Introduction
Hepatocellular carcinoma (HCC) represents one of the
most commonly seen malignancies in the world, and has a high
incident rate in China. According to the World Cancer Report of
2014 of the World Health Organization, the number of newly
diagnosed cases and HCC-related death in China ranks the first in
the world. It accounts for ~50% of the global statistics. Since
early detection of HCC is difficult, most HCC patients miss their
optimal treatment opportunities. The efficacy of current routine
chemotherapies for HCC remains unsatisfactory. It is estimated that
~60–70% patients receiving radical resection have metastatic
recurrence within 5 years. The 5-year survival rate is only 7%. The
mechanisms underlying HCC tumorigenesis and progression remain
poorly understood.
The Cks (cyclin-dependent kinase subunit) family
consists of two members, Cks1 and Cks2, which are small protein
molecule that are highly conserved in eukaryotic organisms. The two
Cks proteins shares 81% peptide sequence homology with the yeast
Cks protein (1). Emerging data show
that expression of Cks proteins is closely related to the
pathogenesis and progression of esophageal, prostate, gastric and
colorectal cancers (2–7). We previously reported that Cks mRNA
and protein expression levels in HCC tissues were elevated and
correlated with cancer histopathological grading stages and AFP
levels (8). Although it has been
found that overexpression of the Cks family members is associated
with tumorigenesis and growth, the detailed molecular mechanism by
which CKS contributes to tumor cell growth and metastasis remain
unclear.
Herein we further report that overexpression of Cks1
and Cks2 by transfection increased cell proliferation, and reduced
apoptosis in HepG2 cells. Consistently, depleting Cks1 or Cks2
expression by siRNA in HepG2 cells reduced cell proliferation and
increased apoptosis. The results indicate that overexpressed Cks1
and Cks2 in HCC cells promote proliferation and survival of the
cells and therefore, promotes HCC growth and progression.
Materials and methods
Materials
The human HCC HepG2 cell-line was kindly provided by
the Laboratory of Hepatobiliary Surgery at the Xiamen University
Affiliated Zhongshan Hospital (obtained from Shanghai Cell Bank of
the Chinese Academy of Sciences, Shanghai, China). Dulbecco's
modified Eagle's medium (DMEM) was obtained from Hyclone (Logan,
UT, USA); fetal bovine serum (FBS) was obtained from Gibco-BRL
(Gaithersburg, MD, USA); HiPerfect transfection reagent for siRNA
was from Qiagen (Hilden, Germany), Lipofectamine 2000 was from
Invitrogen (Carlsbad, CA, USA), TRIzol reagent for RNA extraction
was obtained from Tiangen (Beijing, China). Cell Counting Kit-8 was
from Dojindo (Kumamoto, Japan). RevertAid First-Strand cDNA
Synthesis kits were obtained from Fermentas (Hanover, MD, USA),
GoTaq Probe qPCR Master Mix used for real-time fluorescent
quantitative PCR assay was obtained from Promega (Madison, WI,
USA). RIPA lysis buffer was from Solarbio (Beijing, China). Cks1
antibody was obtained from Abcam (Cambridge, UK), and the Cks2
antibody was obtained from Sigma-Aldrich (St. Louis, MO, USA).
Additionally, enhanced chemiluminescent (ECL) detection reagents
for western blotting assays were obtained from Millipore
(Billerica, MA, USA).
Methods
siRNA and overexpressing vectors
The siRNA sequences that were targeted against Cks1
and Cks2 were designed by Qiagen. The Hs-CKS1B-4 siRNA listed below
were used to knock down Cks1: target sequence
5′-AAGTTTGTATGCATTTAA-3′, sense strand 5′-CAUCUUUCUGAUAACAUUATT-3′,
and antisense strand 5′-UAAUGUUAUCAGAAAGAUGTT-3′. The Hs-CKS2-10
siRNA listed below were used to knock down Cks2: target sequence
5′-AACATCTTTCTGATAACATTA-3′, sense strand
5′-GUUUGUAUGUUGCAUUUAATT-3′, and antisense strand
5′-UUAAAUGCAACAUACAAACTT-3′.
The control siRNA provided in the RNAi Human/Mouse
Starter kit (Qiagen) was used for negative control. The AllStars Hs
Cell Death Control siRNA was used as a cell death control. The Cks1
and Cks2 overexpression vectors with the hygromycin-resistance
coding sequences were purchased from Invitrogen.
Cell culture and transfection
HepG2 cells were cultured in DMEM supplemented with
10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin
sulfate at 37°C in a 5% CO2 incubator. Cells were
cultured in 100-mm tissue culture dishes and were subcultured at a
ratio of 1:3 every 2–3 days.
Transfection
HepG2 cells were inoculated in 6-well plates at a
density of 2×105 cells/well just before being
transfected. For gene depletion, siRNAs (300 ng) mixed with 12
µl of HiPerfect transfection reagent was added to each well
of the respective cultured cells. One day following transfection,
the media were replaced with the fresh culture medium and the cells
were cultured at 37°C until being used. For overexpression
experiments, 2.5 µg of each plasmid mixed in 6.5 µl
of Lipofectamine reagent were added to each well. Six hours after
the transfection, fresh culture media were added to each well.
After being cultured at 37°C for 18 h, the cells were subcultured
at a ratio of 1:10. The medium was supplemented with 50
µg/ml hygromycin for selection.
Cell proliferation assays
For CCK-8 cell proliferation assay, the cells were
seeded in 96-well plates at a density of 3×103 cells
well. The cell viability at four selected time-points (24, 48, 72
and 96 h) was analyzed in quadruplicate samples by Cell Counting
Kit-8 on Thermo Fisher Scientific Multiskan FC (Thermo Fisher
Scientific, Waltham, MA, USA). The OD 450 values were measured and
the growth curve was plotted. Alternatively, the cell number was
quantitated with a Sysmex XE-5000 hemocytometer (Kobe, Japan) at
four selected time-points (24, 48, 72 and 96 h) in quadruplicate,
and the growth curve was plotted.
For cell cycle analyses, the cells were
serum-starved for 24 h to synchronization at the G0 phase of the
cell cycle. The cells were then cultured in normal media for 48 h
and stained with PI for fluorescent assisted flow cytometric
analysis.
Assay of cell apoptosis
The cells were seeded in 6-well plates at a density
of 2×105 cells/well. After cells had attached to the
culture plate, cisplatin was added to the control and experimental
groups at a final concentration of 3 µg/ml to induce
apoptosis. The floating and attached cells in each well were
collected at 48 h after the treatment and double-stained with
Annexin V-FITC/PI and Annexin V-EnzoGloden/PI for flow cytometric
analysis.
Real-time PCR
Total RNAs were extracted with TRIzol reagent for
RNA extraction, and the reverse transcription was carried out with
RevertAid First-Strand cDNA synthesis kits. Cks1 and Cks2 primers
and probes were designed using the ABI Primer Express 2 software.
β-actin was used as the internal loading control. The PCR reactions
were performed with the ABI 7500 fluorescent qPCR device.
Western blot analysis
HepG2 cell lysates were prepared by adding 50
µl of RIPA lysis buffer to 106 of cells. The
lysates were separated on SDS-PAGE and transferred to PVDF
membranes for western blot analyses. Mouse anti-human β-actin
antibody was used at a 1:1,000 dilution. Rabbit anti-human Cks1
antibody was used at a 1:1,500 dilution. Rabbit anti-human antibody
was used at a 1:1,000 dilution. Horseradish peroxidase
(HRP)-labeled goat-anti-mouse IgG antibody (targeted to β-actin) or
goat-anti-rabbit IgG antibody (targeted specifically to Cks1 and
Cks2) was used at a 1:10,000 dilution. The specifically bound
antibodies were visualized with the ECL chemiluminescence kit.
Statistical analysis
All data were processed with the SPSS 20.0
statistical software package. The overall data were analyzed with
Student's t-test. Each assay was repeated in triplicate.
Results
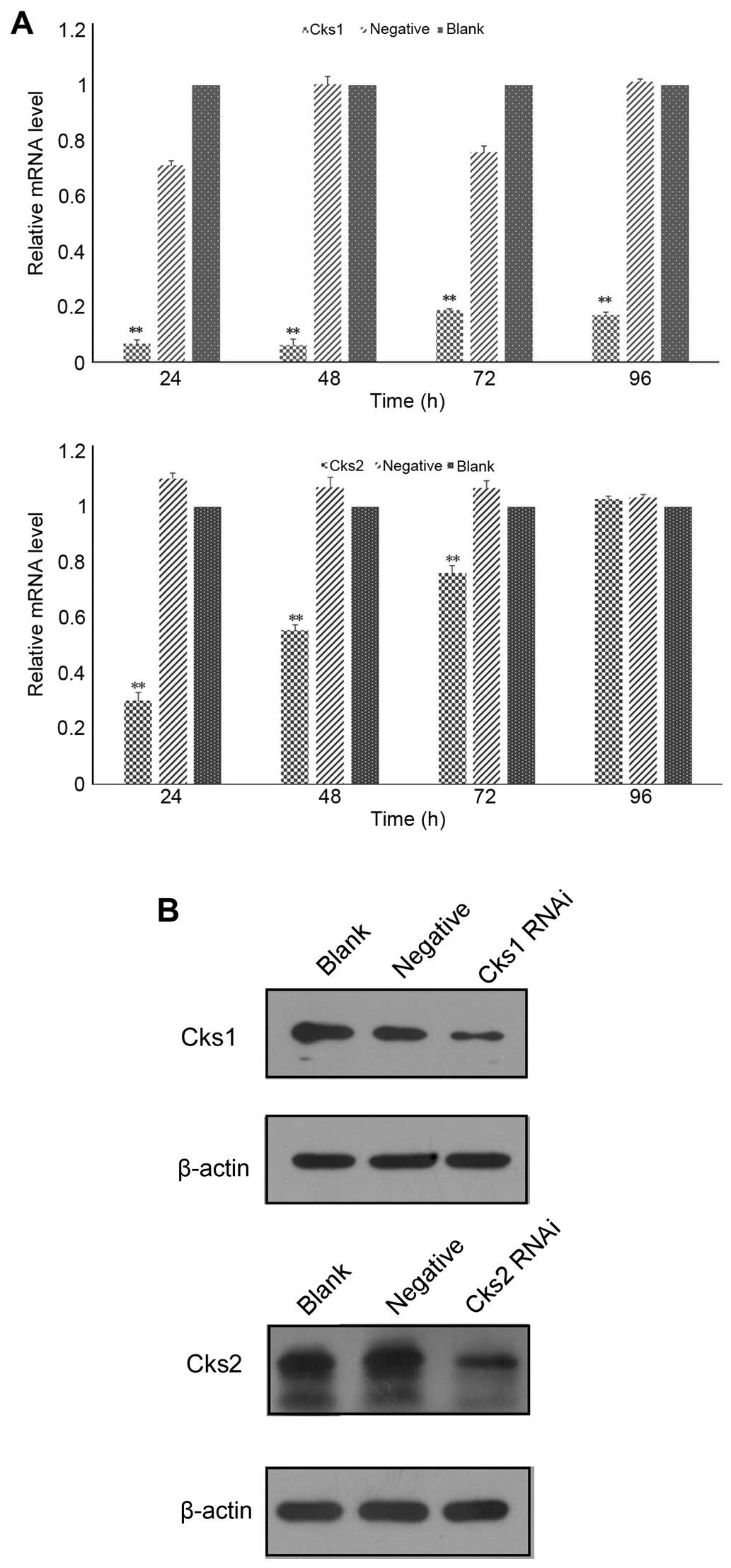
Depletion of Cks1 and Cks2 expression
compromises cell proliferation in HepG2 cells
To assess the role of Cks1/Cks2 in HepG2 cells, the
expression of Cks1/Cks2 was depleted by siRNA specific to Cks1 and
Cks2. The results showed that both Cks1 and Cks2 mRNAs were reduced
with 24 h after the transfection (Fig.
1A). The mRNA levels of Cks1 were downregulated by 95% and Cks2
70%, respectively, at 24 h after the transfection. The knockdown
efficiency gradually decreased over time. Cks1 siRNA maintained a
high knockdown efficiency (81%) even at 96 h after transfection
while the mRNA levels of Cks2 returned to near baseline levels by
96 h after the transfection. Consistently, expression of Cks1 and
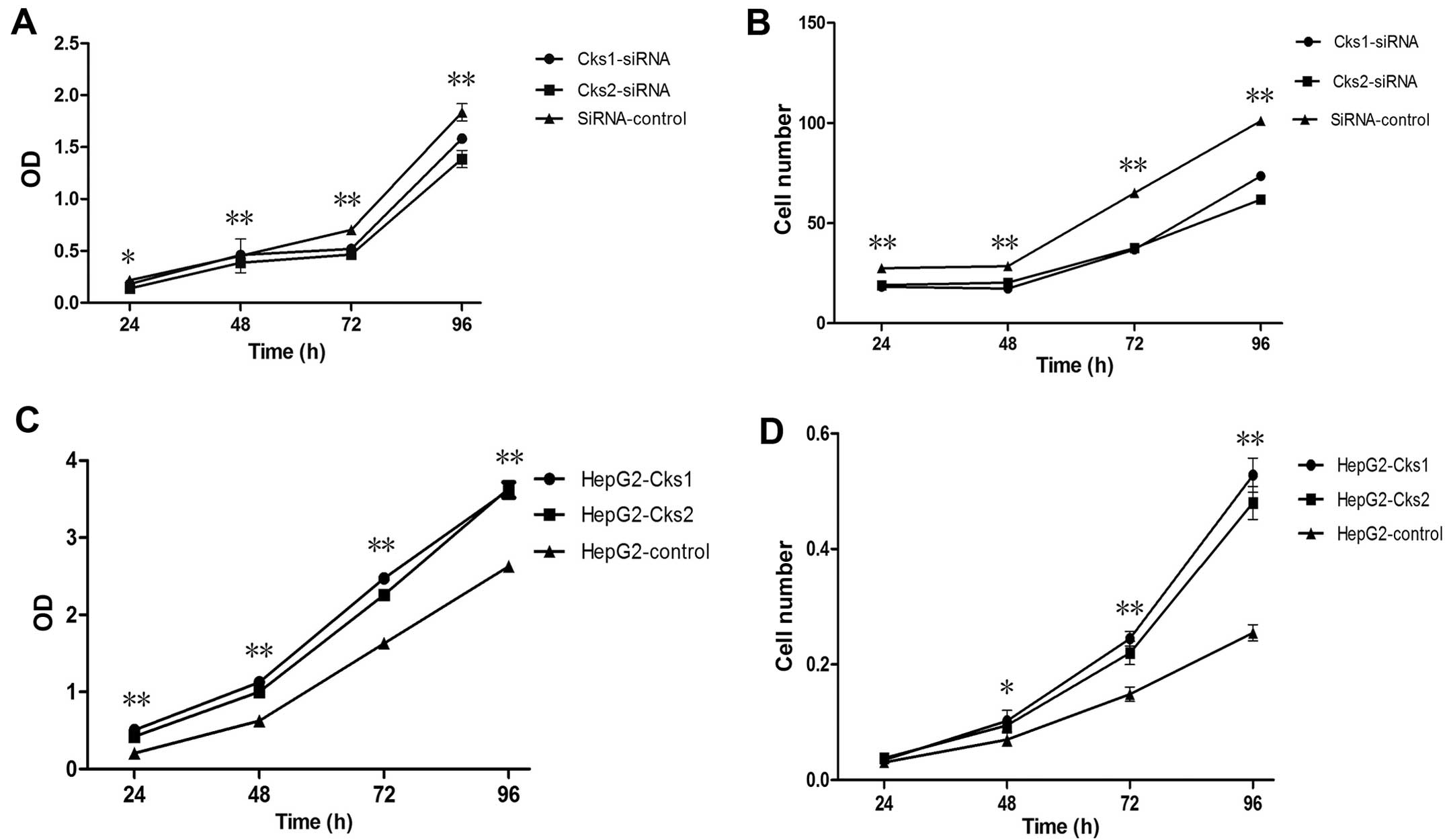
Cks2 at the protein level was also reduced (Fig. 1B). To determine whether depletion of
either Cks1 or Cks2 affects cell proliferation, we used the CCK-8
assay kit to examine cell proliferation at 24, 48, 72 and 96 h
after transfection with Cks1 and Cks2 siRNA (Fig. 2A). It was clear that cell
proliferation was reduced within 24 h after the transfection. The
difference became more obvious over time and remained statistically
significant. Quantitation of cell numbers directly with a cell
counter also revealed that depletion of Cks1 and Cks2 reduced cell
proliferation in HepG2 cells (Fig.
2B). The data suggest that expression of Cks1 and Cks2 has
positive effects on cell proliferation. Consistently,
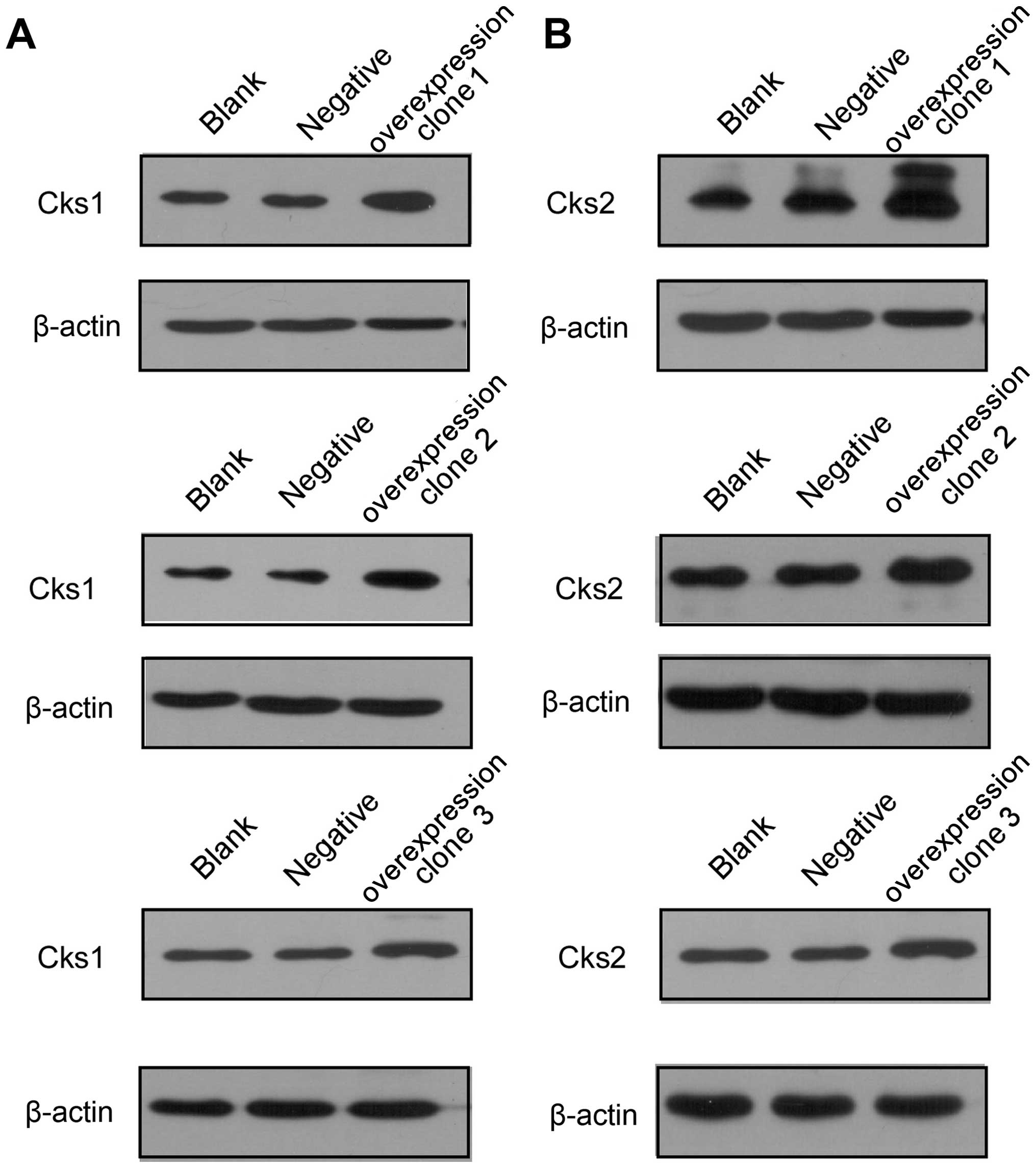
overexpression of Cks1 and Cks2 promote cell proliferation
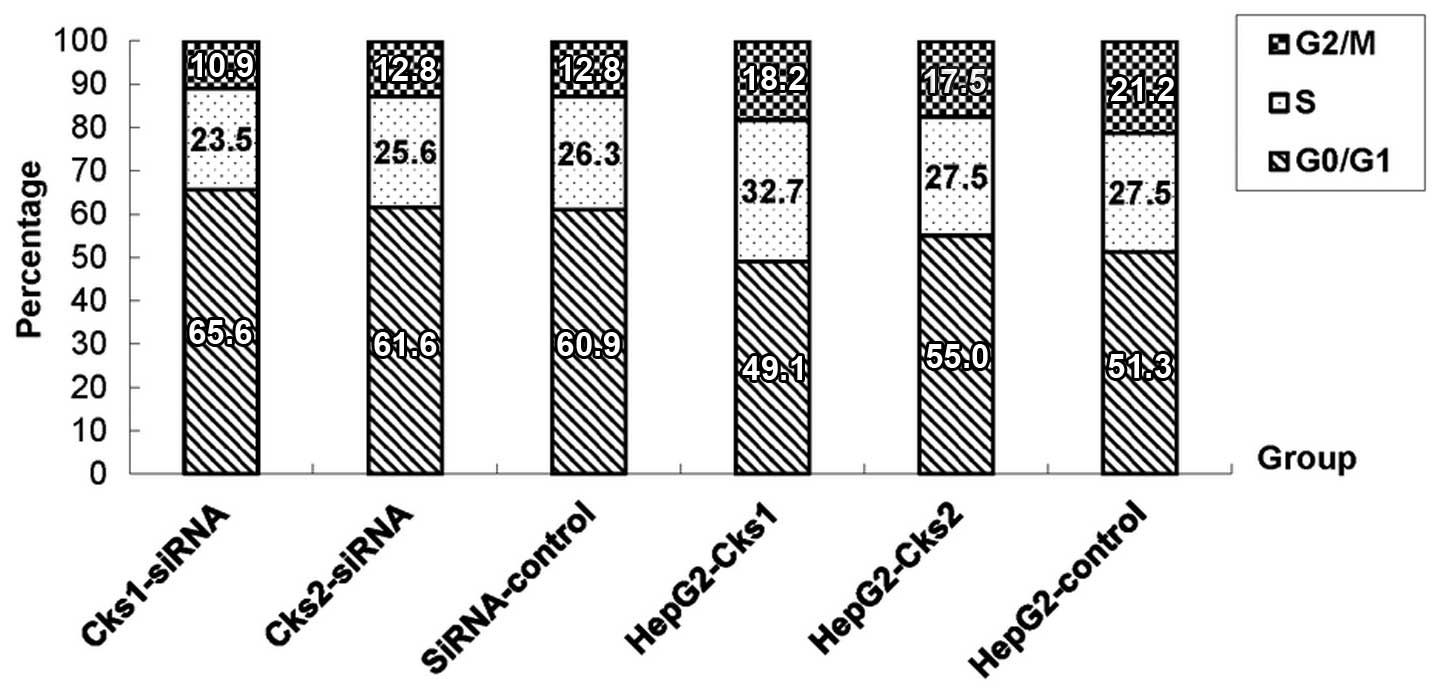
(Figs. 3 and 2C and D). In addition, cell cycle analyses
with the fluorescence-assisted cell sorter (FACS) analyses showed
that overexpression of Cks1 promoted HepG2 cell transition into S
phase (Fig. 4).
Depletion of Cks1 and Cks2 expression
enhances chemotherapy agent-induced apoptosis in HepG2 cells
In order to study the effect of Cks1 and Cks2 on
HepG2 cellular apoptosis, expression of Cks1 and Cks2 was depleted
by siRNA transfection. Twenty-four hours later, the cells were then
treated with cisplatin for another 24 h followed by Annexin V/PI
staining. Flow cytometry was used to analyze the apoptosis rate for
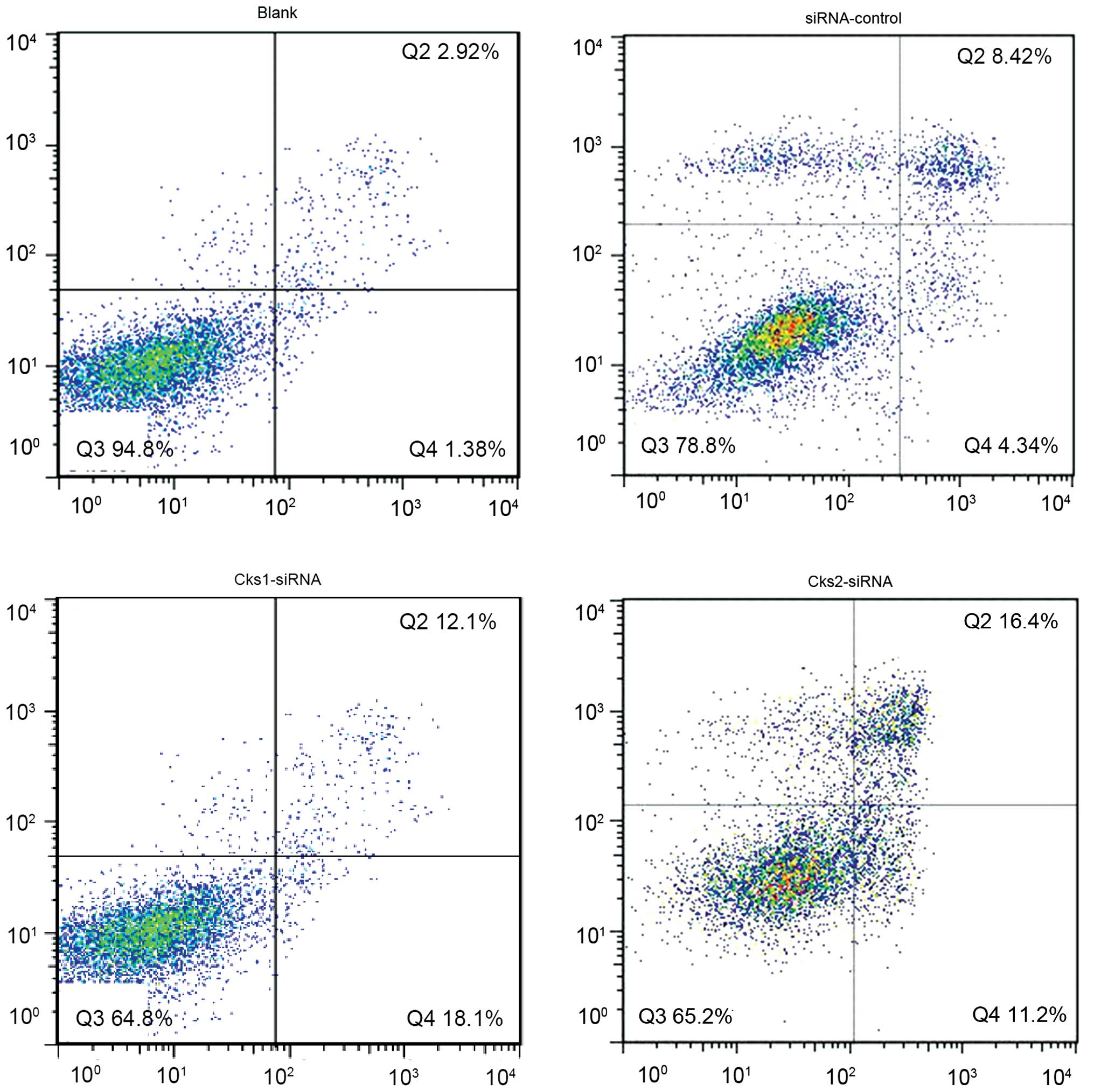
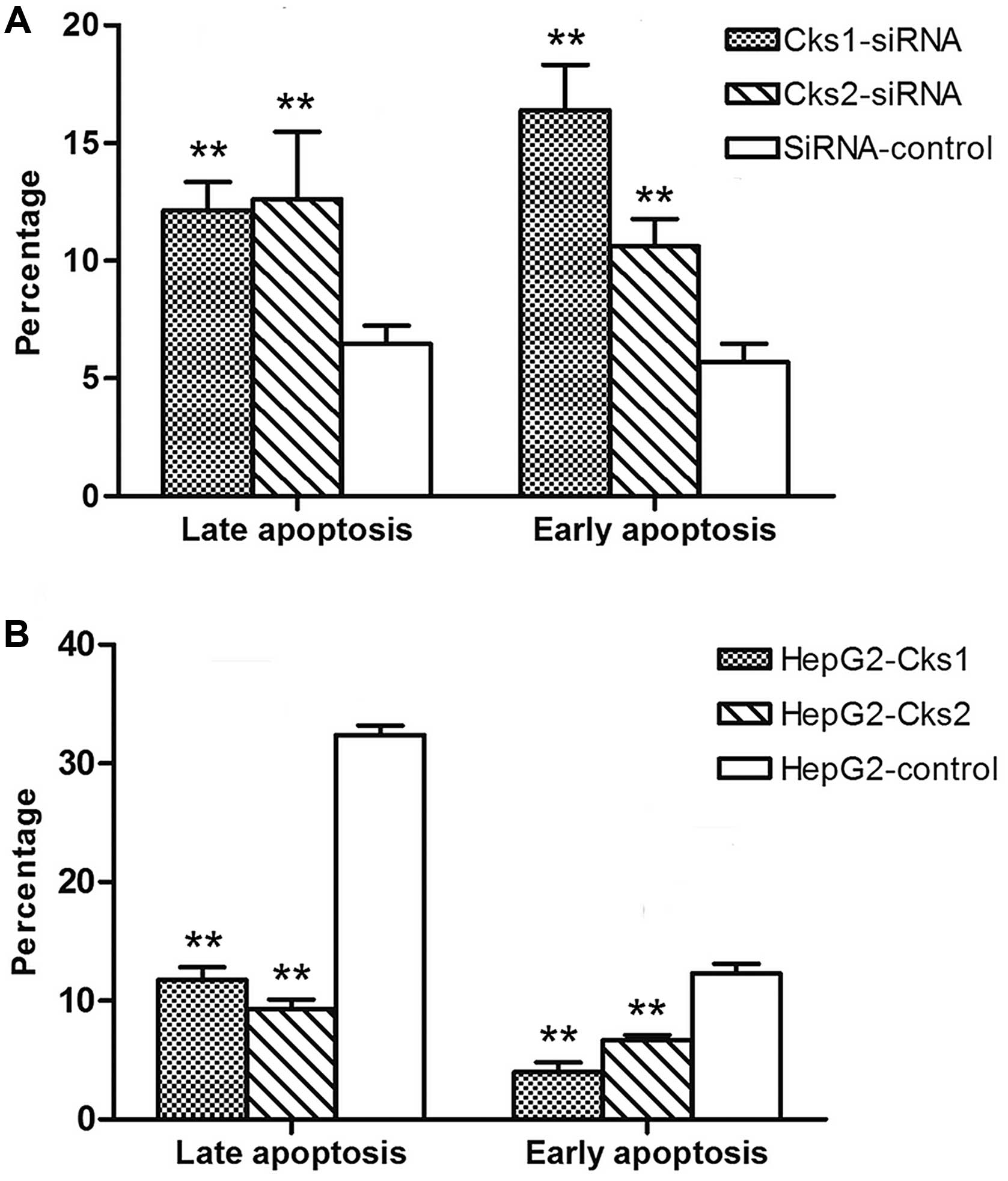
both the control and experimental group (Fig. 5). The results indicate that the
apoptosis rate in Cks1 and Cks2 knockdown groups was higher than
that in the control group upon cisplatin treatment at both early
and late stages (P<0.01, Fig.
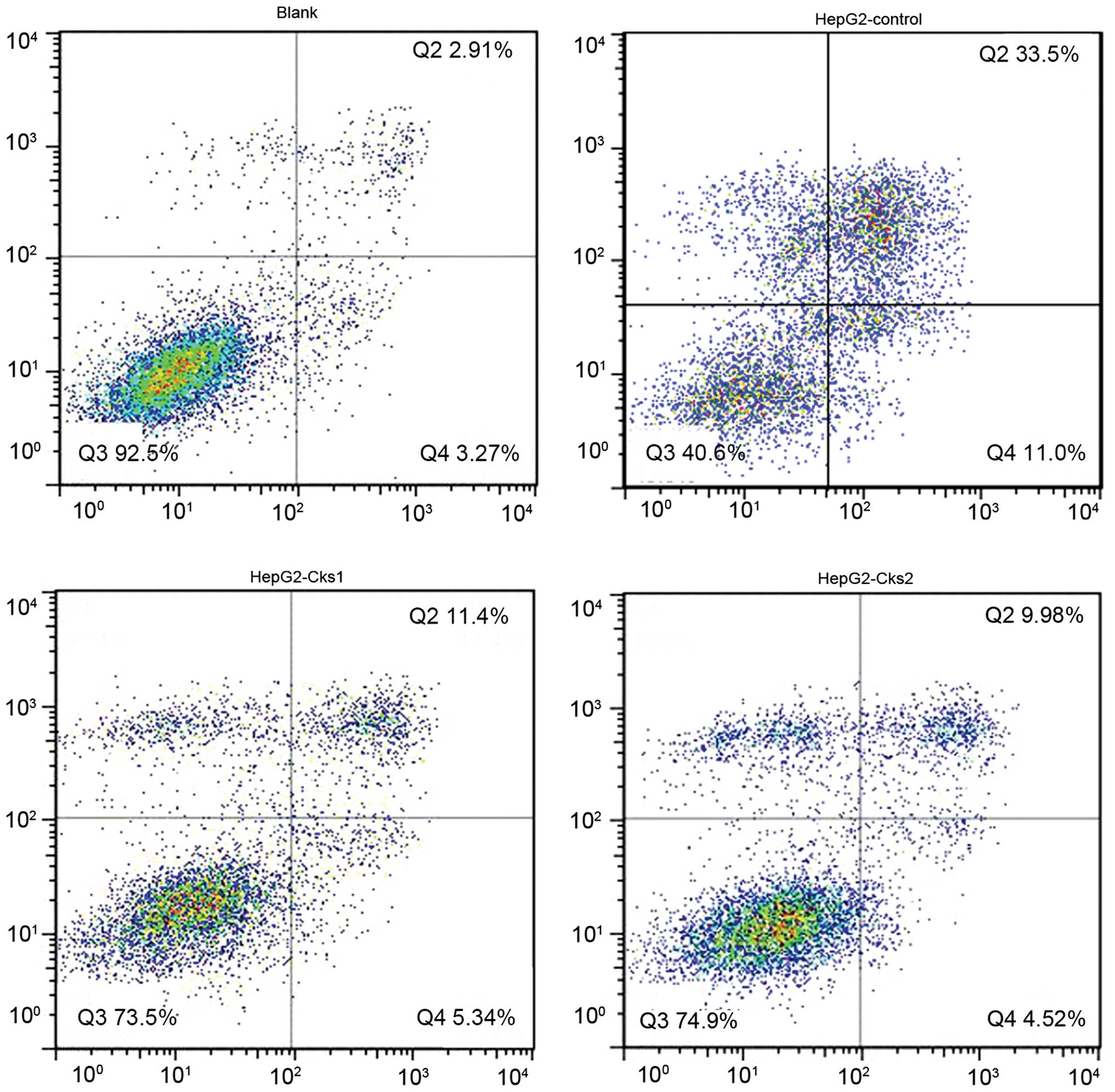
7A). Consistently, overexpression of Cks1 and Cks2 resulted in
a lower cisplatin-induced apoptosis rate at both the early and the
late stages as compared with the control (P<0.01) (Figs. 6 and 7B). Together, the results demonstrate that
high expression level of Cks1 and Cks2 protects the cells from
undergoing apoptosis upon chemotherapy drug treatment.
Depletion of either Cks1 or Cks2 promotes
AKT and GSK-3β phosphorylation in HepG2 cells
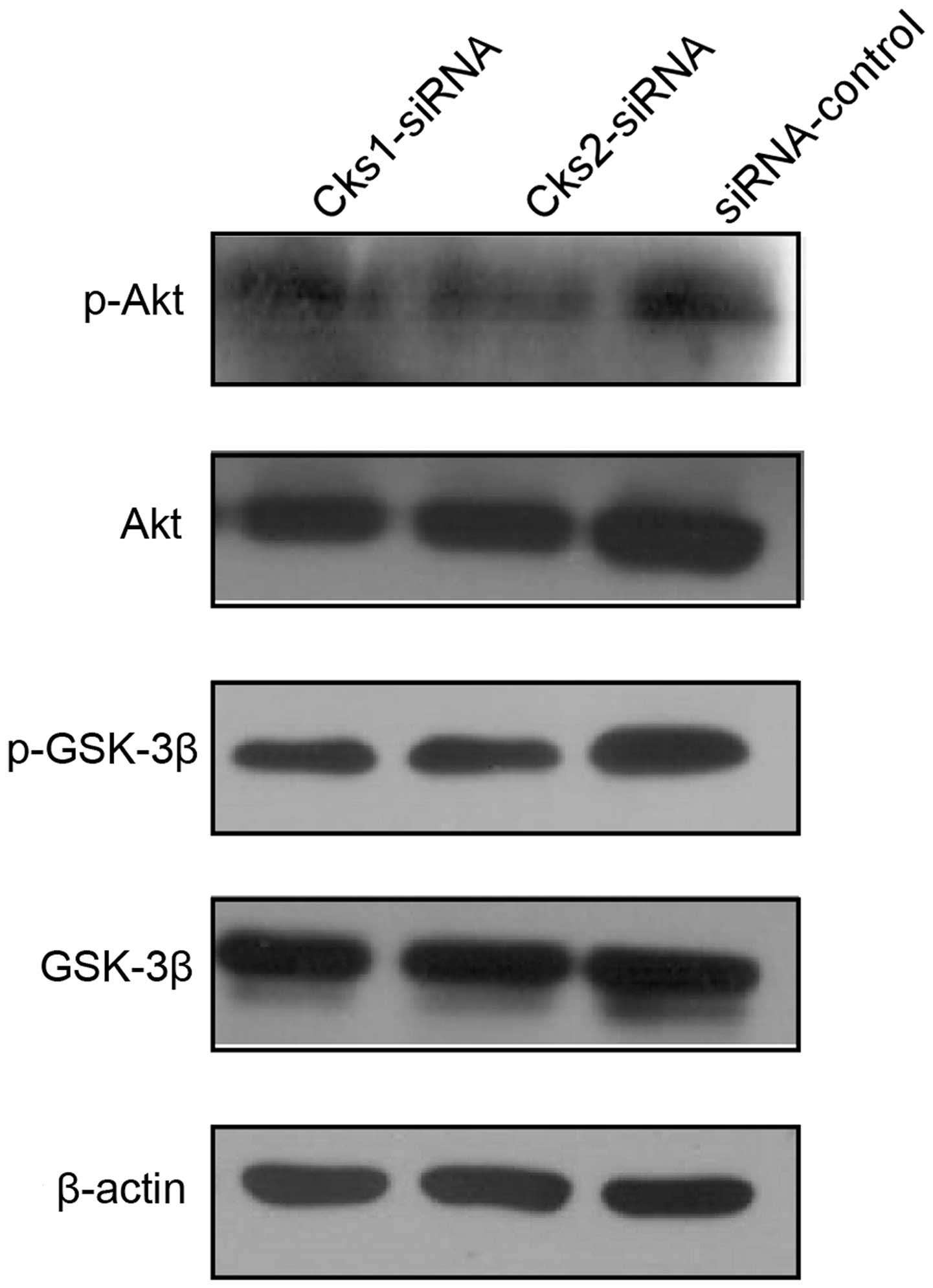
To investigate the mechanism by which depletion of
Cks1 and Cks2 affects cell apoptosis, we then assessed activation
of AKT and GSK activation in Cks depleted cells by western blot
analyses. The results revealed that depletion of either Cks1 or
Cks2 reduced AKT and GSK-3β phosphorylation (Fig. 8). Since activity of AKT and Wnt
signaling has been shown to suppress cell apoptosis, the results
suggested that high expression of Cks proteins protects HCC cells
from apoptosis via the AKT and Wnt pathways.
Discussion
Currently, only a limited number of cancer targeting
drugs are available for HCC treatment (9). However, liver cancer frequently
develops tolerance to the currently available molecular targeting
drugs (10,11). Thus, it is imperative to develop new
targeting drugs for HCC. In the present study, we described that
depleting Cks1 and Cks2 in HepG2 cells reduces cell proliferation
and increase apoptosis induced by the common chemotherapy reagents.
The results suggest that targeting Cks1 and Cks2 in conjunction
with chemotherapy treatment can be a more effective therapy for
HCC.
Although Cks1 promoted p27 ubiquitination and
degradation (12), and therefore,
promotes cell proliferation, the function of Cks2 is largely
unknown in somatic cells. Furthermore, how depletion of Cks1 and
Cks2 increase sensitivity to cancer chemotherapy drugs are still
not clear. Further efforts are needed to characterize the molecular
mechanism by which Cks1 and Cks2 protect cells from undergoing
apoptosis upon chemotherapy treatment. The caspase family of
proteins play a critical role in mediating cellular apoptosis. It
has been reported that Cks2 downregulation reduces cell
proliferation and increase caspase-3 activation and Bax expression
in gastric cancer cells (6).
Depleting Cks2 expression with siRNA in esophageal squamous cell
carcinoma also results in reducing cell proliferation (13).
Activation of Akt via phosphorylation plays an
important role in cell survival and apoptosis (14–16).
It suppresses apoptosis and promotes cell survival, and regulates
glycogen synthesis via GSK-3β and the cell cycle by blocking
GSK-3β-mediated cyclin D phosphorylation and degradation (17). In addition, Akt plays a key role in
cell proliferation. Our results showed that downregulation of Cks1
and Cks2 protein expression led to a decrease in Akt and GSK-3β
phosphorylation. It is speculated that the Cks1- and Cks2-regulated
Akt and GSK-3β signaling activity underlies the oncogenic
activities of both Cks1 and Cks2.
In summary, Cks1 and Cks2 promote HCC cell
proliferation and suppress HCC cell apoptosis induced by
chemotherapy drugs. Therefore, suppressing Cks1 and Cks2 activity
likely will increase efficacy of chemotherapy for HCC, which
deserves further investigation.
Acknowledgments
We appreciate the support of all the staff at the
Central Laboratory of Xiamen University Affiliated Zhongshan
Hospital during the conduct of the experiments. In addition, the
cell lines that we used in this study were generously provided by
the Hepatobiliary Surgery Laboratory at the Xiamen University
Affiliated Zhongshan Hospital. This study was supported by the
National Natural Science Foundation of China (Grant No.
81072016).
Abbreviations:
|
PI
|
propidium iodide
|
|
HCC
|
hepatocellular carcinoma
|
|
Cks
|
cyclin-dependent kinase subunit
|
|
ECL
|
enhanced chemiluminescent
|
|
FACS
|
fluorescence-assisted cell sorter
|
References
|
1
|
Richardson HE, Stueland CS, Thomas J,
Russell P and Reed SI: Human cDNAs encoding homologs of the small
p34Cdc28/Cdc2-associated protein of Saccharomyces cerevisiae
and Schizosaccharomyces pombe. Genes Dev. 4:1332–1344.
1990.
|
|
2
|
Wang JJ, Fang ZX, Ye HM, You P, Cai MJ,
Duan HB, Wang F and Zhang ZY: Clinical significance of
overexpressed cyclin-dependent kinase subunits 1 and 2 in
esophageal carcinoma. Dis Esophagus. 26:729–736. 2013.
|
|
3
|
Lan Y, Zhang Y, Wang J, Lin C, Ittmann MM
and Wang F: Aberrant expression of Cks1 and Cks2 contributes to
prostate tumorigenesis by promoting proliferation and inhibiting
programmed cell death. Int J Cancer. 123:543–551. 2008.
|
|
4
|
Yu M, Zhong M and Qiao Z: Expression and
clinical significance of cyclin kinase subunit 2 in colorectal
cancer. Oncol Lett. 6:777–780. 2013.
|
|
5
|
Lee SW, Kang SB, Lee DS and Lee JU: Akt
and Cks1 are related with lymph node metastasis in gastric
adenocarcinoma. Hepatogastroenterology. 60:932–937. 2013.
|
|
6
|
Tanaka F, Matsuzaki S, Mimori K, Kita Y,
Inoue H and Mori M: Clinicopathological and biological significance
of CDC28 protein kinase regulatory subunit 2 overexpression in
human gastric cancer. Int J Oncol. 39:361–372. 2011.
|
|
7
|
Shen DY, Zhan YH, Wang QM, Rui G and Zhang
ZM: Oncogenic potential of cyclin kinase subunit-2 in
cholangiocarcinoma. Liver Int. 33:137–148. 2013.
|
|
8
|
Shen DY, Fang ZX, You P, Liu PG, Wang F,
Huang CL, Yao XB, Chen ZX and Zhang ZY: Clinical significance and
expression of cyclin kinase subunits 1 and 2 in hepatocellular
carcinoma. Liver Int. 30:119–125. 2010.
|
|
9
|
Li F, Zhao C and Wang L:
Molecular-targeted agents combination therapy for cancer:
Developments and potentials. Int J Cancer. 134:1257–1269. 2014.
|
|
10
|
Ricke J, Bulla K, Kolligs F,
Peck-Radosavljevic M, Reimer P, Sangro B, Schott E, Schütte K,
Verslype C, Walecki J, et al: SORAMIC study group: Safety and
toxicity of radioembolization plus sorafenib in advanced
hepatocellular carcinoma: Analysis of the European multicentre
trial SORAMIC. Liver Int. 35:620–626. 2015.
|
|
11
|
Zhu AX, Duda DG, Ancukiewicz M, di Tomaso
E, Clark JW, Miksad R, Fuchs CS, Ryan DP and Jain RK: Exploratory
analysis of early toxicity of sunitinib in advanced hepatocellular
carcinoma patients: Kinetics and potential biomarker value. Clin
Cancer Res. 17:918–927. 2011.
|
|
12
|
Ganoth D, Bornstein G, Ko TK, Larsen B,
Tyers M, Pagano M and Hershko A: The cell-cycle regulatory protein
Cks1 is required for SCF(Skp2)-mediated ubiquitinylation of p27.
Nat Cell Biol. 3:321–324. 2001.
|
|
13
|
Kita Y, Nishizono Y, Okumura H, Uchikado
Y, Sasaki K, Matsumoto M, Setoyama T, Tanoue K, Omoto I, Mori S, et
al: Clinical and biological impact of cyclin-dependent kinase
subunit 2 in esophageal squamous cell carcinoma. Oncol Rep.
31:1986–1992. 2014.
|
|
14
|
Franke TF, Kaplan DR and Cantley LC: PI3K:
Downstream AKTion blocks apoptosis. Cell. 88:435–437. 1997.
|
|
15
|
Burgering BM and Coffer PJ: Protein kinase
B (c-Akt) in phos-phatidylinositol-3-OH kinase signal transduction.
Nature. 376:599–602. 1995.
|
|
16
|
Franke TF, Yang SI, Chan TO, Datta K,
Kazlauskas A, Morrison DK, Kaplan DR and Tsichlis PN: The protein
kinase encoded by the Akt proto-oncogene is a target of the
PDGF-activated phosphatidylinositol 3-kinase. Cell. 81:727–736.
1995.
|
|
17
|
Hajduch E, Litherland GJ and Hundal HS:
Protein kinase B (PKB/Akt) - a key regulator of glucose transport?
FEBS Lett. 492:199–203. 2001.
|