Gastric cancer is the second leading cause of
cancer-related mortality worldwide and is the most common
malignancy in China and Japan, affecting approximately one million
individuals every year (1,2). The highest ratio, up to 69 cases per
100,000 individuals per year, has been determined in males in
Northeast Asia (3). Prevention and
personalized treatment are regarded as the best options to reduce
gastric cancer mortality rates. Gastric cancer is usually a result
of a high intake of various traditional salt-preserved foods and
salt, concomitant with a low consumption of fresh fruit and
vegetables (4,5). Analysis of the signaling pathways in
cancer cells provides novel biomarkers for diagnosis and drug
targets for treatment.
miRNAs are a novel class of small non-coding RNAs,
typically 22 nucleotides in length (6). miRNAs suppress gene expression by
directly binding to mRNAs, causing translation repression or mRNA
cleavage (7,8). They are single-stranded RNAs that
negatively regulate gene expression at the post-transcriptional
level (6,9,8).
Longer precursor transcripts with hairpin structures are first
synthesized by RNA polymerase II, followed by processing of
precursors by Drosha and Dicer. To date, miRNAs have been found to
be involved in various physiological and pathological processes,
including cell metabolism, tumorigenesis, cell growth and
apoptosis, aging, organic development and the immune response
(10,11). A single RNA may have hundreds or
more targets, therefore it is difficult to distinguish functions to
specific miRNAs.
Studies have demonstrated that miRNAs can function
as oncogenes or tumor suppressors by regulating the expression of
cancer-related genes (12). During
the latest decade, a set of cancer regulator miRNAs have emerged
and these are divided into oncomiRs or anti-oncomiRs. Specific
miRNA profiles have been observed in both tumor tissues and plasma
in various types of cancers (13–16).
These miRNAs are believed to be potential biomarkers for the
diagnosis and may also support the prognosis or treatment of cancer
(17). Recent projects have
attempted to decipher the differential expression of miRNAs in
specific cancers; however, the complex pathways comprising miRNAs
and their target genes remain unclear.
Identification of the molecular causes of cancer
represents a major breakthrough in the history of medicine, moving
the discipline from pattern recognition and therapeutic strategies
based on molecular mechanisms and therapies derived from clinical
trials (18). The development of
pathway strategies for the analysis of gastric cancer makes it
possible to use these approaches for future clinical treatment.
We previously reported that 5 significant miRNAs are
overexpressed in gastric cancer and serve as a fingerprint for
gastric cancer diagnosis (14). In
the present study, the protein classes, molecular functions,
biological functions and canonical pathways comprising the targets
of each gastric cancer-related miRNA as well as four main canonical
pathways were identified and analyzed, offering novel drug targets
for gastric cancer therapy.
The web-based functional annotation tool Database
for Annotation, Visualization and Integrated Discovery (DAVID) v6.7
(http://david.abcc.ncifcrf.gov/tools.jsp) has key
components for disease analysis, gene ontology analysis and pathway
analysis (22).
The signaling pathways and processes were explored
using the systems biology tool KEGG Mapper (http://www.genome.jp/kegg/tool/map_pathway2.html)
which is a collection of tools for KEGG mapping: KEGG pathway
mapping, BRITE mapping and MODULE mapping (23). The KEGG database consists of 16 main
databases (systems information, KEGG PATHWAY, KEGG BRITE, KEGG
MODULE, KEGG DISEASE, KEGG DRUG and KEGG ENVIRON; genomic
information, KEGG ORTHOLOGY, KEGG GENOME, KEGG GENES, KEGG SSDB and
KEGG; chemical information, KEGG COMPOUND, KEGG GLYCAN, KEGG
REACTION, KEGG RPAIR, KEGG RCLASS and KEGG ENZYME).
Based on the experimental data in our previous
study, 5 miRNAs were selected as GC-related miRNAs (Table I). These miRNAs were found to be
clearly increased in GC, and their dysregulation is believed to
promote tumorigenesis.
miR-20a belongs to the miR-17-92 cluster, also known
as oncomiR-1, which includes 6 microRNAs: miR-17-5p, miR-18a,
miR-19a, miR-19b, miR-20a and miR-92a-16 (35). It contributes to the regulation of
many types of tumors as a tumor-suppressor in oral squamous cell
cancer (36) and hepatic cancer
(37) or a tumorpromotor in
osteosarcoma (38), bladder cancer
(39), GC (40), prostate cancer (41) and cervical cancer (42). It promotes the growth, migration,
and invasion of GC cells by inhibiting the early growth response 2
(EGR2) signaling pathway (40), and
may be related with the malignant process of cervical cancer,
particularly invasion and metastasis by targeting autophagy-related
protein 7 (ATG7), tissue inhibitors of metalloproteinase 2 (TIMP2)
and tankyrase 2 (TNKS2) (43). On
the other hand, miR-20a is involved in the tumor inhibition of
cutaneous squamous cell carcinoma (CSCC) by targeting LIM kinase-1
gene (LIK1), a metastasis promoter (44). Moreover, it targets MHC class I
chain-related molecule A and B (MICA/B) to avoid NKG-mediated
immune attack, thus enhancing the survival of ovarian cancer cells
by immune escape (45). It has been
shown that miR-20a inhibits the proliferation and metastasis of
pancreatic carcinoma cells by directly downregulating Stat3 which
is related to various physiological functions (46).
Previous studies have demonstrated that miR-27a acts
as an oncogenic miRNA. Its role in promoting cell proliferation,
invasion and metastasis has been verified in many malignancies,
such as breast cancer (47), HCC
(48), non-small cell lung cancer
(NSCLC) (49), osteosarcoma
(50) and renal cancer (51). In HCC, miR-27a promotes cell
proliferation through suppression of its target gene peroxisome
proliferator-activated receptor γ (PPAR-γ) (48). Direct and indirect mechanisms by
which miR-27a can regulate both MET and EGFR, thus contributing to
tumor progression, was discovered in NSCLC (49). Other studies emphasize the role of
miR-27a expression in sensitivity to anticancer therapies,
including chemotherapy (52–58),
radiotherapy (59) and thermal
therapy (60). Downregulation of
miR-27a is significantly associated with the expression of
P-glycoprotein and multidrug resistance gene 1 (MDR1), leading to
increased chemosensitivity through different targets, for example,
FZD7/β-catenin pathway (52),
homeodomain-interacting protein kinase 2 (HIPK2) (54) and RUNX1 (53). Notably, miR-27 can indirectly affect
chemosensitivity by acting on the tumor microenvironment through
transformation of normal fibroblasts to cancer-associated
fibroblasts (55). Single
nucleotide polymorphism rs11671784 is in the loop of pre-miR-27a
and the G/A variation can significantly decrease the expression of
mature miR-27a, followed by increased RUNX-1 expression and
weakened chemosensitivity (53).
CDC27 is a target of miR-27a by which radiosensitivity is modulated
in triple-negative breast cancer (59). miR-27a may even contribute to
thermal sensitivity by modulating HSP expression (60).
miR-34a is one of the earliest known tumor
suppressors and is commonly downregulated in various solid cancers
by targeting numerous oncogenes related to proliferation, apoptosis
and invasion (61–68). miR-34a is downregulated in many
cancers due to chromosomal deletion or CpG island methylation
(69). As a direct transcriptional
target of p53, decreased expression of miR-34a is partially due to
mutations of p53 in several tumors (70). Ectopic expression of miR-34a can
lead to cell cycle arrest, apoptosis or senescence, mimicking p53
activation (71). And miR-34a can
suppress tumor metastasis via the downregulation of MET, meanwhile
it is associated with the clinicopathological features of gastric
carcinoma and can be a valuable predictor of patient prognosis
(67). Epithelial-mesenchymal
transition (EMT) is a key step in tumor progression. miR-34a
inhibits EMT by targeting Smad4 through the transforming growth
factor-β/Smad pathway in human cholangiocarcinoma (72). The role of miR-34a in regulating
tumor biology has been extensively studied, while being an
effective biomarker is another function of miR-34a. Serum miR-34a
can be a potentially useful diagnostic biomarker of pancreatic
ductal adenocarcinoma (73).
Furthermore, miR-34a is likely to be involved in the treatment
response of lung metastases of HCC to sorafenib (74).
The miR-423 family, identified as oncogenic miRNAs,
has been explored in various types of cancers. There are two
members of the miR-423 family, miR-423-3p and miR-423-5p. miR-423
plays a role in promoting cell growth and trefoil factor 1 (TFF1)
in GC cells (77). The expression
level of miR-423-5p is aberrant in many cancers, including GC
(14), colon carcinoma (78), pancreatic cancer (79) and breast cancer (80). A five-microRNA signature identified
from genome-wide serum microRNA expression profiling, including
miR-423-5p, can serve as a fingerprint for GC diagnosis (14). In stage I–II colorectal cancer,
serum miR-423-5p was found to be significantly elevated compared
with a healthy control, suggesting its value as a tool for early
diagnosis (78). Furthermore, other
than the diagnostic value of miR-423-5p, its role in prediction of
the cancer therapeutic effect has been discovered. A classifier
consisting of seven miRNAs, including overexpression of miR-423-5p,
was able to identify a subgroup of glioblastoma patients who were
resistant to temozolomide (81). On
the contrary, the elevation of secretory miR-423-5p can be a
favorable marker of the effect of sorafenib in HCC patients
(76).
A single miRNA has hundreds of potential targets in
physiological and pathological conditions; therefore, investigation
of miRNA-target genes provides a better description of the
miRNA-involved pathways. All of the predicted targets should be
analyzed to fully understand the functions of GC-related miRNAs. As
shown in Table I, GC miRNAs or
miRNA family has the ability to directly target between 180 and
1,218 mRNAs of genes; moreover, a unique miRNA was observed to have
multiple binding sites in the 3′UTR of the mRNAs. In total, 4,032
genes are regarded as downstream targets of the 5 significant GC
miRNAs. Most of the genes are potential targets of oncomirs, and
these genes are most likely to be downregulated in GC cells. Among
all of the predicted genes, several important regulators include:
BCL3, CD69, VIP, BMP3, MAPK1, BCL9L, BCL11B, PTeN; these genes are
well known to be involved in cell apoptosis, cell proliferation,
cell metastasis and angiogenesis.
In agreement with our research, some groups have
reported that miRNA-related oncogenes and tumor suppressors clearly
show different patterns in function, expression, chromosome
distribution, molecule size, free energy, targets and transcription
factors (82–85).
To provide a direct look at the pathways implicated
in all targets of the 5 miRNAs, all the targets were used for
further pathway analysis. As shown in Table II, the important molecular function
and biological processes are almost identical to the potential
targets of the 5 miRNAs. miR-1 contributes to the biological
process of binding (34.4%), catalytic activity (27.4%), nucleic
acid binding transcription (12%), transporter activity (6.4%),
enzyme regulator activity (6.4%) and receptor activity (6.4%).
miR-20a-related targets play a role in binding (35.8%), catalytic
activity (28.6%), nucleic acid binding transcription (13.1%),
enzyme regulator activity (6.8%) and receptor activity (5.3%). The
potential target genes of miR-27a are also involved in binding
(32.4%), catalytic activity (29.1%), nucleic acid binding
transcription (12.1%), receptor activity (7.6%) and enzyme
regulator activity (7.2%). The miR-34a-related genes are expected
to contribute to binding, catalytic activity, nucleic acid binding
transcription, enzyme regulator activity and receptor activity.
Furthermore, the target genes of miR-423-5p participate in binding
(34.5%), catalytic activity (25.2%), nucleic acid binding
transcription (16%), receptor activity (6.8%) and transporter
activity (Table II).
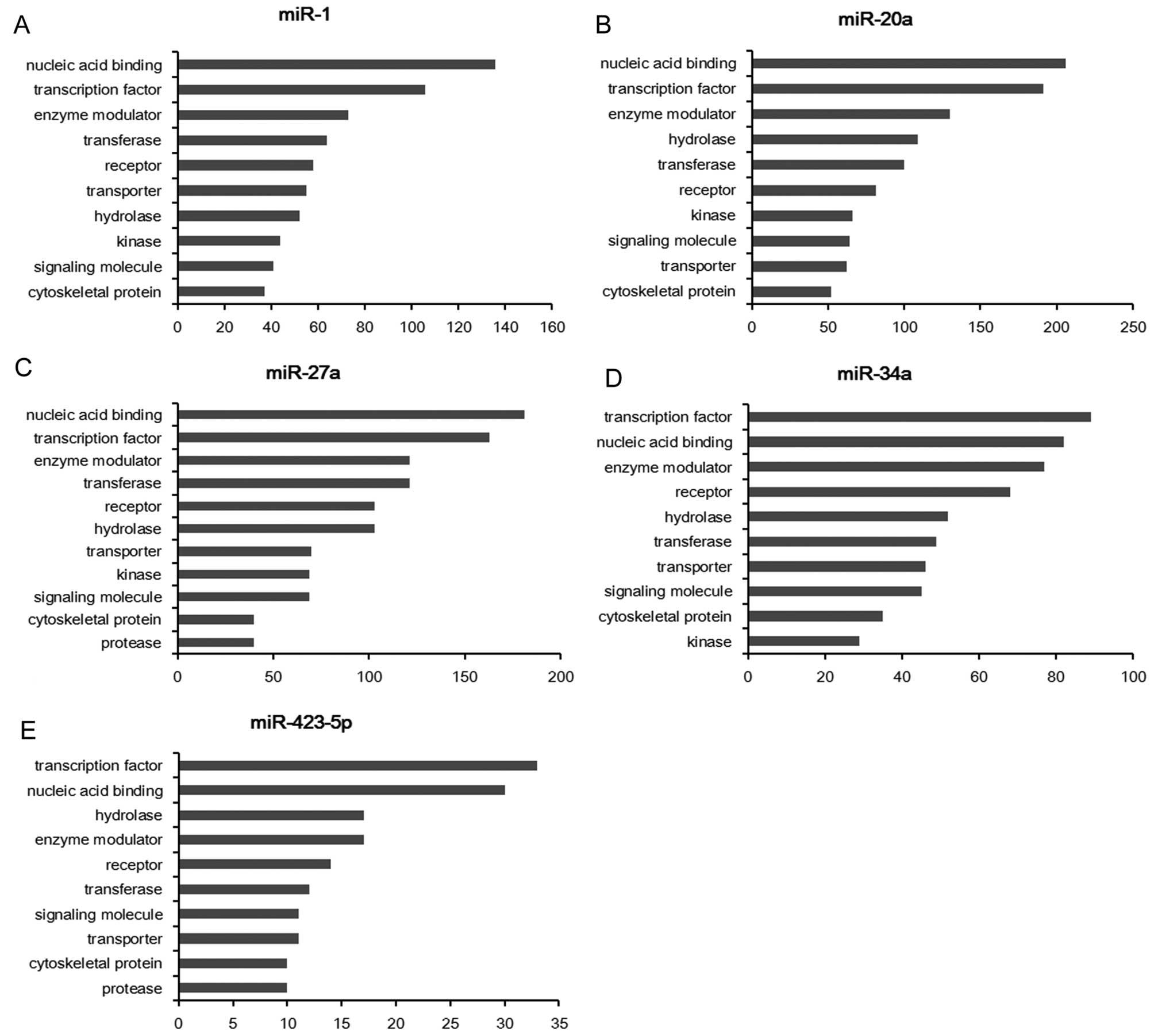
The potential target genes of each miRNA were
classified into several main groups, and the top 10 classes are
respectively shown in Table III.
miR-1, miR-27a, miR-34a and miR-423-5p are found to participate in
the gonadotropin release hormone receptor pathway; miR-1, miR-20a,
miR-27a and miR-34a are involved in the pathway of angiogenesis;
miR-1, miR-27a and miR-423-5p are involved in Wnt signaling pathway
(Table III). As predicted, miR-1
contributes to the pathways of EGF receptor signaling, VEGF
signaling, PDGF signaling and FGF signaling, thus, the
dysregulation of miR-1 is believed to play the most important role
in the tumorigenesis in gastric cancer.
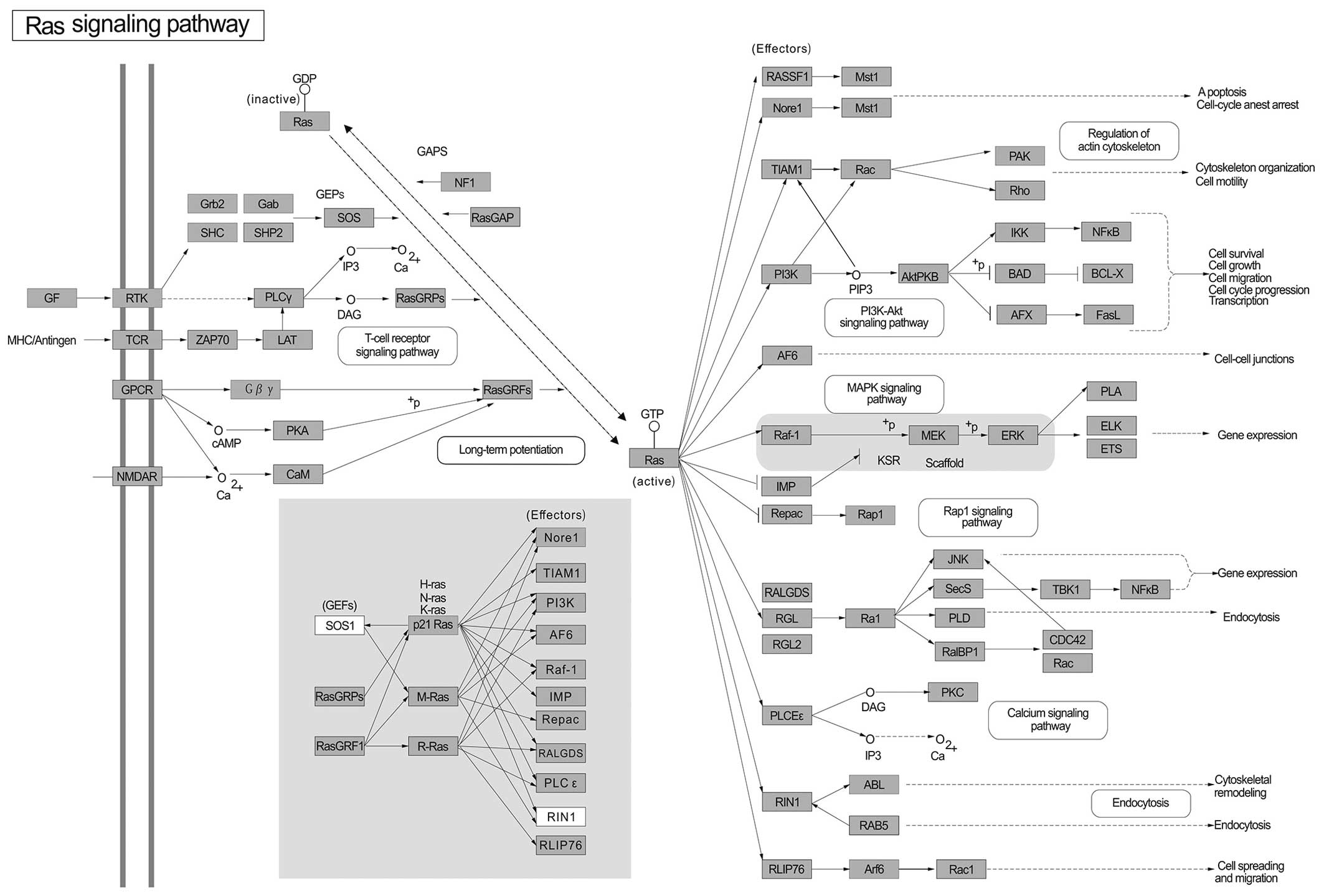
To give a direct view of the GC miRNA-related genes,
all of the targets of miR-1, miR-20a, miR-27a, miR-34a and
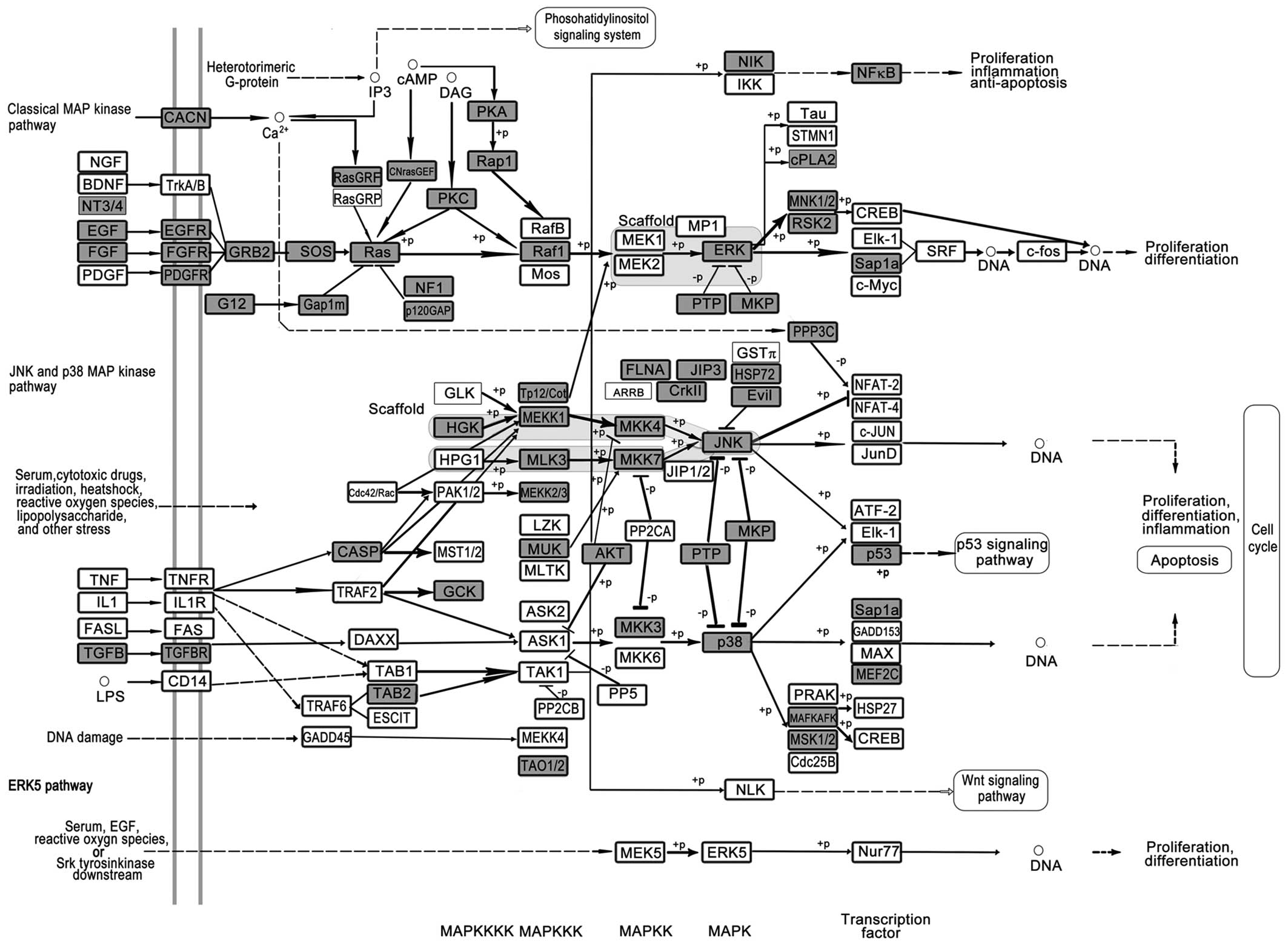
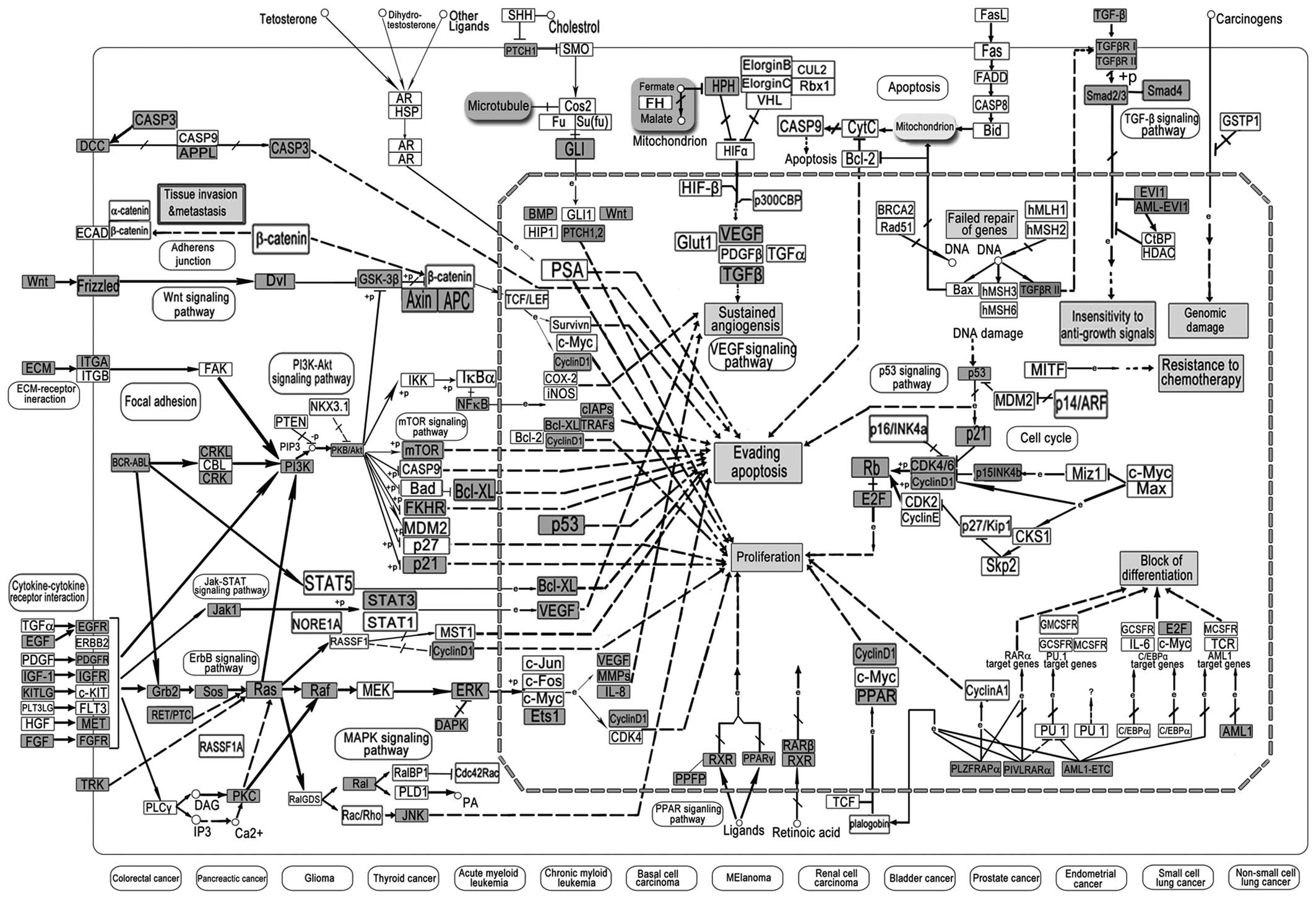
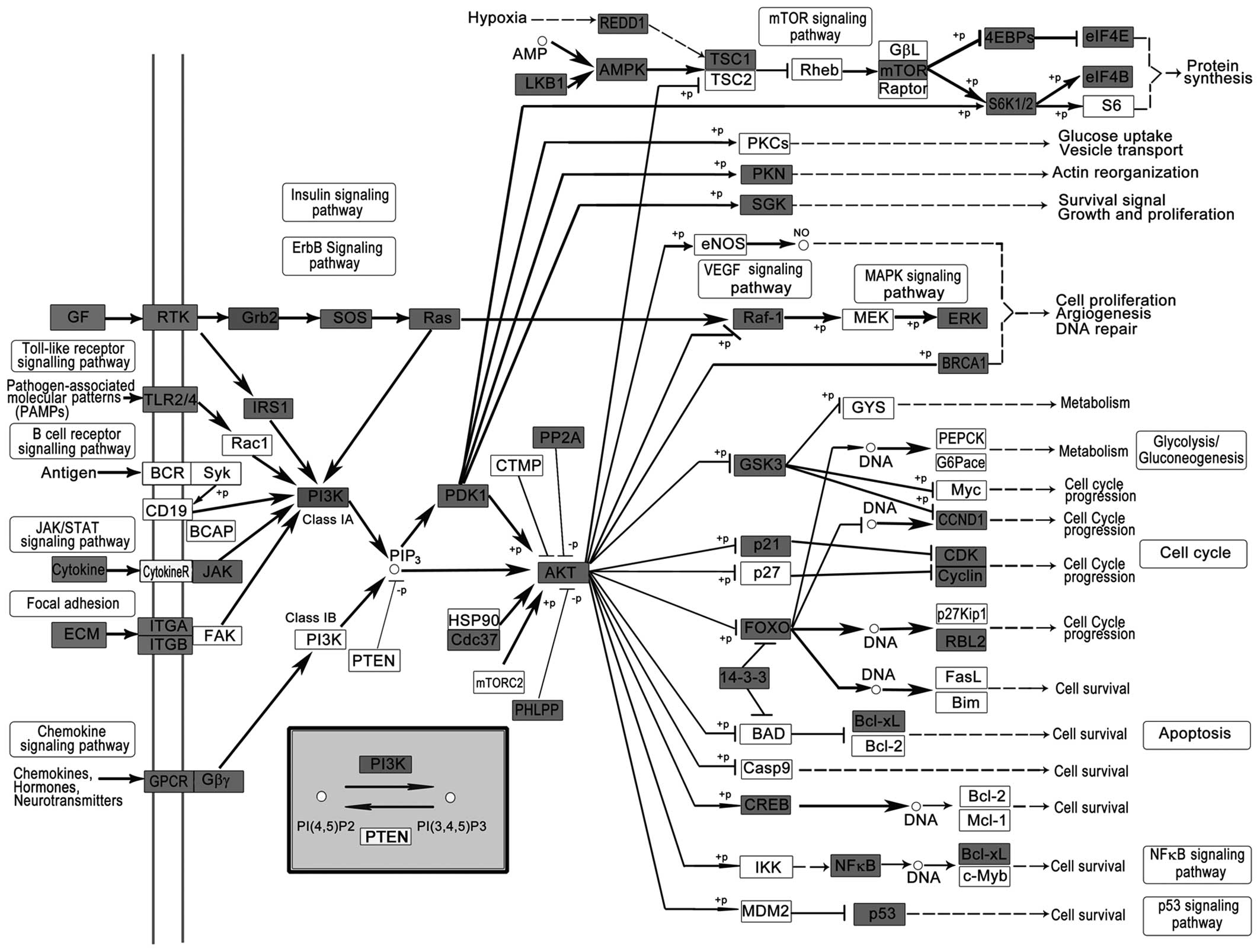
miR-423-5p were used for further pathway analysis. The 5 most
important pathways include the MAPK signaling pathway (Fig. 2), PI3K-Akt signaling pathway
(Fig. 3), pathways in cancer
(Fig. 4), HTLV-I infection
(Fig. 5) and the Ras signaling
pathway (Fig. 6). These pathways
are well known to play important roles in cell growth, cell
metastasis, cell invasion and intercellular communication in
various types of cancer (86–92).
Pathways in cancer are more closely linked with
tumorigenesis, and include the MAPK, PI3K-Akt, VEGF, p53, PPAR and
TGF-β signaling pathways (Fig. 3).
As shown in Fig. 3, the pathways in
cancer have demonstrated the acquisition of biological capabilities
such as blockade of differentiation, resistance to apoptosis,
unlimited replicative potential, sustained angiogenesis, tissue
invasion and metastasis for the transformation of normal cells into
highly malignant tumor cells. The important GC-miRNA-related genes
are Wnt, STAT3, p21, p53, BCL-2, FAS and TGF-β; however, more
common abnormalities in oncogenes and tumor-suppressor genes
regulated by GC miRNAs can be used as potential therapeutic targets
(Fig. 3).
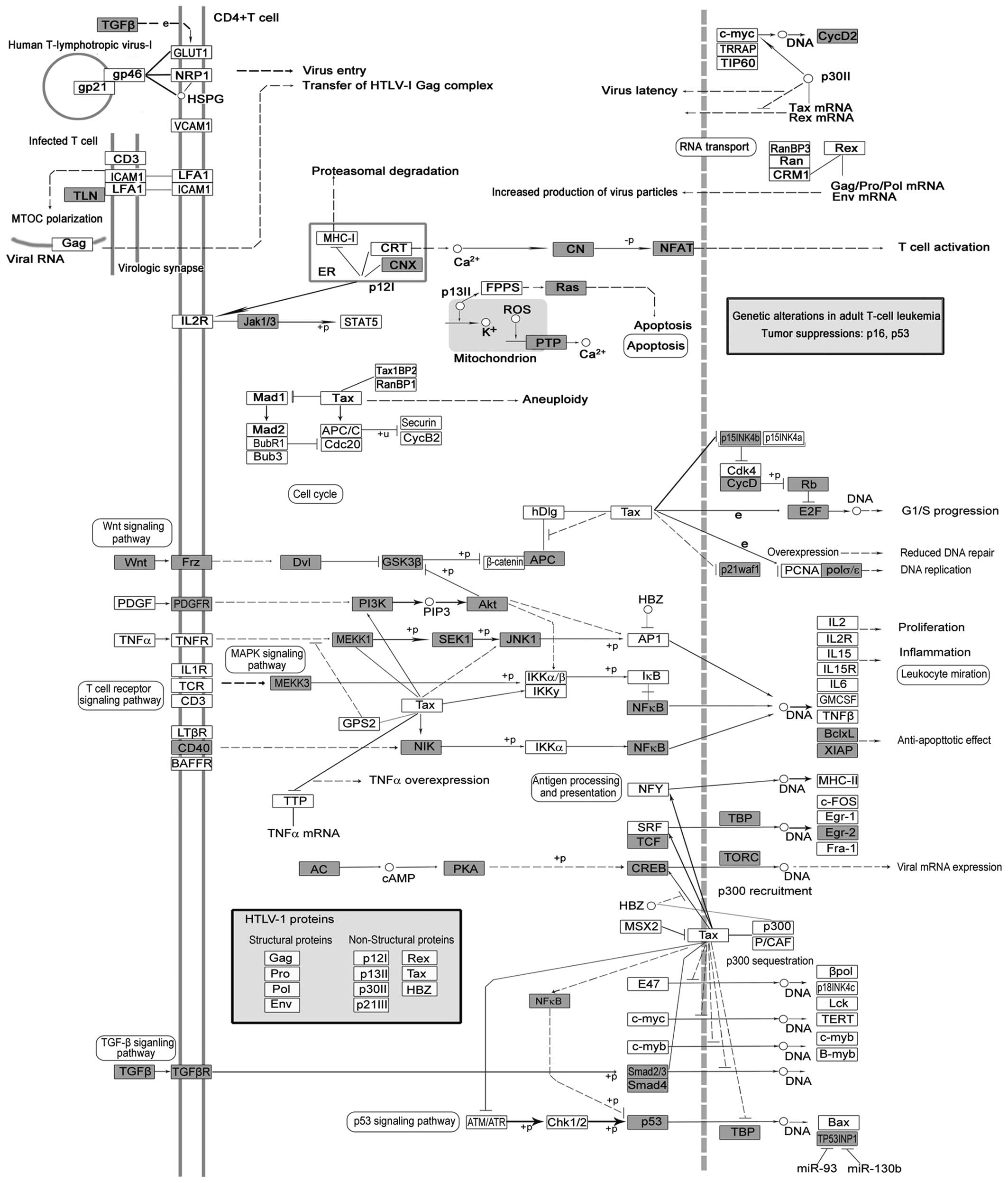
Human T cell leukemia virus 1 (HTLV-1) is a
retrovirus that causes adult T cell leukemia (ATL) and neurological
disorder, the tropical spastic paraparesis (HAM/TSP) (Fig. 5). The pathogenesis apparently
results from the pleiotropic function of Tax protein, which is a
key regulator of viral replication (Fig. 5). Tax encoded by HTLV-1 has been
implicated in tumorigenesis, and is involved in the dysregulation
of anti-apoptosis and cell proliferation (108). Recently, miR-28-3p has been
reported to be a cellular restriction factor that inhibits HTLV-1
replication and infection (109).
However, the role of miRNA and HTLV-1 in cancer, particularly in
GC, remains unclear.
In the present study, we constructed a detailed
biological frame by in-depth analysis of the complex network
comprising GC-miRNA-related targets. These results are believed to
provide a better understanding of the miRNA-regulated pathways in
gastric cancer and identify novel potential targets for future
clinical use.
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81201946 and
81372394) and the National Research Platform of Clinical evaluation
Technology for New Anticancer Drugs (no. 2013ZX09303001). This
study was also supported by the Chinese Ministry of education
Research Fund for doctoral programs (no. 2012202120013), Tianjin
City High School Science and Technology Fund Planning Project (no.
20130122) and Science Foundation of Medical University Of Tianjin
(no. 2011KY15).
|
1
|
Fernández-Fernández FJ and Sesma P:
Gastric cancer. Lancet. 374(1594): author reply. 1594–1595. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hartgrink HH, Jansen EP, van Grieken NC
and van de Velde CJ: Gastric cancer. Lancet. 374:477–490. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamaoka Y, Kato M and Asaka M: Geographic
differences in gastric cancer incidence can be explained by
differences between Helicobacter pylori strains. Intern Med.
47:1077–1083. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsugane S and Sasazuki S: Diet and the
risk of gastric cancer: Review of epidemiological evidence. Gastric
Cancer. 10:75–83. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bae JM, Lee EJ and Guyatt G: Citrus fruit
intake and stomach cancer risk: A quantitative systematic review.
Gastric Cancer. 11:23–32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 131:11–29. 2007.
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Koturbash I, Zemp FJ, Pogribny I and
Kovalchuk O: Small molecules with big effects: The role of the
microRNAome in cancer and carcinogenesis. Mutat Res. 722:94–105.
2011. View Article : Google Scholar
|
|
10
|
Peláez N and Carthew RW: Biological
robustness and the role of microRNAs: A network perspective. Curr
Top Dev Biol. 99:237–255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang H, Yang H, Zhang C, Jing Y, Wang C,
Liu C, Zhang R, Wang J, Zhang J, Zen K, et al: Investigation of
microRNA expression in human serum during the aging process. J
Gerontol A Biol Sci Med Sci. 70:102–109. 2015. View Article : Google Scholar
|
|
12
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M,
et al: Human microRNA genes are frequently located at fragile sites
and genomic regions involved in cancers. Proc Natl Acad Sci USA.
101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu R, Chen X, Du Y, Yao W, Shen L, Wang
C, Hu Z, Zhuang R, Ning G, Zhang C, et al: Serum microRNA
expression profile as a biomarker in the diagnosis and prognosis of
pancreatic cancer. Clin Chem. 58:610–618. 2012. View Article : Google Scholar
|
|
14
|
Liu R, Zhang C, Hu Z, Li G, Wang C, Yang
C, Huang D, Chen X, Zhang H, Zhuang R, et al: A five-microRNA
signature identified from genome-wide serum microRNA expression
profiling serves as a fingerprint for gastric cancer diagnosis. Eur
J Cancer. 47:784–791. 2011. View Article : Google Scholar
|
|
15
|
Wang C, Hu J, Lu M, Gu H, Zhou X, Chen X,
Zen K, Zhang CY, Zhang T, Ge J, et al: A panel of five serum miRNAs
as a potential diagnostic tool for early-stage renal cell
carcinoma. Sci Rep. 5:76102015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo Y, Wang C, Chen X, Zhong T, Cai X,
Chen S, Shi Y, Hu J, Guan X, Xia Z, et al: Increased serum and
urinary microRNAs in children with idiopathic nephrotic syndrome.
Clin Chem. 59:658–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Esquela-Kerscher A and Slack FJ: oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Loscalzo J, Kohane I and Barabasi AL:
Human disease classification in the postgenomic era: A complex
systems approach to human pathobiology. Mol Syst Biol. 3:1242007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Taneja SS, Goddy G, Kibel AS, Penson DF
and Wei JT: Prostate cancer detection using a novel computerized
three-dimensional prostate biopsy template (Targetscan (Tm)):
Results of a multi-center prospective data registry. J Urol.
181:712. 2009. View Article : Google Scholar
|
|
20
|
Mi H and Thomas P: PANTHER pathway: An
ontology-based pathway database coupled with data analysis tools.
Methods Mol Biol. 563:123–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mi H, Guo N, Kejariwal A and Thomas PD:
PANTHER version 6: Protein sequence and function evolution data
with expanded representation of biological pathways. Nucleic Acids
Res. 35:D247–D252. 2007. View Article : Google Scholar
|
|
22
|
Huang DW, Sherman BT, Tan Q, Collins JR,
Alvord WG, Roayaei J, Stephens R, Baseler MW, Lane HC and Lempicki
RA: The DAVID Gene Functional Classification Tool: A novel
biological module-centric algorithm to functionally analyze large
gene lists. Genome Biol. 8:R1832007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kanehisa M, Araki M, Goto S, Hattori M,
Hirakawa M, Itoh M, Katayama T, Kawashima S, okuda S, Tokimatsu T,
et al: KEGG for linking genomes to life and the environment.
Nucleic Acids Res. 36:D480–D484. 2008. View Article : Google Scholar :
|
|
24
|
Han C, Yu Z, Duan Z and Kan Q: Role of
microRNA-1 in human cancer and its therapeutic potentials. Biomed
Res Int. 2014:4283712014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nohata N, Hanazawa T, enokida H and Seki
N: microRNA-1/133a and microRNA-206/133b clusters: Dysregulation
and functional roles in human cancers. Oncotarget. 3:9–21. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu YN, Yin JJ, Abou-Kheir W, Hynes PG,
Casey OM, Fang L, Yi M, Stephens RM, Seng V, Sheppard-Tillman H, et
al: miR-1 and miR-200 inhibit EMT via Slug-dependent and
tumorigenesis via Slug-independent mechanisms. Oncogene.
32:296–306. 2013. View Article : Google Scholar
|
|
27
|
Reid JF, Sokolova V, Zoni E, Lampis A,
Pizzamiglio S, Bertan C, Zanutto S, Perrone F, Camerini T, Gallino
G, et al: miRNA profiling in colorectal cancer highlights miR-1
involvement in MET-dependent proliferation. Mol Cancer Res.
10:504–515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan D, Dong XE, Chen X, Wang L, Lu C, Wang
J, Qu J and Tu L: MicroRNA-1/206 targets c-Met and inhibits
rhabdomyosarcoma development. J Biol Chem. 284:29596–29604. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nasser MW, Datta J, Nuovo G, Kutay H,
Motiwala T, Majumder S, Wang B, Suster S, Jacob ST and Ghoshal K:
Down-regulation of micro-RNA-1 (miR-1) in lung cancer. Suppression
of tumorigenic property of lung cancer cells and their
sensitization to doxorubicin-induced apoptosis by miR-1. J Biol
Chem. 283:33394–33405. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang F, Song G, Liu M, Li X and Tang H:
miRNA-1 targets fibronectin1 and suppresses the migration and
invasion of the Hep2 laryngeal squamous carcinoma cell line. FEBS
Lett. 585:3263–3269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Leone V, D'Angelo D, Rubio I, de Freitas
PM, Federico A, Colamaio M, Pallante P, Medeiros-Neto G and Fusco
A: miR-1 is a tumor suppressor in thyroid carcinogenesis targeting
CCND2, CXCR4, and SDF-1alpha. J Clin endocrinol Metab.
96:E1388–E1398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li D, Liu Y, Li H, Peng JJ, Tan Y, Zou Q,
Song XF, Du M, Yang ZH, Tan Y, et al: MicroRNA-1 promotes apoptosis
of hepatocarcinoma cells by targeting apoptosis inhibitor-5
(API-5). FEBS Lett. 589:68–76. 2015. View Article : Google Scholar
|
|
33
|
Li D, Yang P, Li H, Cheng P, Zhang L, Wei
D, Su X, Peng J, Gao H, Tan Y, et al: MicroRNA-1 inhibits
proliferation of hepatocarcinoma cells by targeting endothelin-1.
Life Sci. 91:440–447. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tominaga E, Yuasa K, Shimazaki S and
Hijikata T: MicroRNA-1 targets Slug and endows lung cancer A549
cells with epithelial and anti-tumorigenic properties. Exp Cell
Res. 319:77–88. 2013. View Article : Google Scholar
|
|
35
|
Jung YJ, Kim JW, Park SJ, Min By, Jang ES,
Kim NY, Jeong SH, Shin CM, Lee SH, Park YS, et al: c-Myc-mediated
overexpression of miR-17-92 suppresses replication of hepatitis B
virus in human hepatoma cells. J Med Virol. 85:969–978. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chang CC, Yang YJ, Li YJ, Chen ST, Lin BR,
Wu TS, Lin SK, Kuo My and Tan CT: MicroRNA-17/20a functions to
inhibit cell migration and can be used a prognostic marker in oral
squamous cell carcinoma. Oral oncol. 49:923–931. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fan MQ, Huang CB, Gu Y, Xiao Y, Sheng JX
and Zhong L: Decrease expression of microRNA-20a promotes cancer
cell proliferation and predicts poor survival of hepatocellular
carcinoma. J Exp Clin Cancer Res. 32:212013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
No authors listed. miR-20a facilitates
metastasis of osteosarcoma cells to lung tissue. Bonekey Rep.
1:762012.PubMed/NCBI
|
|
39
|
Yoshino H, Seki N, Itesako T, Chiyomaru T,
Nakagawa M and Enokida H: Aberrant expression of microRNAs in
bladder cancer. Nat Rev Urol. 10:396–404. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li X, Zhang Z, Yu M, Li L, Du G, Xiao W
and Yang H: Involvement of miR-20a in promoting gastric cancer
progression by targeting early growth response 2 (eGR2). Int J Mol
Sci. 14:16226–16239. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li X, Pan JH, Song B, Xiong EQ, Chen ZW,
Zhou ZS and Su YP: Suppression of CX43 expression by miR-20a in the
progression of human prostate cancer. Cancer Biol Ther. 13:890–898.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao S, Yao DS, Chen JY and Ding N:
Aberrant expression of miR-20a and miR-203 in cervical cancer.
Asian Pac J Cancer Prev. 14:2289–2293. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhao S, Yao D, Chen J, Ding N and Ren F:
miR-20a promotes cervical cancer proliferation and metastasis in
vitro and in vivo. PLoS One. 10:e01209052015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhou J, Liu R, Luo C, Zhou X, Xia K, Chen
X, Zhou M, Zou Q, Cao P and Cao K: miR-20a inhibits cutaneous
squamous cell carcinoma metastasis and proliferation by directly
targeting LIMK1. Cancer Biol Ther. 15:1340–1349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xie J, Liu M, Li Y, Nie Y, Mi Q and Zhao
S: ovarian tumor-associated microRNA-20a decreases natural killer
cell cytotoxicity by downregulating MICA/B expression. Cell Mol
Immunol. 11:495–502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yan H, Wu J, Liu W, Zuo Y, Chen S, Zhang
S, Zeng M and Huang W: MicroRNA-20a overexpression inhibited
proliferation and metastasis of pancreatic carcinoma cells. Hum
Gene Ther. 21:1723–1734. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Guttilla IK and White BA: Coordinate
regulation of FOXO1 by miR-27a, miR-96, and miR-182 in breast
cancer cells. J Biol Chem. 284:23204–23216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li S, Li J, Fei BY, Shao D, Pan Y, Mo ZH,
Sun BZ, Zhang D, Zheng X, Zhang M, et al: miR-27a promotes
hepatocellular carcinoma cell proliferation through suppression of
its target gene peroxisome proliferator-activated receptor γ. Chin
Med J (Engl). 128:941–947. 2015. View Article : Google Scholar
|
|
49
|
Acunzo M, Romano G, Palmieri D, Laganá A,
Garofalo M, Balatti V, Drusco A, Chiariello M, Nana-Sinkam P and
Croce CM: Cross-talk between MET and EGFR in non-small cell lung
cancer involves miR-27a and Sprouty2. Proc Natl Acad Sci USA.
110:8573–8578. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Salah Z, Arafeh R, Maximov V, Galasso M,
Khawaled S, Abou-Sharieha S, Volinia S, Jones KB, Croce CM and
Aqeilan RI: miR-27a and miR-27a* contribute to metastatic
properties of osteosarcoma cells. Oncotarget. 6:4920–4935. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Peng H, Wang X, Zhang P, Sun T, Ren X and
Xia Z: miR-27a promotes cell proliferation and metastasis in renal
cell carcinoma. Int J Clin exp Pathol. 8:2259–2266. 2015.PubMed/NCBI
|
|
52
|
Chen Z, Ma T, Huang C, Zhang L, Lv X, Xu
T, Hu T and Li J: miR-27a modulates the MDR1/P-glycoprotein
expression by inhibiting FZD7/β-catenin pathway in hepatocellular
carcinoma cells. Cell Signal. 25:2693–2701. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Deng Y, Bai H and Hu H: rs11671784 G/A
variation in miR-27a decreases chemo-sensitivity of bladder cancer
by decreasing miR-27a and increasing the target RUNX-1 expression.
Biochem Biophys Res Commun. 458:321–327. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Li Z, Hu S, Wang J, Cai J, Xiao L, Yu L
and Wang Z: miR-27a modulates MDR1/P-glycoprotein expression by
targeting HIPK2 in human ovarian cancer cells. Gynecol oncol.
119:125–130. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tanaka K, Miyata H, Sugimura K, Fukuda S,
Kanemura T, Yamashita K, Miyazaki Y, Takahashi T, Kurokawa Y,
Yamasaki M, et al: miR-27 is associated with chemoresistance in
esophageal cancer through transformation of normal fibroblasts to
cancer-associated fibroblasts. Carcinogenesis. 36:894–903. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang H, Li M, Han Y, Hong L, Gong T, Sun
L and Zheng X: Down-regulation of miR-27a might reverse multidrug
resistance of esophageal squamous cell carcinoma. Dig Dis Sci.
55:2545–2551. 2010. View Article : Google Scholar
|
|
57
|
Zhao X, Yang L and Hu J: Down-regulation
of miR-27a might inhibit proliferation and drug resistance of
gastric cancer cells. J Exp Clin Cancer Res. 30:552011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhu H, Wu H, Liu X, evans BR, Medina DJ,
Liu CG and Yang JM: Role of microRNA miR-27a and miR-451 in the
regulation of MDR1/P-glycoprotein expression in human cancer cells.
Biochem Pharmacol. 76:582–588. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ren YQ, Fu F and Han J: miR-27a modulates
radiosensitivity of triple-negative breast cancer (TNBC) cells by
targeting CDC27. Med Sci Monit. 21:1297–1303. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kariya A, Furusawa Y, Yunoki T, Kondo T
and Tabuchi Y: A microRNA-27a mimic sensitizes human oral squamous
cell carcinoma HSC-4 cells to hyperthermia through downregulation
of Hsp110 and Hsp90. Int J Mol Med. 34:334–340. 2014.PubMed/NCBI
|
|
61
|
Coutinho-Camillo CM, Lourenço SV, de
Araújo Lima L, Kowalski LP and Soares FA: expression of
apoptosis-regulating miRNAs and target mRNAs in oral squamous cell
carcinoma. Cancer Genet. 208:382–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hong JH, Roh KS, Suh SS, Lee S, Sung SW,
Park JK, Byun JH and Kang JH: The expression of microRNA-34a is
inversely correlated with c- MET and CDK6 and has a prognostic
significance in lung adenocarcinoma patients. Tumour Biol. Jun
25–2015, (Epub ahead of print) http://dx.doi.org/10.1007/s13277-015-3428-9.
View Article : Google Scholar
|
|
63
|
Isosaka M, Niinuma T, Nojima M, Kai M,
Yamamoto E, Maruyama R, Nobuoka T, Nishida T, Kanda T, Taguchi T,
et al: A screen for epigenetically silenced microRNA genes in
gastrointestinal stromal tumors. PLoS One. 10:e01337542015.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lin L, Jiang H, Huang M, Hou X, Sun X,
Jiang X, Dong X, Sun X, Zhou B and Qiao H: Depletion of histone
deacetylase 1 inhibits metastatic abilities of gastric cancer cells
by regulating the miR-34a/CD44 pathway. Oncol Rep. 34:663–672.
2015.PubMed/NCBI
|
|
65
|
Liu C, Kelnar K, Liu B, Chen X,
Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, et
al: The microRNA miR-34a inhibits prostate cancer stem cells and
metastasis by directly repressing CD44. Nat Med. 17:211–215. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Lu G, Sun Y, An S, Xin S, Ren X, Zhang D,
Wu P, Liao W, Ding Y and Liang L: MicroRNA-34a targets FMNL2 and
E2F5 and suppresses the progression of colorectal cancer. Exp Mol
Pathol. 99:173–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wei B, Huang QE, Huang SR, Mai W and Zhong
XG: MicroRNA 34a attenuates the proliferation, invasion and
metastasis of gastric cancer cells via downregulation of MET. Mol
Med Rep. 12:5255–5261. 2015.PubMed/NCBI
|
|
68
|
Yu L, Xiong J, Guo L, Miao L, Liu S and
Guo F: The effects of lanthanum chloride on proliferation and
apoptosis of cervical cancer cells: Involvement of let-7a and
miR-34a microRNAs. Biometals. 28:879–890. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Hermeking H: The miR-34 family in cancer
and apoptosis. Cell Death Differ. 17:193–199. 2010. View Article : Google Scholar
|
|
70
|
Kasinski AL and Slack FJ: miRNA-34
prevents cancer initiation and progression in a therapeutically
resistant K-ras and p53-induced mouse model of lung adenocarcinoma.
Cancer Res. 72:5576–5587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
He L, He X, Lowe SW and Hannon GJ:
microRNAs join the p53 network - another piece in the
tumour-suppression puzzle. Nat Rev Cancer. 7:819–822. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Qiao P, Li G, Bi W, Yang L, Yao L and Wu
D: microRNA-34a inhibits epithelial mesenchymal transition in human
cholangiocarcinoma by targeting Smad4 through transforming growth
factor-beta/Smad pathway. BMC Cancer. 15:4692015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Alemar B, Izetti P, Gregório C, Macedo GS,
Castro MA, Osvaldt AB, Matte U and Ashton-Prolla P: miRNA-21 and
miRNA-34a are potential minimally invasive biomarkers for the
diagnosis of pancreatic ductal adenocarcinoma. Pancreas. Aug
10–2015.Epub ahead of print. PubMed/NCBI
|
|
74
|
Shi Y and Huang A: effects of sorafenib on
lung metastasis in rats with hepatocellular carcinoma: The role of
microRNAs. Tumour Biol. May 31–2015.(Epub ahead of print)
http://dx.doi.org/10.1007/s13277-015-3565-1.
View Article : Google Scholar
|
|
75
|
Lin J, Huang S, Wu S, Ding J, Zhao Y,
Liang L, Tian Q, Zha R, Zhan R and He X: MicroRNA-423 promotes cell
growth and regulates G(1)/S transition by targeting p21Cip1/Waf1 in
hepatocellular carcinoma. Carcinogenesis. 32:1641–1647. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Stiuso P, Potenza N, Lombardi A,
Ferrandino I, Monaco A, Zappavigna S, Vanacore D, Mosca N,
Castiello F, Porto S, et al: MicroRNA-423-5p promotes autophagy in
cancer cells and is increased in serum from hepatocarcinoma
patients treated with sorafenib. Mol Ther Nucleic Acids.
4:e2332015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Liu J, Wang X, Yang X, Liu Y, Shi Y, Ren J
and Guleng B: miRNA423-5p regulates cell proliferation and invasion
by targeting trefoil factor 1 in gastric cancer cells. Cancer Lett.
347:98–104. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lu X and Lu J: The significance of
detection of serum miR-423-5p and miR-484 for diagnosis of
colorectal cancer. Clin Lab. 61:187–190. 2015.PubMed/NCBI
|
|
79
|
Ali S, Saleh H, Sethi S, Sarkar FH and
Philip PA: MicroRNA profiling of diagnostic needle aspirates from
patients with pancreatic cancer. Br J Cancer. 107:1354–1360. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhao H, Gao A, Zhang Z, Tian R, Luo A, Li
M, Zhao D, Fu L, Fu L, Dong JT, et al: Genetic analysis and
preliminary function study of miR-423 in breast cancer. Tumour
Biol. 36:4763–4771. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Yan W, Liu Y, Yang P, Wang Z, You Y and
Jiang T: MicroRNA profiling of Chinese primary glioblastoma reveals
a temozolomide-chemoresistant subtype. Oncotarget. 6:11676–11682.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wang D, Qiu C, Zhang H, Wang J, Cui Q and
Yin Y: Human microRNA oncogenes and tumor suppressors show
significantly different biological patterns: From functions to
targets. PLoS One. 5:e130672010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Lv H, Pei J, Liu H, Wang H and Liu J: A
polymorphism site in the pre-miR-34a coding region reduces miR-34a
expression and promotes osteosarcoma cell proliferation and
migration. Mol Med Rep. 10:2912–2916. 2014.PubMed/NCBI
|
|
84
|
Kisseljov FL: MicroRNAs and cancer. Mol
Biol. 48:197–206. 2014. View Article : Google Scholar
|
|
85
|
Tutar L, Tutar E and Tutar Y: MicroRNAs
and cancer; an overview. Curr Pharm Biotechnol. 15:430–437. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Li Y, Che Q, Bian Y, Zhou Q, Jiang F, Tong
H, Ke J, Wang K and Wan XP: Autocrine motility factor promotes
epithelial-mesenchymal transition in endometrial cancer via MAPK
signaling pathway. Int J oncol. 47:1017–1024. 2015.PubMed/NCBI
|
|
87
|
Zhao L, Wang Y, Yan Q, Lv W, Zhang Y and
He S: Exogenous hydrogen sulfide exhibits anti-cancer effects
though p38 MAPK signaling pathway in C6 glioma cells. Biol Chem.
396:1247–1253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Yan H, Xin S, Wang H, Ma J, Zhang H and
Wei H: Baicalein inhibits MMP-2 expression in human ovarian cancer
cells by suppressing the p38 MAPK-dependent NF-κB signaling
pathway. Anticancer Drugs. 26:649–656. 2015.PubMed/NCBI
|
|
89
|
Nakareangrit W, Thiantanawat A,
Visitnonthachai D, Watcharasit P and Satayavivad J: Sodium arsenite
inhibited genomic estrogen signaling but induced pERα (Ser118) via
MAPK pathway in breast cancer cells. Environ Toxicol. Mar
2–2015.Epub ahead of print. View Article : Google Scholar
|
|
90
|
Chang L, Graham PH, Ni J, Hao J, Bucci J,
Cozzi PJ and Li Y: Targeting PI3K/Akt/mTOR signaling pathway in the
treatment of prostate cancer radioresistance. Crit Rev Oncol
Hematol. Jul 18–2015.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Xia P and Xu Xy: PI3K/Akt/mTOR signaling
pathway in cancer stem cells: From basic research to clinical
application. Am J Cancer Res. 5:1602–1609. 2015.PubMed/NCBI
|
|
92
|
Wang L, Wu J, Lu J, Ma R, Sun D and Tang
J: Regulation of the cell cycle and PI3K/Akt/mTOR signaling pathway
by tanshinone I in human breast cancer cell lines. Mol Med Rep.
11:931–939. 2015.
|
|
93
|
Zuidervaart W, van Nieuwpoort F, Stark M,
Dijkman R, Packer L, Borgstein AM, Pavey S, van der Velden P, Out
C, Jager MJ, et al: Activation of the MAPK pathway is a common
event in uveal melanomas although it rarely occurs through mutation
of BRAF or RAS. Br J Cancer. 92:2032–2038. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
O'Connell RM, Taganov KD, Boldin MP, Cheng
G and Baltimore D: MicroRNA-155 is induced during the macrophage
inflammatory response. Proc Natl Acad Sci USA. 104:1604–1609. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Mateescu B, Batista L, Cardon M, Gruosso
T, de Feraudy Y, Mariani O, Nicolas A, Meyniel JP, Cottu P,
Sastre-Garau X, et al: miR-141 and miR-200a act on ovarian
tumorigenesis by controlling oxidative stress response. Nat Med.
17:1627–1635. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Shao N, Lu Z, Zhang Y, Wang M, Li W, Hu Z,
Wang S and Lin Y: Interleukin-8 upregulates integrin β3 expression
and promotes estrogen receptor-negative breast cancer cell invasion
by activating the PI3K/Akt/NF-κB pathway. Cancer Lett. 364:165–172.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Yue S, Li J, Lee SY, Lee HJ, Shao T, Song
B, Cheng L, Masterson TA, Liu X, Ratliff TL, et al: Cholesteryl
ester accumulation induced by PTEN loss and PI3K/AKT activation
underlies human prostate cancer aggressiveness. Cell Metab.
19:393–406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Graham RM, Middleton A, Benito DA, Uddin
R, Zhang B, Walters W, Bregy A, Vanni S and Komotar RJ: Targeting
cancer stem cells via inhibition of PI3K/AKT pathway alone and in
combination with autophagy blockade. Mol Cancer Ther. 14:B392015.
View Article : Google Scholar
|
|
100
|
Prasad SB, Yadav SS, Das M, Modi A, Kumari
S, Pandey LK, Singh S, Pradhan S and Narayan G: PI3K/AKT
pathway-mediated regulation of p27(Kip1) is associated with cell
cycle arrest and apoptosis in cervical cancer. Cell oncol (Dordr).
38:215–225. 2015. View Article : Google Scholar
|
|
101
|
Mabuchi S, Kuroda H, Takahashi R and
Sasano T: The PI3K/AKT/MTOR pathway as a therapeutic target in
ovarian cancer. Gynecol oncol. 137:173–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Jin D, Yang JP, Hu JH, Wang LN and Zuo JL:
MCP-1 stimulates spinal microglia via PI3K/Akt pathway in bone
cancer pain. Brain Res. 1599:158–167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Fang F and Wang L, Zhang S, Fang Q, Hao F,
Sun Y, Zhao L, Chen S, Liao H and Wang L: CD147 modulates autophagy
through the PI3K/Akt/mTOR pathway in human prostate cancer PC-3
cells. Oncol Lett. 9:1439–1443. 2015.PubMed/NCBI
|
|
104
|
Cárdenas A, Kong M, Alvarez A, Valdivia A,
Quest AF and Leyton L: PAR-3 and Syndecan-4 are involved in
astrocyte adhesion induced by neuronal Thy-1 ocyte adhesion. Glia.
63:E102. 2015.
|
|
105
|
Xie M, He J, He C and Wei S: γ secretase
inhibitor BMS-708163 reverses resistance to eGFR inhibitor via the
PI3K/Akt pathway in lung cancer. J Cell Biochem. 116:1019–1027.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Xue B, Huang W, Yuan X, Xu B, Lou Y, Zhou
Q, Ran F, Ge Z, Li R and Cui J: YSY01A, a novel proteasome
inhibitor, induces cell cycle arrest on G2 phase in MCF-7 cells via
eRα and PI3K/Akt pathways. J Cancer. 6:319–326. 2015. View Article : Google Scholar
|
|
107
|
Niu NK, Wang ZL, Pan ST, Ding HQ, Au GH,
He ZX, Zhou ZW, Xiao G, Yang YX, Zhang X, et al: Pro-apoptotic and
pro-autophagic effects of the Aurora kinase A inhibitor alisertib
(MLN8237) on human osteosarcoma U-2 OS and MG-63 cells through the
activation of mitochondria-mediated pathway and inhibition of p38
MAPK/PI3K/Akt/mTOR signaling pathway. Drug Des Devel Ther.
9:1555–1584. 2015.PubMed/NCBI
|
|
108
|
Gatza ML, Watt JC and Marriott SJ:
Cellular transformation by the HTLV-I Tax protein, a
jack-of-all-trades. Oncogene. 22:5141–5149. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Bai XT and Nicot C: miR-28-3p is a
cellular restriction factor that inhibits human T cell leukemia
virus, type 1 (HTLV-1) replication and virus infection. J Biol
Chem. 290:5381–5390. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Drosten M, Sum EY, Lechuga CG,
Simón-Carrasco L, Jacob HK, García-Medina R, Huang S, Beijersbergen
RL, Bernards R and Barbacid M: Loss of p53 induces cell
proliferation via Ras-independent activation of the Raf/Mek/erk
signaling pathway. Proc Natl Acad Sci USA. 111:15155–15160. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Manousaridis I, Mavridou S, Goerdt S,
Leverkus M and Utikal J: Cutaneous side effects of inhibitors of
the RAS/RAF/MeK/eRK signalling pathway and their management. J Eur
Acad Dermatol Venereol. 27:11–18. 2013. View Article : Google Scholar
|
|
112
|
Noser JA, Sakuma R, Lee PWK and Ikeda Y:
The Ras/Raf-1/MeK/eRK signaling pathway dictates host cell
permissiveness to VSV infection. Mol Ther. 13:S371. 2006.
View Article : Google Scholar
|
|
113
|
Dancey JE: Agents targeting ras signaling
pathway. Curr Pharm Des. 8:2259–2267. 2002. View Article : Google Scholar : PubMed/NCBI
|