Introduction
CLL is the most common form of leukemia in adults,
and is particularly common in Western countries (1). However, the incidence of this disease
is gradually increasing also in China (2). It is characterized by an accumulation
of abnormal B cells, resulting from the dysregulation of
proliferation and apoptosis (3,4).
Despite attempts to develop new treatment strategies, CLL is
currently incurable (5,6).
Recent studies have suggested that most CLL cells
are arrested in the G0/G1 phase of the cell cycle. This is caused,
in part, by the aberration of cell cycle related gene such as
cyclin D family, which blocks the transition of G1 to S (5,7,8).
However, there are several studies supporting that a small fraction
of CLL cells display rapid growth (9–11),
which are accumulated in special structures of the bone marrow and
lymphatic nodes, the proliferation centers (12). CLL cells in those areas are
characterized by a higher expression of Ki-67, survivin and Bcl-2
factors typically associated with proliferation (13).
COP1 was first defined as a central regulator of
photomorphogenic development in plants (14), later in mammals (15). It consists of 3 functional domains,
a RING-finger domain, a coiled-coil domain, and a WD40 domain and
is highly conserves from plants to mammals (16). COP1 possesses E3 ubiquitin ligase
activity, which targets key transcription factors for
proteasome-dependent degradation and plays an important function in
many biological responses in mammals (17). As reported, the substrates of
mammalian COP1 include c-Jun, ETV1, p53, acetyl-CoA carboxylase
(ACC) (18), TORC2 and FoxO1,
suggesting that COP1 is involved in tumorigenicity, lipid
metabolism and gluconeogenesis (19–24).
COP1 is frequently overexpressed in breast, ovarian, hepatoma and
gastric cancer, and promotes tumorigenicity via inhibition of p53
activity (25–27). In contrast, COP1 targets
CCAAT/enhancer-binding protein (C/EBPa) for degradation and induces
acute myeloid leukemia via Trib1 (28), while COP1 constitutively regulates
c-Jun protein stability and functions as a tumor suppressor in mice
(29). Although COP1 is involved in
many biological processes in mammalian cells, its role in
tumorigenicity remains controversial (18). Besisdes, the relative abundance and
physiological function of COP1 is unclear in the progression of
CLL.
In the present study, we addressed the role of COP1
in CLL cell proliferation and tumorigenicity. We collected 23
samples from CLL patients at diagnosis, and assessed the
correlation between the expression of COP1 and clinical stage,
chromosomal abnormalities and prognostic indicators in CLL
patients. We found that the level of COP1 was positively correlated
with clinical staging of CLL patients and ZAP-70 expression. We
also showed that overexpression of COP1 facilitated to cell colony
formation and proliferation, further promoted cell cycle transition
from G0/G1 to S phase by inhibition of the FoxO1 and p21 activity.
Moreover, overexpression of COP1 accelerated tumorigenicity and
promoted xenograft growth. These results suggest that COP1 may play
a positive role in the progression of CLL, and that determination
of its expression may be helpful for monitoring condition changes
in CLL patients.
Materials and methods
Patients and samples
CLL samples were obtained from the Affiliated
Hospital of Xuzhou Medical College according to the diagnostic
criteria for CLL between August 2013 and July 2015, while 3 healthy
volunteers served as a normal control. PBMCs were isolated from
heparinized blood obtained from 23 CLL patients, and normal B cells
were separated from healthy donor via CD19+ labeled
MicroBeads according to the manufacturer's instructions.
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards.
Cell line and reagent
CLL cell line HG3, gifted by Anders Rosén (Linköping
University, Sweden) (30), was
cultured in RPIM-1640 medium supplied with 10% (v/v) fetal bovine
serum (FBS; Gibco, Grand Island, NY, USA) at 37°C in 5%
CO2 incubator (Thermo Fisher Scientific, Waltham, MA,
USA). COP1 antibody was purchased from Bethyl Laboratories, Inc.
(Montgomery, TX, USA). Polyclonal antibodies specific for FoxO1 and
ZAP-70 were purchased from Santa Cruz Biotechnology (Santa Cruz,
CA, USA), and monoclonal antibodies for p21 and Ki-67 were,
respectively, purchased from Cell Signaling Technology (Danvers,
MA, USA) and Abcam (Cambridge, UK), anti-GAPDH was purchased from
EnoGene Biotech Co., Ltd. (Nanjing, China). Goat anti-rabbit IgG
and rabbit anti-mouse IgG were obtained from Sigma-Aldrich (St.
Louis, MO, USA). Cell Counting kit (CCK-8) was from Dojindo
Molecular Technologies (Xiongben, Japan), CD19+ labeled
MicroBeads were from Miltenyi Biotec (Auburn, CA, USA).
Animals
Female BALB/C nude mice of 6 weeks of age were
obtained from Vital River Laboratories Co., Ltd. (Beijing, China).
All experimental procedures were performed in accordance with the
guidelines for laboratory animals established by the Xuzhou Medical
College Animal Care and Use Committee.
Plasmid construction and lentiviral
production
For overexpression of COP1, the COP1 cDNA was a gift
from Han Jiahuai laboratory and cloned into lentiviral plasmid
pWPXLd-GFP. For viral production, the pWPXLd-GFP-COP1 plasmid was
cotransfected into 293FT human embryonic kidney cells together with
packaging and envelope protein plasmids with PolyJet (Signagen
Laboratories, Gaithersburg, MD, USA) as described in the
manufacturer's protocol. Culture supernatants containing lentiviral
particles were harvested 72 h after transfection and used for
infection.
Establishment of the stable cell
lines
For stable overexpression of COP1, the HG3 CLL cells
were infected with pWPXLd-GFP or pWPXLd-GFP-COP1 viruses,
respectively. Forty-eight hours after infection, the cells were
continuously cultured in the medium containing 2.5 µg/ml of
puromycin (Sigma). The surviving cells were cultured into cell
lines stably expressing GFP or GFP-COP1.
Soft-agar clone formation assay
The CLL cells were harvested and pipetted to
single-cell suspension with 1640 medium containing 20% FBS in a
given concentration at 1×104/ml. Melted 1.2% agar by
microwave was mixed with pre-warmed 1640 medium containing 20% FBS
and put into a 6-well plate for it to become solid (total volume 1
ml/well), and then, melted 0.6% agar was mixed quickly with cell
suspension equally and added the bottom solid agar (total volume 1
ml/well). Additional 100 µl medium was added to the top
layer and incubated for 2 weeks, at 37°C. The number of colonies
was calculated under a microscope in 3-wells of two groups.
CCK-8 analysis for cell vitality
Three thousand cells in 100 µl of medium were
seeded into 96-well plates with three replicates. CCK-8 reagent (5
µl) was added into each well at the time-point 0, 6, 12, 18,
24 and 30 h. Following incubation for additional 4 h, the cells
were exposed to measure the absorbance at 450 nm by a microplate
reader (Wellscan MK3; Labsystems Dragon, Helsinki, Finland).
Cell cycle analysis
The normal cultured and serum starved CLL cells were
collected and fixed in 70% ethanol on ice for 10 min, rinsed with
PBS and incubated with 100 mg/ml RNase A (0.25 mg/ml) for 15 min.
After washing with PBS two times, the cells were further incubated
with 50 µg/ml propidium iodide (PI) at room temperature for
10 min, and then exposed to cell cycle analysis.
Immunoblotting
The CLL patient samples and cultured cells were
collected and proteins extracted for western blotting. Equal amount
of protein lysates were subjected to 10–12% SDS-PAGE and then
transferred to 0.45 µm pore size PVDF membrane (Millipore,
Billerica, MA, USA). After blocking with 5% non-fat milk, the
membrane was probed with primary antibodies at 4°C overnight and
secondary antibodies at room temperature for 1 h. Bound antibodies
were detected by the Pierce ECL Plus Western Blotting Substrate
(Thermo Fisher Scientific) and visualized by ImageQuant LAS 4000
(GE, Fairfield, CT, USA).
Tumor formation in BALB/C nude mice
Experiments were performed with 6-weeks-old female
BALB/C nude mice (n=6). Both HG3-GFP and HG3-COP1 cells were
cultured in 10-cm dishes in RPMI-1640 medium with 10% FBS and
gently harvested by washing with PBS. The cells were centrifuged
and re-suspended into a suitable concentration at
5×107/ml. An equal volume of the cell suspension (100
µl) was inoculated subcutaneously in the right flank of the
mice. At day 14 after inoculation, tumor size was assessed by
external measurement of the length (L) and width (W) of the tumors
using a vernier caliper. Tumor volume was calculated by using the
following equation: TV = (L × W2)/2. After 30 days, the
tumor was isolated and fixed in 4% PFA at 4°C overnight and
dehydrated gradient in sucrose, after entrapment with embedding
material, the tumor was cut into frozen slices.
Immunohistochemistry
Briefly, tissue sections (~2 mm) were deparaffinized
in xylene, rehydrated in graded alcohols (100, 95, 85 and 75%) and
washed in distilled water. The immunohistochemistry was performed
according to the S-P immunohistochemistry kit (Maixin-Bio, Fuzhou,
China). For each tissue specimen, three horizons were randomly
selected and photographed by microscopy for statistical
analysis.
Statistical analysis
The results are representative of experiments
repeated at least three times and quantitative data were expressed
as means ± SEM. Student's t-test was used to analyze the difference
between test samples and control, and Fisher's exact test was used
for correlation analysis of small samples. P<0.05 was considered
statistically significant, and P<0.01 as very significant. All
statistical analyses were performed using Graphpad Prism 6.0.
Results
Expression of COP1 positively correlates
with Binet classification in CLL patients
We analyzed the expression of COP1 in normal and CLL
patients by isolating mononuclear cells from peripheral blood and
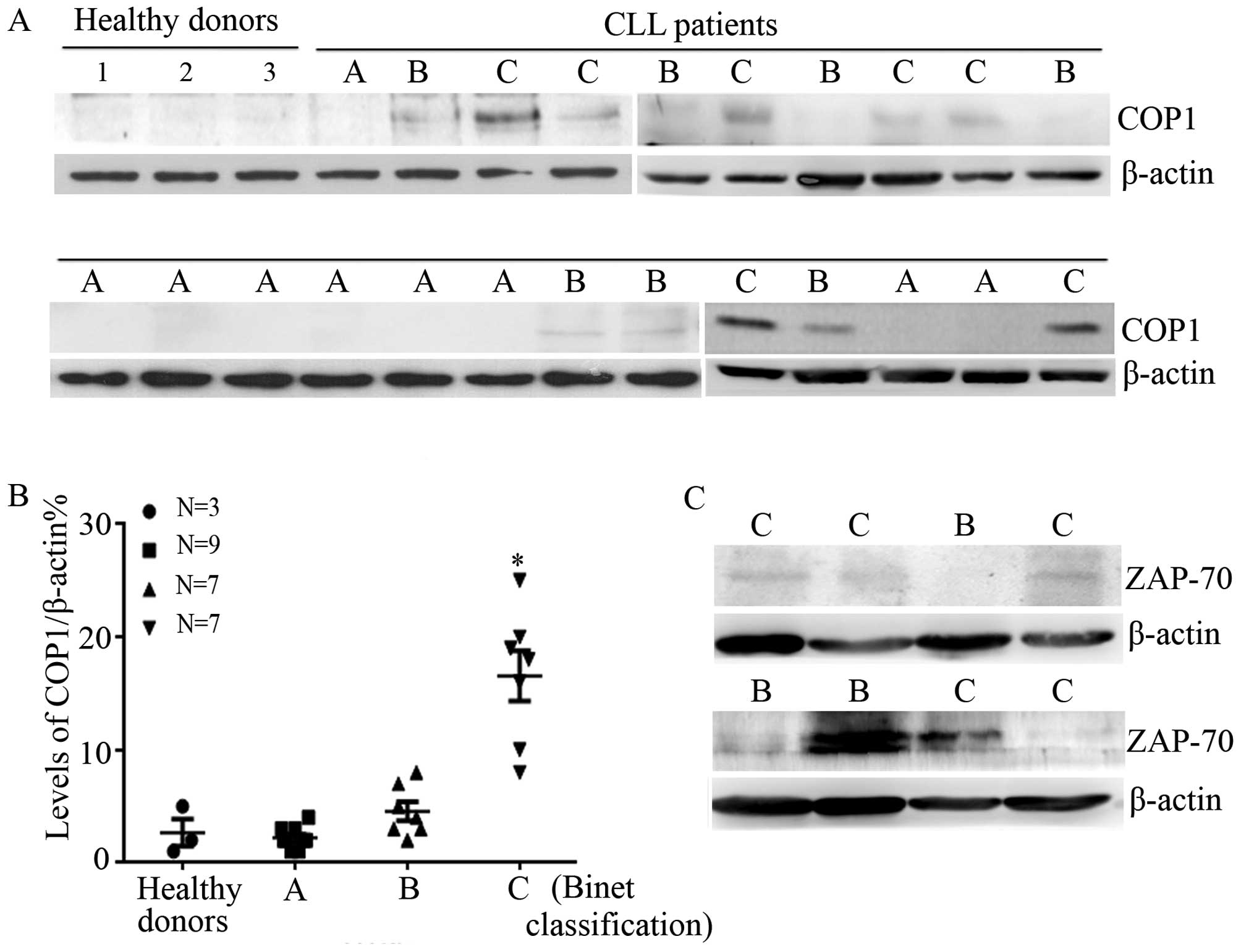
extracting protein for western blotting. Fig. 1A shows that COP1 levels were high in
CLL patients compared with normal donors. Further statistical
analysis indicated that COP1 levels were significantly increased in
CLL cells of Benit C patients (P<0.05) (Fig. 1B).
We next measured the incidence of del(13q14) and
del(17q-), as well as ZAP-70 expression to examine whether the
expression of COP1 was correlated with chromosomal abnormality or
molecular marker in CLL patients. As shown in Table I and Fig. 1C, no apparent correlation was
observed between COP1 expression and del(13q14) (P=0.28), del(17q-)
(P=0.39). Nevertheless, the expression of COP1 was strongly
positively correlated with ZAP-70 expression (P=0.02).
 | Table ICorrelation analysis between the
expression of COP1 and del(13q14), del(17q-), ZAP-70 expression in
CLL patients. |
Table I
Correlation analysis between the
expression of COP1 and del(13q14), del(17q-), ZAP-70 expression in
CLL patients.
| COP1 expression |
|---|
| Low (P<6.8%) | High (≥6.8%) | P-value |
|---|
| ZAP-70
expression | | | 0.02 |
| Positive | 1 | 5 | |
| Negative | 13 | 4 | |
| 13q14 | | | 0.28 |
| Positive | 1 | 2 | |
| Negative | 13 | 7 | |
| 17q- | | | 0.39 |
| Positive | 0 | 1 | |
| Negative | 14 | 8 | |
Overexpression of COP1 contributes to the
proliferation of HG3 cells
We further investigated the role of COP1 in CLL
progression by transfecting HG3 cells with lentiviruses expressing
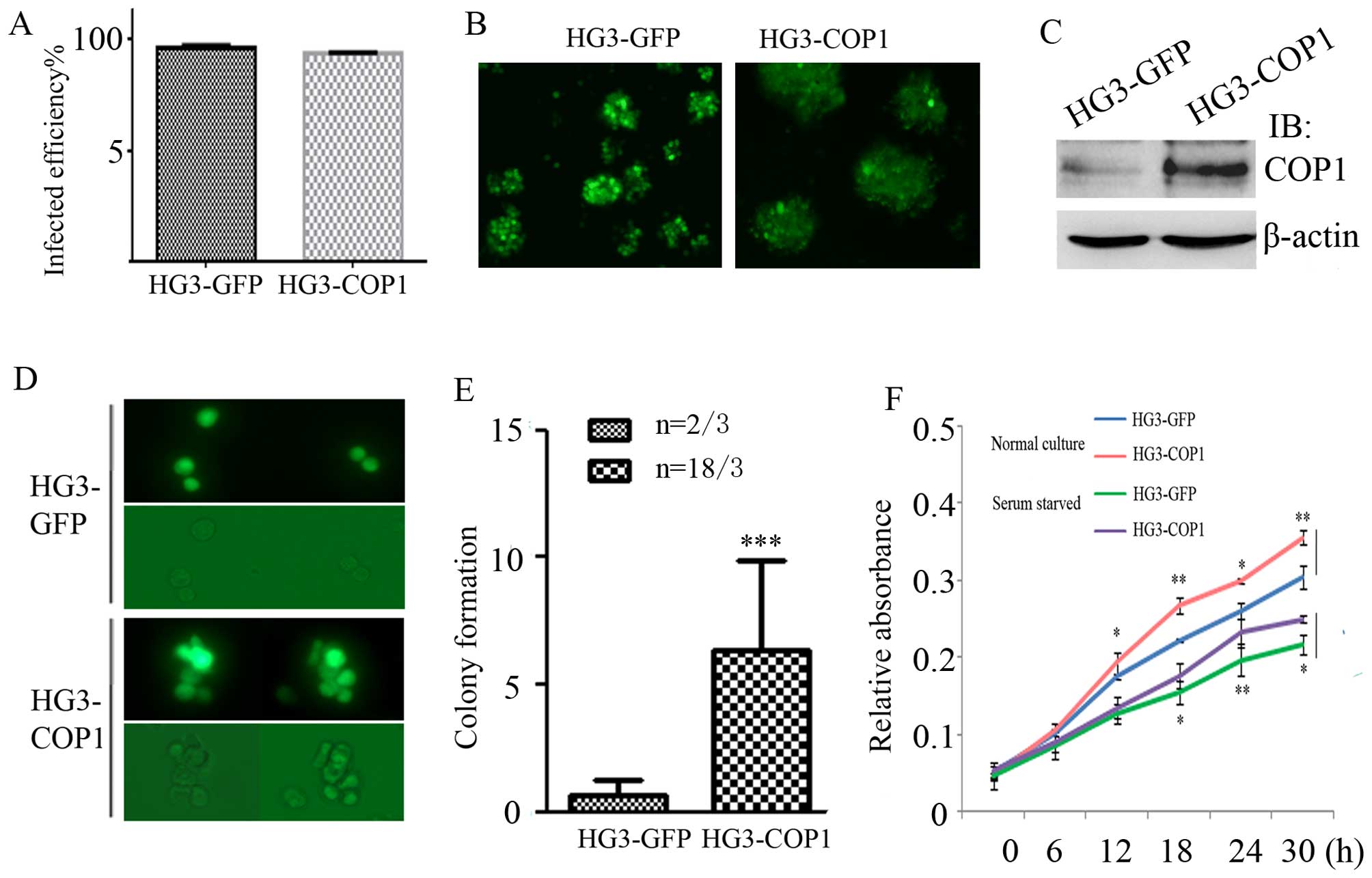
GFP or GFP-COP1. After sustaining the selection with puromycin,
flow cytometry and western blotting showed the infected efficiency
reached >95% (Fig. 2A–C).
Thereafter, the well-established HG3-GFP and HG3-COP1 cells were
first grown in soft agar to observe the colony formation. During
the cultivation, the two groups of CLL cells grew slowly in soft
agar compared to complete culture medium. Notably, HG3-COP1 cells
proliferated slowly and formed cell colonies, while most HG3-GFP
cells did not growth or proliferate except for very few cells. In
the further observation for two weeks, there were 18 colonies in
HG3-COP1 vs. 2 small colonies in HG3-GFP group (Fig. 2D and E).
Then the growth rates of both cells were measured
quantitatively using a CCK-8 kit at 0, 6, 12, 18 and 24 h after
lentivirus infection. HG3-COP1 cells showed significantly higher
absorbance compared with HG3-GFP cells in normal cultivation or
serum starvation treatment, suggesting increased cell viability
(Fig. 2F) (P<0.05).
Overexpression of COP1 promotes the
transition of HG3 cells from G1 to S phase
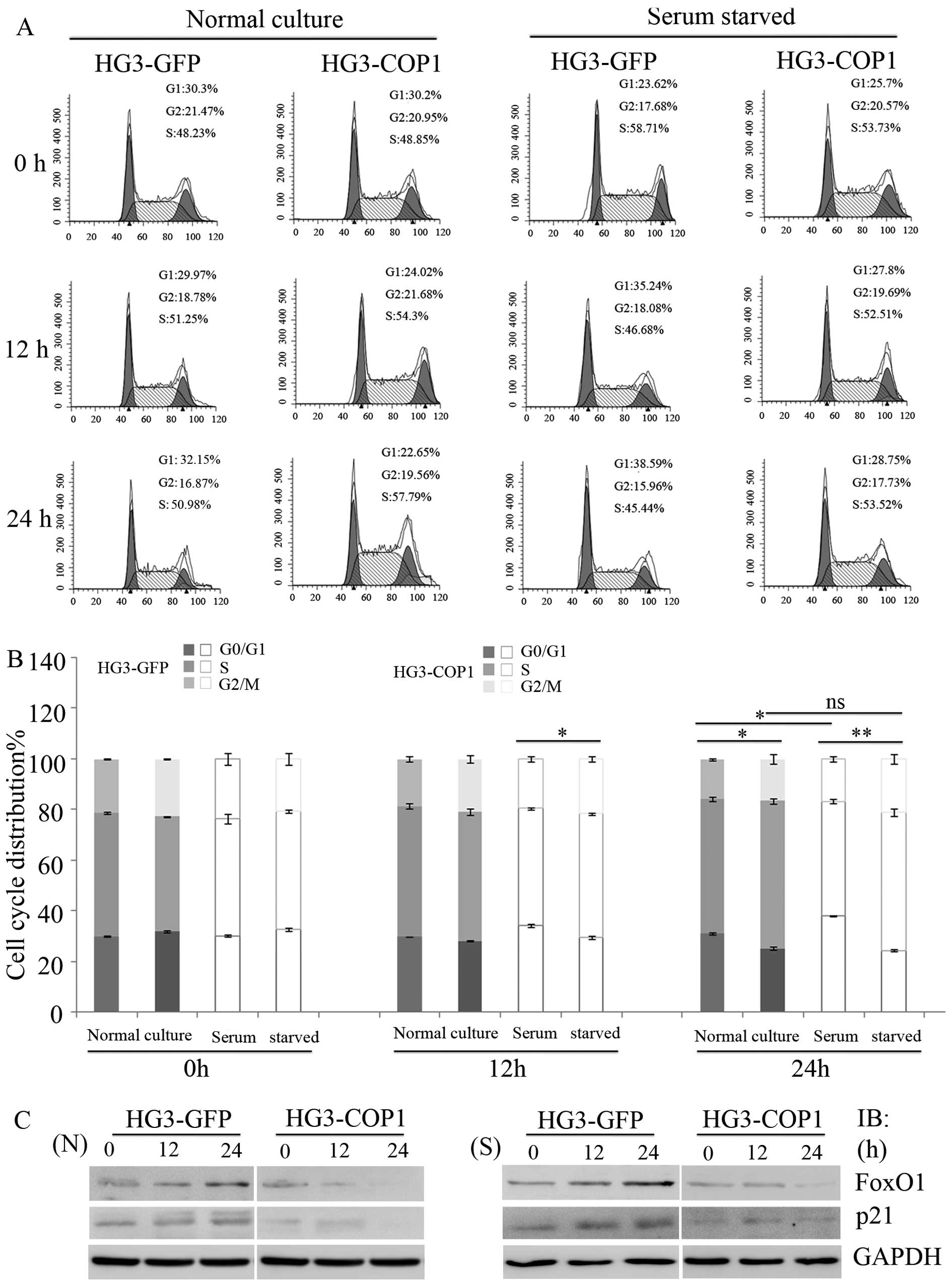
To investigate the role of COP1 in cell cycle
progression, cells were collected and stained with propidium iodide
for cell cycle analysis via flow cytometry. Similar distribution of
the cell cycle were observed in two groups (30–35% in G1, 41–50% in
S and 20–24% in G2/M phases) after normal cultivation for 12 h,
while S-phase HG3-COP1 cells were 55±1% compared with 46±0.9% of
HG3-GFP cells after 24 h, displaying a faster and more
proliferative phenotype (P<0.05). Moreover, it can be seen that
HG3-GFP cells were grown arrested at G0/G1 phase (38±1 and 39±3%,
respectively) upon serum starvation for 12 or 24 h compared to
untreated cells (30±1%), showing an increase of 8 and 9%, while in
G0/G1-phase HG3-COP1 cells (27.6±4 and 28.4±1%, respectively) no
changes were observed with the same treatment compared with cells
at 0 h (25.5±2%) (Fig. 3A and
B).
To study the underlying mechanisms responsible for
the fast growth of HG3-COP1 cells, we detected the expression of
FoxO1, as a polyubiquitinated substate of COP1, and its downstream
genes associated with cell cycle progression. Western blotting
showed that FoxO1 and p21 level in HG3 cells were significantly
inhibited after overexpression of COP1 in normal cultivation for 24
h. In addition, we found that raising COP1 level blocked the
elevation of FoxO1 and p21 level upon serum starvation treatment
(Fig. 3C).
Overexpression of COP1 promotes
tumorigenicity of HG3 cells
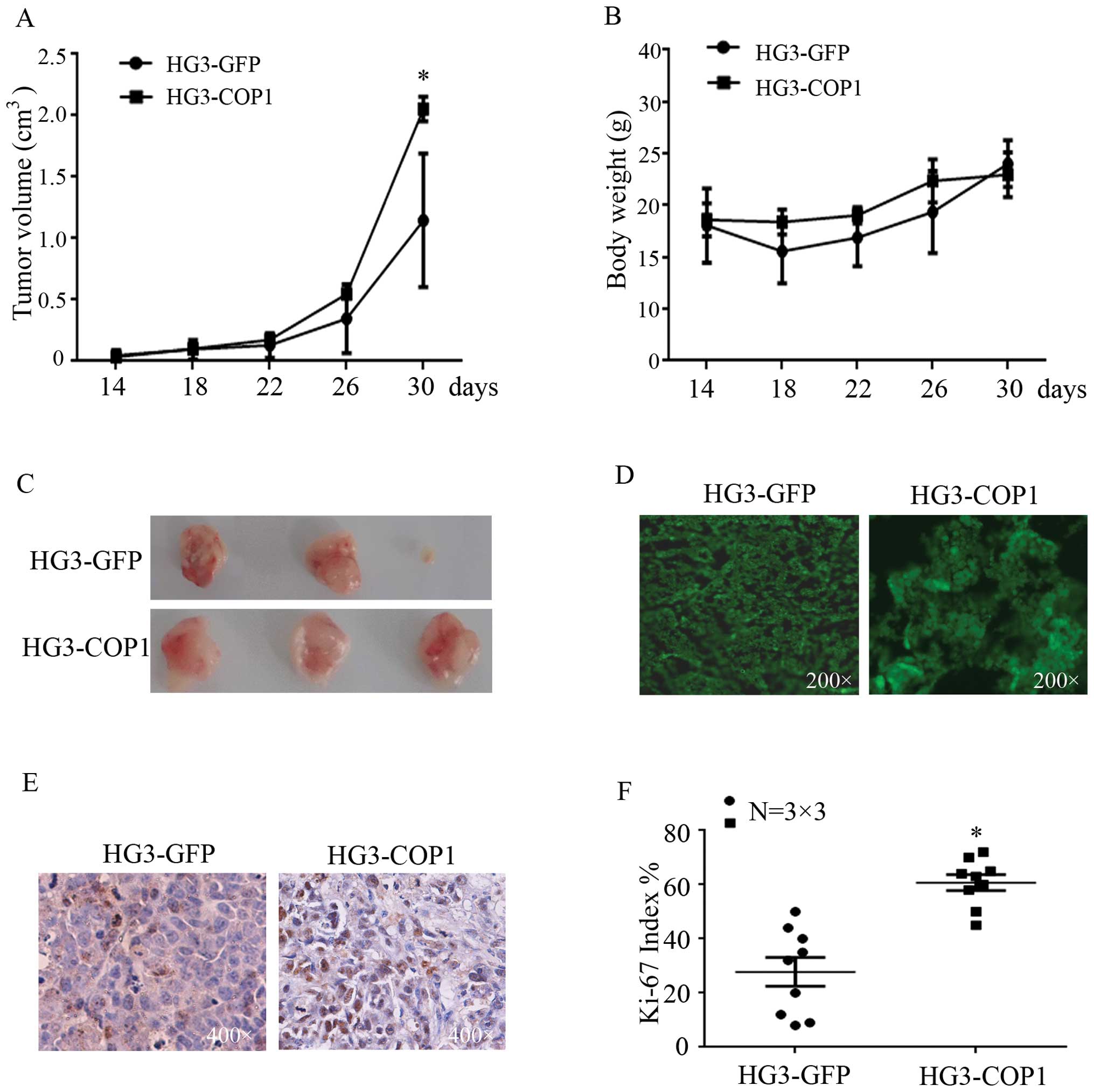
To study the effect of COP1 in CLL cells in
vivo, female BALB/C nude mice were subcutaneously inoculated
with equal numbers of HG3-GFP or HG3-COP1 cells in the right flank.
At day 14 after inoculation, all three mice in the HG3-COP1 group
had developed tumors, whereas, only two mice inoculated with
HG3-GFP cells developed tumors, and the third mouse developed a
tumor at day 26 after inoculation (Table II).
| Table IITumorigenicity of HG3-GFP and
HG3-COP1 cells in BALB/C nude mice. |
Table II
Tumorigenicity of HG3-GFP and
HG3-COP1 cells in BALB/C nude mice.
| Groups | N | Tumorigenicity |
|---|
| 14 | 18 | 22 | 26 | 30 (Days) |
|---|
| HG3-GFP | 3 | 1/3 | 2/3 | 2/3 | 3/3 | 3/3 |
| HG3-COP1 | 3 | 3/3 | 3/3 | 3/3 | 3/3 | 3/3 |
Moreover, the tumor size and body weight of mice
were measured once per four days. The mice inoculated with HG3-COP1
cells showed remarkably increased tumor growth, the final tumor
volume was 2.2 cm3, significantly higher than control
ones with 1.4 cm3 of tumor volume (Fig. 4A). Moreover, the body weight was not
changed in these mice (Fig. 4B).
Thirty days after measurement, both groups had developed tumors
(Fig. 4C and D), whereas, the tumor
tissue from HG3-COP1 group showed stronger Ki-67 staining compared
with the control (Fig. 4E and
F).
Discussion
CLL is the most common human leukemia, representing
30% of all cases (31), which are
characterized as accumulation of B cells due to an imbalance
between activation of cell proliferation and inhibition of
apoptosis (3,4). Most CLL B cells are characterized by
high expression of the p27 protein, which blocks the cell cycle
progression (32). Interestingly, a
small but significant fraction of all leukemic cells proliferates
with higher Ki-67 expression (33),
this discovery makes the pathogenetic mechanisms of CLL more
complicated.
COP1 has been reported to be expressed in most
tumors and normal matching tissues except for partial deletions
(<8% in all cancer types analyzed) in various tumors (29). COP1, as an oncogene, plays an
important role in breast, ovarian, gastric and hepatocellular
carcinoma, its high expression is significantly correlated with
poor survival in gastric cancer patients (13,17,18).
In the present study, we found that COP1 expression was upregulated
in human Benit C-phase CLL patients compared with normal donors.
Moreover, this expression was positively correlated with CLL
clinical stage and ZAP-70 expression, suggesting that it may be an
important indicator of CLL progression.
A recent study showed that COP1 silencing greatly
suppressed the proliferation of human hepatocellular carcinoma
cells, as well as tumorigenicity (17). While overexpression of COP1
accelerates development of acute myeloid leukemia by affecting the
upstream tumor suppressor C/EBPα, suggesting its ligase activity is
crucial for leukemogenesis (33).
Conversely, Migliorini et al (29) reported that a significant decrease
or deletion of COP1 expression are found in prostate carcinoma or
ALL (acute lymphocytic leukemia), respectively. COP1 knockdown
promotes prostate carcinoma cell proliferation by increasing the
basal c-Jun protein levels. In particular, COP1 deficiency
stimulates mouse embryonic fibroblast proliferation in a
c-Jun-dependent manner, and Cop1 hypomorphic mice spontaneously
developed lymphomagenesis when exposed to radiation induction
(21). Notably, we found that
overexpression of COP1 significantly facilitated colony formation
and proliferation of HG3 cells. In particular, it promoted the
cellular transition from G1 to S phase by downregulation of FoxO1.
As known, FoxO1, as a substrate of COP1, directly targets p21 to
regulate the cell cycle process (34). The results showed upregulation of
COP1 also inhibited p21 level in normal cultivation, and blocked
the elevation of FoxO1 and p21 level upon serum starvation.
Importantly, the overexpression of COP1 accelerated tumorigenicity
of HG3 cells and promoted xenograft growth.
In conclusion, the present study revealed that COP1
is an important indicator of CLL processes, while its expression
was shown to be associated with CLL clinical stage and ZAP-70
expression. Moreover, overexpression of COP1 promoted the cell
proliferation and tumorigenicity. These results suggest that COP1
may play a positive role in CLL progression.
Acknowledgments
We are grateful to Dr Anders Rosén (Linköping
University, Sweden) for generously providing HG3 cells. The present
study was supported by the National Natural Science Foundation of
China (81400127), the National Natural Science Foundation of China
(81201264), the National Natural Science Foundation of China
(81200376) and the Certificate of China Postdoctoral Science
Foundation Grant (2015M571818).
References
|
1
|
Malek SN: The biology and clinical
significance of acquired genomic copy number aberrations and
recurrent gene mutations in chronic lymphocytic leukemia. Oncogene.
32:2805–2817. 2013. View Article : Google Scholar :
|
|
2
|
Zhu DX, Zhu W, Fang C, Fan L, Zou ZJ, Wang
YH, Liu P, Hong M, Miao KR, Liu P, et al: miR-181a/b significantly
enhances drug sensitivity in chronic lymphocytic leukemia cells via
targeting multiple anti-apoptosis genes. Carcinogenesis.
33:1294–1301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bianchi S, Dighiero G and Pritsch O:
Selected topics in chronic lymphocytic leukemia pathogenesis.
Chronic Lymphocytic Leukemia. InTech; Rijeka: pp. 3–18. 2012
|
|
4
|
Wójtowicz M and Wołowiec D: Dysregulation
of apoptosis and proliferation in CLL cells. Chronic Lymphocytic
Leukemia. InTech; Rijeka: pp. 37–62. 2012
|
|
5
|
Wang P, Pavletic ZS and Joshi SS:
Increased apoptosis in B-chronic lymphocytic leukemia cells as a
result of cyclin D3 down regulation. Leuk Lymphoma. 43:1827–1835.
2002. View Article : Google Scholar
|
|
6
|
Razavi R, Gehrke I, Gandhirajan RK,
Poll-Wolbeck SJ, Hallek M and Kreuzer KA: Nitric oxide-donating
acetylsalicylic acid induces apoptosis in chronic lymphocytic
leukemia cells and shows strong antitumor efficacy in vivo. Clin
Cancer Res. 17:286–293. 2011. View Article : Google Scholar
|
|
7
|
Halina A, Artur P, Barbara MK, Joanna S
and Anna D: Alterations in TP53, cyclin D2, c-Myc, p21WAF1/CIP1 and
p27KIP1 expression associated with progression in B-CLL. Folia
Histochem Cytobiol. 48:534–541. 2010.
|
|
8
|
Decker T, Hipp S, Ringshausen I, Bogner C,
Oelsner M, Schneller F and Peschel C: Rapamycin-induced G1 arrest
in cycling B-CLL cells is associated with reduced expression of
cyclin D3, cyclin E, cyclin A, and survivin. Blood. 101:278–285.
2003. View Article : Google Scholar
|
|
9
|
Decker T, Schneller F, Hipp S, Miething C,
Jahn T, Duyster J and Peschel C: Cell cycle progression of chronic
lymphocytic leukemia cells is controlled by cyclin D2, cyclin D3,
cyclin-dependent kinase (cdk) 4 and the cdk inhibitor p27.
Leukemia. 16:327–334. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Damle RN, Calissano C and Chiorazzi N:
Chronic lymphocytic leukaemia: A disease of activated monoclonal B
cells. Best Pract Res Clin Haematol. 23:33–45. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sainz-Perez A, Gary-Gouy H, Gaudin F,
Maarof G, Marfaing-Koka A, de Revel T and Dalloul A: IL-24 induces
apoptosis of chronic lymphocytic leukemia B cells engaged into the
cell cycle through dephosphorylation of STAT3 and stabilization of
p53 expression. J Immunol. 181:6051–6060. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Caligaris-Cappio F and Ghia P: Novel
insights in chronic lymphocytic leukemia: Are we getting closer to
understanding the pathogenesis of the disease? J Clin Oncol.
26:4497–4503. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Soma LA, Craig FE and Swerdlow SH: The
proliferation center microenvironment and prognostic markers in
chronic lymphocytic leukemia/small lymphocytic lymphoma. Hum
Pathol. 37:152–159. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schwechheimer C and Deng XW: The
COP/DET/FUS proteins-regulators of eukaryotic growth and
development. Semin Cell Dev Biol. 11:495–503. 2000. View Article : Google Scholar
|
|
15
|
Yi C, Wang H, Wei N and Deng XW: An
initial biochemical and cell biological characterization of the
mammalian homologue of a central plant developmental switch, COP1.
BMC Cell Biol. 3:302002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yi C and Deng XW: COP1 - from plant
photomorphogenesis to mammalian tumorigenesis. Trends Cell Biol.
15:618–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marine JC: Spotlight on the role of COP1
in tumorigenesis. Nat Rev Cancer. 12:455–464. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wei W and Kaelin WG Jr: Good COP1 or bad
COP1? In vivo veritas. J Clin Invest. 121:1263–1265. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wertz IE, O'Rourke KM, Zhang Z, Dornan D,
Arnott D, Deshaies RJ and Dixit VM: Human De-etiolated-1 regulates
c-Jun by assembling a CUL4A ubiquitin ligase. Science.
303:1371–1374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dornan D, Wertz I, Shimizu H, Arnott D,
Frantz GD, Dowd P, O'Rourke K, Koeppen H and Dixit VM: The
ubiquitin ligase COP1 is a critical negative regulator of p53.
Nature. 429:86–92. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vitari AC, Leong KG, Newton K, Yee C,
O'Rourke K, Liu J, Phu L, Vij R, Ferrando R, Couto SS, et al: COP1
is a tumour suppressor that causes degradation of ETS transcription
factors. Nature. 474:403–406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi L, Heredia JE, Altarejos JY, Screaton
R, Goebel N, Niessen S, Macleod IX, Liew CW, Kulkarni RN, Bain J,
et al: TRB3 links the E3 ubiquitin ligase COP1 to lipid metabolism.
Science. 312:1763–1766. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dentin R, Liu Y, Koo SH, Hedrick S, Vargas
T, Heredia J, Yates J III and Montminy M: Insulin modulates
gluconeogenesis by inhibition of the coactivator TORC2. Nature.
449:366–369. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kato S, Ding J, Pisck E, Jhala US and Du
K: COP1 functions as a FoxO1 ubiquitin E3 ligase to regulate
FoxO1-mediated gene expression. J Biol Chem. 283:35464–35473. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dornan D, Bheddah S, Newton K, Ince W,
Frantz GD, Dowd P, Koeppen H, Dixit VM and French DM: COP1, the
negative regulator of p53, is overexpressed in breast and ovarian
adeno-carcinomas. Cancer Res. 64:7226–7230. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee YH, Andersen JB, Song HT, Judge AD,
Seo D, Ishikawa T, Marquardt JU, Kitade M, Durkin ME, Raggi C, et
al: Definition of ubiquitination modulator COP1 as a novel
therapeutic target in human hepatocellular carcinoma. Cancer Res.
70:8264–8269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li YF, Wang DD, Zhao BW, Wang W, Huang CY,
Chen YM, Zheng Y, Keshari RP, Xia JC and Zhou ZW: High level of
COP1 expression is associated with poor prognosis in primary
gastric cancer. Int J Biol Sci. 8:1168–1177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yoshida A, Kato JY, Nakamae I and
Yoneda-Kato N: COP1 targets C/EBPα for degradation and induces
acute myeloid leukemia via Trib1. Blood. 122:1750–1760. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Migliorini D, Bogaerts S, Defever D, Vyas
R, Denecker G, Radaelli E, Zwolinska A, Depaepe V, Hochepied T,
Skarnes WC, et al: Cop1 constitutively regulates c-Jun protein
stability and functions as a tumor suppressor in mice. J Clin
Invest. 121:1329–1343. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rosén A, Bergh AC, Gogok P, Evaldsson C,
Myhrinder AL, Hellqvist E, Rasul A, Björkholm M, Jansson M,
Mansouri L, et al: Lymphoblastoid cell line with B1 cell
characteristics established from a chronic lymphocytic leukemia
clone by in vitro EBV infection. Oncoimmunology. 1:18–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pekarsky Y, Zanesi N and Croce CM:
Molecular basis of CLL. Semin Cancer Biol. 20:370–376. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wolowiec D, Wojtowicz M, Ciszak L,
Kosmaczewska A, Frydecka I, Potoczek S, Urbaniak-Kujda D,
Kapelko-Slowik K and Kuliczkowski K: High intracellular content of
cyclin-dependent kinase inhibitor p27Kip1 in early- and
intermediate stage B-cell chronic lymphocytic leukemia lymphocytes
predicts rapid progression of the disease. Eur J Haematol.
82:260–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Giné E, Martinez A, Villamor N,
López-Guillermo A, Camos M, Martinez D, Esteve J, Calvo X,
Muntañola A, Abrisqueta P, et al: Expanded and highly active
proliferation centers identify a histological subtype of chronic
lymphocytic leukemia (accelerated chronic lymphocytic leukemia)
with aggressive clinical behavior. Haematologica. 95:1526–1533.
2010. View Article : Google Scholar
|
|
34
|
Huang H and Tindall DJ: Dynamic FoxO
transcription factors. J Cell Sci. 120:2479–2487. 2007. View Article : Google Scholar : PubMed/NCBI
|