Introduction
Onconase (P-30 protein), which is purified from
embryos of the Northern leopard frog (Rana pipiens), is a
member of the bovine pancreatic ribonuclease A (RNase A)
superfamily (1,2). It has a molecular weight of 12.0 kDa
and is composed of 104 amino acid (aa) residues (3). Although it is similar in molecular
structure to RNase A, it has two unique features: the presence of a
Cys87-Cys104 disulphide bond at the C-terminus; and presence of the
N-terminal pyroglutamic acid residue, which results from
cyclization of the glutamine residue (Gln) in the N-terminal and is
an integral part of the active site. Recombinant onconase, which
has an N-terminal methionine, has only l% activity compared to the
native protein. In addition, it has three disulfide bonds that
stabilize the protein structure and play a significant role in the
resistance to hydrolysis by proteases and heat (4,5).
Onconase has cytotoxicity towards various malignant
tumors such as breast, pancreatic, and non-small cell lung cancers,
as well as malignant mesothelioma (6–9).
Recombinant onconase was the first ribonuclease to be evaluated as
a therapeutic drug in humans, and has progressed to phase III
clinical trials in patients with unresectable mesothelioma. In
vivo and in vitro experiments have shown that onconase
has a synergistic effect with many other drugs including chemicals,
synthetic drugs, and cytokines (10–13).
Many cell surface receptors are overexpressed in multiple tumor
types including the HER2 receptor, luteinizing hormone-releasing
hormone receptor, and CXC chemokine receptor 4 (CXCR4) (14,15).
CXCR4 is overexpressed in more than 23 types of cancer, including
melanoma, glioma, acute myelogenous leukemia, chronic lymphocytic
leukemia, and breast, prostate, renal, pancreatic, ovarian,
cervical, colon, and small-cell lung cancers (16,17).
DV3 is the binding sequence of the CXCR4 ligand, thus it has the
potential to be used as a target to discern cancer cells in drug
design.
Cell-penetrating peptides (CPPs) penetrate the cell
membrane barrier and deliver various biological molecule cargoes
into the cytoplasm without receptors, including proteins, DNA, RNA,
and even nanoparticles (18–20).
As a typical CPP with the amino acid sequence PFVYLI, PFV delivers
fluorescent probes, RNA, and apoptotic peptides into a multiple of
cancer cells (21–23).
The methylotrophic yeast Pichia pastoris has
developed into a highly successful expression system for the
production of a variety of heterologous proteins using its strong
and tightly regulated AOX1 promoter (24,25).
This AOX1 promoter-controlled expression system depends on the
generation of biomass in medium without the use of methanol
(25). Despite the remarkable
achievements made with this expression system, the use of methanol
as the only carbon source can result in a series of issues, such as
the consumption of large amounts of dissolved oxygen, the need for
pure oxygen to maintain industrial production, and the proteolysis
of recombinant proteins during high density fermentation.
Furthermore, because methanol is an inflammable, explosive, and
toxic substance, it can be dangerous to use (26).
The glyceraldehyde-3-phosphate dehydrogenase (GAP)
promoter can be used for the constitutive expression of recombinant
heterologous proteins in P. pastoris cells using glucose,
glycerol, and methanol as carbon sources. Since the GAP
promoter-derived expression system allows for continuous production
of the recombinant product while avoiding methanol fed-batch
fermentation by P. pastoris, this system was proposed to be
more suitable for large-scale fermentation and production of
exogenous recombinant proteins (27,28).
The most used and best studied secretion leader in yeast is the
α-mating factor (α-MF) pre-pro-leader originating from S.
cerevisiae, which has been used for production of secretory
proteins with and without EAEA (Glu-Ala)2 overhang in
various yeast species. Of the three types of leading sequences that
can be used [pre sequence (19 aa), pre+pro sequence, pre+pro+EAEA],
only the pre sequence resulted in high onconase bioactivity.
Materials and methods
Materials
All restriction enzymes were purchased from New
England Biolabs (Beverly, MA, USA) and DNA polymerase was obtained
from Takara (Dalian, Liaoning, China). Yeast extracts and peptone
were purchased from Oxoid (Basingstoke, UK). Luria Bertani (LB)
medium and yeast extract peptone glucose (YPD) components were
purchased from Beijing Chemical Industry (Beijing, China).
Escherichia coli TOP10 cells were used for plasmid
maintenance. Zeocin, P. pastoris X33, and pGAPzαA were
purchased from Invitrogen (Carlsbad, CA, USA) and used for the
expression process.
Growth media and conditions
YPD broth (1% yeast extract, 2% peptone, and 2%
glucose) containing Zeocin (100, 200, 300, and 400 µg/ml)
was employed for selection of P. pastoris transformants at
30°C. Low salt LB (1% tryptone, 0.5% yeast extract, and 0.5% NaCl,
pH 7.0) with Zeocin (25 µg/ml) were employed to culture
E. coli TOP10 transformants at 37°C.
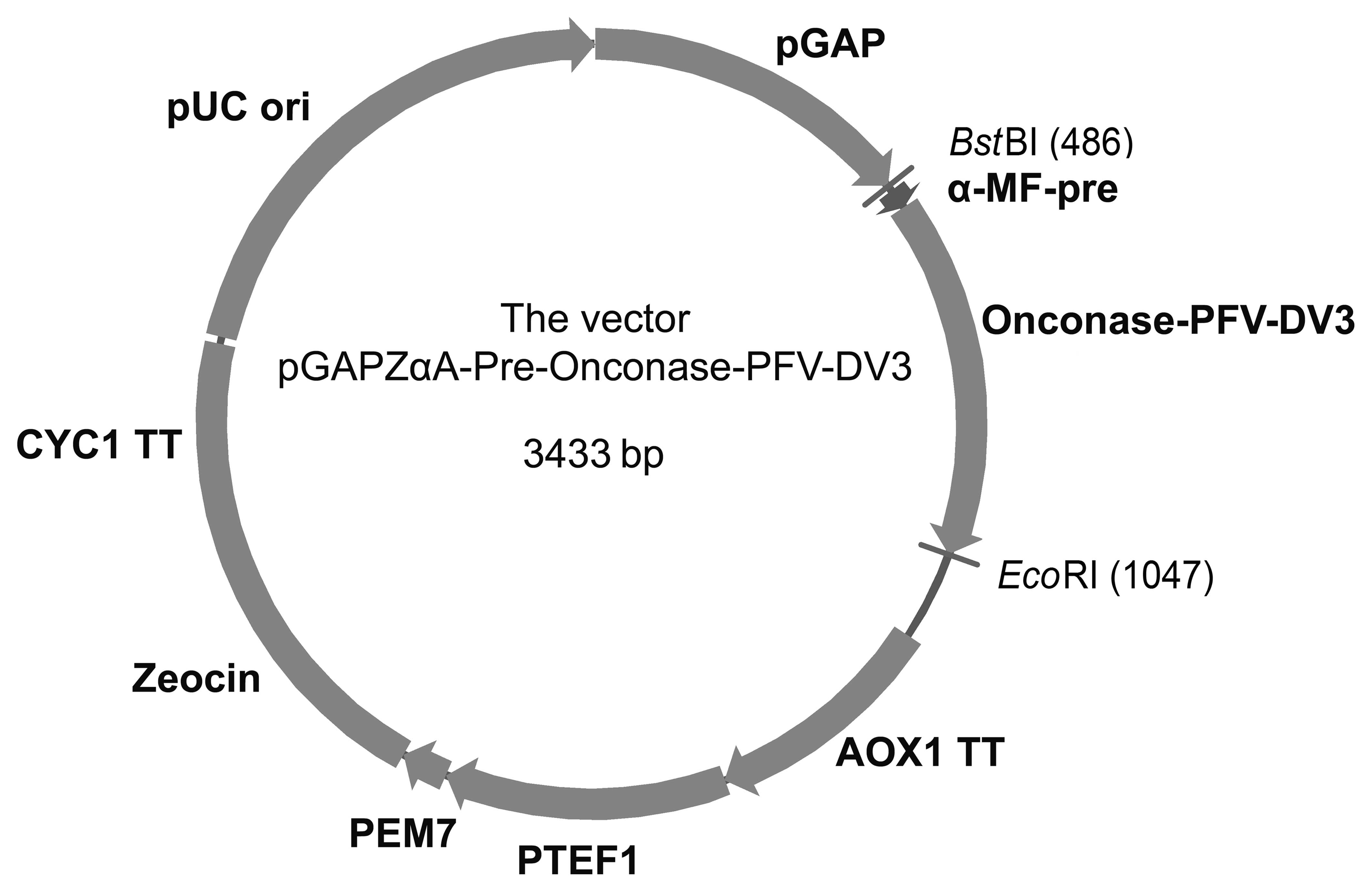
Plasmid and strains
A 560-base pair (bp) Pre-Onc-DV3 fusion gene
fragment was synthesized by GenScript Corporation (Nanjing, China)
after optimization. Both pGAPzαA and the Pre-Onc-DV3 gene were
double digested by BstBI and EcoRI, and the digested
gene was inserted into the open reading frame under control of the
GAP promoter, resulting in the pGAPZαA-Pre-onconase-PFV-DV3
expression vector (Fig. 1). The
recombinant plasmid was transferred into E. coli Top10
cells, and screened on low salt LB containing 25 µg/ml
Zeocin. The extracted recombinant plasmid was identified by
digesting with BglII and EcoRI, linearized with
BlnI, and transformed into P. pastoris X33 cells by
electroporation. The electrocompetent P. pastoris X33 cells
were prepared according to standard methods, and using an
electroporator (Scientz, Ningbo, China) following the
manufacturer's instructions. The electroporation conditions were
1.5 kV, 25 µF, and 200 Ω.
Screening of clones
The transformed X33 clones were screened on YPD agar
plates containing 100 µg/ml zeocin for 3 days. The chosen
transformants were screened for Zeocin-level clones from the YPD
agar plates containing a higher Zeocin concentration (200, 300, and
400 µg/ml). All of these high-level secretion clones were
confirmed by PCR using X33 transformant DNA, and selected according
to the Onc-DV3 recombinant protein concentration on SDS-PAGE. The
promising X33 transformants were grown in 10 ml YPD medium in a
100-ml flask for 24 h. Then, the culture was transferred to 100 ml
YPD medium for 24 h in a 1,000-ml flask. The inoculums were
cultivated to OD600 of 0.5 as starting seed cells to
investigate the effects of culture parameters: i) carbon source
cells were cultured in 2% (w/v) of different carbon sources
(glucose, glycerol, methanol, sucrose, sorbitol); ii) nitrogen
source cells were cultured in 2% (w/v) of different nitrogen
sources [tryptone, peptone, yeast extract, beef extract,
(NH4)2SO4]; iii) the cells were
cultured under different temperatures (21, 24, 27, 30, and 33°C);
and (iv) the cells were cultured under a different pH (4.5, 5.0,
5.5, 6.0, 6.5, 7.0, 7.5, 8.0).
The concentration of the Onc-DV3 recombinant protein
was determined by SDS-PAGE and RNA hydrolytic activity. All of the
data are expressed as the mean ± standard deviation (SD) of
independent triplicate determinations.
Western blot analysis
Western blot analysis was performed to determine the
concentration of Onc-DV3. Briefly, samples were collected at
various time-points (35, 45, 55, 65, and 75 h), and resolved by
SDS-PAGE. Then, the proteins were electrophoretically transferred
to polyvinylidene fluoride (PVDF) membranes (Millipore Co., Ltd.,
Boston, MA, USA) using a DYCZ-40G transfer blotter (Beijing Liuyi
Co., Ltd., Beijing, China). The membranes were incubated in
blocking buffer (10 mM Tris-HCl, 150 mM NaCl, 5% skim milk, pH 7.5)
at room temperature for 2 h, after which they were incubated with a
polyclonal antibody of DV3 at room temperature for 4 h. Next, the
membranes were washed three times with TBS containing 0.05%
Tween-20, and incubated with goat anti-rabbit IgG peroxidase
conjugate (diluted 1:100; Boster Co., Ltd., Wuhan, Hubei, China) at
room temperature for 4 h. The membrane was incubated with a
substrate solution containing 0.06% (w/v) chloronaphthol and 0.01%
H2O2 (both from Beijing Chemicals) after
washing.
Fed-batch fermentation
A 50-l fermentor was applied to evaluate the ability
for larger scale constitutive expression of Onc-DV3 immunotoxin
using the GAP promoter in P. pastoris, with the
aforementioned method. The time-course for biomass and production
of Onc-DV3 in the 50-l fermentor was determined. The production
value of the recombinant immunotoxin increased with biomass until
72 h. The biomass (OD600) was elevated to ~300 during
fed-batch fermentation in a 50-l fermentor. The production of
Onc-DV3 in the 50-l fermentor reached about 2×106 U/l
after 72 h of fermentation, compared to 6×105 U/l in a
shake flask.
Biomass analysis, protein concentration
determination, and cell viability assay
Biomass in the culture broth was determined by cell
density expressed as optical absorbance (OD600). The
protein concentration was determined using the Bradford assay kit
(Sangon, Shanghai). Cell growth was monitored using a model
UV-4802H spectrophotometer (Unico, Shanghai, China) at a wavelength
of 490 nm, and converted to dry cell weight using a predetermined
correlation factor.
Purification of onconase-DV3 fusion
protein
The culture broth was collected after 72 h, and
yeast cells were removed by centrifugation at 8,000 rpm for 30 min
at 4°C. The centrifuged liquid was ultrafiltrated and concentrated
by ultrafiltration using the Amfore ultrafiltration membrane 10000.
The concentrated recombinant immunotoxin was loaded onto a SP
Sepharose Fast Flow column (5.0×20 cm) that was pre-equilibrated
with buffer A (20 mM PB, pH 7.0). The bound recombinant immunotoxin
was eluted with buffer B (1 mol/l NaCl). The eluted protein was
mixed with buffer C (20 mM PB, pH 7.0) and further loaded onto the
Sephadex G-75 column (1.6×100 cm) that was pre-equilibrated with
the same buffer. The purity of the recombinant immunotoxin from the
final eluate was analyzed by SDS-PAGE.
Determination of the biological activity
of the onconase
In order to determine the RNA degradation activity
of onconase in vitro, 10 µg Pichia RNA was
added in a 100-µl reaction buffer (100 mmol/l Tris-HCl, pH
8.0), and then purified onconase solutions at different
concentrations (100, 10, 1, and 0.1 µg/ml) were added. The
tubes were incubated in a water bath at 37°C for 30 min. The
control group was incubated under the same conditions, but standard
RNase A was added to replace the onconase. RNA degradation was
assessed by agarose gel electrophoresis.
Determination of enzymatic parameters of
the expression products using the double reciprocal
With 1 mg/ml solution of yeast transfer RNA (tRNA)
as a substrate, various agents were successively added to assay the
enzymatic activity of the purified Onc-DV3 by drawing a portrait of
change values of OD260 along the RNase activity. The
experiment was repeated at least three times.
Cell lines, culture, and the MTT
assay
The paired highly metastatic and non-metastatic cell
lines used were as follows: i) PC-3M-1E8, a human prostate cell
line (highly metastatic), ii) PC-3M-2B4, a human prostate cell
line, iii) MDA-MB-231, a human breast carcinoma cell line (highly
metastatic), iv) MCF-7, a human breast carcinoma cell line, v)
PG-BE1, a human lung cancer cell line (highly metastatic), vi)
PG-LH7, a human lung cancer cell line, vii) L-02, a human normal
liver cell, and viii) HEK293, a human embryonic kidney cell
line.
All of the cell lines were kept in our laboratory in
RPMI-1640 medium (Invitrogen) with 20% fetal bovine serum and 10%
DMSO at −80°C. Cells were cultured in RPMI-1640 medium with 10%
fetal bovine serum, 100 U/ml penicillin, and 100 µg/ml
streptomycin at 37°C in 5% CO2. The cells (PC-3M-1E8,
PC-3M-2B4, MDA-MB-231, MCF-7, PG-BE1, PG-LH7, L-02, HEK293) were
seeded on 96-well flat-bottomed plates at 3,000 cells/well, and
incubated at 37°C in 5% CO2 atmosphere for 24 h. Then,
the cells were treated with Onc-DV3 after removing the medium at
serial dilutions (0.05, 0.10, 0.20, 0.40, 0.80, 1.60, and 3.20
µM). There were six individual wells for each dose. The
blank control was the dilution medium without added sample. The
cells were incubated in 96-well flat-bottomed plates at 37°C in 5%
CO2 atmosphere for 72 h, after which the supernatant was
removed, and the cells were washed with PBS (pH 7.4). The cells
were incubated in medium containing 20 µl MTT solution (5
mg/ml) at 37°C for 4 h. In order to dissolve the formed formazan
crystals, 100 µl dimethyl sulfoxide (DMSO) was added after
removing the supernatant. The OD was measured using the ELISA
microplate reader at 490 nm. Cytotoxicity to the cancer cells was
determined by survival rate (%). The experiment was repeated at
least three times. Data are presented as mean ± SD.
Results
Construction of an expression vector
A synthesized 560 bp Pre-Onc-DV3 gene was inserted
into the pGAPzαA vector under control of the GAP promoter, by
integrating the coding gene in frame with the S. cerevisiae
α-factor pre-secretion signal sequence. The constitutive expression
of pGAPzα-Pre-Onc-DV3 is shown (Fig.
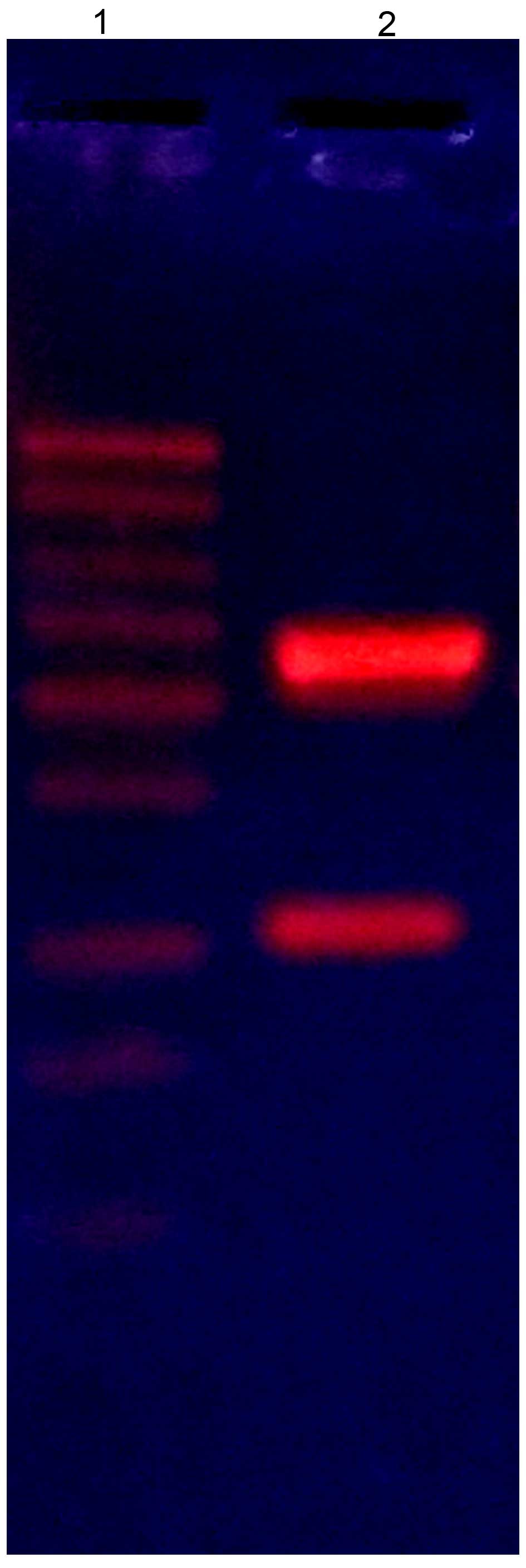
1). The extracted recombinant plasmid was identified by
digesting with BglII and EcoRI, producing a DNA
fragment ~1,050 bp (Fig. 2). To
confirm validity of the open reading frame, the full-length
promoter and Pre-Onc-DV3 was sequenced (data not shown). Zeocin was
used to screen the E. coli TOP10 transformants.
Screening of constitutive Pichia X33
clones of recombinant pGAP
The reconstructed vector was extracted and digested
with BlnI to obtain a linear recombinant vector. Then, the
linear vector of pGAPZαA-Pre-Onc-DV3 was employed to transform
P. pastoris X33 by electroporation. The X33 transformants
harboring the recombinant vector were screened depending on their
resistance to Zeocin on YPD plates containing different
concentrations of Zeocin. The 21 transformants were selected on YPD
plates containing 400 µg/ml Zeocin, and were confirmed by
genomic DNA PCR analysis. The expression ability of these
transformants was determined by measuring the medium supernatant
for RNA degradation efficiency.
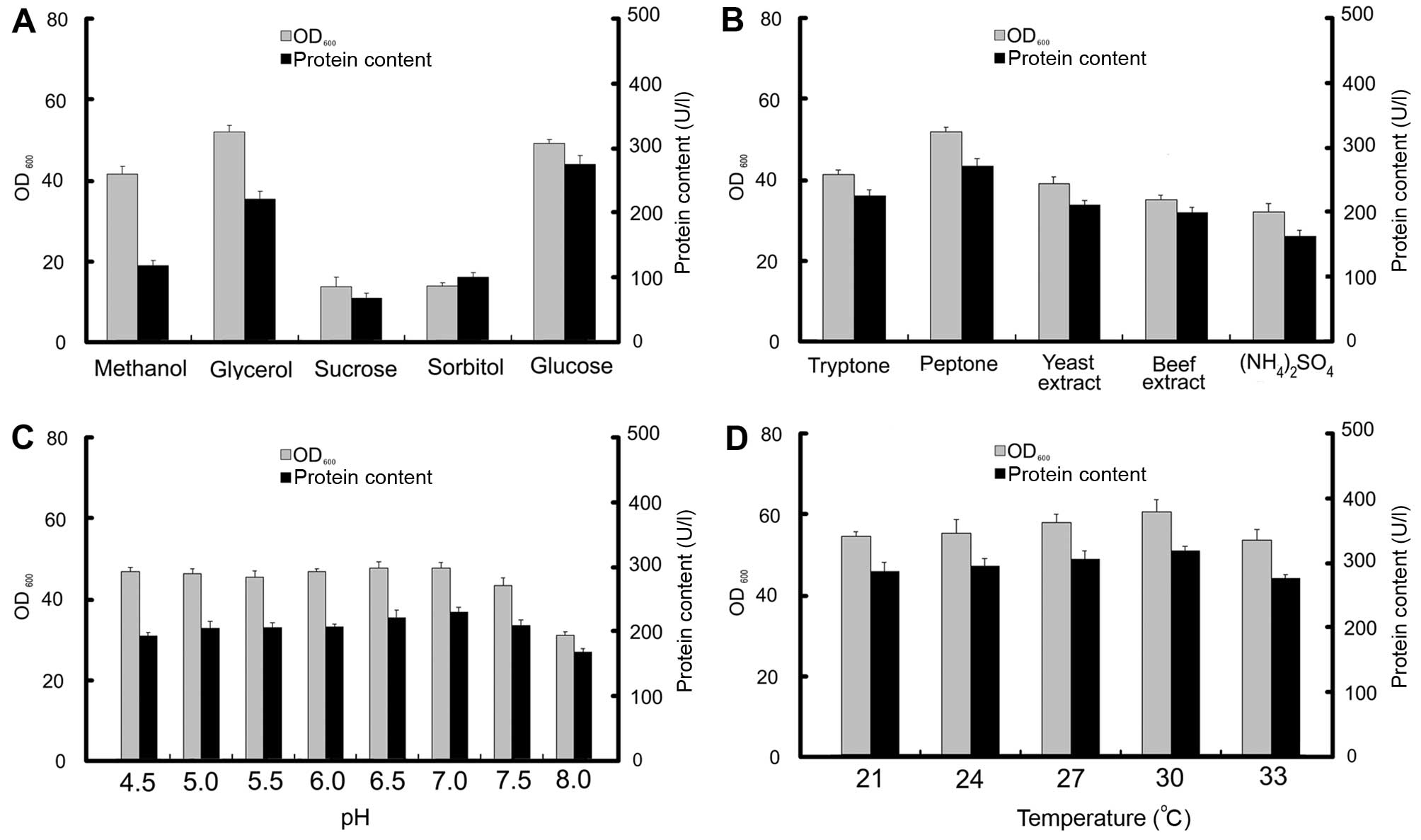
Effects of carbon source, temperature,
and pH on the production of onconase-DV3 immunotoxin
The effects of a few carbon sources on the
production of Onc-DV3 immunotoxin during the fermentation process
were evaluated. The X33 transformant with the highest expression
level of recombinant immunotoxin was selected as an initial seed
and cultured in modified YPD in which the glucose was 2% (w/v), but
was replaced with different carbon sources (glucose, sorbitol,
sucrose, methanol, or glycerol). The results demonstrated that
glucose was the most favorable carbon source, with a maximum
expression level of immunotoxin of ~310 U/l after 60 h of culture.
The results are shown in Fig. 3A.
Significantly, the biomass achieved the highest level when glycerol
was used as a carbon source, and immunotoxin protein production was
much lower than that of glucose as a carbon source. Compared to
glucose and glycerol, both the biomass and production level of
Onc-DV3 immunotoxin were much lower when using methanol, sucrose,
or sorbitol as the carbon source. The main nitrogen sources were
evaluated for their capacity to produce Onc-DV3 immunotoxin
[tryptone, peptone, yeast extract, beef extract,
(NH4)2SO4]. The results
demonstrated that peptone was the most favorable nitrogen source.
The maximum expression level of immunotoxin was ~310 U/l after 60 h
of culture (Fig. 3B). The cells
were grown in YPD medium (pH 7.0) at 21, 24, 27, 30, and 33°C to
evaluate the effects of temperature on biomass and Onc-DV3
immunotoxin production (Fig. 3D).
The biomass and production of Onc-DV3 did not show remarkable
variance under different temperatures. The selected temperature for
Onc-DV3 expression was at 30°C during fermentation, according to
the empirical of higher enzyme activity at higher culture
temperature (8). The effects of
different medium initial pH (4.5, 5.0, 5.5, 6.0, 6.5, 7.0, 7.5,
8.0) at 30°C on biomass and Onc-DV3 production were also checked
during culture. The results showed (Fig. 3C) that the initial pH of the YPD
medium had little effect on the biomass, but Onc-DV3 expression
level was highest at pH 7.0.
The expression levels of recombinant immunotoxin at
different times during the culture were analyzed by western blot
analysis using anti-onconase polyclonal antibody. The results are
shown in Figs. 4 and 5. The production of the recombinant
immunotoxin was presented at 35 h and reached the highest
production at 65 h in a flask culture.
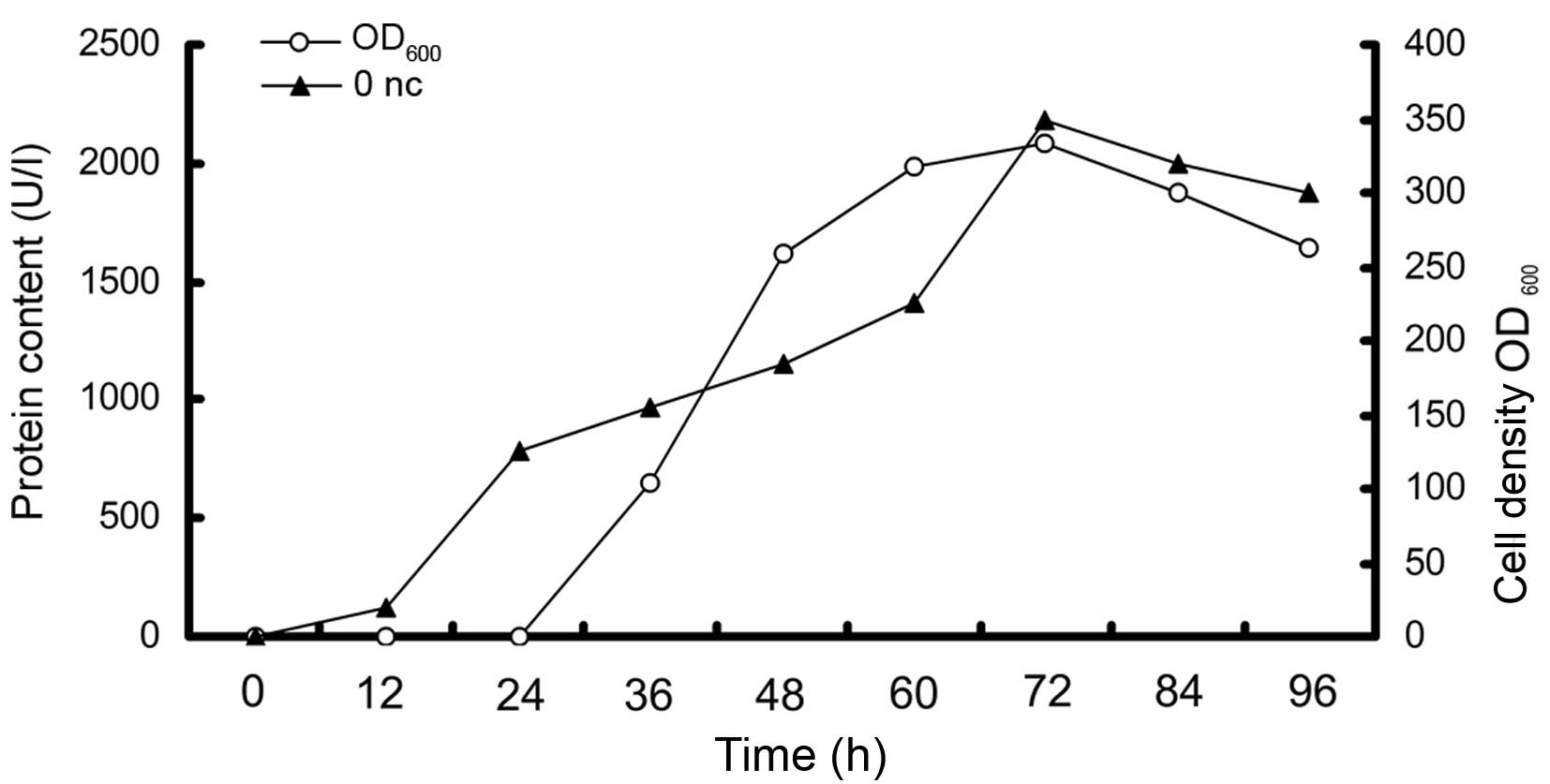
Fed-batch fermentation
By using optimized conditions, a 50-l fermentor was
applied to evaluate the ability for large-scale constitutive
expression of Onc-DV3 immunotoxin using GAP promoter in P.
pastoris, with the above selected method. The time-course for
biomass and production of Onc-DV3 in the 50-l fermentor are shown
in Fig. 6. The production value of
the recombinant immunotoxin increased with the biomass until 72 h.
The biomass (OD600) was elevated to ~300 during
fed-batch fermentation in a 50-l fermentor. The production of
Onc-DV3 in the 50 l fermentor reached about 2×103 U/l
after 72 h of fermentation compared with that in the shake flask
culture of 3.1×102 U/l. The results revealed that the
constitutive expression of the Onc-DV3 recombinant immunotoxin was
less hazardous to P. pastoris. It is viable to efficiently
produce Onc-DV3 recombinant immunotoxin in the 50 l fermentor by
adopting the constitutive fermentation strategy established in this
study.
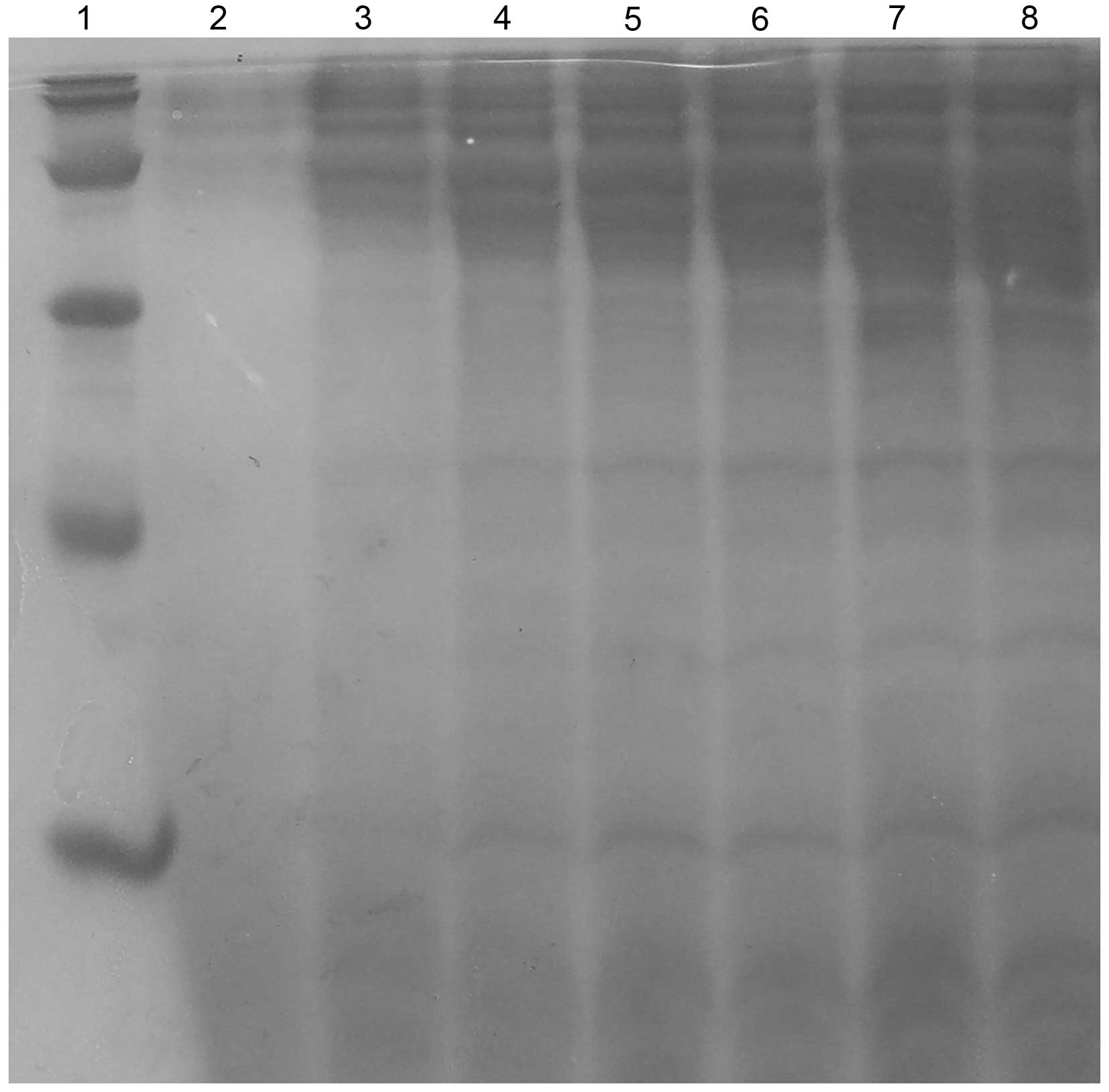
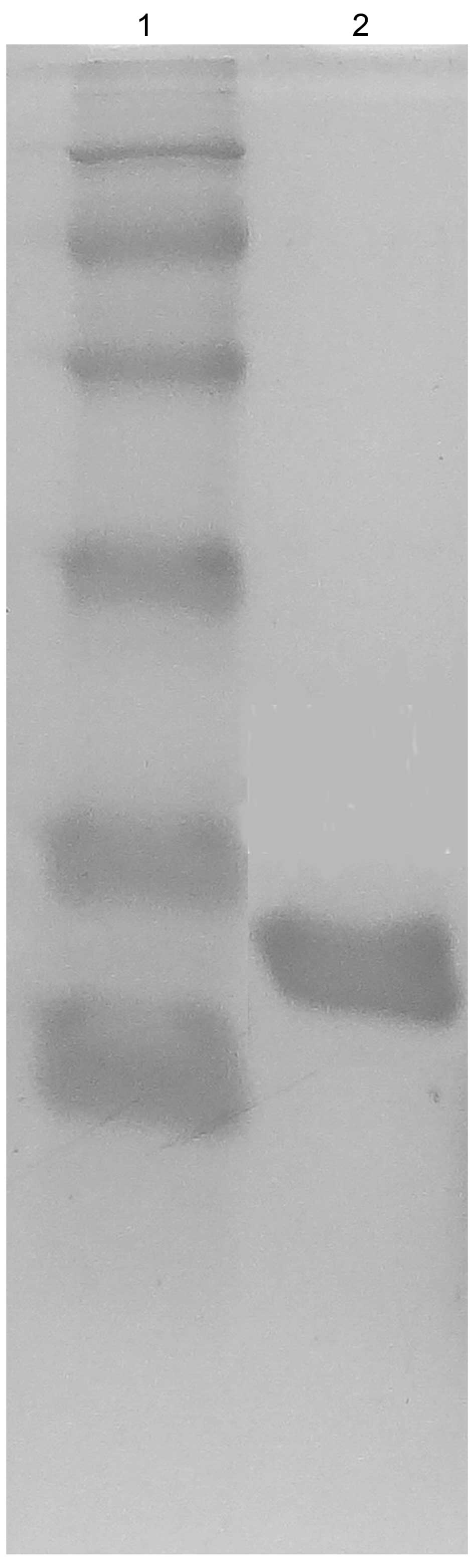
Purification and characterization of
onconase-DV3 recombinant immunotoxin
The recombinant immunotoxin was isolated from the
medium supernatant after ultrafiltration using membranes of 60 and
10 kDa and purified by SP Sepharose ion exchange chromatography and
Sephadex G-75 gel chromatography. The purity of the recombinant
immunotoxin reached 97.0% with the recovery level of 12.8%
(Table I). The purified immunotoxin
showed a single band on SDS-PAGE at ~20.0 kDa (Fig. 7), which was near the theoretical
value for this recombinant protein.
 | Table IA summary of the purification process
for the fusion protein onconase-DV3 produced by P.
pastoris. |
Table I
A summary of the purification process
for the fusion protein onconase-DV3 produced by P.
pastoris.
| Purification
step | Volume
(liters) | Onconase-DV3
(U/l) | Purity (%) | Cumulative yield
(%) |
|---|
| Culture
supernatant | 35 | 2,045 | – | 100.0 |
|
Ultrafiltration | 5 | 12,497 | – | 87.3 |
| SP Sepharose | 3 | 13,026 | 71.2 | 54.6 |
| Sephadex G75 | 5 | 3,836 | 96.1 | 26.8 |
The biological activity assay of Onc-DV3 was carried
out by agrose electrophoresis and increment ultraviolet absorption
(Fig. 8). RNA hydrolytic activity
of the biological activity of Onc-DV3 was analyzed according to
RNase assay from the onconase domain of Onc-DV3 (Table II). The results confirmed that the
recombinant immunotoxin Onc-DV3 produced by the GAP
promoter-derived expression system maintained high biological
activity.
| Table IIDetermination of RNase activity of
the immunotoxin product. |
Table II
Determination of RNase activity of
the immunotoxin product.
| Number | 1 | 2 | 3 | 4 | 5 | 6 | Sample | Sample | Sample |
|---|
| Tris-HCl buffer
(µl) | 850 | 850 | 850 | 850 | 850 | 850 | 850 | 850 | 850 |
| tRNA solution
(µl) | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Time (min) | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 |
| 50 µl Rnase
A (U) | 10 | 8 | 6 | 4 | 2 | 0 | – | – | – |
| Onc-DV3
(µl) | – | – | – | – | – | – | 400 | 400 | 400 |
| Change of
OD260 | 0.92 | 0.76 | 0.58 | 0.42 | 0.22 | 0 | 0.14 | 0.18 | 0.13 |
In conclusion, Onc-DV3 recombinant immunotoxin was
expressed under the control of the GAP promoter in the P.
pastoris expression system with high production and high
biological activity.
MTT assay
Every strain of the tumor cells was cultured and
treated with immunotoxin Onc-DV3 and incubated for 72 h. The
survival rate of the cells as indicated by OD490 is
shown in Table III. The trials of
Onc-DV3 with tumor cells showed high efficient inhibition in highly
metastatic tumor cells (PC-3M-1E8, PG-BE1, MDA-MB-231; the survival
rate varied from 24.79±7.31 to 27.61±9.16), middle efficiency on
poorly metastatic tumor cells (PC-3M-2B4, MCF-7, PG-LH7, the
survival rate varied from 53.38±17.37 to 59.64±8.03), and no effect
on normal cells (L-02, HEK293).
| Table IIIThe survival rate of cells treated
with 0.4 µM Onc-DV3. |
Table III
The survival rate of cells treated
with 0.4 µM Onc-DV3.
| Cell line | Survival rate
(%)a | P-value
(vs. control group) |
|---|
| PC-3M-1E8 | 24.79±7.31 | 0.007 |
| PG-BE1 | 25.85±6.72 | 0.005 |
| MDA-MB-231 | 27.61±9.16 | 0.032 |
| PC-3M-2B4 | 53.38±17.37 | 0.016 |
| MCF-7 | 55.19±10.85 | 0.028 |
| PG-LH7 | 59.64±8.03 | 0.019 |
| L-02 | 107.13±21.80 | 0.041 |
| HEK293 | 125.79±19.32 | 0.027 |
Discussion
The metastasis of tumor cells is the main cause of
cancer-related death due to the rapid proliferation, the poor
differentiation and the limited clinical efficacy of traditional
treatments on tumor metastasis. Targeted cancer therapies based on
immunological recognition of tumor cell-specific targets are
expected to be more effective than traditional treatments and less
harmful to normal cells.
The DV3 can selectively bind to metastatic tumors,
but has no cytotoxicity to induce tumor cell apoptosis. Therefore,
it needs to be linked with a molecule which is capable of causing
tumor cell apoptosis.
Onconase has demonstrated its antitumor activity on
several types of tumors and has been developed as a drug for
malignant pleural mesothelioma (MPM).
A bivalent targeted domain has been reported to be
more effective than using a single targeted domain for targeting
against the receptor. The dimer DV3 markedly improved the tumor
cell killing activity than the single one. Since multi-copy seems
to cause highly effective internalization (29–33),
(DV3)2 with onconase may provide a more efficient
approach for delivering RNase payloads into the cytosol of
metastatic tumor cells.
The main systemic toxicity of onconase is the damage
of proximal kidney tubular cells in which onconase exhibits highly
non-specific uptake by tissues (34,35).
The linkage of onconase and (DV3)2 is crucial so that it
can specifically target metastatic tumor tissue.
In the present study, two copies of DV3 were
selected to be linked to onconase to prepare an antitumor drug for
metastatic tumors.
The best method of preparation was to construct the
fusion gene of (DV3)2 and onconase, but formation of the
cyclization of the glutamine residue (Gln) in the N-terminal
contained by onconase may be difficult to be expressed in P.
pastoris. Guided by the α-mating factor, high bioactivity and
production were obtained.
Since it was not clear whether the carbon source,
the nitrogen source, the temperature, or pH affect the expression
level during the fermentation of Onc-(DV3)2, in order to
ensure a high level of expression of Onc-DV3, these parameters were
evaluated.
We demonstrated that the combination of onconase to
a bivalent DV3 caused selective and specific cytotoxicity only
towards metastatic tumor cells whereas onconase alone mediated
cytotoxicity towards both metastatic and non-metastatic tumor
cells.
The results of the cytotoxicity experiments of the
DV3 + onconase mixture, onconase-single DV3, and onconase-double
DV3 revealed a progressive antitumor effect, suggesting that double
DV3 could deliver more target Onc to metastatic tumor cells and
selectively enhance the cytotoxicity of onconase.
According to its specific and selective antitumor
efficacy on metastatic tumor cells, we expect that further studies
of Onc-DV3 could act in its development as a clinical drug.
Acknowledgments
This study was supported by Jilin Province Science
and Technology Key Problem of China (20140203014YY).
References
|
1
|
Darzynkiewicz Z, Carter SP, Mikulski SM,
Ardelt WJ and Shogen K: Cytostatic and cytotoxic effects of Pannon
(P-30 protein), a novel anticancer agent. Cell Tissue Kinet.
21:169–182. 1988.PubMed/NCBI
|
|
2
|
Mosimann SC, Ardelt W and James MN:
Refined 1.7 A X-ray crystallographic structure of P-30 protein, an
amphibian ribonuclease with anti-tumor activity. J Mol Biol.
236:1141–1153. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ardelt W, Mikulski SM and Shogen K: Amino
acid sequence of an anti-tumor protein from Rana pipiens oocytes
and early embryos. Homology to pancreatic ribonucleases. J Biol
Chem. 266:245–251. 1991.PubMed/NCBI
|
|
4
|
Gorbatyuk VY, Tsai CK, Chang CF and Huang
TH: Effect of N-terminal and Met23 mutations on the structure and
dynamics of onconase. J Biol Chem. 279:5772–5780. 2004. View Article : Google Scholar
|
|
5
|
Bretscher LE, Abel RL and Raines RT: A
ribonuclease A variant with low catalytic activity but high
cytotoxicity. J Biol Chem. 275:9893–9896. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu Y, Mikulski SM, Ardelt W, Rybak SM and
Youle RJ: A cytotoxic ribonuclease. Study of the mechanism of
onconase cytotoxicity. J Biol Chem. 268:10686–10693.
1993.PubMed/NCBI
|
|
7
|
Saxena SK, Sirdeshmukh R, Ardelt W,
Mikulski SM, Shogen K and Youle RJ: Entry into cells and selective
degradation of tRNAs by a cytotoxic member of the RNase A family. J
Biol Chem. 277:15142–15146. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suhasini AN and Sirdeshmukh R: Transfer
RNA cleavages by onconase reveal unusual cleavage sites. J Biol
Chem. 281:12201–12209. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rodríguez M, Torrent G, Bosch M, Rayne F,
Dubremetz JF, Ribó M, Benito A, Vilanova M and Beaumelle B:
Intracellular pathway of onconase that enables its delivery to the
cytosol. J Cell Sci. 120:1405–1411. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Newton DL, Hansen HJ, Mikulski SM,
Goldenberg DM and Rybak SM: Potent and specific antitumor effects
of an anti-CD22-targeted cytotoxic ribonuclease: Potential for the
treatment of non-Hodgkin lymphoma. Blood. 97:528–535. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pavlakis N and Vogelzang NJ: Ranpirnase -
an antitumour ribonuclease: Its potential role in malignant
mesothelioma. Expert Opin Biol Ther. 6:391–399. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee I, Kalota A, Gewirtz AM and Shogen K:
Antitumor efficacy of the cytotoxic RNase, ranpirnase, on A549
human lung cancer xenografts of nude mice. Anticancer Res.
27:299–307. 2007.PubMed/NCBI
|
|
13
|
Chang CH, Gupta P, Michel R, Loo M, Wang
Y, Cardillo TM and Goldenberg DM: Ranpirnase (frog RNase) targeted
with a humanized, internalizing, anti-Trop-2 antibody has potent
cytotoxicity against diverse epithelial cancer cells. Mol Cancer
Ther. 9:2276–2286. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu XF and Xing PX: Discovery and
validation of new molecular targets for ovarian cancer. Curr Opin
Mol Ther. 5:625–630. 2003.
|
|
15
|
Ménard S, Pupa SM, Campiglio M and
Tagliabue E: Biologic and therapeutic role of HER2 in cancer.
Oncogene. 22:6570–6578. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Balkwill F: The significance of cancer
cell expression of the chemokine receptor CXCR4. Semin Cancer Biol.
14:171–179. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Snyder EL, Saenz CC, Denicourt C, Meade
BR, Cui XS, Kaplan IM and Dowdy SF: Enhanced targeting and killing
of tumor cells expressing the CXC chemokine receptor 4 by
transducible anticancer peptides. Cancer Res. 65:10646–10650. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saar K, Lindgren M, Hansen M, Eiríksdóttir
E, Jiang Y, Rosenthal-Aizman K, Sassian M and Langel U:
Cell-penetrating peptides: A comparative membrane toxicity study.
Anal Biochem. 345:55–65. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu S, Tager LA, Chitale S and Riley LW: A
cell-penetrating peptide derived from mammalian cell uptake protein
of Mycobacterium tuberculosis. Anal Biochem. 353:7–14. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Snyder EL and Dowdy SF: Cell penetrating
peptides in drug delivery. Pharm Res. 21:389–393. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rhee M and Davis P: Mechanism of uptake of
C105Y, a novel cell-penetrating peptide. J Biol Chem.
281:1233–1240. 2006. View Article : Google Scholar
|
|
22
|
Watkins CL, Brennan P, Fegan C, Takayama
K, Nakase I, Futaki S and Jones AT: Cellular uptake, distribution
and cytotoxicity of the hydrophobic cell penetrating peptide
sequence PFVYLI linked to the proapoptotic domain peptide PAD. J
Control Release. 140:237–244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park JW, Bang EK, Jeon EM and Kim BH:
Complexation and conjugation approaches to evaluate siRNA delivery
using cationic, hydrophobic and amphiphilic peptides. Org Biomol
Chem. 10:96–102. 2012. View Article : Google Scholar
|
|
24
|
Macauley-Patrick S, Fazenda ML, McNeil B
and Harvey LM: Heterologous protein production using the Pichia
pastoris expression system. Yeast. 22:249–270. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Damasceno LM, Huang CJ and Batt CA:
Protein secretion in Pichia pastoris and advances in protein
production. Appl Microbiol Biotechnol. 93:31–39. 2012. View Article : Google Scholar
|
|
26
|
Sinha J, Plantz BA, Inan M and Meagher MM:
Causes of proteolytic degradation of secreted recombinant proteins
produced in methylotrophic yeast Pichia pastoris: Case study with
recombinant ovine interferontau. Biotechnol Bioeng. 89:102–112.
2005. View Article : Google Scholar
|
|
27
|
Vassileva A, Chugh DA, Swaminathan S and
Khanna N: Expression of hepatitis B surface antigen in the
methylotrophic yeast Pichia pastoris using the GAP promoter. J
Biotechnol. 88:21–35. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang X, Sun Y, Ke F, Zhao H, Liu T, Xu L,
Liu Y and Yan Y: Constitutive expression of yarrowia lipolytica
lipase LIP2 in Pichia pastoris using GAP as promoter. Appl Biochem
Biotechnol. 166:1355–1367. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pedersen MW, Jacobsen HJ, Koefoed K, Hey
A, Pyke C, Haurum JS and Kragh M: Sym004: A novel synergistic
anti-epidermal growth factor receptor antibody mixture with
superior anticancer efficacy. Cancer Res. 70:588–597. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Q, Villeneuve G and Wang Z: Control
of epidermal growth factor receptor endocytosis by receptor
dimerization, rather than receptor kinase activation. EMBO Rep.
6:942–948. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fan Z, Lu Y, Wu X and Mendelsohn J:
Antibody-induced epidermal growth factor receptor dimerization
mediates inhibition of autocrine proliferation of A431 squamous
carcinoma cells. J Biol Chem. 269:27595–27602. 1994.PubMed/NCBI
|
|
32
|
Perez-Torres M, Guix M, Gonzalez A and
Arteaga CL: Epidermal growth factor receptor (EGFR) antibody
down-regulates mutant receptors and inhibits tumors expressing EGFR
mutations. J Biol Chem. 281:40183–40192. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vasandani VM, Burris JA and Sung C:
Reversible nephrotoxicity of onconase and effect of lysine pH on
renal onconase uptake. Cancer Chemother Pharmacol. 44:164–169.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mikulski S, Grossman A, Carter P, Shogen K
and Costanzi J: Phase-I human clinical-trial of onconase(r) (p-30
protein) administered intravenously on a weekly schedule in
cancer-patients with solid tumors. Int J Oncol. 3:57–64.
1993.PubMed/NCBI
|
|
35
|
Vasandani VM, Wu YN, Mikulski SM, Youle RJ
and Sung C: Molecular determinants in the plasma clearance and
tissue distribution of ribonucleases of the ribonuclease A
superfamily. Cancer Res. 56:4180–4186. 1996.PubMed/NCBI
|