Introduction
Lung cancer is the second leading cause of
cancer-related mortality worldwide, and more than 1.6 million cases
are diagnosed every year (1).
Tobacco smoking and exposure to environmental carcinogens have been
found to be the major risk factors in the development of this
disease (2). Most lung cancer
patients are diagnosed in an advance stage with an overall survival
of five years (1). Despite
considerable advances in our knowledge and experience in the
treatment of lung cancer patients, our capacity to effectively
fight and treat this disease is still limited (3). Treatment of lung cancer patients only
manages to reduce the burden of the primary lesion but rarely is
effective to completely eradicate the tumour cells which in turn
leads to relapse and fatality (2).
These facts and limitations highlight the need for the greater
understanding of the cellular and molecular events that drive
tumourigenesis. Thus, therapeutic strategies can be tailored for
better treatment efficacies.
Lung cancer can be classically subdivided into small
cell lung cancer (SCLC) and three types of non-small cell lung
cancer (NSCLC), which include squamous cell carcinoma,
adenocarcinoma and large cell carcinoma (4). The existence of several lung
epithelial progenitor cells that initiate diverse lung epithelial
subtypes and functions is thought to be responsible for this tumour
variety (5). The cancer stem cell
(CSC) theory suggests that mutations in the progenitor cells lead
to the formation of CSCs resulting in cellular hierarchy and clonal
expansion within a tumour (6,7). CSCs
are known to share common properties with normal epithelial stem
cells including self-renewal, proliferation and capacity for
lineage differentiation (6,8,9).
However, CSCs may not necessarily be homogeneous in general as they
often evolve subsequently by accumulating additional mutations,
which in turn results in a complex clonal heterogeneity (10). CSCs are also believed to be the
driving source of the malignant phenotype (resistance to
chemotherapy, distant metastasis and relapse) in the primary tumour
(11). Therefore, therapies that
target chemoresistant tumour cells and distant tumour metastasis,
which are characteristic of CSCs, may be an effective and yet
powerful treatment strategy to eradicate the primary tumour
(12,13).
Curcumin (diferuloymethane), a naturally occurring
polyphenol extract from the rhizome Curcuma longa (Tumeric),
possesses biological activities against many types of tumours
(14–18). Curcumin modulates numerous target
proteins including transcription factors, receptors, kinases,
cytokines, enzymes and growth factors (19). Curcumin was found to downregulate
the expression of several drugresistance proteins such as
ATP-binding cassette (ABC) drug transporters, P-glycoproteins and
multi-drug resistant (MDR) proteins, which resulted in the
sensitivity of tumour cells to chemotherapy (20–22).
Pre-clinical studies have shown that curcumin acts synergistically
with conventional chemotherapeutic drugs to eradicate resistant
lung cancer cell lines (20,23,24).
Similar findings with different tumours have also been reported
in vitro as well as in experimental animal models (25–28).
In a human breast cancer xenograft model, administration of
curcumin markedly decreased the metastasis of breast tumour cells
to the lung and suppressed the expression of vascular endothelial
growth factor (VEGF), matrix metalloproteinase-9 (MMP-9) and
intercellular adhesion molecule-1, which reduced the invasive and
metastatic phenotype of the tumour cells (29). Furthermore, curcumin has been found
to be safe when administered at ≤10 g/day in humans, thus reducing
the difficulty of reaching an effective dose due to dose-limiting
toxicity (30).
The antitumour efficacy of curcumin has also been
studied recently, either alone or in combination with other
antitumour agents on stem-like cells isolated from several tumours
using in vitro CSC assays (sphere formation, enzyme
activity, side population and cell-surface marker expression) as
well as in vivo animal models. In breast cancer models, 5
µM of curcumin treatment reduced mammosphere formation by
50%, while complete elimination of mammospheres and reduction in
aldehyde dehydrogenase 1 (ALDH) enzyme activity (a selective marker
noted in most CSCs) were noted as the concentration of curcumin was
increased to 10 µM (31,32). A
study conducted by Fong et al using an in vivo glioma
model reported that daily treatment of 5 µM curcumin
resulted in the reduction of the side population as analysed by
flow cytometry (33). Furthermore,
curcumin also reduced the expression of CD133 and nestin (neural
stem/progenitor markers) indicating the differentiation of gliomal
CSCs that eventually led to deregulation of the self-renewal
capability of CSCs (34).
CD133 was recently reported as a promising CSC
marker noted in prostate cancer (35–37),
brain tumours (38–41), colon cancer (42–44)
and hepatic carcinoma (45–48). However, in the context of lung
cancer stem cells, the utility of the marker appears limited due to
the low expression detected in most lung cancer samples (49,50),
and the discrepancy of the findings in regards to CD133 in most
studies have questioned the prognostic value of this marker in
clinical application (50–52). It is therefore important to identify
markers that are commonly expressed in most lung cancer samples;
hence it can be applied in a larger fraction of lung tumour
samples. Other studies established that CSC markers such as CD326
(EpCAM) and CD166 are more robust compared to CD133 as these
markers are highly detected in most NSCLC cancer samples (53,54).
Furthermore, CD166+/Lin− markers were also
found to be prominent in NSCLC patients suggesting the
applicability of CD166 as a selective marker for CSCs in NSCLC
(54). We previously identified and
characterised, based on in vivo tumourigenicity, a novel
CD166+/EpCAM+ CSC subpopulation isolated from
NSCLC cell lines, and showed that this subpopulation has
self-renewal capacity, higher mobility, resistance to apoptosis and
exhibits mesenchymal lineage differentiation based on gene
expression profiling (55). In the
present study, we investigated the anticancer effects of curcumin
(either alone or in combination with cisplatin) as a drug
sensitiser and metastatic inhibitor on both unsorted and sorted
(CD166 and EpCAM) cancer stem-like populations derived from NSCLC
cell lines. This study will provide further insight into the
potential of using curcumin as a sensitiser of CSCs to
cisplatin-induced cell death.
Materials and methods
All of the cell lines were purchased from the
American Type Culture Collection (ATCC, Manassas, VA, USA). The
research protocol was approved by our Institutional Review Boards
(Medical Research Ethics Committee/MREC, Ministry of Health,
Malaysia).
Cell culture
NSCLC cell lines, A549 (ATCC® CRL-185™)
and H2170 (ATCC® CRL-5928™) were cultured in RPMI-1640
(Invitrogen, Carlsbad, CA, USA) medium containing 10% fetal bovine
serum (FBS), 100 IU/ml penicillin and 100 µg/ml streptomycin
(all purchased from ATCC) and grown at 37°C in a humidified 5%
CO2 atmosphere. Human lung fibroblast (IMR-90) cells
were cultured in MEM-α (1x)-Glutamax medium containing 10% FBS, 100
IU/ml penicillin and 100 µg/ml streptomycin (ATCC). Cells
were maintained in T75 tissue culture flasks, and the medium was
changed three times a week. Confluent cells were harvested by
washing in phosphate-buffered saline (PBS) followed by
trypsinisation (0.25% in EDTA) for subculturing. All of the cell
lines were purchased from ATCC, and culture reagents were purchased
from Gibco-Life Technologies (Grand Island, NY, USA) unless
otherwise stated.
Sorting of
CD166+/EpCAM+ and
CD166−/EpCAM− NSCLC cell populations
The NSCLC cell lines (A549 and H2170) were harvested
upon incubation with 0.25% trypsin (Life Technologies, Foster City,
CA, USA) and washed with phosphate-buffered solution with 2% FBS.
The CD166-PE and EpCAM-FITC (BD Biosciences, San Jose, CA, USA)
antibodies were used for CSC identification by flow cytometry.
Briefly, cells were trypsinised, counted by a haemocytometer and
transferred to 75-mm polystyrene round-bottom test tubes (BD
Falcon, NJ, USA) at a cell concentration of 1×106
cells/ml and subsequently stained with 10 µl of antibodies
in the dark at 4°C. The cells were then washed and filtered through
a 40-µm cell strainer to obtain a single-cell suspension
before sorting. The expression of cancer stem cell markers (CD166
and EpCAM) was analysed and sorted using FACSAria III (BD,
Biosciences). Gating used for the sorting of
CD166+/EpCAM+ (Q2) and
CD166−/EpCAM− (Q3) NSCLC cell lines is
depicted in Fig. 3.
Spheroid assay and self-renewal
capacity
Sorted lung tumour cells (1.0×103
cells/ml) were suspended in serum-free medium containing DMEM F12
(Gibco) supplemented with 10 ng/ml fibroblast growth factor (bFGF),
1% of B27, 20 ng/ml of EGF, 1% antibiotic-antimycotic, (all
purchased from Life Technologies) and seeded in an 96-well
ultra-low attachment (ULA) dish. Spheroid formation was assessed by
light microscopy after 20 days of culture. Self-renewal
capabilities were also evaluated by monitoring single cells using
the live cell analyser (JuLI™ Br; NanoEnTek, CA, USA).
Preparation of curcumin and cisplatin
stock
Curcumin (Sigma-Aldrich, St. Louis, MO, USA) was
dissolved in 1 ml DMSO to make a stock solution of 10 mM. The
curcumin stock was then diluted in complete RPMI-1640 medium to
provide a substock and final working concentrations. Cisplatin
(Sigma-Aldrich) was prepared as a 10 mM stock in 0.9% sodium
chloride (NaCl) and was diluted in complete RPMI-1640 medium to
provide a substock. The solution was filtered through a
0.22-µm membrane, aliquoted and stored at −20°C until
further use.
Inhibitory concentration
(IC50) of single treatments (curcumin and/or cisplatin)
in the NSCLC cell lines
IC50 values for the single treatment with
either curcumin and cisplatin of NSCLC cell lines were assessed by
the MTS [3-(4,5-dimethylthiazol-2-yl)-2H-tetrazolium, inner salt]
assay purchased from Promega (Madison, WI, USA). Tumour cells were
plated at a density of 1×104 cells/well in 96-well
plates and incubated overnight in humidified air with 5%
CO2 at 37°C. NSCLC cells were then treated with a
working concentration of curcumin (10, 20, 30 and 40 µM) and
cisplatin (5, 10, 15, 20 and 25 µM) for 48 h. After a 48-h
incubation, 15 µl of MTS solution was added to each well and
incubated for another 4 h. Solubilisation solution (100 µl)
was later added to the cells, and the absorbance at 570 nm was
measured using Odyssey® SA Imaging System (Li-Cor,
Lincoln, NE, USA), using wells without cells as the blank. Cell
viability was calculated according to the following formula: Cell
viability (%) = cells (sample)/cells (control) × 100 and
IC50 was calculated using log formula.
IC50 of curcumin sensitisation
prior to cisplatin treatment in the NSCLC cell lines
In order to evaluate the efficacy of curcumin
sensitisation prior to cisplatin treatment in NSCLC cell lines,
both A549 and H2170 cells were initially sensitised/cultured with
different doses of curcumin (10, 20, 30 and 40 µM) for 24 h,
followed by low dose cisplatin (<3 µM) for another 24 h.
Briefly, the tumour cells were seeded (1.0×105
cells/well) in 6-well plates and sensitised with different doses of
curcumin for 24 h. On the following day, the cells were harvested
and seeded again in 96-well plates (1.0×104 cells/well)
with medium containing cisplatin (low dose) for another 24 h. At
the end of the experiment, 15 µl of MTS solution was added
to each well and incubated for another 4 h. Solubilisation solution
(100 µl) was later added to the cells, and the absorbance at
570 nm was measured using Odyssey SA Imaging System, using wells
without cells as the blank.
Toxicity of curcumin and cisplatin in the
human lung fibroblast (IMR-90) cell line
The IC50 values of both curcumin and
cisplatin in the A549 and H2170 cells were tested on IMR-90 cells
to evaluate the toxic effect of curcumin and cisplatin on normal
cells. IMR-90 cells were seeded overnight in 96-well plates at a
density of 1.0×104 cells/well in 100 µl complete
MEM-α. Subsequently, 100 µl of either curcumin and/or
cisplatin (concentration based on IC50 of A549 and
H2170) was added to the cells and incubated for 48 h. The viability
of the IMR-90 cells was assessed by adding 10 µl of Presto
Blue (BD Pharmingen, Franklin Lakes, NJ, USA) to each well and
incubated for 2 h before the absorbance was measured at 570 nm.
Apoptosis assay
The apoptosis assay was conducted using the Annexin
V/propidium iodide (PI) apoptosis kit purchased from BD Pharmingen.
In brief, 9.0×105 cells/well of sorted and unsorted
NSCLC cells were seeded into 6-well plates and incubated overnight.
Direct combination (synergistic effects) of both curcumin and
cisplatin on the NSCLC cell lines was performed by incubation of
the cells in medium containing the single treatment (cisplatin or
curcumin) and combination of both using the IC50 doses
for 48 h. Indirect combination (sensitising effects) of curcumin
was performed by incubating the NSCLC cell lines with curcumin
(IC50 value) for 24 h, followed by incubation with low
dose cisplatin (3 µM) for another 24 h. After treatments for
48 h (synergistic and sensitisation), both NSCLC cell lines were
harvested by trysinisation and collected by centrifugation. The
cell pellet was suspended in 100 µl of 1X Annexin V binding
buffer (Becton Dickinson BD) and 1 µl of Annexin V-FITC was
added. Antibody incubation was performed at 4°C for 20 min, and 1
µl of PI was later added before FACS acquisition. Stained
cells were subjected to flow cytometric analysis using a
FACSCalibur instrument (Becton Dickinson BD), and a total of 10,000
events were acquired and analyzed using Cell Quest software (Becton
Dickinson BD).
Scratch-wound/migration assay
Briefly, sorted and unsorted NSCLC cells were seeded
at a density of ~3–4×105 cells/well in complete medium
and grown overnight to a 90% confluent monolayer. The next day, the
cells were treated with colcemide (10 µg/ml) for 2 h for
cell synchronisation. After incubation, a scratch wound was
inflicted using a sterile 200-µl pipette tip and gentle
washing was carried out twice using PBS to remove debris. Cells
were then incubated with 2 ml of media containing both single
treatments (cisplatin and curcumin) and/or the combination of both
for another 48 h. The concentration of curcumin and cisplatin
(single treatments and/or combination) used for the assay was based
on the IC50 values evaluated on both A549 and H2170
cells. Images of migrated cells (five fields of each triplicate
well) were captured using relief contrast microscopy at ×40
magnification (Olympus IX 71; Olympus, Tokyo, Japan) and analysed
for 48 h. The numbers of cells that migrated into the wound area
were evaluated using the formula: Percentage of migrated cells =
[initial scratch (0 h) − final scratch (48 h)]/initial (0 h) ×
100.
Post-treatment effects on CSC marker
expression (CD326 and CD166) in NSCLC cells analysed by FACS
NSCLC cells (A549 and H2170) were seeded in 6-well
plates at a density of 9.0×105 cells/well and incubated
overnight. After incubation, the cells were treated with
IC50 values of curcumin and cisplatin, respectively for
48 h. Cells were then harvested and washed with ice-cold PBS.
Antibodies (CD326 and CD166) (10 µl) were added to each tube
and incubated for another 20 min in the dark at 4°C. The cells were
then suspended in ice-cold PBS supplemented with 2% FBS, and a
total of 10,000 events were acquired and analysed using Cell Quest
software (Becton Dickinson BD).
Quantitative real time-polymerase chain
reaction (RT-qPCR)
Initially, total RNA was extracted and evaluated for
purity as previously described. Transcriptor First Strand cDNA
synthesis kit (Roche Applied Science, Nonnenwald, Penzberg,
Germany) was used to synthesise cDNA according to the protocols
recommended by the manufacturer. Quantitative RT-PCR (qRT-PCR) was
performed using the Light Cycler 480 (Roche, Mannheim, Germany), on
sorted lung tumour cells subsequent to treatment either by single
agent (cisplatin or curcumin) or direct combination of both
(synergistic effects) based on the IC50 values. The
qRT-PCR reaction was prepared using SYBR 1 Master Mix (Roche
Applied Science, Penzberg, Germany) and primers as stated in
Table I. PCR conditions were set
under the following cycle conditions: pre-denaturation for 4 min at
95°C followed by 40 cycles consisting of denaturation at 95°C for
15 sec, annealing at 60°C for 30 sec and extension at 72°C for 30
sec followed by dissociation curve. The basic relative gene
expression (RQ) was calculated using the ΔΔCt formula and the
efficiency (E) of primer binding equal to 2.
 | Table IHuman primer sequences used for
qRT-PCR. |
Table I
Human primer sequences used for
qRT-PCR.
| Gene | Accession | Sense primer | Antisense
primer | Product size
(bp) |
|---|
| Apaf 1 | NM_013229.2 |
CACGTTCAAAGGTGGCTGAT |
TGGTCAACTGCAAGGACCAT | 214 |
| Cytochrome
c | NM_018947.5 |
GGAGGCAAGCATAAGACTGG |
GTCTGCCCTTTCTCCCTTCT | 267 |
| Caspase-9 | XM_005246014.1 |
TGTGGTGGTCATCCTCTCTCA |
GTCACTGGGGGTAGGCAAACT | 331 |
| p21 | NM_000389.4 |
CTCAGAGGAGGCGCCATG |
GGGCGGATTAGGGCTTCC | 517 |
| Cyclin D1 | XM_006718653.1 |
CGGAGGACAACAAACAGATC |
GGGTGTGCAAGCCAGGTCCA | 350 |
| GAPDH | NM_001289746.1 |
TGAAGGTCGGAGTCAACGGATT |
CATGTGGGCCATGAGGTCCACCAC | 530 |
Statistical analysis
All data are expressed as the mean ± standard
deviation (SD) of three independent experiments. Comparison between
two groups was performed using the two-tailed t-test. P-values of
<0.01 were considered to indicate statistically significant
differences.
Results
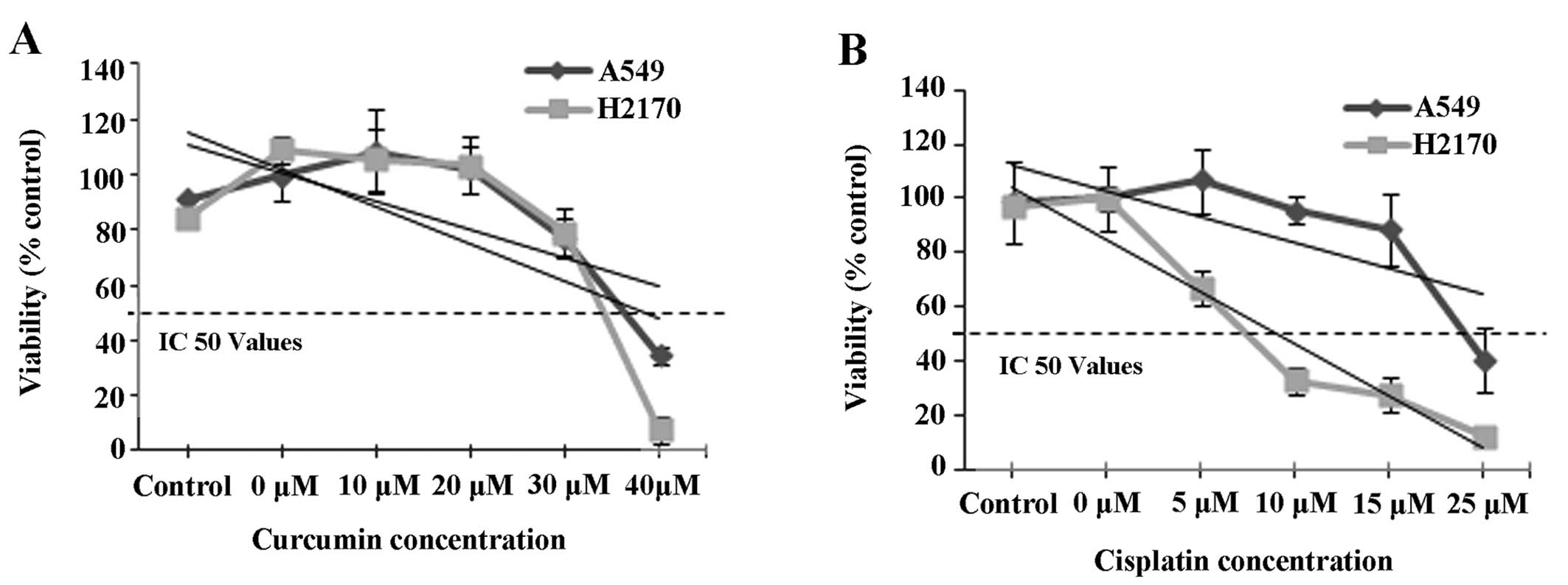
The IC50 values of curcumin
and cisplatin for both A549 and H2170 cell lines
The IC50 values of A549 and H2170 cells
treated with curcumin and cisplatin were assessed by MTS assay at
48 h. The results showed that exposure of NSCLC cell lines (A549
and H2170) to a range of curcumin (≤40 µM) and cisplatin
(≤25 µM) concentrations resulted in IC50 values
of 41 and 30 µM and 33 and 7 µM, respectively
(Fig. 1A and B). Furthermore, we
noted that the IC50 values for both A549 and H2170 cells
to curcumin were almost equal. However, the IC50 value
of cisplatin in the H2170 cells was markedly lower compared to that
for the A549 cells indicating higher sensitivity of H2170 cells to
cisplatin-induced inhibition. Based on the IC50 value
indicated, the combination of 41 µM curcumin and 30
µM cisplatin was selected for A549 cells and 33 µM of
curcumin and 7 µM of cisplatin were selected for H2170 cells
for further downstream study (Table
II).
| Table IIIC50 values were
determined by proliferation assays as specified in Materials and
methods. |
Table II
IC50 values were
determined by proliferation assays as specified in Materials and
methods.
| NSCLC cell
lines | IC50
values (µM) for the treatments
|
|---|
| Curcumin | Cisplatin |
|---|
| A549 | 40±9.3 | 30±5.0 |
| H2170 | 30±8.8 | 7±0.8 |
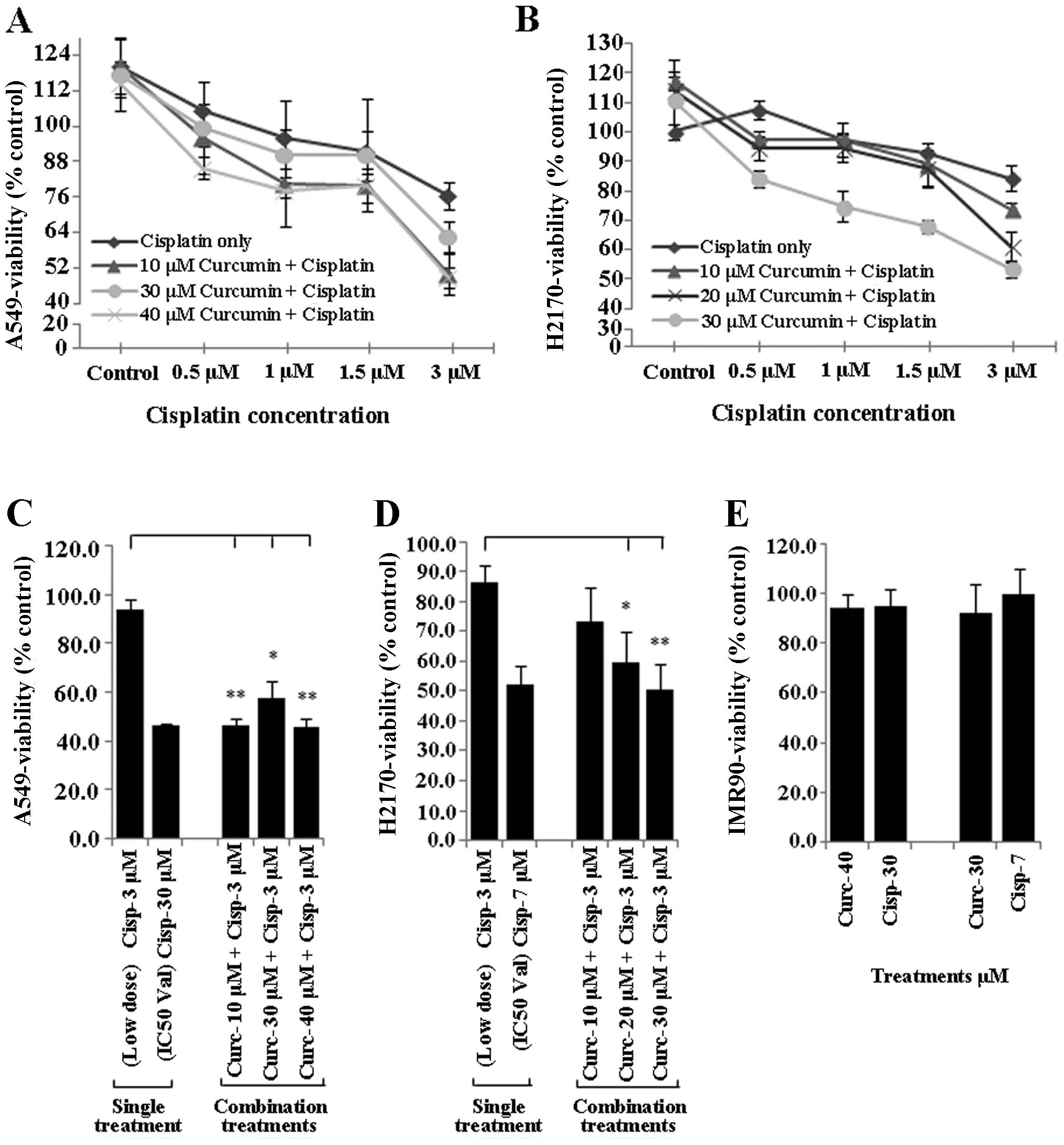
Curcumin sensitisation enhances the
tumour growth inhibitory effect of low dose cisplatin
To determine whether curcumin sensitises NSCLC cell
lines to the tumour inhibitory effect of low dose cisplatin (≤3
µM), NSCLC cells (A549 and H2170) were incubated overnight
with different doses of curcumin (10, 20, 30 and 40 µM),
harvested and seeded again with low dose cisplatin (≤3 µM)
for another 24 h. Treated NSCLC cells were subsequently evaluated
for cell viability using the MTS assay. Treatment of the A549 and
H2170 cells with curcumin (10–40 µM) markedly enhanced the
sensitivity of both NSCLC cell lines to cisplatin (Fig. 2A and B).
Treatment of both A549 and H2170 cells with 3
µM cisplatin alone (low dose) was found to be ineffective to
induce inhibition of growth in the NSCLC cell lines (tumour
viability ~80%). However, this treatment became highly effective
similar to the IC50 concentration (tumour viability
~50%) when both A549 and H2170 were initially sensitised to
different concentrations of curcumin (10–40 µM) (Fig. 2C and D). Moreover, exposure of
IMR-90 cells to the IC50 values of curcumin and
cisplatin in both NSCLC cell lines did not induce toxicity to the
cells, as the percentage of viability was higher than 90% (Fig. 2E).
Isolation of
CD166+/EpCAM+ and
CD166−/EpCAM− subpopulations from the NSCLC
cell lines
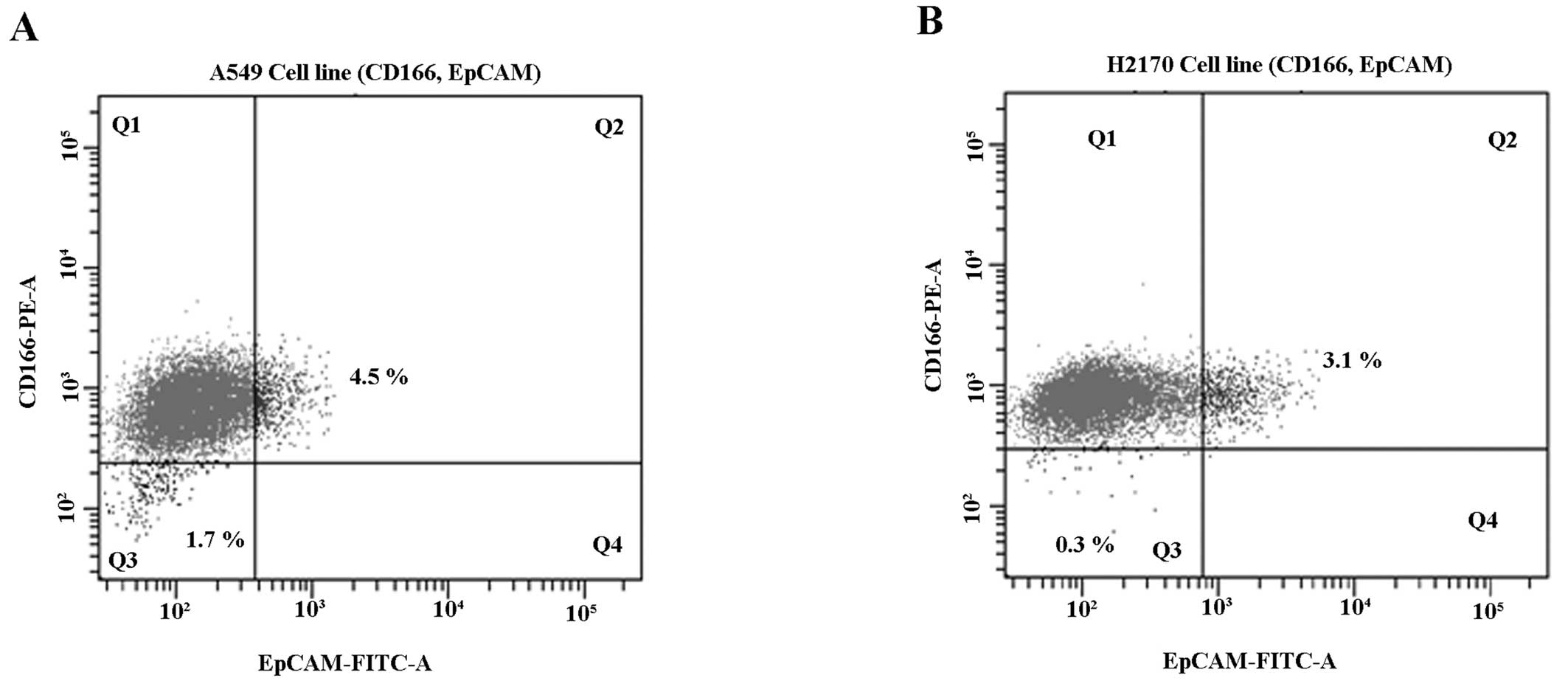
The expression of CSC markers (CD166 and EpCAM) in
the NSCLC cell lines was studied and we found a small population of
NSCLC cells (A549 and H2170) that showed positivity for CD166 and
EpCAM (CD166+/EpCAM+) (Fig. 3). The NSCLC cells showed consistent
double-positive expression of CD166+/EpCAM+
ranging from 3.0 to 4.5% (Fig. 3).
This was consistent with the characteristics of CSCs in a tumour
population indicating that the expression of CSC markers should be
within ~4% of the total population (56). Moreover, the double-negative
(CD166−/EpCAM−) population was much lower in
the NSCLC cell lines with 1.7 and 0.3% in both the A549 and H2170
cells, respectively (Fig. 3A and
B). The CD166+/EpCAM+ and
CD166−/EpCAM− populations were sorted from
the A549 and H2170 cells into a 15-ml tube containing complete
medium and transferred to a T75 flask for expansion and further
downstream study.
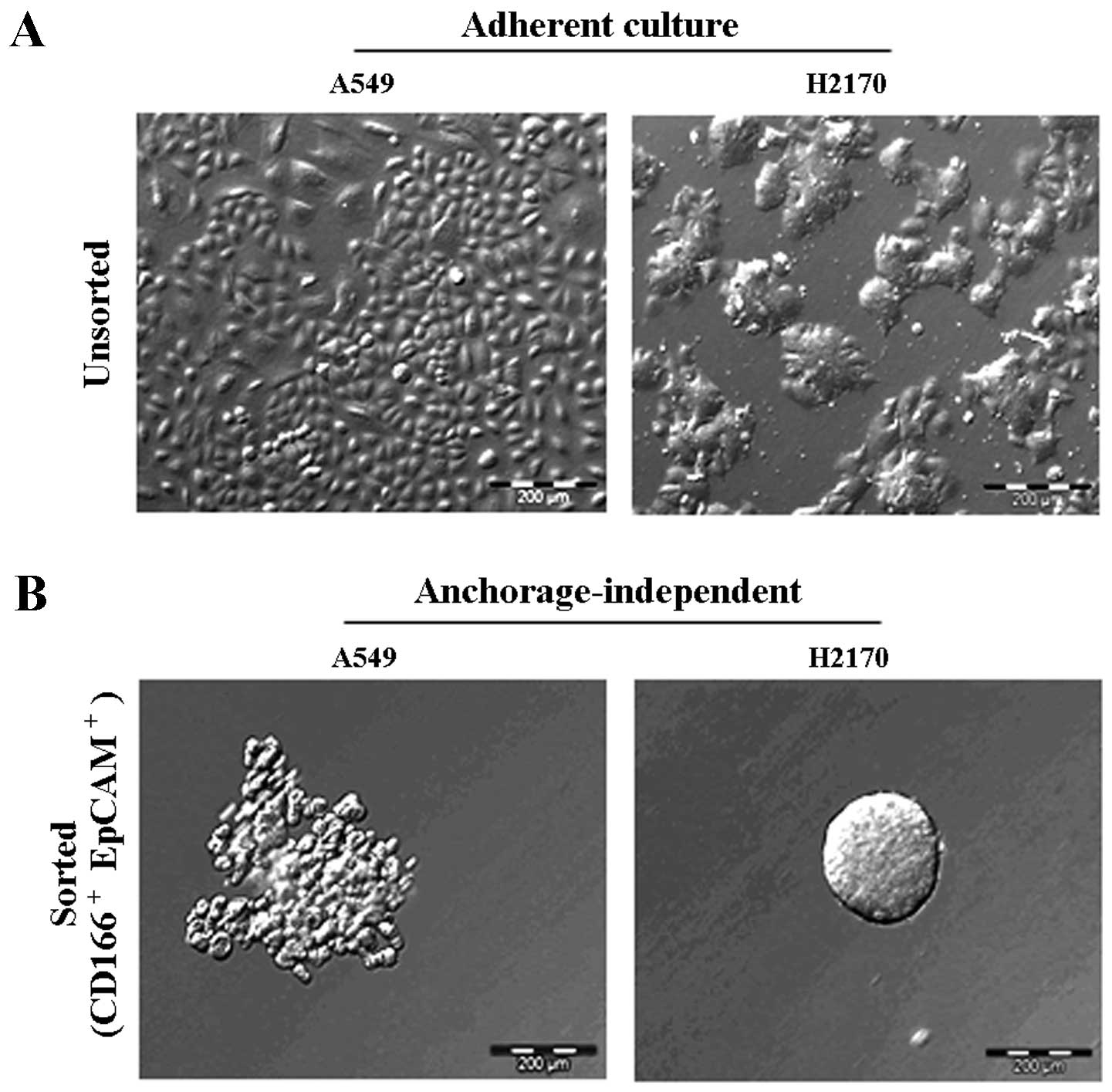
Tumour sphere formation and self-renewal
capacity of the CD166+/EpCAM+
subpopulation
The ability of the double-positive
(CD166+/EpCAM+) subpopulation sorted from the
NSCLC cell lines to form three-dimensional spheres in serum-free
medium containing stem cell growth factors (EGF and bFGF) on
non-adherent plates was examined. The sorted NSCLC cells grew as
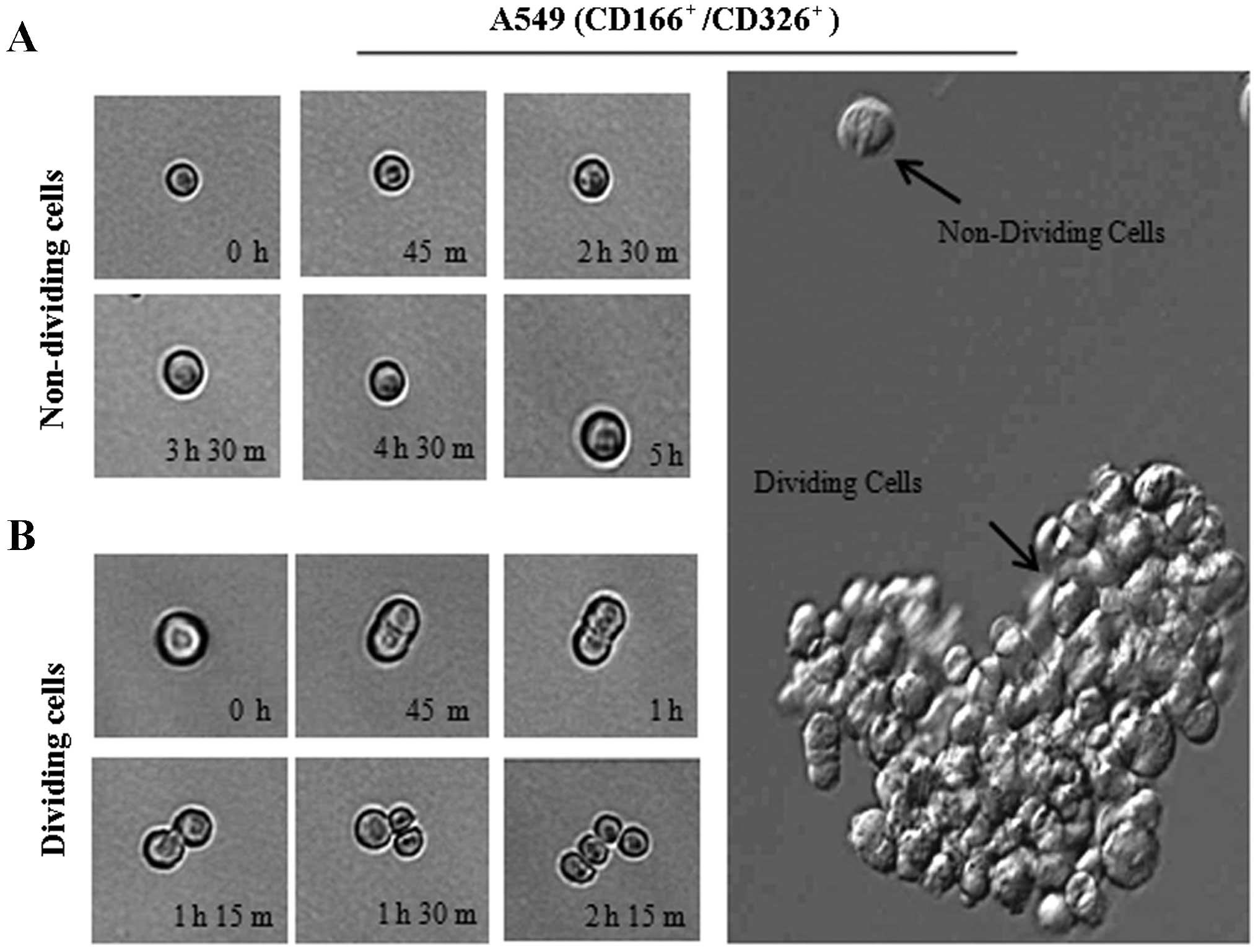
anchorage-independent spheres after 14 days of culture (Fig. 4B). We observed that the isolated
CD166+/EpCAM+ subpopulations of both A549 and
H2170 cells were able to form tumour spheres with an average size
ranging from 50 to 200 µm in diameter (Fig. 4B). We also noted that the
CD166+/EpCAM+ subpopulation isolated from
A549 cells had the ability for self-renewal and produced daughter
cells, which is an important characteristic of CSCs (Fig. 5B). However, there were cells
observed as non-dividing (Fig. 5A),
which were a dormant phenotype of CSCs.
Combination of curcumin and cisplatin
enriches the CD166+/EpCAM+ CSC
subpopulation
In order to study the combination effects of
curcumin and cisplatin on the regulation of CSC subpopulations, the
expression of two combination markers (CD166 and EpCAM) which were
previously described as markers of lung cancer CSCs, were evaluated
in the NSCLC cell lines (A549 and H2170) after treatment with
either a single agent of curcumin or cisplatin, or a combination of
both. Treatment of A549 and H2170 cells with the combination of
curcumin and cisplatin led to an average increase of ~10 and ~6%
expression of the double-positive
(CD166+/EpCAM+) CSC subpopulation,
respectively, as compared to both curcumin and cisplatin treatment
alone (Table III). Combination
treatment only led to ~2% increase and a slight reduction (~0.5%)
in the CD166−/EpCAM− subpopulation noted in
the A549 and H2170 cells, correspondingly (Table III), suggesting that the
combination of both curcumin and cisplatin synergistically acted to
enrich the double-positive (CD166+/EpCAM+)
subpopulation as compared to each single treatment alone.
| Table IIIThe percentage of subpopulations in
NSCLC cells by CSC marker expression post-treatment. |
Table III
The percentage of subpopulations in
NSCLC cells by CSC marker expression post-treatment.
| Cell lines | Subpopulations | Treatments
|
|---|
| Curcumin (%) | Cisplatin (%) | Combination
(%) |
|---|
| A549 |
CD166+/EpCAM+ | 25.2±3.2 | 25.1±7.4 | 34.6±0.7 |
|
CD166−/EpCAM− | 14.9±3.6 | 21.4±4.2 | 23.6±12.6 |
| H2170 |
CD166+/EpCAM+ | 47.4±18.4 | 50.1±14.3 | 55.0±0.4 |
|
CD166−/EpCAM− | 1.3±0.2 | 2.2±0.2 | 1.7±0.4 |
Curcumin enhances the sensitivity of the
CSC subpopulation of CD166+/EpCAM+ cells to
cisplatin-induced apoptosis
The apoptotic effects of the combined treatment of
curcumin either by synergism with cisplatin or sensitising effects
prior to treatment with low dose cisplatin in the double-positive
(CD166+/EpCAM+) CSC subpopulation was
examined using the apoptosis assay (Annexin V/PI) 48 h
post-treatment. The unsorted A549 and H2170 cells were used as a
control to indicate the basal level of apoptosis following the
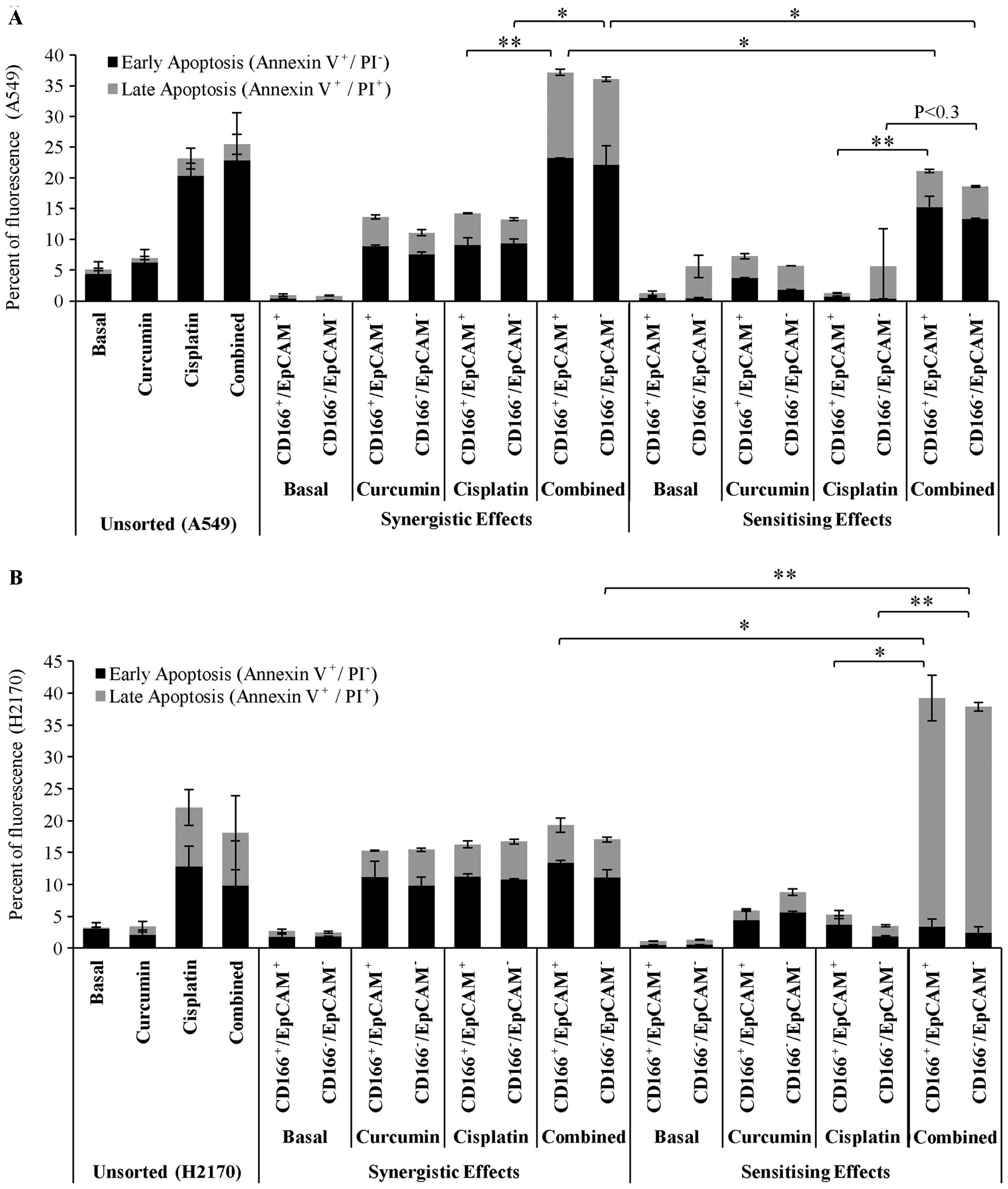
treatments (Fig. 6). As shown for
the synergistic effect of curcumin (Fig. 6A); the results indicated that single
treatments of curcumin and cisplatin induced apoptosis in the
double-positive (CD166+/EpCAM+) CSC
subpopulation of A549 cells to an average of 14 and 15% following
48 h of treatments. Moreover, the apoptotic effect was
significantly increased to an average of ~37% in the
double-positive (CD166+/EpCAM+) CSC
subpopulation as these two treatments were applied simultaneously.
The results also showed that curcumin sensitisation prior to
treatment with low dose cisplatin in the double-positive
(CD166+/EpCAM+) CSC subpopulation of A549
cells substantially increased the percentage of apoptosis by ~20%
as compared to treatment with low dose cisplatin with only 2%
apoptosis (Fig. 6A).
There were no significant changes in the percentage
of apoptosis between the single treatment of curcumin or cisplatin,
and the combination treatments by synergistic effects on sorted
H2170 cells (Fig. 6B). This result
suggests that the cells are highly sensitive to both curcumin and
cisplatin; the IC50 concentrations of both treatments
given to the cells have already induced a maximal response.
Interestingly, we noted that by sensitising the double-positive
(CD166+/EpCAM+) CSC subpopulation of H2170
cells to curcumin, prior to treatment with low dose cisplatin
notably enhanced its apoptotic effect by 40%, compared to only 20%
apoptosis as observed for the synergistic treatments. Combination
treatments by sensitisation of the double-positive
CD166+/EpCAM+ CSC subpopulation of H2170
cells to curcumin, also significantly increased the percentage of
late apoptosis to 35%, compared to treatment with low dose
cisplatin with only 4% detected (Fig.
6B).
Curcumin enhances the cisplatin-induced
inhibition of the metastasis of the highly migratory CSC
subpopulation (CD166+/EpCAM+) in the NSCLC
cell lines
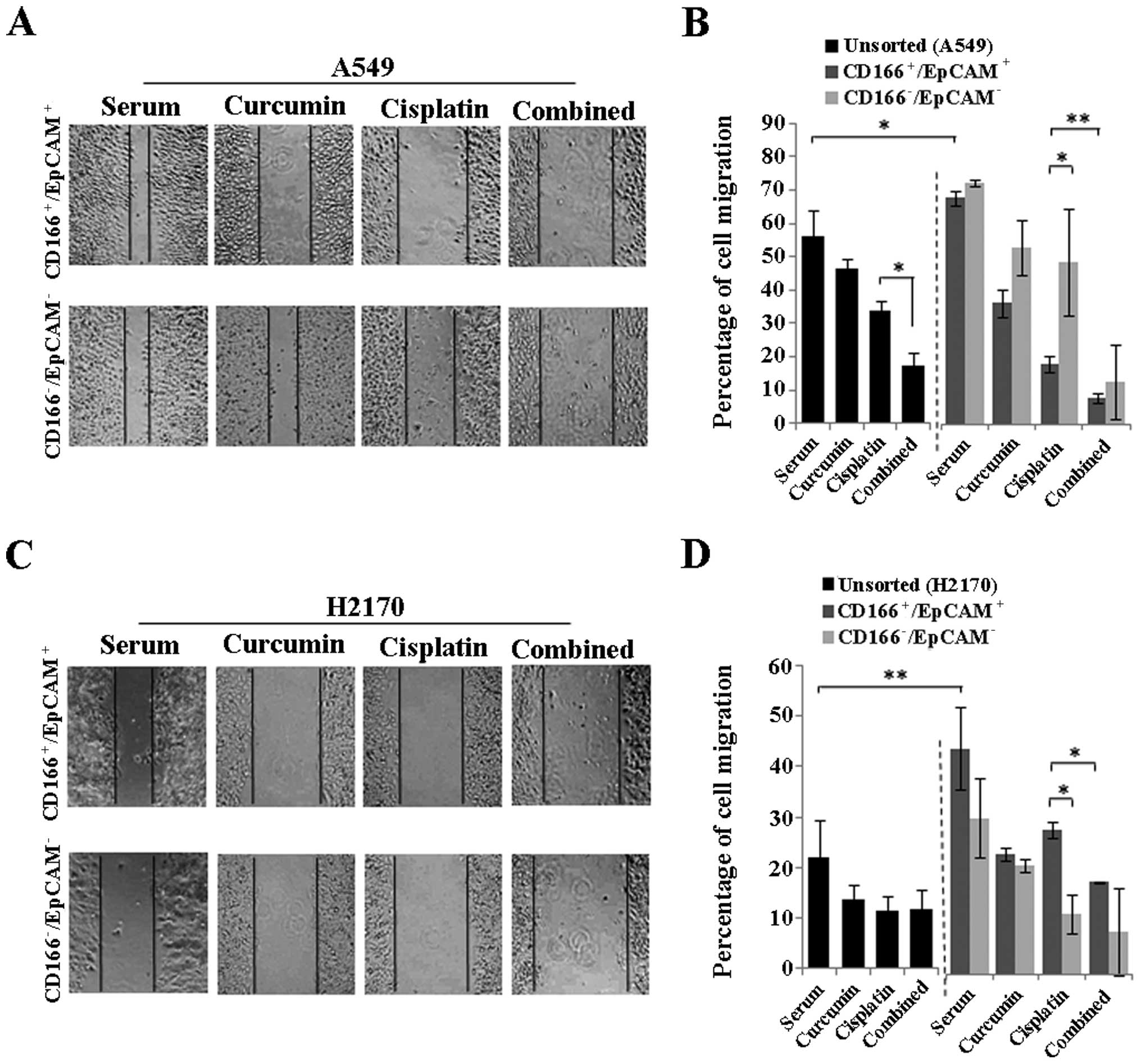
To evaluate the migratory potential of the CSC
subpopulation in the NSCLC cell lines (A549 and H2170) and the
effects of curcumin either alone or in combination with cisplatin
to inhibit the migration of these cells; scratch wound (migration)
assay was performed in the sorted
(CD166+/EpCAM+ and
CD166−/EpCAM−) and unsorted NSCLC cell lines
48 h post-treatment. As depicted in Fig. 7B and D, the double-positive
(CD166+/EpCAM+) CSC subpopulation had a
significantly higher migratory potential as compared to the
unsorted cells observed for both NSCLC cell lines. The combination
of curcumin and cisplatin reduced the percentage of cell migration
from 33.8 (cisplatin) to 17.3% (combined) in the unsorted A549
cells (Fig. 7B) with no apparent
changes as noted in the H2170 cells (Fig. 7D). Furthermore, combined treatment
markedly inhibited the migration of the
CD166+/EpCAM+ subpopulation from 19.6 to 8.7%
in the A549 cells and from 32.6 to 20.9% in the H2170 cells as
compared to cisplatin treatment alone (Fig. 7B and D). Moreover, curcumin alone
was able to inhibit the migration of the
CD166+/EpCAM+ subpopulation in both the A549
and H2170 cells signifying the potential of curcumin on CSC
inhibition. A higher cell migration was also noted in the
CD166−/EpCAM− compared to the
CD166+/EpCAM+ subpopulation for the cisplatin
treatment alone in A549 cells, while an opposite effect was noted
in the H2170 cells (Fig. 7B and D).
However, there were no differences in the percentage of cell
migration between the CD166+/EpCAM+ and
CD166−/EpCAM− subpopulation for the
combination treatment in both NSCLC cell lines (Fig. 7B and D).
Curcumin together with cisplatin
increases the positive expression of apoptotic and cell
cycle-regulating genes in the sorted cells
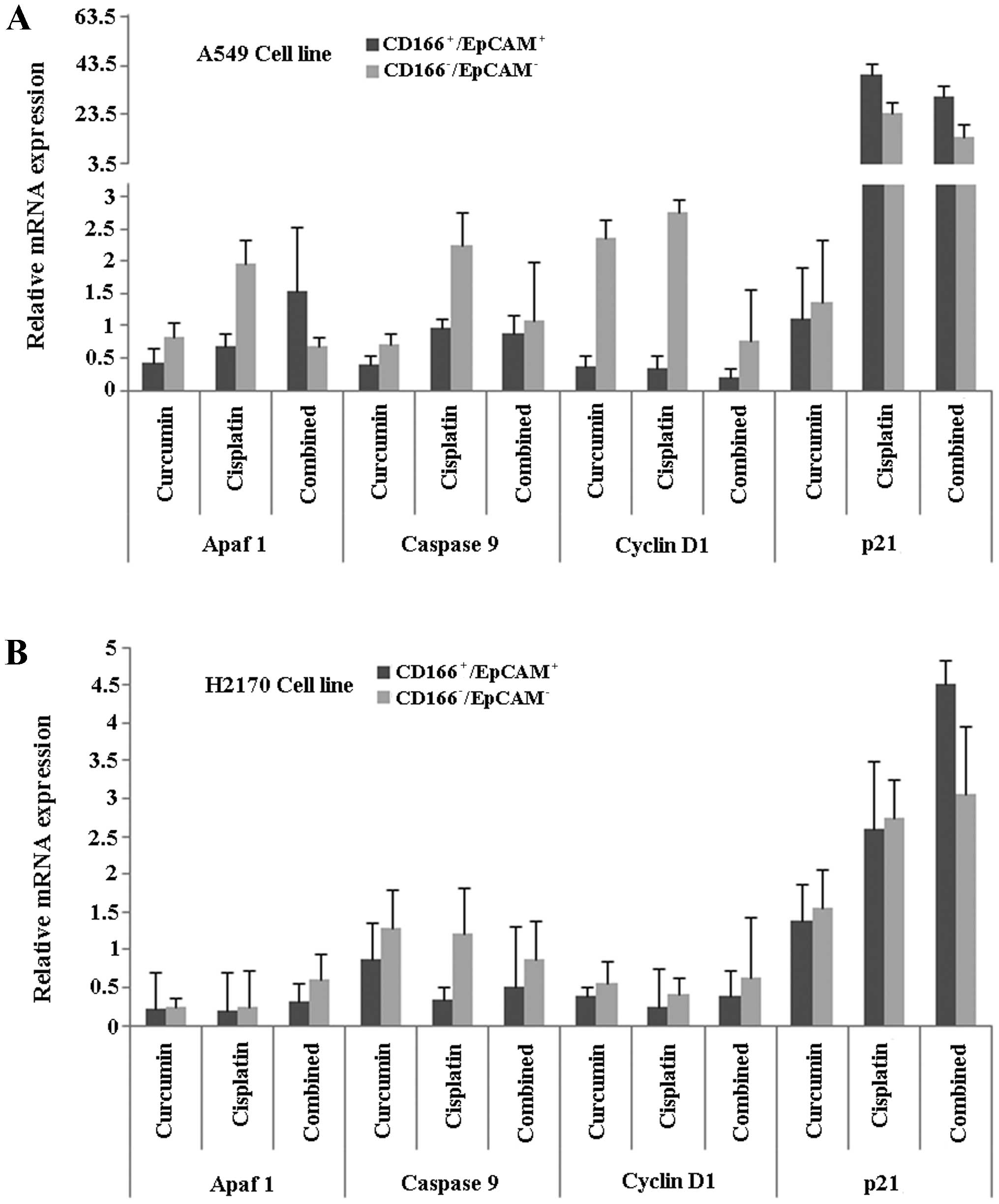
Finally, in order to understand the mechanisms
behind the process, we investigated specific genes involved in
apoptosis (Apaf1, cytochrome c and caspase-9) and cell cycle
regulation (cyclin D1 and p21) in the double-positive
(CD166+/EpCAM+) CSC subpopulation of both
A549 and H2170 cells, after induction of treatments using either
curcumin or cisplatin, and the combination of both. The results
showed that the relative gene expression level of Apaf1 was higher
in the combined treatment group compared to the single treatments
(curcumin or cisplatin) in the CD166+/EpCAM+
subpopulation of A549 cells (Fig.
8A). Furthermore, the expression of p21 was high, with low
expression of the cyclin D1 gene, in the
CD166+/EpCAM+ subpopulation of both the A549
and H2170 cells, as compared to the
CD166−/EpCAM− subpopulation in the combined
treatment group (Fig. 8A and B).
Combined treatments induced high expression of caspase-9 in the
CD166+/EpCAM+ subpopulation of A549, compared
to single treatments of curcumin (Fig.
8A). On the other hand, the expression of caspase-9 was
consistently low in the CD166+/EpCAM+
subpopulation of H2170 cells for all of the treatments (Fig. 8B).
Discussion
The existence of chemoresistant tumour cells is one
of the major obstacles reducing the efficacies of antitumour agents
for cancer treatments. Studies have demonstrated that CSCs, as the
main component in the tumour that drives tumour invasion,
metastasis and relapse, are also believed to be the main reason for
the chemoresistant phenotype. Currently, cisplatin and other
platinum-based compounds are the most effective agents for the
treatment of lung cancer patients, and they are usually combined
with other agents such as docetaxel, gemcitabine and paclitaxel to
yield higher efficacies (57).
However the use of conventional drugs is limited due to the side
effects and the resistant phenotype acquired by tumours (58,59).
Active compounds derived from plants, microbes and marine organisms
have been the interest of many investigators recently. These active
compounds either in their crude or purified extracts have been
shown to either have synergistic effects with chemotherapy or
sensitising effects on CSCs, thus yielding superior efficacy as
compared to chemotherapy alone (60,61).
It is also suggested that the sensitising effects of these active
compounds, might be useful to reduce the toxicity in patients by
high dose chemotherapy.
In the present study, we studied the efficacy of
curcumin, a natural compound extracted from Curcuma longa,
either alone or in combination with cisplatin on the inhibition of
double-positive (CD166+/EpCAM+) CSC
subpopulation sorted from NSCLC cell lines (A549 and H2170), that
we previously characterised (55).
Curcumin cytotoxicity in NSCLC cell lines indicated by the
IC50 values (Table II)
showed that the agent, similar to cisplatin, is able to inhibit
NSCLC cell proliferation. Moreover, sensitisation of both NSCLC
cell lines using curcumin prior to treatment with low dose
cisplatin, significantly reduced the percentage of viability in
both the NSCLC cell lines compared to the treatment with low dose
cisplatin (Fig. 2C and D). These
results are in agreement with studies that have shown similar
findings on the ability of curcumin to enhance the effects of
cisplatin in NSCLC cell lines (23,62).
These findings might also suggest that through the sensitising
effects of curcumin, a combination of both curcumin and cisplatin
could potentially be used as a treatment strategy to compliment the
effects of low dose ciplatin. Thus, a higher therapeutic efficacy
with lower toxicity can be achieved. Moreover, the IC50
value of curcumin, as well as cisplatin, that were evaluated in the
NSCLC cell lines did not induce cytotoxicity on normal epithelial
cells (IMR-90) (Fig. 2E) indicating
that the target effects of both agents are tumour-specific.
The presence of CSCs as part of the tumour
population has recently become the interest of many investigators.
Most studies believe that by specifically targeting these
subpopulations, the efficacy of treatments could be enhanced and
eventually might reduce the chances for relapse (56). However, the major obstacles to this
approach are the resistance characteristics of CSCs upon treatment
(63). We noted that curcumin
enhanced the induction of apoptosis by cisplatin in the
CD166+/EpCAM+ subpopulation in both A549 and
H2170 cells by either sensitising or synergistic effects (Fig. 6). Interestingly, in H2170 cells,
synergistic effect of curcumin by direct combination with cisplatin
did not induce significant changes in the percentage of apoptosis
in the CD166+/EpCAM+ subpopulation of this
cell line, as compared to cisplatin alone (Fig. 6B). However, the percentage of
apoptosis was significantly increased when the subpopulation was
initially sensitised to curcumin, prior to treatment with low dose
cisplatin (Fig. 6B). Based on this
observation, we believe that curcumin has the potential to alter
the phenotype of NSCLC cells to treatments by enhancing the
sensitivity of CSCs to chemotherapy. This theory is supported by
few studies that have shown the same effects in several tumour
models such as breast tumours and colon cancer, where these
investigators have attributed the effects of curcumin on the
inhibition of CSCs (64,65). These results also suggest that the
approach of utilising curcumin either by direct combination
(synergistic) or indirect combination (sensitising) with cisplatin
should be taken into consideration if efficacy of the combined
treatment is to be optimal on inhibiting the CSC subpopulation.
Moreover, analysis on the gene expression level in the
CD166+/EpCAM+ subpopulation of both A549 and
H2170 cells from our previous study also demonstrated that there
are variations in the tumourigenic mRNA expression between this
subpopulation that we believed could further be attributed to the
heterogeneity of CSCs to treatment outcomes (55).
The properties of CSCs, a subpopulation of cells
that exhibit stem cell characteristics and contribute to treatment
resistance, have been suggested as a candidate for mediating
metastatic progression (66). In
contrary, other cancer cells which do not exhibit stem cell
characteristics and metastasise into distant tissue and confront an
entirely new microenvironment may often be unable to colonise and
grow. Only CSCs with high EMT (epithelial to mesenchymal
transition) characteristics and a resistant phenotype have the
capacity to metastasise and survive long enough and arrive at
distant sites (67). Our results
presented here showed that the CD166+/EpCAM+
subpopulation of both NSCLC cell lines have substantially higher
migratory potential as compared to the unsorted cells (Fig. 7), consistent with the high
metastatic characteristics of CSCs, that have been shown by several
studies (68,69). Furthermore, a combination of both
curcumin and cisplatin markedly inhibited the migration of the
CD166+/EpCAM+ subpopulation in both A549
(Fig. 7A and B) and H2170 cells
(Fig. 7C and D) compared to
cisplatin treatment alone, indicating that the combination
treatment induced superior effects on inhibiting the migration of
CSCs. This finding might also indicate that the synergistic effects
of both treatments could be utilised as a treatment strategy to
combat the highly migratory CSC subpopulation. Thus, the
probability of metastatic progression after chemotherapy in NSCLC
may be reduced.
We observed that the combination treatment reduced
the mRNA expression of cyclin D1 and induced p21 expression in the
CD166+/EpCAM+ subpopulation in both the A549
and H2170 cells that eventually halted the growth of these cells
(Fig. 8). These results are
consistent with previous studies, demonstrating that curcumin by
itself, has the potential to alter cyclin D1 and p21 expression and
in combination with common chemotherapeutic drugs, the inhibitory
effects were enhanced through the inhibition of CSCs (70,71).
Although we noted that the combination treatment enhanced mRNA
expression of Apaf1 and caspase-9 in the CSC subpopulation of A549
cells, compared to cisplatin treatment alone, the combination
treatment did not influence Apaf1 and caspase-9 expression in the
CSC subpopulation of H2170 cells. We believed that this is due to
the heterogeneity of CSCs that leads to the different sensitivity
of the double-positive (CD166+/EpCAM+)
subpopulation to the treatment outcome.
In conclusion, we showed that curcumin is able to
increase the efficacy of low dose cisplatin in unsorted NSCLC cell
lines. Through our investigation of the sorted NSCLC cell lines, we
also found that curcumin had the capacity to enhance
cisplatin-induced metastatic inhibition and apoptosis of the highly
migratory CSC subpopulation (CD166+/EpCAM+)
in the NSCLC cell lines suggesting that curcumin might be useful as
a complement to common chemotherapy for inhibiting tumour
progression and reducing metastasis.
Acknowledgments
The authors wish to thank the Director General of
Health, Malaysia for his permission to publish this paper. This
study was supported by a Ministry of Health grant,
JPP-IMR-12-023.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012.
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011.
|
|
3
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000.
|
|
4
|
Mountain CF, Lukeman JM, Hammar SP,
Chamberlain DW, Coulson WF, Page DL, Victor TA and Weiland LH: Lung
cancer classification: The relationship of disease extent and cell
type to survival in a clinical trials population. J Surg Oncol.
35:147–156. 1987.
|
|
5
|
Curtis SJ, Sinkevicius KW, Li D, Lau AN,
Roach RR, Zamponi R, Woolfenden AE, Kirsch DG, Wong K-K and Kim CF:
Primary tumor genotype is an important determinant in
identification of lung cancer propagating cells. Cell Stem Cell.
7:127–133. 2010.
|
|
6
|
Tan BT, Park CY, Ailles LE and Weissman
IL: The cancer stem cell hypothesis: A work in progress. Lab
Invest. 86:1203–1207. 2006.
|
|
7
|
Gil J, Stembalska A, Pesz KA and Sasiadek
MM: Cancer stem cells: The theory and perspectives in cancer
therapy. J Appl Genet. 49:193–199. 2008.
|
|
8
|
Li L and Neaves WB: Normal stem cells and
cancer stem cells: The niche matters. Cancer Res. 66:4553–4557.
2006.
|
|
9
|
Mukherjee S, Kong J and Brat DJ: Cancer
stem cell division: When the rules of asymmetry are broken. Stem
Cells Dev. 24:405–416. 2015.
|
|
10
|
Chen S and Huang EH: The colon cancer stem
cell microenvironment holds keys to future cancer therapy. J
Gastrointest Surg. 18:1040–1048. 2014.
|
|
11
|
O'Flaherty JD, Barr M, Fennell D, Richard
D, Reynolds J, O'Leary J and O'Byrne K: The cancer stem-cell
hypothesis: Its emerging role in lung cancer biology and its
relevance for future therapy. J Thorac Oncol. 7:1880–1890.
2012.
|
|
12
|
Hong IS, Lee HY and Nam JS: Cancer stem
cells: The 'Achilles heel' of chemo-resistant tumors. Recent
Patents Anticancer Drug Discov. 10:2–22. 2015.
|
|
13
|
Orian-Rousseau V and Ponta H: Perspectives
of CD44 targeting therapies. Arch Toxicol. 89:3–14. 2015.
|
|
14
|
Ma J, Fang B, Zeng F, Pang H, Zhang J, Shi
Y, Wu X, Cheng L, Ma C, Xia J, et al: Curcumin inhibits cell growth
and invasion through up-regulation of miR-7 in pancreatic cancer
cells. Toxicol Lett. 231:82–91. 2014.
|
|
15
|
Mukherjee S, Mazumdar M, Chakraborty S,
Manna A, Saha S, Khan P, Bhattacharjee P, Guha D, Adhikary A,
Mukhjerjee S, et al: Curcumin inhibits breast cancer stem cell
migration by amplifying the E-cadherin/β-catenin negative feedback
loop. Stem Cell Res Ther. 5:1162014.
|
|
16
|
Sarkar R, Mukherjee A, Mukherjee S, Biswas
R, Biswas J and Roy M: Curcumin augments the efficacy of antitumor
drugs used in leukemia by modulation of heat shock proteins via
HDAC6. Environ Pathol Toxicol Oncol. 33:247–263. 2014.
|
|
17
|
Xiao C, Wang L, Zhu L, Zhang C and Zhou J:
Curcumin inhibits oral squamous cell carcinoma SCC-9 cells
proliferation by regulating miR-9 expression. Biochem Biophys Res
Commun. 454:576–580. 2014.
|
|
18
|
Ye M and Zhang J and Zhang J, Miao Q, Yao
L and Zhang J: Curcumin promotes apoptosis by activating the
p53-miR-192-5p/215-XIAP pathway in non-small cell lung cancer.
Cancer Lett. 357:196–205. 2015.
|
|
19
|
Anand P, Sundaram C, Jhurani S,
Kunnumakkara AB and Aggarwal BB: Curcumin and cancer: An 'old-age'
disease with an 'age-old' solution. Cancer Lett. 267:133–164.
2008.
|
|
20
|
Andjelkovic T, Pesic M, Bankovic J, Tanic
N, Markovic ID and Ruzdijic S: Synergistic effects of the purine
analog sulfinosine and curcumin on the multidrug resistant human
non-small cell lung carcinoma cell line (NCI-H460/R). Cancer Biol
Ther. 7:1024–1032. 2008.
|
|
21
|
Ebert B, Seidel A and Lampen A:
Phytochemicals induce breast cancer resistance protein in Caco-2
cells and enhance the transport of benzo(a)pyrene-3-sulfate.
Toxicol Sci. 96:227–236. 2007.
|
|
22
|
Hou X-L, Takahashi K, Tanaka K, Tougou K,
Qiu F, Komatsu K, Takahashi K and Azuma J: Curcuma drugs and
curcumin regulate the expression and function of P-gp in Caco-2
cells in completely opposite ways. Int J Pharm. 358:224–229.
2008.
|
|
23
|
Chanvorachote P, Pongrakhananon V,
Wannachaiyasit S, Luanpitpong S, Rojanasakul Y and Nimmannit U:
Curcumin sensitizes lung cancer cells to cisplatin-induced
apoptosis through superoxide anion-mediated Bcl-2 degradation.
Cancer Invest. 27:624–635. 2009.
|
|
24
|
Li S, Liu Z, Zhu F, Fan X, Wu X, Zhao H
and Jiang L: Curcumin lowers erlotinib resistance in non-small cell
lung carcinoma cells with mutated EGF receptor. Oncol Res.
21:137–144. 2013.
|
|
25
|
Chan MM, Fong D, Soprano KJ, Holmes WF and
Heverling H: Inhibition of growth and sensitization to
cisplatin-mediated killing of ovarian cancer cells by polyphenolic
chemopreventive agents. J Cell Physiol. 194:63–70. 2003.
|
|
26
|
Chan MM, Soprano KJ, Weinstein K and Fong
D: Epigallo catechin-3-gallate delivers hydrogen peroxide to induce
death of ovarian cancer cells and enhances their cisplatin
susceptibility. J Cell Physiol. 207:389–396. 2006.
|
|
27
|
Duarte VM, Han E, Veena MS, Salvado A, Suh
JD, Liang LJ, Faull KF, Srivatsan ES and Wang MB: Curcumin enhances
the effect of cisplatin in suppression of head and neck squamous
cell carcinoma via inhibition of IKKβ protein of the NFκB pathway.
Mol Cancer Ther. 9:2665–2675. 2010.
|
|
28
|
Notarbartolo M, Poma P, Perri D, Dusonchet
L, Cervello M and D'Alessandro N: Antitumor effects of curcumin,
alone or in combination with cisplatin or doxorubicin, on human
hepatic cancer cells. Analysis of their possible relationship to
changes in NF-κB activation levels and in IAP gene expression.
Cancer Lett. 224:53–65. 2005.
|
|
29
|
Aggarwal BB, Shishodia S, Takada Y,
Banerjee S, Newman RA, Bueso-Ramos CE and Price JE: Curcumin
suppresses the paclitaxel-induced nuclear factor-kappaB pathway in
breast cancer cells and inhibits lung metastasis of human breast
cancer in nude mice. Clin Cancer Res. 11:7490–7498. 2005.
|
|
30
|
Cheng AL, Hsu CH, Lin JK, Hsu MM, Ho YF,
Shen TS, Ko JY, Lin JT, Lin BR, Ming-Shiang W, et al: Phase I
clinical trial of curcumin, a chemopreventive agent, in patients
with high-risk or pre-malignant lesions. Anticancer Res.
21:2895–2900. 2001.
|
|
31
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007.
|
|
32
|
Kakarala M, Brenner DE, Korkaya H, Cheng
C, Tazi K, Ginestier C, Liu S, Dontu G and Wicha MS: Targeting
breast stem cells with the cancer preventive compounds curcumin and
piperine. Breast Cancer Res Treat. 122:777–785. 2010.
|
|
33
|
Fong D, Yeh A, Naftalovich R, Choi TH and
Chan MM: Curcumin inhibits the side population (SP) phenotype of
the rat C6 glioma cell line: Towards targeting of cancer stem cells
with phytochemicals. Cancer Lett. 293:65–72. 2010.
|
|
34
|
Zhuang W, Long L, Zheng B, Ji W, Yang N,
Zhang Q and Liang Z: Curcumin promotes differentiation of
glioma-initiating cells by inducing autophagy. Cancer Sci.
103:684–690. 2012.
|
|
35
|
Pellacani D, Oldridge EE, Collins AT and
Maitland NJ: Prominin-1 (CD133) expression in the prostate and
prostate cancer: A marker for quiescent stem cells. Adv Exp Med
Biol. 777:167–184. 2013.
|
|
36
|
Reyes EE, Kunovac SK, Duggan R, Kregel S
and Vander Griend DJ: Growth kinetics of CD133-positive prostate
cancer cells. Prostate. 73:724–733. 2013.
|
|
37
|
Vander Griend DJ, Karthaus WL, Dalrymple
S, Meeker A, DeMarzo AM and Isaacs JT: The role of CD133 in normal
human prostate stem cells and malignant cancer-initiating cells.
Cancer Res. 68:9703–9711. 2008.
|
|
38
|
Bi CL, Fang JS, Chen FH, Wang YJ and Wu J:
Chemoresistance of CD133(+) tumor stem cells from human brain
glioma. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 32:568–573. 2007.In
Chinese.
|
|
39
|
Choi SA, Wang KC, Phi JH, Lee JY, Park CK,
Park SH and Kim SK: A distinct subpopulation within CD133 positive
brain tumor cells shares characteristics with endothelial
progenitor cells. Cancer Lett. 324:221–230. 2012.
|
|
40
|
Li MC, Deng YW, Wu J, Chen FH, Liu JF and
Fang JS: Isolation and characterization of brain tumor stem cells
in human medulloblastoma. Ai Zheng. 25:241–246. 2006.In
Chinese.
|
|
41
|
Singh S and Dirks PB: Brain tumor stem
cells: Identification and concepts. Neurosurg Clin N Am. 18:31–38.
2007.
|
|
42
|
Kozovska Z, Gabrisova V and Kucerova L:
Colon cancer: Cancer stem cells markers, drug resistance and
treatment. Biomed Pharmacother. 68:911–916. 2014.
|
|
43
|
Mărgaritescu C, Pirici D, Cherciu I,
Bărbălan A, Cârtână T and Săftoiu A: CD133/CD166/Ki-67 triple
immunofluorescence assessment for putative cancer stem cells in
colon carcinoma. J Gastrointestin Liver Dis. 23:161–170. 2014.
|
|
44
|
Vincent Z, Urakami K, Maruyama K,
Yamaguchi K and Kusuhara M: CD133-positive cancer stem cells from
Colo205 human colon adenocarcinoma cell line show resistance to
chemotherapy and display a specific metabolomic profile. Genes
Cancer. 5:250–260. 2014.
|
|
45
|
Cogliati B, Aloia TP, Bosch RV, Alves VA,
Hernandez-Blazquez FJ and Dagli ML: Identification of hepatic
stem/progenitor cells in canine hepatocellular and
cholangiocellular carcinoma. Vet Comp Oncol. 8:112–121. 2010.
|
|
46
|
Tomuleasa C, Soritau O, Rus-Ciuca D, Pop
T, Todea D, Mosteanu O, Pintea B, Foris V, Susman S, Kacsó G, et
al: Isolation and characterization of hepatic cancer cells with
stem-like properties from hepatocellular carcinoma. J
Gastrointestin Liver Dis. 19:61–67. 2010.
|
|
47
|
Yang XR, Xu Y, Yu B, Zhou J, Qiu SJ, Shi
GM, Zhang BH, Wu WZ, Shi YH, Wu B, et al: High expression levels of
putative hepatic stem/progenitor cell biomarkers related to tumour
angio-genesis and poor prognosis of hepatocellular carcinoma. Gut.
59:953–962. 2010.
|
|
48
|
Zhang L, Sun H, Zhao F, Lu P, Ge C, Li H,
Hou H, Yan M, Chen T, Jiang G, et al: BMP4 administration induces
differentiation of CD133+ hepatic cancer stem cells,
blocking their contributions to hepatocellular carcinoma. Cancer
Res. 72:4276–4285. 2012.
|
|
49
|
Bertolini G, Roz L, Perego P, Tortoreto M,
Fontanella E, Gatti L, Pratesi G, Fabbri A, Andriani F, Tinelli S,
et al: Highly tumor-igenic lung cancer CD133+ cells
display stem-like features and are spared by cisplatin treatment.
Proc Natl Acad Sci USA. 106:16281–16286. 2009.
|
|
50
|
Salnikov AV, Gladkich J, Moldenhauer G,
Volm M, Mattern J and Herr I: CD133 is indicative for a resistance
phenotype but does not represent a prognostic marker for survival
of non-small cell lung cancer patients. Int J Cancer. 126:950–958.
2010.
|
|
51
|
Sullivan JP, Spinola M, Dodge M, Raso MG,
Behrens C, Gao B, Schuster K, Shao C, Larsen JE, Sullivan LA, et
al: Aldehyde dehydrogenase activity selects for lung adenocarcinoma
stem cells dependent on notch signaling. Cancer Res. 70:9937–9948.
2010.
|
|
52
|
Woo T, Okudela K, Mitsui H, Yazawa T,
Ogawa N, Tajiri M, Yamamoto T, Rino Y, Kitamura H and Masuda M:
Prognostic value of CD133 expression in stage I lung
adenocarcinomas. Int J Clin Exp Pathol. 4:32–42. 2010.
|
|
53
|
Sterlacci W, Savic S, Fiegl M, Obermann E
and Tzankov A: Putative stem cell markers in non-small-cell lung
cancer: A clini-copathologic characterization. J Thorac Oncol.
9:41–49. 2014.
|
|
54
|
Zhang WC, Shyh-Chang N, Yang H, Rai A,
Umashankar S, Ma S, Soh BS, Sun LL, Tai BC, Nga ME, et al: Glycine
decarboxylase activity drives non-small cell lung cancer
tumor-initiating cells and tumorigenesis. Cell. 148:259–272.
2012.
|
|
55
|
Zakaria N, Yusoff NM, Zakaria Z, Lim MN,
Baharuddin PJN, Fakiruddin KS and Yahaya B: Human non-small cell
lung cancer expresses putative cancer stem cell markers and
exhibits the transcriptomic profile of multipotent cells. BMC
Cancer. 15:842015.
|
|
56
|
Chen K, Huang YH and Chen JL:
Understanding and targeting cancer stem cells: Therapeutic
implications and challenges. Acta Pharmacol Sin. 34:732–740.
2013.
|
|
57
|
Xu X-M, Zhang Y, Qu D, Liu H-B, Gu X, Jiao
G-Y and Zhao L: Combined anticancer activity of osthole and
cisplatin in NCI-H460 lung cancer cells in vitro. Exp Ther
Med. 5:707–710. 2013.
|
|
58
|
Douillard J-Y, Eckardt J and Scagliotti
GV: Challenging the platinum combinations in the chemotherapy of
NSCLC. Lung Cancer. 38(Suppl 4): 21–28. 2002.
|
|
59
|
Stewart DJ: Mechanisms of resistance to
cisplatin and carboplatin. Crit Rev Oncol Hematol. 63:12–31.
2007.
|
|
60
|
Burnett J, Newman B and Sun D: Targeting
cancer stem cells with natural products. Curr Drug Targets.
13:1054–1064. 2012.
|
|
61
|
Mondal S, Bandyopadhyay S, Ghosh MK,
Mukhopadhyay S, Roy S and Mandal C: Natural products: Promising
resources for cancer drug discovery. Anticancer Agents Med Chem.
12:49–75. 2012.
|
|
62
|
Ye MX, Zhao YL, Li Y, Miao Q, Li ZK, Ren
XL, Song LQ, Yin H and Zhang J: Curcumin reverses cisplatin
resistance and promotes human lung adenocarcinoma A549/DDP cell
apoptosis through HIF-1alpha and caspase-3 mechanisms.
Phytomedicine. 19:779–787. 2012.
|
|
63
|
Zhou B-BS, Zhang H, Damelin M, Geles KG,
Grindley JC and Dirks PB: Tumour-initiating cells: Challenges and
opportunities for anticancer drug discovery. Nat Rev Drug Discov.
8:806–823. 2009.
|
|
64
|
Buhrmann C, Kraehe P, Lueders C, Shayan P,
Goel A and Shakibaei M: Curcumin suppresses crosstalk between colon
cancer stem cells and stromal fibroblasts in the tumor
microen-vironment: Potential role of EMT. PLoS One.
9:e1075142014.
|
|
65
|
Charpentier MS, Whipple RA, Vitolo MI,
Boggs AE, Slovic J, Thompson KN, Bhandary L and Martin SS: Curcumin
targets breast cancer stem-like cells with microtentacles that
persist in mammospheres and promote reattachment. Cancer Res.
74:1250–1260. 2014.
|
|
66
|
Wang X, Zhu Y, Ma Y, Wang J, Zhang F, Xia
Q and Fu D: The role of cancer stem cells in cancer metastasis: New
perspective and progress. Cancer Epidemiol. 37:60–63. 2013.
|
|
67
|
Fazilaty H, Gardaneh M, Bahrami T,
Salmaninejad A and Behnam B: Crosstalk between breast cancer stem
cells and metastatic niche: Emerging molecular metastasis pathway?
Tumour Biol. 34:2019–2030. 2013.
|
|
68
|
Croker AK, Goodale D, Chu J, Postenka C,
Hedley BD, Hess DA and Allan AL: High aldehyde dehydrogenase and
expression of cancer stem cell markers selects for breast cancer
cells with enhanced malignant and metastatic ability. J Cell Mol
Med. 13:2236–2252. 2009.
|
|
69
|
Hermann PC, Huber SL, Herrler T, Aicher A,
Ellwart JW, Guba M, Bruns CJ and Heeschen C: Distinct populations
of cancer stem cells determine tumor growth and metastatic activity
in human pancreatic cancer. Cell Stem Cell. 1:313–323. 2007.
|
|
70
|
Srivastava RK, Chen Q, Siddiqui I, Sarva K
and Shankar S: Linkage of curcumin-induced cell cycle arrest and
apoptosis by cyclin-dependent kinase inhibitor p21(/WAF1/CIP1).
Cell Cycle. 6:2953–2961. 2007.
|
|
71
|
Zhang H, Yu T, Wen L, Wang H, Fei D and
Jin C: Curcumin enhances the effectiveness of cisplatin by
suppressing CD133(+) cancer stem cells in laryngeal carcinoma
treatment. Exp Ther Med. 6:1317–1321. 2013.
|