCTCATCC | 132 | 540 | 100 |
| 24-2R |
TAGAAAGGCTAGGACCAAACCT | 652 | | |
Mitochondrial ultrastructure
examination
Electron microscopy was performed on H446 and
H446/CDDP cells in order to investigate whether the difference of
mitochondrial structure was related to the varied sensitivity of
H446 and H446/CDDP cells to cisplatin. Cell mass was collected by
centrifugal method, and 1 µm thick sections were stained to
locate significant areas for electron microscopic examination.
Ultrathin sections were stained with uranyl acetate, and
mitochondria were examined under a TEM-100CX electron microscope
(Japan Electron Optics Laboratory, Tokyo, Japan) after the cell
mass was post-fixed in osmium tetroxide for 1 h, dehydrated in
alcohol, and embedded in epoxy resin.
Mitochondrial staining
MitoTracker probe (Life Technologies Corporation,
USA) was used to dye mitochondria in the H446 and H446/CDDP cells,
and the procedure was carried out according to the manufacturer's
instructions. Briefly, medium was removed from the H446 and
H446/CDDP cells in logarithmic growth phase, and the cells were
washed 3 times with phosphate-buffered saline (PBS; Life
Technologies Corporation). The cells were then incubated with
warmed (37°C) staining solution containing MitoTracker probe (100
nM) for 30 min. After that, the cells were washed again with PBS
and examined with an inverted fluorescence microscope (DMI6000B;
Leica, Germany) within half an hour.
Detection of lactic acid secretion
In order to determine the metabolic difference of
H446 and H446/CDDP cells, the lactic acid content in the cultural
medium with or without 5 µg/ml cisplatin stimulation was
detected with a lactic acid assay kit (Jiancheng Bioengineering
Institute, Nanjing, China) at different time-points. The procedure
was carried out according to the manufacturer's instructions, and
the data were collected and are expressed as mean ± standard
deviation.
Reactive oxygen species (ROS)
detection
In order to evaluate the function of mitochondria in
the H446 and H446/CDDP cells, ROS levels in the cells with or
without cisplatin stimu lation were detected with DCFH-DA (Beyotime
Institute of Biotechnology, China). Briefly, the cells were seeded
in laser confocal Petri dishes at a density of
1x104cells/well and incubated for 48 h. Then, 500
µl of DCFH-DA working liquid (1:1,500 diluted in RPMI-1640
medium and filtered through a 0.22-mm membrane) was added into each
plate. Inverted confocal microscope (FV1000 IX81; Olympus) was used
to capture the fluorescence intensity every 5 min a time since
cells were challenged by cisplatin. Finally, fluorescence intensity
in the images were digitalized and analyzed by Image-Pro Plus
software.
Analysis of mitochondrial membrane
potential (MMP)
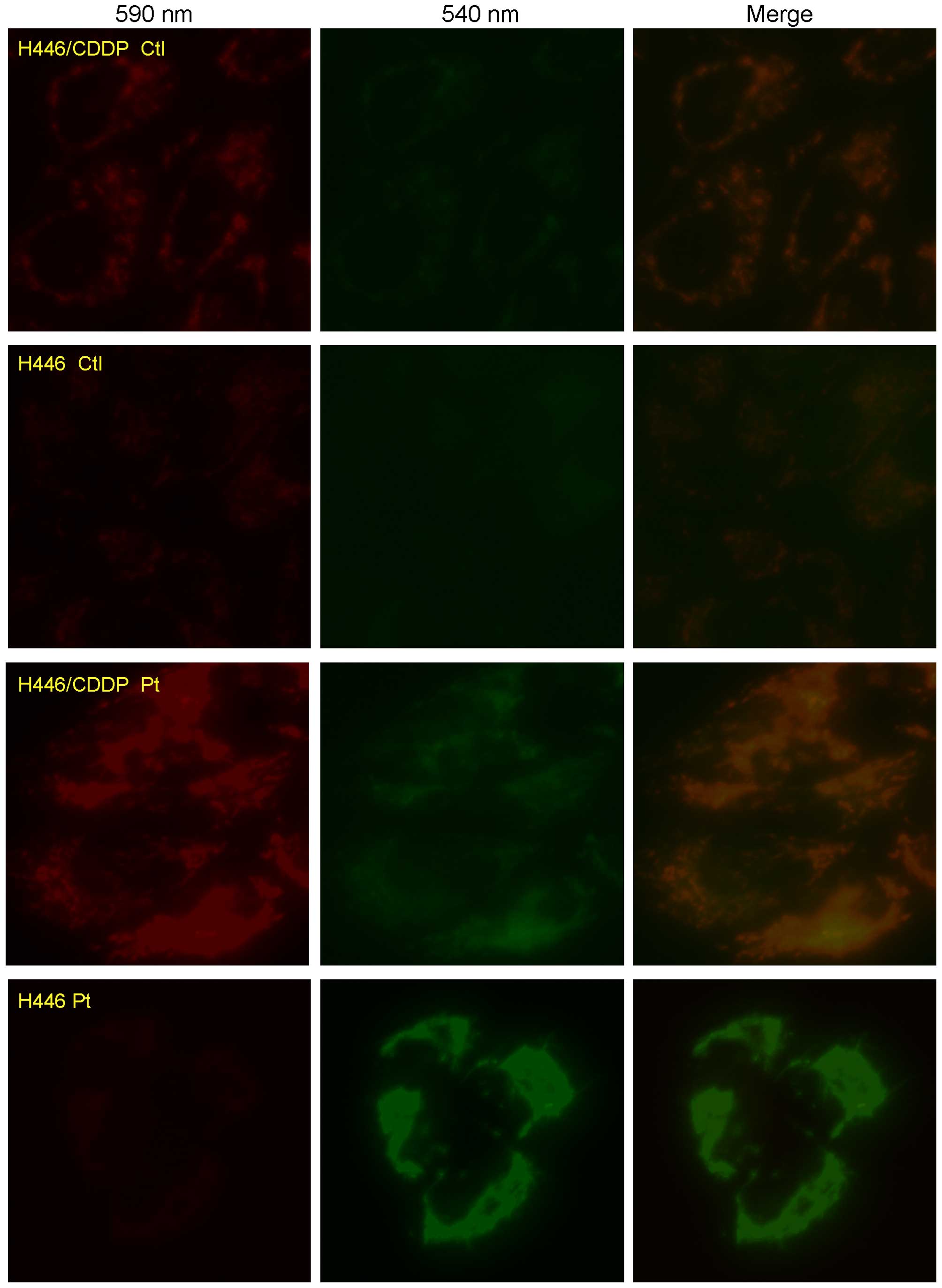
Tetrachloro-tetraethylbenzimidazol carbocyanine
iodide (JC-1; Beyotime Institute of Biotechnology) is a
mitochondrial-specific cationic dye. It is a monomer when the
mitochondrial membrane potential is <120 mV and emits a green
light (540 nm) following excitation by blue light (490 nm). When
JC-1 is converted to J-aggregates, a red light (590 nm) is emitted
following excitation by green light (540 nm) (20). In brief, H446 and H446/CDDP cells
were plated at a density of 2×105 cells/well in a
24-well plate, and cells were incubated with 5 µM JC-1 for
30 min after stimulation by cisplatin for 24 h in dark at room
temperature. Finally, fluorescence was captured with an inverted
fluorescence microscope (DMI6000B; Leica), and changes in the
fluorescence intensity ratio between the measurements at
wavelengths of 590 nm (red) and 540 nm (green) were used to
evaluate the MMP.
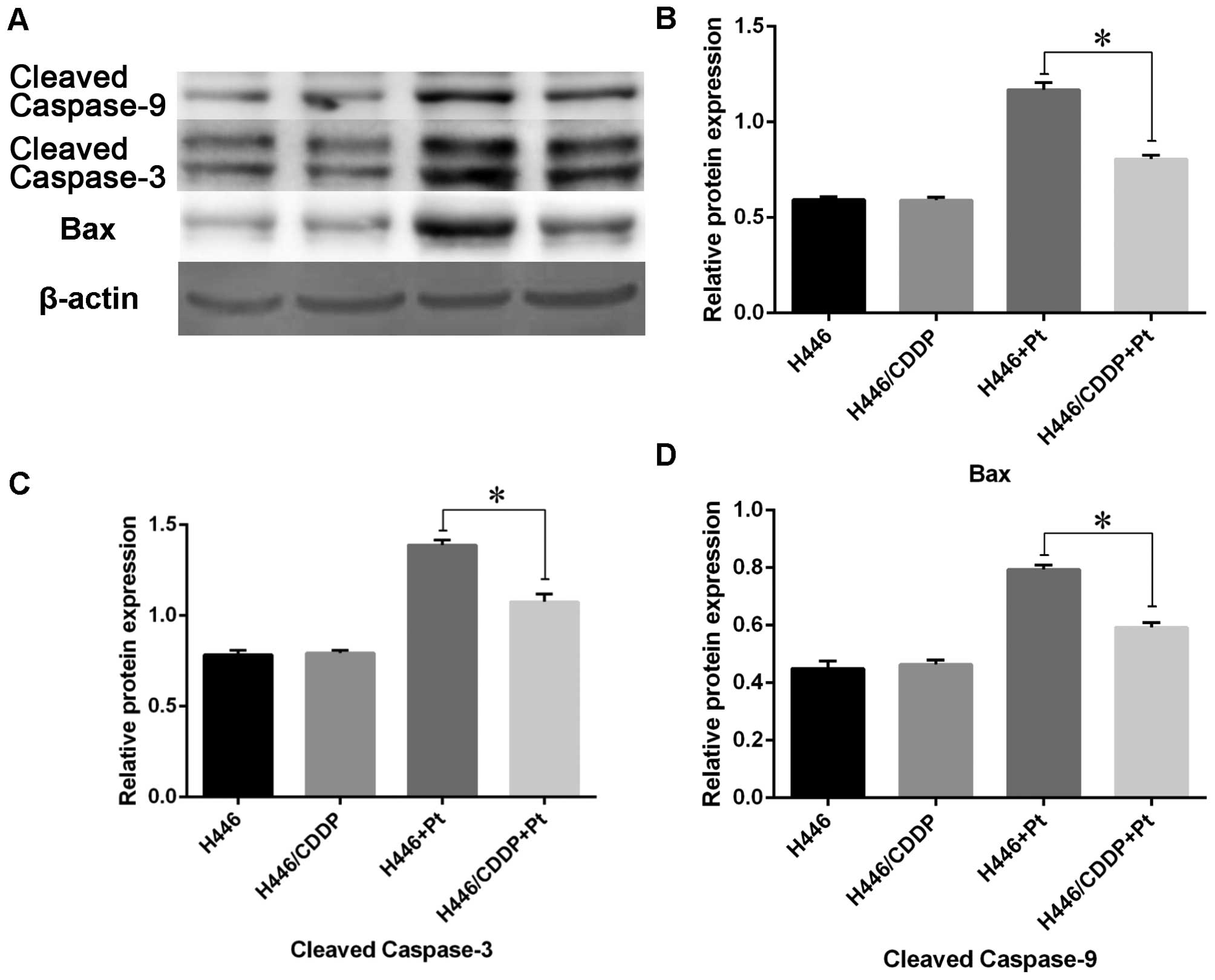
Western blot analysis
In order to confirm the role of mitochondrial
dysfunction in MDR of H446/CDDP cells, activation of the
mitochondrial apoptotic pathway was evaluated by western blot
analysis. Briefly, H446 and H446/CDDP cells with or without
stimulation of cisplatin for 24 h were collected by centrifugation,
and the total protein was extracted by a repeated freezing and
thawing method. Equal amounts of protein from each group were
separated on 12% dodecyl sulfate, sodium salt (SDS)-polyacrylamide
gel electrophoresis (SDS-PAGE) gels and transferred to a PVDF
membrane (Amersham Biosciences, Piscataway, NJ, USA). The membranes
were soaked in blocking buffer {5% skimmed milk melted in TBS-T [25
mM Tris (pH 7.6), 138 mM NaCl and 0.05% Tween-20 for 2 h and then
probed with Bax, cleaved caspase-3, cleaved caspase-9 and β-actin
(1:1,000-1:5,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA)
antibodies overnight at 4°C. After that, the membranes were further
incubated with anti-rabbit IgG HRP-conjugated secondary antibody
(1:5,000). Finally, immune-reactive signals were detected using an
ECL detection system (Amersham Pharmacia Biotech).
Statistical analysis
Statistical analysis was performed with SPSS 17.0.
Numerical variables are expressed as means ± SD. Statistical
differences between the experimental groups were analyzed by
one-way analysis of variance (ANOVA) followed by Dunnett's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
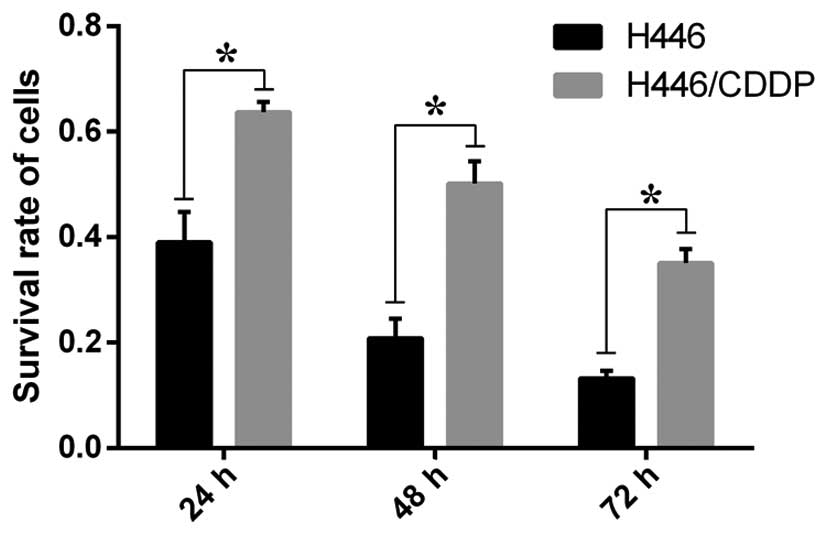
Sensitivity of the H446 and H446/CDDP
cells to cisplatin
The survival rate of the control group was taken as
1, and the results from the different time groups were compared
with the control in order to calculate the relative survival rates.
The survival rates of the H446 and H446/CDDP cells treated with 5
µg/ml cisplatin were significantly different (P<0.01) at
all three time-points (Fig. 1). The
difference in the survival rate between H446 and H446/CDDP cells
increased with a decreasing p-value as the time of stimulation was
prolonged.
Sequencing results of mtDNA from the H446
and H446/CDDP cells
Amplified mtDNAs of the H446 and H446/CDDP cells
were sequenced and compared with rCRS. The mtDNA sequences of the
H446 and H446/CDDP cells belonged to haploid type G (Table II), and several specific base
mutations were detected in the mtDNA sequence of both H446 and
H446/CDDP cells, which could be related to types of amino acid
protein in the mitochondrial respiratory chain and the secondary
structure of tRNAs and rRNAs. It was also found that the mtDNA
sequence of the H446 cell line was the same as that of the
H446/CDDP cells.
| Table IImtDNA mutations and corresponding
changes of amino acids. |
Table II
mtDNA mutations and corresponding
changes of amino acids.
| Cell lines | mtDNA NA type | mtDNA
mutations | Influenced
proteins | Amino acid
changes | tRNA | rRNA |
|---|
| H446 | G | G6366A | CO1 | V155I | tRNA-Cys | |
| H446/CDDP | | A10086G | ND3 | N10D | | |
| | A13105G | ND5 | I257V | | |
| | A15311G | cytB | I189V | | |
| | A15824G | cytB | T360A | | |
| | G5773A | | | | |
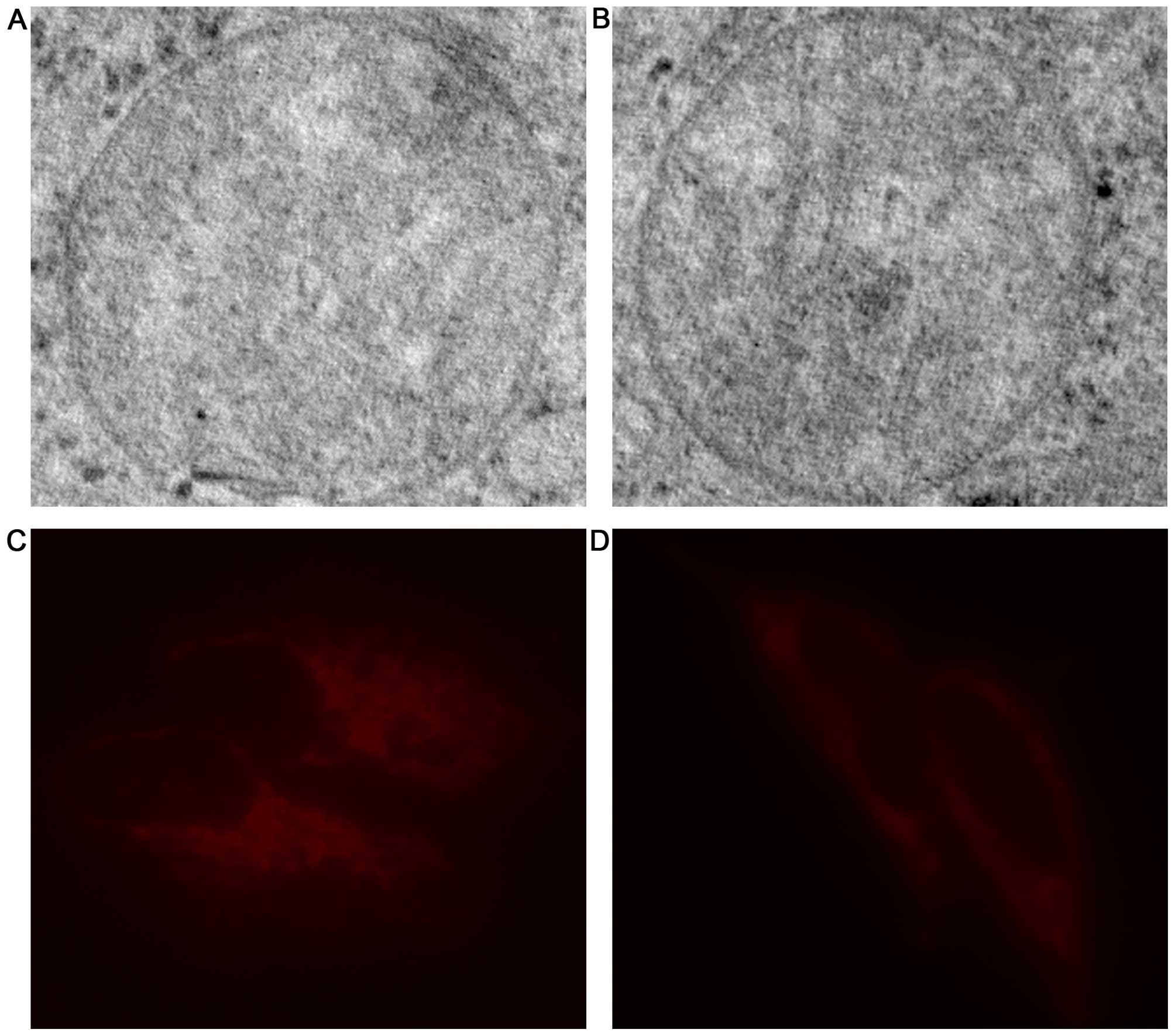
Ultrastructure and content of
mitochondria in the H446 and H446/CDDP cells
Since there was no difference in the sequences of
mtDNA in the H446 and H446/CDDP cells, we explored the structural
and functional differences of mitochondria in these two cell lines.
Results from electron microscopic observation (Fig. 2A and B) revealed no difference in
the ultra-structure of mitochondria in the H446 and H446/CDDP
cells. Then, we compared the number of mitochondria in these two
cell lines. Mitochondrial staining results revealed that the
fluorescence of mitochondria in the H446 cell line (Fig. 2C) was much stronger than that of the
H446/CDDP cells (Fig. 2D).
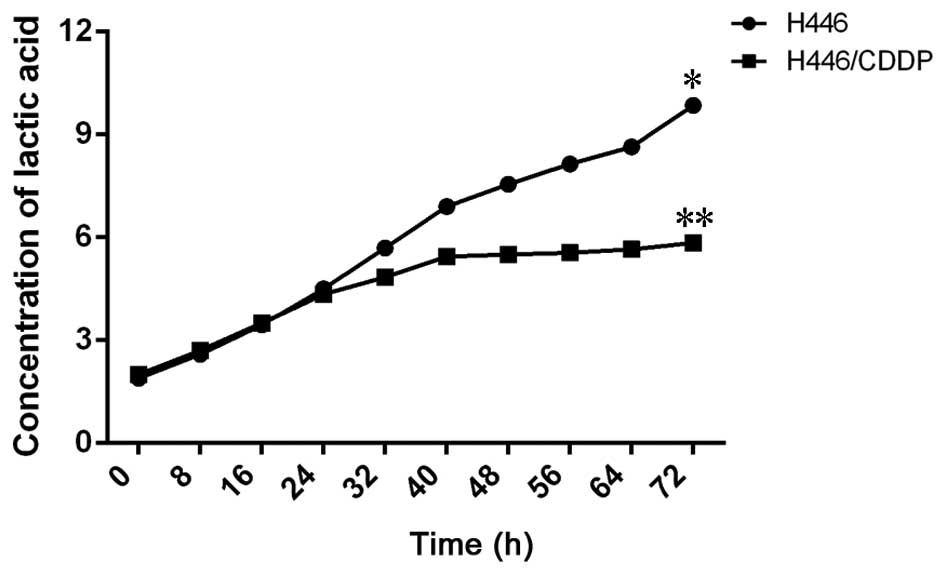
Lactic acid generation in the H446 and
H446/CDDP cells treated with cisplatin
Concentration of lactic acid in the culture
supernatant was detected in order to evaluate the metabolic
difference in the H446 and H446/CDDP cells. The concentration of
lactic acid in the cultural medium from the H446 and H446/CDDP
cells increased at about the same speed when stimulated by
cisplatin (Fig. 3). However, the
generating speed of lactic acid in the H446 cells accelerated 24 h
after stimulation and the concentration of lactic acid in the H446
cells was extremely higher than that in the H446/CDDP cells 72 h
after of cisplatin treatment (P<0.01).
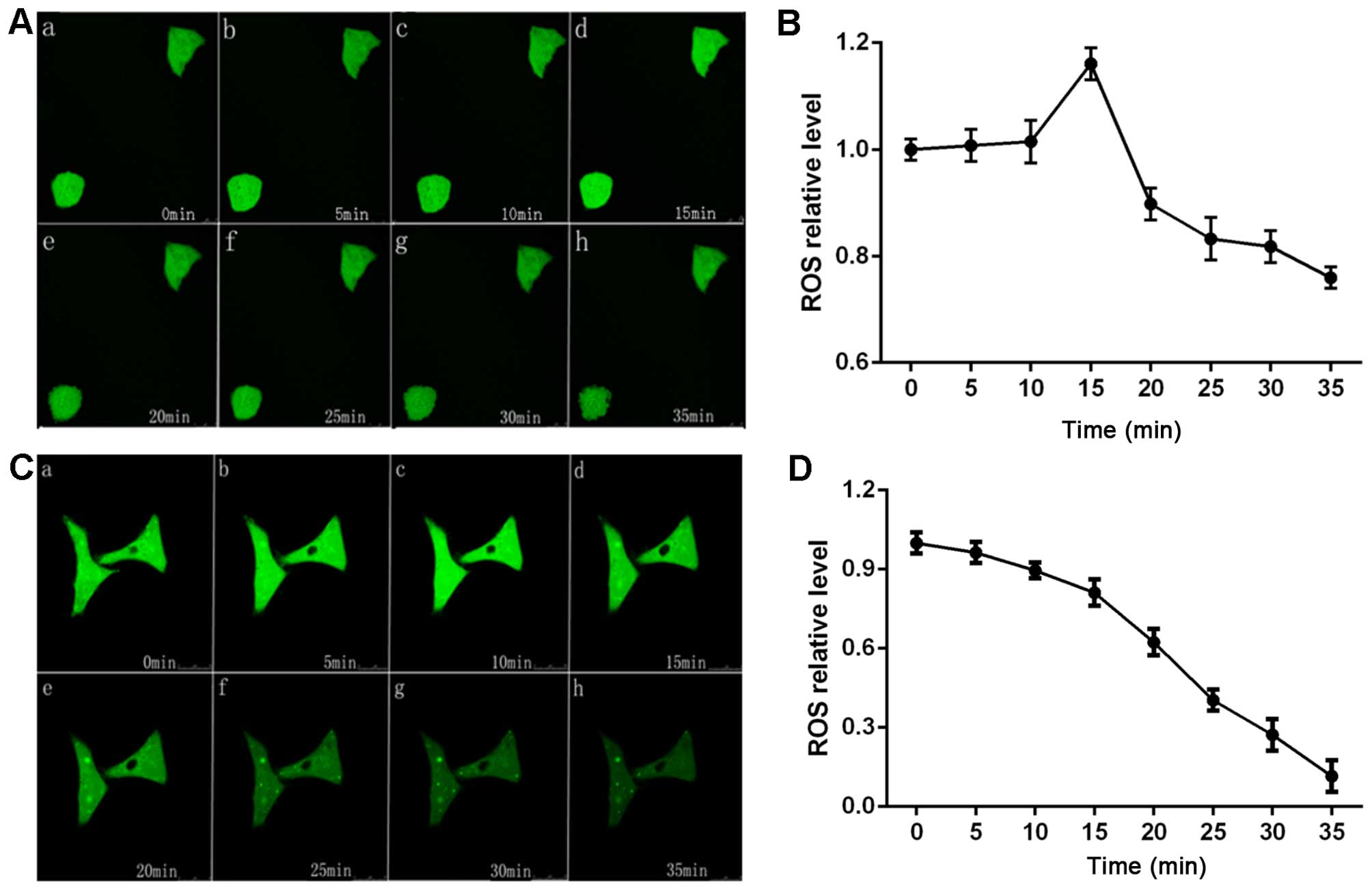
ROS generation in the H446 and H446/CDDP
cells treated with cisplatin
ROS generation in the H446 and H446/CDDP cells was
detected so as to evaluate the ability of these two cell lines to
cope with oxidative stress induced by cisplatin. The ROS content in
the H446 cells increased and peaked at 15 min after being
stimulated by cisplatin, and then the ROS content slowly decreased
(Fig. 4). Meanwhile, H446 cells
became shrunken after being stimulated and it was most obvious 35
min after treatment of cisplatin. On the other hand, ROS in the
H446/CDDP cells decreased at a relatively stable speed when
stimulated with cisplatin and there was no shrinking by this cell
line upon stimulation with cisplatin.
Depolarization of the MMP in the H446 and
H446/CDDP cells
ROS generation is closely related to depolarization
of the MMP which may further lead to cell apoptosis. Depolarization
of MMP in H446 and H446/CDDP cells stimulated by cisplatin was
evaluated by cationic dye JC-1. The green light became stronger in
both the H446 and H446/CDDP cells challenged by cisplatin (Fig. 5). But much stronger green light
against red light was detected in the H446 cells compared with that
of the H446/CDDP cells, which meant that reduction in the MMP in
the H446 cell line was much more obvious than that of the H446/CDDP
cells stimulated by 5 µg/ml cisplatin.
Expression of the mitochondrial apoptosis
pathway in the H446 and H446/CDDP cells treated with cisplatin
Expression of the mitochondrial apoptosis pathway in
the H446 and H446/CDDP cells treated with cisplatin was evaluated
by western blotting. Expression of Bax, cleaved caspase-3 and
cleaved caspase-9 was increased in both the H446 and H446/CDDP
cells when challenged by cisplatin (Fig. 6). However, expression levels of
these molecules in the H446/CDDP cells were much lower than levels
in the H446 cells (P<0.05).
Discussion
Mitochondria are intracellular semi-autonomous
organelles, which possess a self-owned genome and participate in
energy metabolism and homeostasis (21). Human mtDNA is a 16.6-kb
double-stranded, circular DNA molecule encoding for 13 complex
polypeptides belonging to the mitochondrial respiratory chain, 22
transfer RNAs (tRNAs) and 2 ribosomal RNAs (rRNAs) (21). Since mtDNA is essential for cells,
the accumulation of mtDNA mutations and/or reset of mtDNA copy
number are believed to affect energy production, cell proliferation
as well as cell apoptosis (21,22).
It was first investigated, in the present study,
whether mtDNA mutations are associated with the tumorigenesis of
SCLC and the MDR phenotype of H446/CDDP cells. mtDNAs from H446 and
H446/CDDP cells were amplified, sequenced and compared with rCRS.
The results revealed that several spot mutations existed in the
mtDNA of SCLC cell lines, which led to changes of amino acids in
mtDNA-encoded proteins and base-shifts in rRNA. Astonishingly, the
sequences of mtDNA in the H446 cells were the same as that of the
MDR cell line H446/CDDP, which meant that mtDNA mutation in
sequences was not related to the MDR characteristic of the
H446/CDDP cells. Research indicates that both mutations in mtDNA
sequences and low mtDNA copy number are closely related to the
dysfunction of mitochondria. A reduced mtDNA content could result
in impaired mitochondrial respiration and depolarization of the
mitochondrial membrane (23). We
then detected the mitochondrial content in the H446 and H446/CDDP
cells, and found that the fluorescence representing the content of
mitochondria in the H446 cells was much stronger than that of
H446/CDDP cells. This result was consistent with previous reports
indicating that varied mtDNA content markedly increases the risk of
lymph node metastasis and high mortality of patients with
late-stage tumors (24).
Based on the findings that mutations in mtDNA
sequences are responsible for the tumorigenesis of SCLC and varied
mitochondrial content in single cells is related to drug
sensitivity of H466 and H446/CDDP cells, we wondered how this
varied content of mitochondria in the H446 and H446/CDDP cells
influenced their sensitivity to cisplatin. As known, mitochondria
possess many biological functions, including ATP generation,
production of ROS, providing reaction sites for many biological
processes and modulating cell apoptosis (25). Evidence also suggests that an energy
generation-related 'Warburg effect' may be blamed for the survival
benefit of cancer cells (26,27).
Lactic acid is an important metabolite of glycolytic activity which
could be taken as a marker of anaerobic glycolysis. Generation and
secretion of lactic acid from H446 and H446/CDDP cells were
evaluated in order to assess their difference in glycolytic
activity. Results showed that the H446/CDDP cells generated less
lactic acid than that of H446 cells when stimulated by the same
dosage of cisplatin, which indicated that a stronger ability of
lactic acid removal benefited cancer cell survival.
Then we wondered whether this difference in energy
generation was associated with the biological behavior of H446 and
H446/CDDP cells. Naito et al (28) reported that mtDNA depletion promotes
the chemoresistance of breast cancer cells. In addition, HeLa cells
lacking or without mtDNA have been confirmed to become resistant to
adriamycin and photodynamics (29).
More importantly, Park et al (30) showed that hepatoma cells deprived of
mtDNA were less sensitive to hydrogen peroxide and to ROS-inducing
agents and further research illustrated that this drug-resistant
phenotype was associated with increased expression of antioxidant
enzymes manganese superoxide dismutase and glutathione peroxidase.
Based on this knowledge, we investigated the production of ROS in
H446 and H446/CDDP cells stimulated by cisplatin, and the results
showed that more ROS were produced in the H446 cells when
challenged by cisplatin compared with that of the H446/CDDP cells.
Our findings indicated an upgraded antioxidant capacity in
H446/CDDP cells which was also consistent with previous research
illustrating that the insensitivity of cancer cells to
chemotherapeutic drugs paclitaxel and cisplatin is associated with
increased antioxidant expression levels (31,32).
Varied mitochondrial content, difference in lactic
acid secretion and ROS generation were confirmed to be related to
the discriminative sensitivity of H446 and H446/CDDP cells to
cisplatin. While the mechanisms of how those differences
participate in the MDR of SCLC remain unknown. As we know,
mitochondria are crucial centers of metabolism and ROS production
(33). An excess of ROS induces
mitochondrial inner membrane permeabilization leading to
mitochondrial depolarization, swelling, cytochrome c release
and subsequent apoptotic death (34). We further evaluated the activation
of the mitochondrial apoptotic pathway in H446 and H446/CDDP cells
challenged by cisplatin. Molecules of the Bcl-2 family were
evaluated since they are closely related to the mitochondrial
apoptotic pathway, and their abnormal expression leads to loss of
ΔΨm, leakage of apoptogenic proteins, activation of caspase-3, and
finally cell death (35).
Evaluation of the mitochondrial-mediated cell apoptosis pathway
showed that cell apoptosis-promoting proteins, Bax, cleaved
caspase-3 and cleaved caspase-9, were increased in the H446 cells
when compared with the levels in the H446/CDDP cells following
stimulation by cisplatin. This indicated that abnormal generation
of ROS affected the sensitivity of H446 and H446/CDDP cells to
cisplatin probably by interfering with the mitochondrial apoptotic
pathway.
In conclusion, mtDNAs from small cell lung cancer
cell line H446 and its multi-drug-resistant cell line H446/CDDP
were sequenced and compared with rCRS. The results revealed that
mtDNA mutations were responsible for the tumorigenesis but not for
the MDR capacity of SCLC. On the other hand, a difference in lactic
acid and ROS generation confirmed that mitochondrial dysfunction
participated in the MDR of SCLC. Finally, evaluation of the
ROS-related mitochondrial apoptotic pathway in H446 and H446/CDDP
cells challenged by cisplatin explained, at least to some extent,
that mitochondrial dysfunction participated in the MDR of SCLC.
Acknowledgments
The present research was supported by the National
Natural Science Foundation of China (no. 81071933).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jun S, Dimyan M, Jones KD and Ladabaum U:
Obstipation as a paraneoplastic presentation of small cell lung
cancer: Case report and literature review. Neurogastroenterol
Motil. 17:16–22. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sørensen M, Pijls-Johannesma M and Felip
E; ESMO Guidelines Working Group: Small-cell lung cancer: ESMO
Clinical Practice Guidelines for diagnosis, treatment and
follow-up. Ann Oncol. 21(Suppl 5): v120–v125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu X, Zhuang Y and Zhang J: Study on the
molecular mechanism of multi-drug resistance in clinical isolates
of Mycobacterium tuberculosis. Zhonghua Jie He He Hu Xi Za Zhi.
20:332–335. 1997.In Chinese.
|
|
6
|
Murakami S: Molecular mechanism of
multi-drug resistance. Nihon Rinsho. 66:193–203. 2008.In Japanese.
PubMed/NCBI
|
|
7
|
Warburg O, Wind F and Negelein E: The
metabolism of tumors in the body. J Gen Physiol. 8:519–530. 1927.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Warburg O: On the origin of cancer cells.
Science. 3191:309–314. 1956. View Article : Google Scholar
|
|
9
|
Upadhyay M, Samal J, Kandpal M, Singh OV
and Vivekanandan P: The Warburg effect: Insights from the past
decade. Pharmacol Ther. 137:318–330. 2013. View Article : Google Scholar
|
|
10
|
Lee HC, Yin PH, Lin JC, Wu CC, Chen CY, Wu
CW, Chi CW, Tam TN and Wei YH: Mitochondrial genome instability and
mtDNA depletion in human cancers. Ann NY Acad Sci. 1042:109–122.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Compton S, Kim C, Griner NB, Potluri P,
Scheffler IE, Sen S, Jerry DJ, Schneider S and Yadava N:
Mitochondrial dysfunction impairs tumor suppressor p53
expression/function. J Biol Chem. 286:20297–20312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen JZ, Gokden N, Greene GF, Mukunyadzi P
and Kadlubar FF: Extensive somatic mitochondrial mutations in
primary prostate cancer using laser capture microdissection. Cancer
Res. 62:6470–6474. 2002.PubMed/NCBI
|
|
13
|
Heddi A, Faure-Vigny H, Wallace DC and
Stepien G: Coordinate expression of nuclear and mitochondrial genes
involved in energy production in carcinoma and oncocytoma. Biochim
Biophys Acta. 1316:203–209. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gochhait S, Bhatt A, Sharma S, Singh YP,
Gupta P and Bamezai RN: Concomitant presence of mutations in
mitochondrial genome and p53 in cancer development - a study in
north Indian sporadic breast and esophageal cancer patients. Int J
Cancer. 123:2580–2586. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vivekanandan P, Daniel H, Yeh MM and
Torbenson M: Mitochondrial mutations in hepatocellular carcinomas
and fibrolamellar carcinomas. Mod Pathol. 23:790–798. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Larman TC, DePalma SR, Hadjipanayis AG,
Protopopov A, Zhang J, Gabriel SB, Chin L, Seidman CE, Kucherlapati
R and Seidman JG; Cancer Genome Atlas Research Network: Spectrum of
somatic mitochondrial mutations in five cancers. Proc Natl Acad Sci
USA. 109:14087–14091. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guha M, Srinivasan S, Ruthel G, Kashina
AK, Carstens RP, Mendoza A, Khanna C, Van Winkle T and Avadhani NG:
Mitochondrial retrograde signaling induces epithelial-mesenchymal
transition and generates breast cancer stem cells. Oncogene.
33:5238–5250. 2014. View Article : Google Scholar
|
|
18
|
Imanishi H, Hattori K, Wada R, Ishikawa K,
Fukuda S, Takenaga K, Nakada K and Hayashi J: Mitochondrial DNA
mutations regulate metastasis of human breast cancer cells. PLoS
One. 6:e234012011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Andrews RM, Kubacka I, Chinnery PF,
Lightowlers RN, Turnbull DM and Howell N: Reanalysis and revision
of the Cambridge reference sequence for human mitochondrial DNA.
Nat Genet. 23:1471999. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ovadje P, Chatterjee S, Griffin C, Tran C,
Hamm C and Pandey S: Selective induction of apoptosis through
activation of caspase-8 in human leukemia cells (Jurkat) by
dandelion root extract. J Ethnopharmacol. 133:86–91. 2011.
View Article : Google Scholar
|
|
21
|
Wallace DC: Mitochondria and cancer. Nat
Rev Cancer. 12:685–698. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wallace DC: A mitochondrial bioenergetic
etiology of disease. J Clin Invest. 123:1405–1412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guaragnella N, Giannattasio S and Moro L:
Mitochondrial dysfunction in cancer chemoresistance. Biochem
Pharmacol. 92:62–72. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang G, Qu Y, Dang S, Yang Q, Shi B and
Hou P: Variable copy number of mitochondrial DNA (mtDNA) predicts
worse prognosis in advanced gastric cancer patients. Diagn Pathol.
8:1732013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dohi T and Altieri DC: Mitochondrial
dynamics of survivin and 'four dimensional' control of tumor cell
apoptosis. Cell Cycle. 4:21–23. 2005. View Article : Google Scholar
|
|
26
|
Strum SB, Adalsteinsson O, Black RR, Segal
D, Peress NL and Waldenfels J: Case report: Sodium dichloroacetate
(DCA) inhibition of the 'Warburg effect' in a human cancer patient:
Complete response in non-Hodgkin's lymphoma after disease
progression with rituximab-CHOP. J Bioenerg Biomembr. 45:307–315.
2013. View Article : Google Scholar
|
|
27
|
Cui J, Shi M, Xie D, Wei D, Jia Z, Zheng
S, Gao Y, Huang S and Xie K: FOXM1 promotes the Warburg effect and
pancreatic cancer progression via transactivation of LDHA
expression. Clin Cancer Res. 20:2595–2606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Naito A, Carcel-Trullols J, Xie CH, Evans
TT, Mizumachi T and Higuchi M: Induction of acquired resistance to
antiestrogen by reversible mitochondrial DNA depletion in breast
cancer cell line. Int J Cancer. 122:1506–1511. 2008. View Article : Google Scholar
|
|
29
|
Singh KK, Russell J, Sigala B, Zhang Y,
Williams J and Keshav KF: Mitochondrial DNA determines the cellular
response to cancer therapeutic agents. Oncogene. 18:6641–6646.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park SY, Chang I, Kim JY, Kang SW, Park
SH, Singh K and Lee MS: Resistance of mitochondrial DNA-depleted
cells against cell death: Role of mitochondrial superoxide
dismutase. J Biol Chem. 279:7512–7520. 2004. View Article : Google Scholar
|
|
31
|
Hoshida Y, Moriyama M, Otsuka M, Kato N,
Taniguchi H, Shiratori Y, Seki N and Omata M: Gene expressions
associated with chemosensitivity in human hepatoma cells.
Hepatogastroenterology. 54:489–492. 2007.PubMed/NCBI
|
|
32
|
Trachootham D, Alexandre J and Huang P:
Targeting cancer cells by ROS-mediated mechanisms: A radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Degli Esposti M: Mitochondria in
apoptosis: Past, present and future. Biochem Soc Trans. 32:493–495.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Salet C, Moreno G, Ricchelli F and
Bernardi P: Singlet oxygen produced by photodynamic action causes
inactivation of the mitochondrial permeability transition pore. J
Biol Chem. 272:21938–21943. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Du W, Hong J, Wang YC, Zhang YJ, Wang P,
Su WY, Lin YW, Lu R, Zou WP, Xiong H, et al: Inhibition of
JAK2/STAT3 signalling induces colorectal cancer cell apoptosis via
mitochondrial pathway. J Cell Mol Med. 16:1878–1888. 2012.
View Article : Google Scholar
|