Introduction
Gastric cancer is a common gastrointestinal tumor.
Its mortality rate ranks second among men and fourth among women
(1,2). At present, surgery is the primary
therapy for this disease and may achieve satisfactory results when
combined with chemotherapy. However, for advanced gastric cancer,
surgical therapy is not satisfactory.
Natural products are important resources for
anticancer chemical agents. Thus, a significant strategy for
developing anticancer drugs is to identify effective and active
ingredients from natural products.
β-asarone
(C12H16O3, MW 208.5) is the main
volatile oil of Chinese herb Rhizoma Acori Tatarinowii. It
exhibits therapeutic effects on many organs of the human body
displaying many biological activities. Numerous studies have
illustrated that β-asarone regulates the nervous system (3), kills parasites and bacteria (4) and prevents cholesterol synthesis
(5). Previous laboratory research
found that β-asarone significantly inhibited the growth of colon
cancer (6). However, there is no
study concerning its effects on gastric cancer cells and the
related mechanism. Accordingly, the present study aimed to
investigate the impact of β-asarone on human gastric cancer cells
and to explore the possible mechanism.
Materials and methods
Cell culture
Three types of cell lines, low-differentiated human
gastric cancer cell line BGC-823, moderately differentiated human
gastric cancer cell line SGC-7901 and highly differentiated human
gastric cancer cell line MKN-28, were obtained from the Type
Culture Collection, Chinese Academy of Sciences (Shanghai, China),
and were routinely cultured in RPMI-1640 medium containing 10%
bovine serum at 37°C with 5% CO2.
Drugs and reagents
β-asarone, methyl thiazolyl tetrazolium and β-actin
were purchased from Sigma-Aldrich Co. LLC. RPMI-1640 medium was
purchased from HyClone, USA, bovine serum was purchased from
Hangzhou Sijiqing Biological Engineering Materials Co., Ltd. and
0.25% trypsin was purchased from Biosharp Biological Technology
Co., Ltd. Annexin V/PI was purchased from KeyGen Biotech Co. Ltd.
and TRIzol and SYBR-Green were obtained from Life Science and
Technology Co. Real-time PCR reverse transcription kit was
purchased from Takara Co. Ltd. and antibodies against caspase-3,
-8, and -9, Bax, Bak, Bcl-xL, MMP-2, MMP-9, MMP-14, RECK,
E-cadherin, N-cadherin and β-actin were purchased from Cell
Signaling Co., USA. Bcl-2 was purchased from GeneTex, Inc. and NR2B
and GluR2 as the secondary antibodies were purchased from ZSGB-Bio
Co. All of the reagents were analytically pure.
MTT assay
The growth inhibitory effect of β-asarone was
measured by methyl thiazolyl tetrazolium (MTT) assay. Human gastric
cancer cells in a logarithm growth phase were inoculated into a
96-well plate at a density of 6×103/well. After the
cells adhered for 24 h, the medium in the plate was discarded, and
β-asarone was mixed with RPMI-1640 medium. The mixed solution with
β-asarone at a concentration of 0, 0.48, 0.24, 0.12, 0.06 or 0.03
mM were added into the 96-well plate and the cells were further
incubated at 37°C with 5% CO2 for 24 h. Afterwards, 5
µl MTT (5 mg/ml) was added into each well. Four hours later,
the dark blue formazan precipitates were dissolved, and 150
µl dimethylsulfoxide (DMSO) was added into each well. The
absorbance value was detected by an enzyme-linked immunosorbent
assay (ELISA) at a wavelength of 490 nm to calculate the inhibitory
rate of β-asarone for cancer growth. The assay was repeated three
times.
Flow cytometric analysis
The gastric cancer cells were treated with 0.12 and
0.24 mM β-asarone and 5-fluorouracil (5-FU) (50 µg/ml) under
the same conditions as the MTT assay. After treatment with
β-asarone for 24 h, the cells were collected, washed with cold
phosphate-buffered saline (PBS), and resuspended in binding buffer
at a density of 8×105/ml. Subsequently, the cells were
stained with 5 µl Annexin V and 5 µl PI for 15 min in
the absence of light. Apoptosis of the cells was detected by flow
cytometry within 1 h.
Fluorescent immunocytochemistry test
Three types of gastric cancer cells were plated into
a 6-well plate at a density of 3×105/well. After
adhering for 24 h, the cells were treated with β-asarone (0.12 and
0.24 mM) and 5-FU (50 µg/ml) for 24 h and washed with PBS
twice. To detect the nuclei, cells were incubated with Hoechst for
10 min, stained by Hoechst 33342 solution for 20 min and washed
with PBS three times. Afterwards, the nuclei were photographed
using a fluorescence microscope.
Electron microscopic analysis
After treatment with β-asarone (0.24 mM) for 24 h,
the cells were fixed in 2.5% glutaraldehyde, 4% paraformaldehyde,
post-fixed with 1% osmium tetroxide, dehydrated and in-layed by
epoxy resin. The cells were cut into 80-nm sections which were
stained with lead uranyl acetate and detected by a JEM-1010
electron microscope (JEOL, Japan) at 80 kV.
Transwell assay
Low-differentiated human gastric cancer BGC-823
cells with stronger invasive potential were inoculated in a culture
dish for 24 h. Cells (6×105) in media supplemented with
1% FBS were plated onto the upper chambers coated with Matrigel,
and 600 µl media supplemented with 10% FBS was added into
the lower chambers. After treatment with β-asarone (0.12 and 0.24
mM) or 50 µg/ml 5-FU for 24 h, the cells were fixed and
stained with 0.1% crystal violet. The non-migrated cells were wiped
off and the migrated cells were photographed.
Wound-healing assay
BGC-823 cells were cultured into a 24-well plate
(1×106/well). Once 100% confluence was observed, the
cell monolayer was scratched with a pipette tip and washed with PBS
twice. The cells were treated with β-asarone (0.12 and 0.24 mM) or
50 µg/ml 5-FU for 24 h. The width of the scratches was
photographed for analysis.
Cell matrix adhesion assay
BGC-823 cells were inoculated in a culture dish
until they adhered to the plate surface. β-asarone (0.12 and 0.24
mM) and 5-FU (50 µg/ml) were added and maintained for 24 h.
The cells were digested and centrifuged. The precipitated cell
suspension was resuspended with culture medium containing the
original medicine. Approximately 1×105 live cells were
seeded into each well of a 96-well plate coated with fibronectin.
The original medicated medium was discarded every 30 min, and the
cells were cultured with RPMI-1640 medium. Afterwards (120 min), 5
µl MTT (5 mg/ml) was added into each well. After 4 h of
cultivation, ELISA was used to read the absorption values at a
490-nm wavelength.
Assessment of mRNA expression
Having been incubated with β-asarone for 24 h, the
human gastric cancer cell lines were assessed by RT-PCR to
determine the effect of β-asarone on the mRNA expression of
apoptosis- and metastasis-associated genes. Total RNA from the
cells was isolated using TRIzol reagent. Subsequently, 1 µg
of the extracted total RNA was reverse-transcribed into
first-strand complementary DNA (cDNA) using the High-Capacity cDNA
Reverse Transcription kit according to the manufacturer's
instructions. They were then performed on 7500 Fast RT-PCR system,
using DNA-binding dye SYBR-Green. The ΔΔCt method was used for
qPCR. The sequences of the primers used to specifically amplify the
genes of interest are shown in Table
I.
 | Table ISequences of the primers used in the
RT-PCR amplifications. |
Table I
Sequences of the primers used in the
RT-PCR amplifications.
| Gene primer | Sequences
(5′-3′) | Length of PCR product
(bp) |
|---|
| Bax | F
TTTGCTTCAGGGTTTCATCC
R GCCACTCGGAAAAAGACCTC | 213 |
| Bak | F
ACGCTATGACTCAGAGTTCC
R CTTCGTACCACAAACTGGCC | 360 |
| Bcl-xL | F
ATGAACTCTTCCGGGATGG
R TGGATCCAAGGCTCTAGGTG | 166 |
| Bcl-2 | F
TCGCCCTGTGGATGACTGAG
R CAGAGTCTTCAGAGACAGCCAGGA | 143 |
| Survivin | F
TTCTCAAGGACCACCGCATC
R GCCAAGTCTGGCTCGTTCTC | 127 |
| N-cadherin | F
GTGCCATTAGCCAAGGGAATTCAGC
R GCGTTCCTGTTCCACTCATAGGAGG | 370 |
| E-cadherin | F
GGTGCTCTTCCAGGAACCTC
R CAGCAACGTGTTTCTGCATTTC | 196 |
| RECK | F
ATCTGTTCACCCTGGAGT
R TGTTGAAGTGTTTGCTTT | 96 |
| MMP-2 | F
CTCCCGGAAAAGATTGATG
R GGTGCTGGCTGAGTAGAT | 96 |
| MMP-9 | F
TCTATGGTCCTCGCCCTGAA
R CATCGTCCACCGGACTCAAA | 219 |
| MMP-14 | F
ATCTGCCTCTGCCTCACCTA
R AAGCCCCATCCAAGGCTAAC | 126 |
| β-actin | F
GGCCAACCGCGAGAAGAT
R CGTCACCGGAGTCCATCA | 134 |
Western blot analysis
After 24 h of incubation with β-asarone, the gastric
cancer cell lines were assessed by western blotting to detect the
activity of the apoptosis- and metastasis-associated proteins. The
cell lysates were separated by 12% SDS-PAGE, and transferred to a
polyvinylidene fluoride membrane, blocked with 5% skimmed milk for
1 h, and incubated with the primary antibodies (1:1,000 dilution)
overnight at 4°C. Having been incubated with horseradish
peroxidase-conjugated secondary antibody, ECL liquid was obtained
for lighting and banding analysis.
Statistical analysis
Data are presented as the mean values ± SD.
Differences between the mean values for individual groups were
assessed using one-way ANOVA followed by Duncan's multiple range
tests. Differences were considered significant at P<0.05. SPSS
13.0 was applied for statistical analysis.
Results
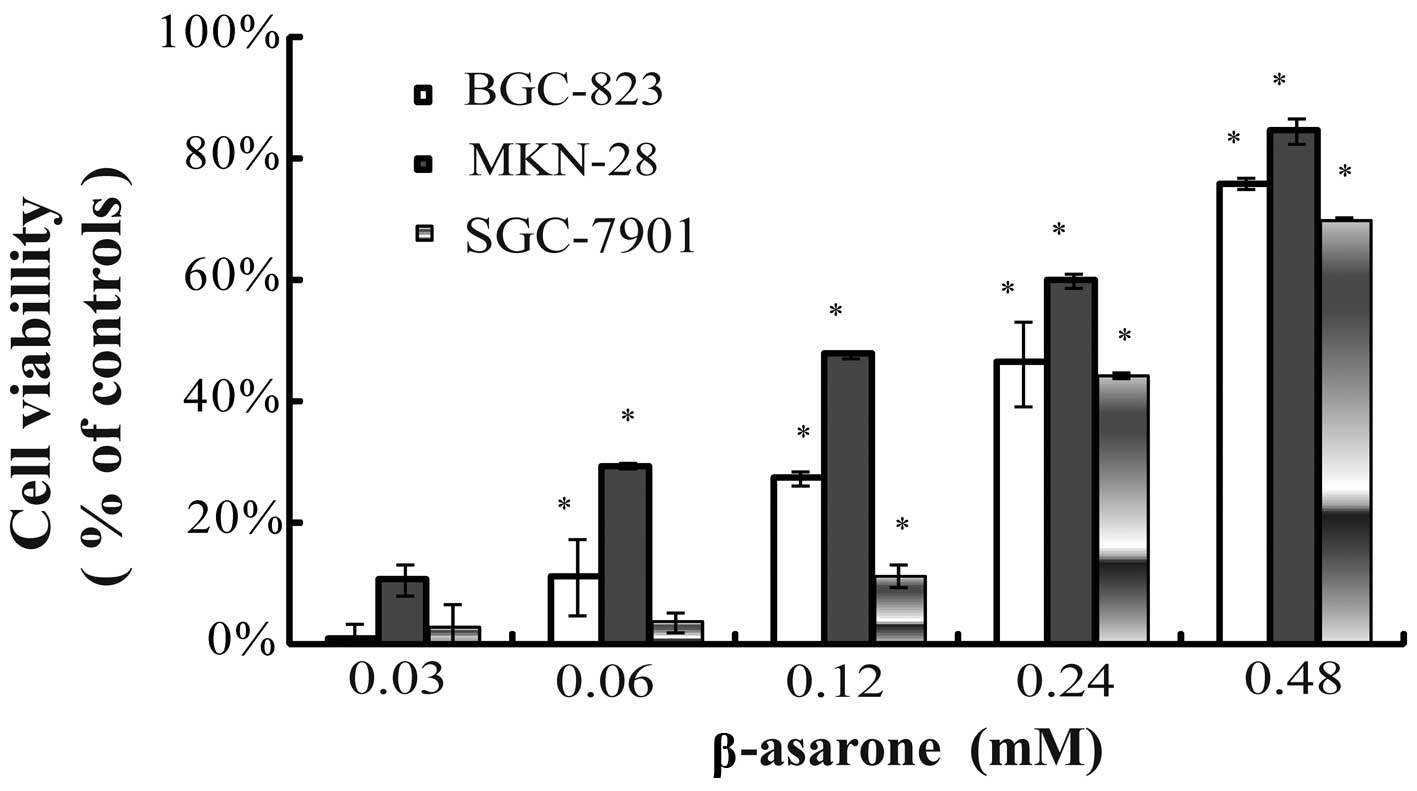
β-asarone inhibits the proliferation of
gastric cancer cells
Human gastric cancer cells were treated with 0,
0.48, 0.24, 0.12, 0.06 and 0.03 mM β-asarone, respectively, for up
to 24 h. β-asarone exhibited an inhibitory effect on the
proliferation of the cells in a dose-dependent manner. The
inhibitory rate of β-asarone at the concentration of 0.48 mM for
BGC-823, MKN-28 and SGC-7901 cells within 24 h was 75.79±3.12,
84.42±0.38 and 69.85±1.91%, respectively (Fig. 1).
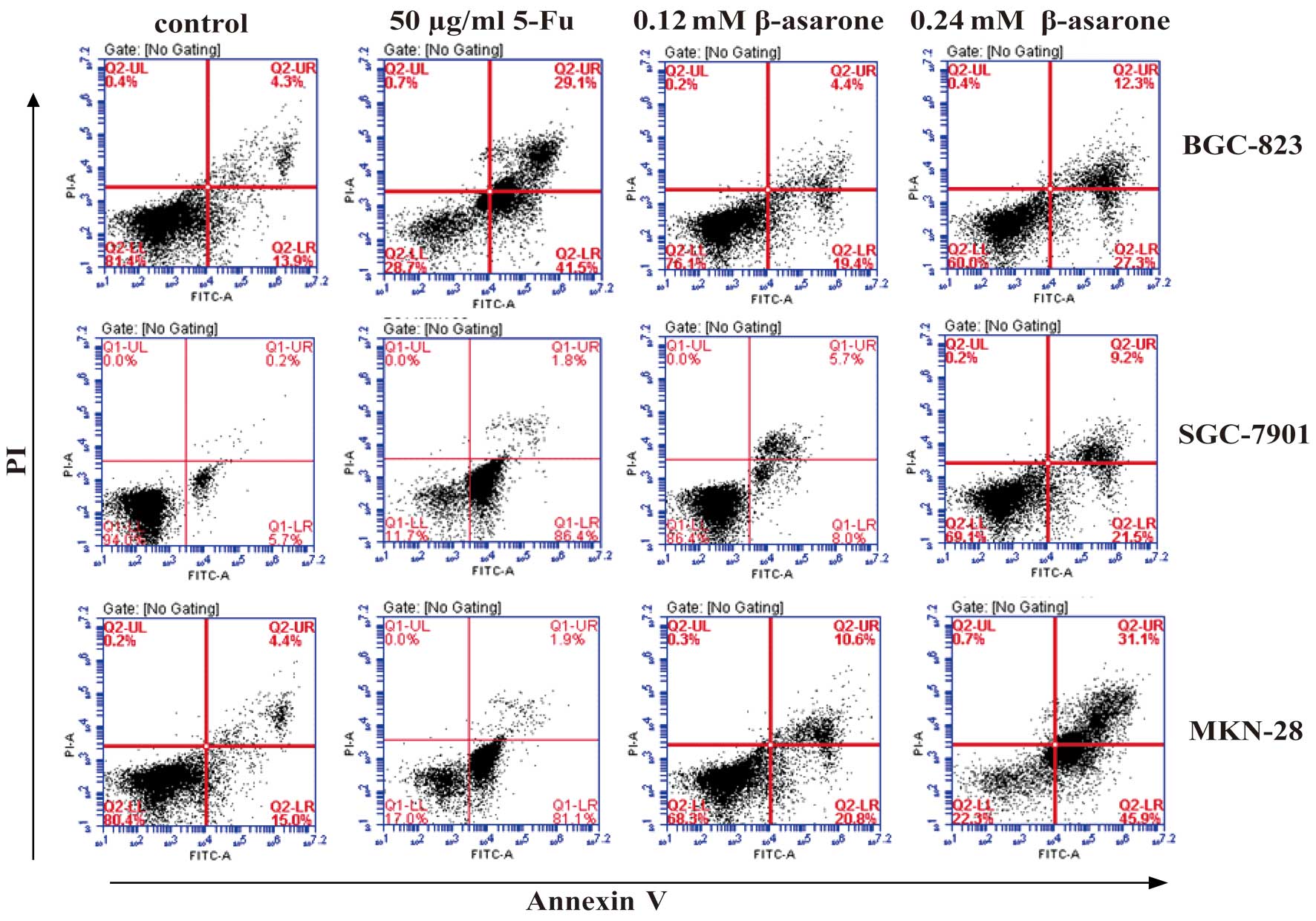
β-asarone induces the apoptosis of
gastric cancer cells
Flow cytometric analysis detected the early
apoptosis of the human gastric cancer cells 24 h after β-asarone
treatment. Following treatment at concentrations of 0.12 and 0.24
mM β-asarone, the early apoptosis rate of the BGC-823, SGC-7901 and
MKN-28 cells rose from 19.4 to 27.3%, 8 to 21.5% and 20.8 to 45.9%,
respectively. Meanwhile, the apoptosis rate of the positive control
group was 41.5% (BGC-823), 86.4% (SGC-7901) and 81.1% (MKN-28)
(Fig. 2).
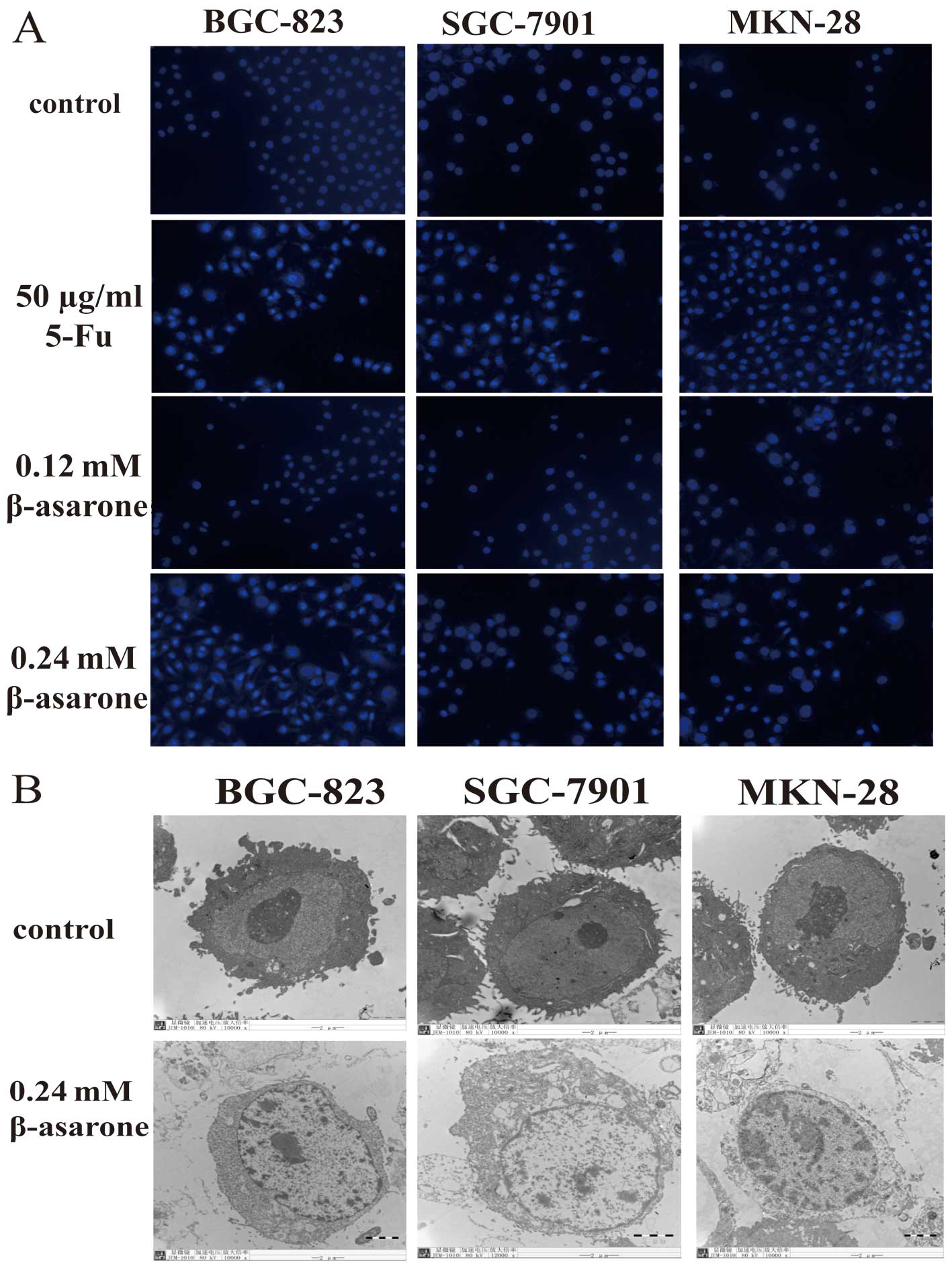
β-asarone alters the structure of gastric
cancer cells
In order to observe the structural changes in the
gastric cancer cells following treatment with β-asarone, both
fluorescence microscopy and transmission electron microscopy were
utilized. The cells stained with Hochest 33342 were observed under
a fluorescence microscope (magnification, x200). The nuclei of the
cells treated by β-asarone were fragmented and shrunken which
indicated apoptosis (Fig. 3A).
Transmission electron microscopy (magnification,
x8000) found nuclear atrophy, cytoplasmic vacuolization,
endoplasmic reticulum expansion and chromatin edge gathering in the
cells, which were also indicative of cell apoptosis (Fig. 3B). Thus, β-asarone significantly
induced apoptosis in the gastric cancer cells.
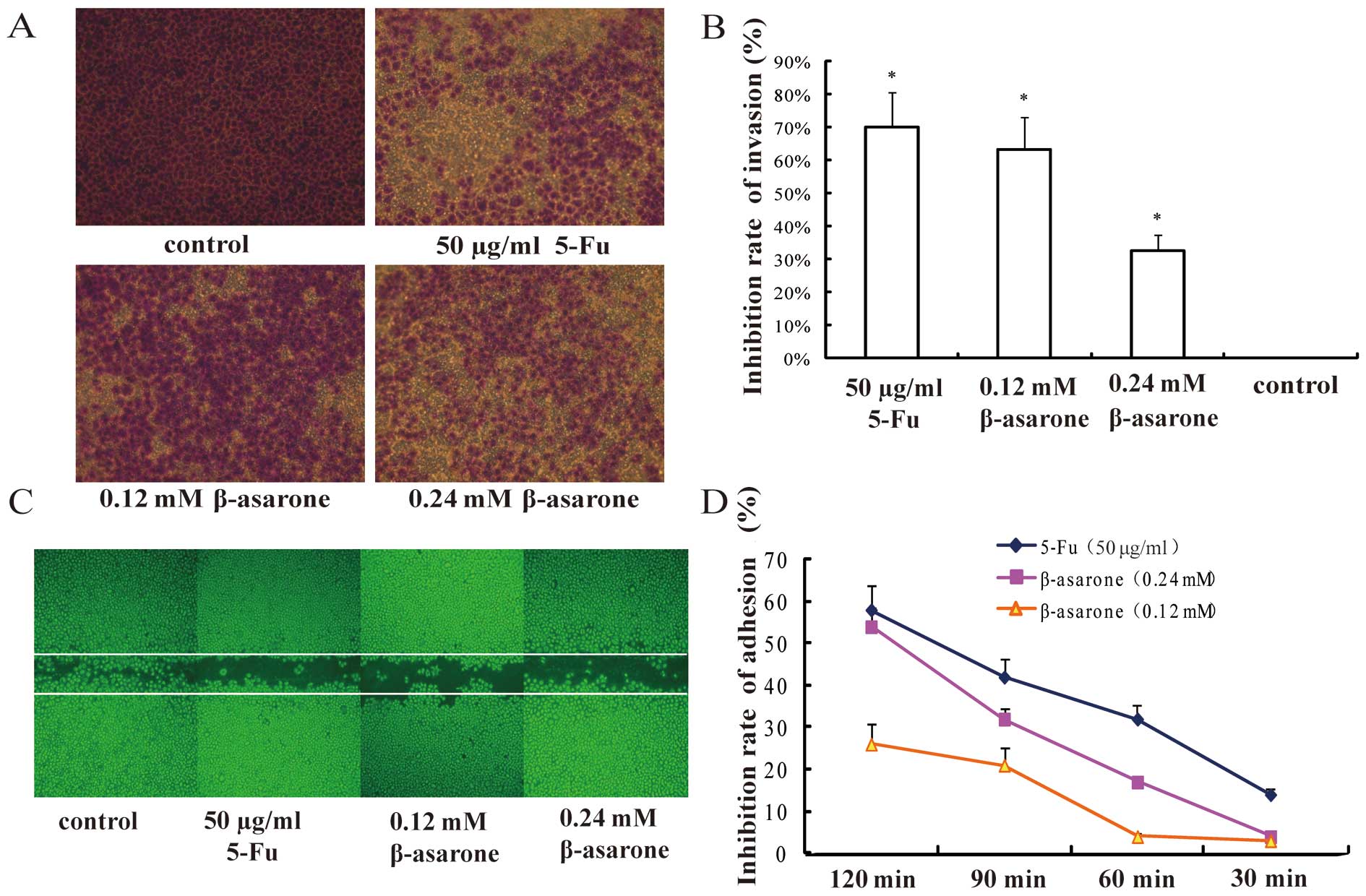
β-asarone prohibits the migration and
invasion of BGC-823 cells
In order to observe the effect of β-asarone on the
migration and invasion of gastric cancer cells, low-differentiated
BGC-823 cells were chosen as the target. Both Transwell (Fig. 4A and B) and wound-healing assays
(Fig. 4C) revealed that β-asarone
inhibited the migration of BGC-823 cells in a
concentration-dependent manner. β-asarone efficiently suppressed
the cell invasion in a dose-dependent manner as determined by the
Matrigel-coated Transwell assay. Furthermore, compared with the
control group, the area of the treated cells in the 6-well plate
which migrated into the whole wound area was markedly decreased,
suggesting that β-asarone also suppressed the migration of BGC-823
cells in a dose-dependent manner.
β-asarone inhibits the adhesive ability
of the BGC-823 cells
Cell matrix adhesion assay was performed to evaluate
the effect of β-asarone on gastric cancer cell adhesion. We found
that the adhesion rate at each time point was lower than that of
the control group. Further data demonstrated that the inhibitory
rate of adhesion increased as the concentration of β-asarone and
the treatment time increased (Fig.
4D). Therefore, β-asarone significantly prevented gastric
cancer cells from adhering to fibronectin, and the inhibition rate
was exhibited in a dose- and time-dependent manner.
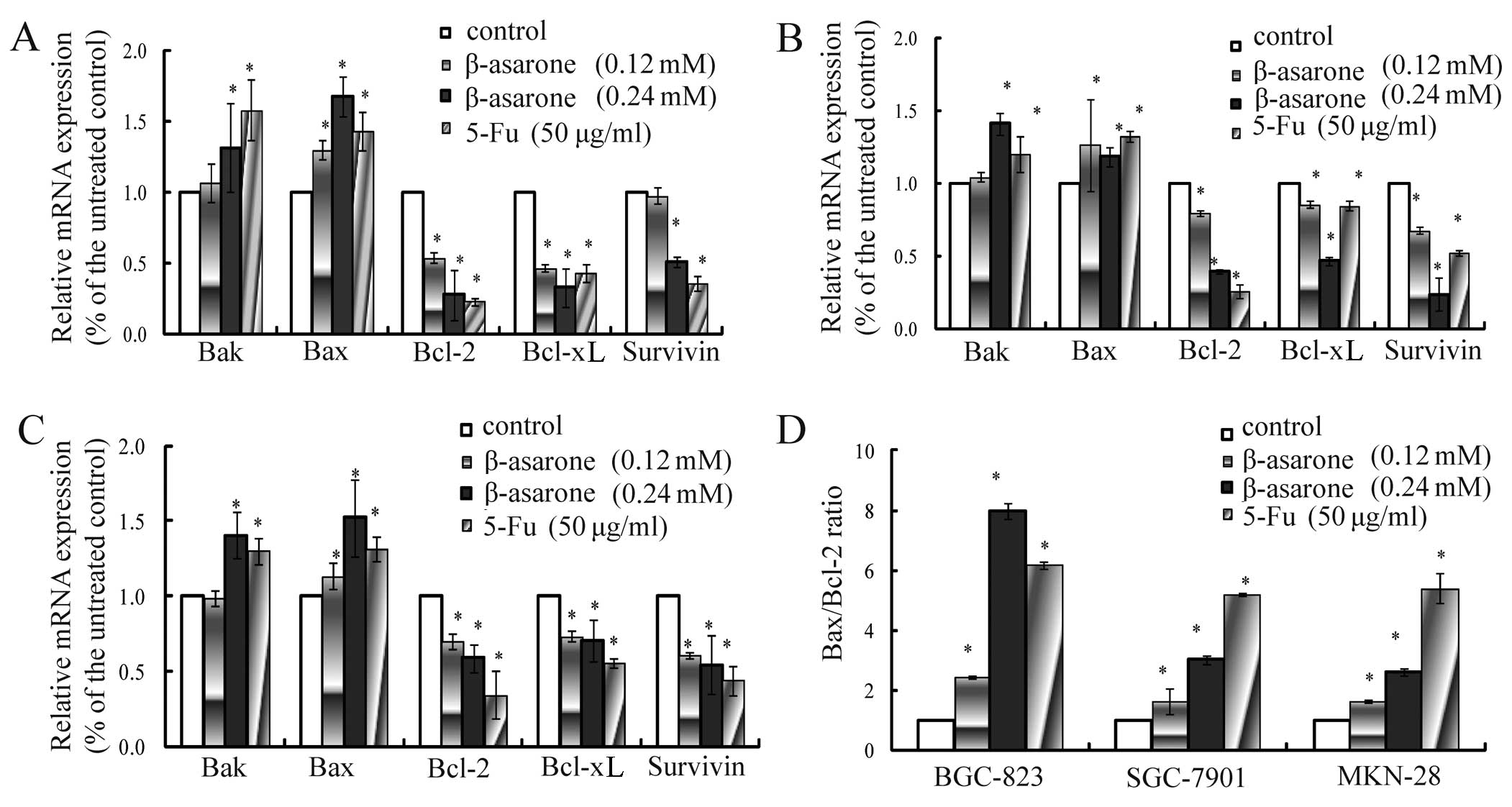
Changes in the mRNA and protein
expression of apoptosis-related genes
In order to further study the mechanism of β-asarone
action on the apoptosis of gastric cancer cells, RT-PCR and western
blotting were applied to analyze changes in the mRNA and protein
levels. RT-PCR showed that after 24 h of incubation of BGC-823
(Fig. 5A), SGC-7901 (Fig. 5B), MNK-28 (Fig. 5C) cells with β-asarone (0.12 and
0.24 mM), the mRNA expression levels of Bax and Bak were increased,
the mRNA expression levels of Bcl-2, Bcl-xL and survivin were
reduced and the Bax/Bcl-2 ratio was increased in a dose-dependent
manner (Fig. 5D). In addition,
western blotting revealed a gradual increase in the expression of
Bax, caspase-3, caspase-8 and caspase-9 and a reduction in the
expression of Bcl-2 and Bcl-xL in a dose-dependent manner (Fig. 6A). These results indicated that
β-asarone induced the apoptosis of gastric cancer cells by
regulating Bax expression and downregulating the expression of
Bcl-2 and Bcl-xL as well as activating the caspase-cascade
response.
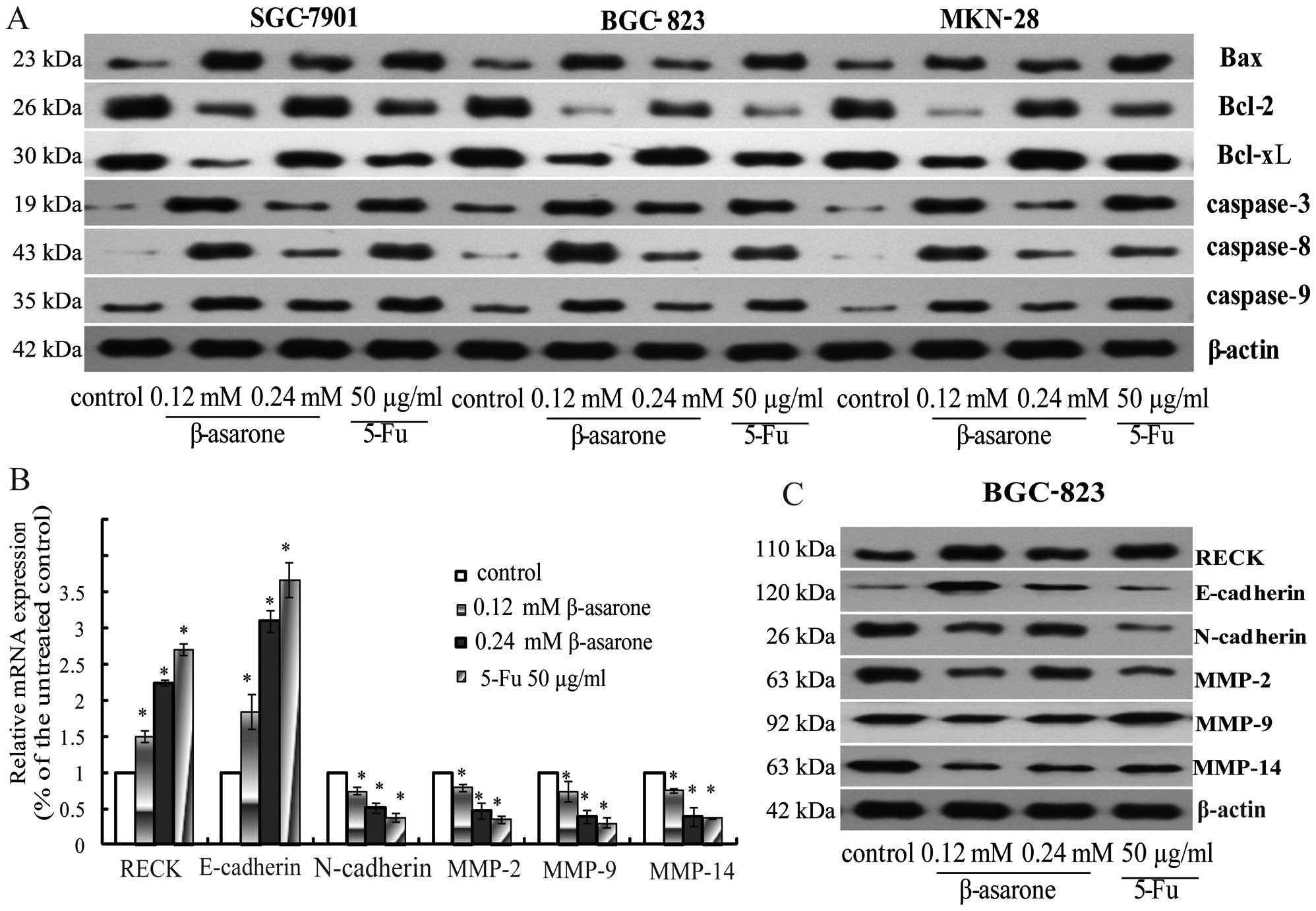 | Figure 6(A) After BGC-823, SGC-7901 and MKN-28
cells were treated with β-asarone (0.12 and 0.24 mM) for 24 h, the
levels of apoptosis-related proteins (Bax, Bcl-2, Bcl-xL,
caspase-3, -8 and -9) were determined by western blotting. β-actin
was taken as a loading control and 5-FU (50 µg/ml) as a
positive control. (B) RT-PCR analysis of invasion-related
molecules. After BGC-823 cells were treated with β-asarone (0.12
and 0.24 mM) for 24 h, the mRNA levels of invasion and EMT-related
genes (RECK, E-cadherin, N-cadherin, MMP-2, MMP-9 and MMP-14) were
detected. β-actin was used as an internal control.
*P<0.05. (C) After BGC-823 cells were treated with
β-asarone (0.12 and 0.24 mM) for 24 h, the expression changes in
the proteins closely associated with invasion and adhesion (RECK,
E-cadherin, N-cadherin, MMP-2, MMP-9 and MMP-14) were detected.
β-actin was used as an internal control and 5-FU (50 µg/ml)
as a positive control. |
Changes in the mRNA and protein
expression of invasion-related genes
In order to investigate the mechanism of β-asarone
in the invasion, migration and adhesion of gastric cancer cells, we
assessed changes in the mRNA and protein expression levels of
invasion-related genes. The results of both RT-PCR (Fig. 6B) and western blotting (Fig. 6C) revealed that the expression of
RECK was significantly increased, while the expression levels of
MMP-2, MMP-9 and MMP-14 were decreased in the cells following
treatment with β-asarone as the dose increased. β-asarone was found
to markedly prevent gastric cancer cells from adhering to
fibronectin. Therefore, the expression of E-cadherin and N-cadherin
which are closely associated with adhesion was further detected. We
found that the BGC-823 cells exhibited a dose-dependent increase in
E-cadherin and a decrease in N-cadherin following treatment with
β-asarone.
Discussion
β-asarone is one of the main bioactive constituents
of the essential oil of Rhizoma Acori Tatarinowii. The
inhibitory effect of β-asarone on gastric cancer cells has been
poorly reported, and the detailed underlying anticancer mechanism
remains unclear. Here, for the first time, we investigated the
potential abilities of β-asarone to resist gastric cancer growth.
The present study found that β-asarone efficiently suppressed
gastric cancer cell viability, induced cell apoptosis and inhibited
invasive, migratory and adhesive abilities.
Apoptosis refers to programmed cell death. A dynamic
balance between apoptosis and proliferation is crucial to
maintaining cell stability and preventing carcinogenesis. At
present, the choice of anticancer drugs is often based on
biochemical properties of apoptosis. Thus, once inhibition of the
gastric cancer cell proliferation by β-asarone was confirmed, its
effect on the apoptosis of gastric cancer cells was examined. The
study found obvious morphological apoptotic changes in the gastric
cancer cells following treatment with β-asarone; meanwhile, flow
cytometry also revealed that obvious apoptosis occurred in the
gastric cancer cell lines 24 h after β-asarone treatment.
Caspases play a crucial role in cell apoptosis. They
are a type of protease hydrolysates, usually existing in the form
of procaspases. According to different initiative factors and
pathways, cell apoptosis is divided into two categories: exogenous
apoptosis and endogenous apoptosis. In regards to exogenous
apoptosis, the interaction between death ligands and receptors
causes intracellular structural change to attract pro-caspase-8 to
cut and activate itself. Activated caspase-8 then activates
downstream caspases such as caspase-3, caspase-6 and caspase-7
which induces cell apoptosis by shearing cytoplasm and nuclear
substrates (7). Endogenous
apoptosis referring to a condition when the cell is stimulated,
promotes the release of apoptotic proteins, such as cytochrome
c, from the mitochondria to the cytoplasm recruiting
pro-caspase-9 to construct protein complexes and apoptotic bodies.
Meanwhile, activated caspase-9 induces activation of caspase-3,
caspase-6 and caspase-7 belonging to downstream caspases and
finally intiating the apoptosis process (8–10).
Therefore, it is the active caspase family that is the main
executor of the apoptotic process. In the present study, western
blot results showed that β-asarone increased the expression of
caspase-3, caspase-8 and caspase-9 proteins. The inducing effect of
β-asarone on gastric cancer cells was probably due to endogenous
and exogenous apoptosis.
In the process of cell apoptosis, Bcl-2 is another
crucial gene family. The Bcl-2 family mainly participates in
apoptosis regulation through coordination with mitochondria
(11). They are divided into two
types based on different functions: anti-apoptotic protein,
represented by Bcl-2 and Bcl-xL and pro-apoptotic proteins, Bax and
Bak. Bcl-2 regulates transport across the reticulum and prevents
the release of cytochrome c, thus the cascade reaction
process of apoptosis is inhibited. However, Bax is a pro-apoptotic
gene, and the increasing expression of Bax improves antagonistic
Bcl-2 and Bcl-xL to promote apoptosis (12). The balance between Bcl-2 and Bax
determines the start of cell apoptosis (13). Once the ratio of Bax/Bcl-2
increases, a series of complicated cascade reactions and the signal
transmissions in the cell induce the start of apoptosis which leads
to cell death (14,15). The present experiment showed that at
24 h after β-asarone treatment in the gastric cancer cells, Bcl-2
and Bcl-xL were downregulated, while Bax and Bak were upregulated
and the Bax/Bcl-2 ratio increased in a concentration-dependent
manner. This was possibly an aspect of the molecular mechanisms of
apoptosis in gastric cancer cells induced by β-asarone.
In addition to apoptosis inhibition, local invasion
and distant metastasis are the most important biological features
of malignant tumors. To elucidate the effect of β-asarone on the
motility of gastric cancer cells, their migration and invasion were
detected by wound-healing migration and chamber invasion assays,
respectively. β-asarone was found to have a significant effect on
the inhibition of cell migration and invasion.
Epithelial-mesenchymal transition (EMT) plays a
pivotal role in tumor metastasis (16). The transition of cells from an
epithelial phenotype into a mesenchymal phenotype endows tumor
cells with greater invasiveness and motility by resulting in the
loss of polarity of the cells, damaging the cell connection and
reorganizing the intracellular actin cytoskeleton (17,18).
Typical molecular markers of EMT include a decrease
in the expression of epithelial markers such as E-cadherin and an
increase in the expression of mesenchymal markers such as
N-cadherin (19). In the present
study, β-asarone was found to reverse the process of EMT by
lowering N-cadherin and raising E-cadherin levels.
Moreover, the expression and secretion of several
ECM-degrading proteolytic proteases such as matrix
metal-loproteinases (MMPs), play an important role in promoting the
process of metastasis (20). Of the
MMP family, MMP-2, MMP-9 and MMP-14 are important for degrading the
natural barrier basement membrane (21,22).
RECK, as a critical MMP suppressor, potently inhibits tumor
angiogenesis and metastasis (23).
Recent studies have demonstrated that RECK inhibited at least three
types of MMPs, including MMP-2, MMP-9 and MMP-14, to prevent tumosr
from transmembrane metastasis (24,25).
The present study found that the expression of MMP-2, MMP-9 and
MMP-14 in the gastric cancer cells was decreased, while RECK was
increased in a concentration-dependent manner after treatment with
β-asarone. This may be an important mechanism of the inhibition by
β-asarone of gastric cancer cell invasion and metastasis. However,
as a complex molecular process, invasion and metastasis of cancer
cells need further study.
Collectively, the present study demonstrated that
β-asarone inhibited the gastric cancer cell growth by upregulating
the expression of caspase-3, caspase-8 and caspase-9, Bax, Bak,
RECK, E-caderin and downregulating the expression of Bcl-2, Bcl-xL,
MMP-2, MMP-9, MMP-14 and N-caderin. Additionally, β-asarone
inhibited cancer cell growth by inducing apoptosis and prohibiting
migration. The anticancer effect of β-asarone thus warrants further
in vivo testing, and its specific effective concentration
and its mechanism in detail require further investigation.
Acknowledgments
This study was supported by the Foundation of the
Priority Academic Program Development of Jiangsu Higher Education
Institutions (PAPD), by the National Natural Science Foundation of
China (nos. 81202954 and 81473605).
References
|
1
|
Gomceli I, Demiriz B and Tez M: Gastric
carcinogenesis. World J Gastroenterol. 18:5164–5170.
2012.PubMed/NCBI
|
|
2
|
Dong H, Gao Z, Rong H, Jin M and Zhang X:
β-asarone reverses chronic unpredictable mild stress-induced
depression-like behavior and promotes hippocampal neurogenesis in
rats. Molecules. 19:5634–5649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang YX, Chen YT, Zhou XJ, Hong CL, Li CY
and Guo JY: Beta-asarone, a major component of Acorus tatarinowii
Schott, attenuates focal cerebral ischemia induced by middle
cerebral artery occlusion in rats. BMC Complement Altern Med.
13:2362013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu XC, Zhou LG, Liu ZL and Du SS:
Identification of insecticidal constituents of the essential oil of
Acorus calamus rhizomes against Liposcelis bostrychophila Badonnel.
Molecules. 18:5684–5696. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee SH, Kim KY, Ryu SY, Yoon Y, Hahm DH,
Kang SA, Cho SH, Lim JS, Moon EY, Yoon SR, et al: Asarone inhibits
adipogenesis and stimulates lipolysis in 3T3-L1 adipocytes. Cell
Mol Biol (Noisy-le-grand). 56(Suppl): OL1215–OL1222. 2010.
|
|
6
|
Zou X, Liu SL, Zhou JY, Wu J, Ling BF and
Wang RP: Beta-asarone induces LoVo colon cancer cell apoptosis by
up-regulation of caspases through a mitochondrial pathway in vitro
and in vivo. Asian Pac J Cancer Prev. 13:5291–5298. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Garg S, Narula J and Chandrashekhar Y:
Apoptosis and heart failure: Clinical relevance and therapeutic
target. J Mol Cell Cardiol. 38:73–79. 2005. View Article : Google Scholar
|
|
8
|
Ghavami S, Hashemi M, Ande SR, Yeganeh B,
Xiao W, Eshraghi M, Bus CJ, Kadkhoda K, Wiechec E, Halayko AJ, et
al: Apoptosis and cancer: Mutations within caspase genes. J Med
Genet. 46:497–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Majors BS, Betenbaugh MJ and Chiang GG:
Links between metabolism and apoptosis in mammalian cells:
Applications for anti-apoptosis engineering. Metab Eng. 9:317–326.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 7:719–727. 2005. View Article : Google Scholar
|
|
11
|
Autret A and Martin SJ: Emerging role for
members of the Bcl-2 family in mitochondrial morphogenesis. Mol
Cell. 36:355–363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brooks C and Dong Z: Regulation of
mitochondrial morphological dynamics during apoptosis by Bcl-2
family proteins: A key in Bak? Cell Cycle. 6:3043–3047. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ghoneum M, Matsuura M, Braga M and
Gollapudi S: S. cerevisiae induces apoptosis in human metastatic
breast cancer cells by altering intracellular Ca2+ and
the ratio of Bax and Bcl-2. Int J Oncol. 33:533–539.
2008.PubMed/NCBI
|
|
14
|
Salakou S, Kardamakis D, Tsamandas AC,
Zolota V, Apostolakis E, Tzelepi V, Papathanasopoulos P, Bonikos
DS, Papapetropoulos T, Petsas T, et al: Increased Bax/Bcl-2 ratio
up-regulates caspase-3 and increases apoptosis in the thymus of
patients with myasthenia gravis. In Vivo. 21:123–132.
2007.PubMed/NCBI
|
|
15
|
Zhang H and Rosdahl I: Bcl-xL and bcl-2
proteins in melanoma progression and UVB-induced apoptosis. Int J
Oncol. 28:661–666. 2006.PubMed/NCBI
|
|
16
|
Acloque H, Thiery JP and Nieto MA: The
physiology and pathology of the EMT. Meeting on the
epithelial-mesenchymal transition. EMBO Rep. 9:322–326. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wells A, Chao YL, Grahovac J, Wu Q and
Lauffenburger DA: Epithelial and mesenchymal phenotypic switchings
modulate cell motility in metastasis. Front Biosci. 16:815–837.
2011. View Article : Google Scholar
|
|
19
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tsareva SA, Moriggl R, Corvinus FM,
Wiederanders B, Schütz A, Kovacic B and Friedrich K: Signal
transducer and activator of transcription 3 activation promotes
invasive growth of colon carcinomas through matrix
metalloproteinase induction. Neoplasia. 9:279–291. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Seiler R, Thalmann GN and Fleischmann A:
MMP-2 and MMP-9 in lymph-node-positive bladder cancer. J Clin
Pathol. 64:1078–1082. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Daniele A, Zito AF, Giannelli G, Divella
R, Asselti M, Mazzocca A, Paradiso A and Quaranta M: Expression of
metal-loproteinases MMP-2 and MMP-9 in sentinel lymph node and
serum of patients with metastatic and non-metastatic breast cancer.
Anticancer Res. 30:3521–3527. 2010.PubMed/NCBI
|
|
23
|
Clark JC, Thomas DM, Choong PF and Dass
CR: RECK - a newly discovered inhibitor of metastasis with
prognostic significance in multiple forms of cancer. Cancer
Metastasis Rev. 26:675–683. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Omura A, Matsuzaki T, Mio K, Ogura T,
Yamamoto M, Fujita A, Okawa K, Kitayama H, Takahashi C, Sato C, et
al: RECK forms cowbell-shaped dimers and inhibits matrix
metalloproteinase-catalyzed cleavage of fibronectin. J Biol Chem.
284:3461–3469. 2009. View Article : Google Scholar
|
|
25
|
Takemoto N, Tada M, Hida Y, Asano T, Cheng
S, Kuramae T, Hamada J, Miyamoto M, Kondo S and Moriuchi T: Low
expression of reversion-inducing cysteine-rich protein with Kazal
motifs (RECK) indicates a shorter survival after resection in
patients with adenocarcinoma of the lung. Lung Cancer. 58:376–383.
2007. View Article : Google Scholar : PubMed/NCBI
|