Introduction
Hepatocellular carcinoma (HCC) represents the most
common type of primary liver cancer (1). HCC is a highly prevalent and lethal
neoplasia, and the management of HCC has significantly improved
during the last few years (2). A
better understanding of the pathogenesis and the molecular
mechanisms facilitate a better prediction of prognosis and the most
appropriate treatment approach. At present, curative therapies
including resection and transplantation have increased the survival
rate of HCC patients at an early stage, while those diagnosed at
intermediate and advanced stages still suffered from a high risk of
side-effects (3–5). Investigation of the cellular origins
and the early molecular events resulting in HCC are urgently
necessary for further understanding HCC disease progression.
The transforming growth factor β (TGF-β) superfamily
of cytokines is composed of TGF-βs, activins, inhibins, bone
morphogenetic proteins (BMPs) and anti-Müllerian hormone (AMH) and
are expressed in all multicellular organisms (6). In mammals, the TGF-β family exerts
regulatory functions on cell growth, differentiation, apoptosis,
immunity, extracellular matrix (ECM) production, adhesion and even
embryonic development. Alterations of the components of the TGF-β
signaling pathway and abnormal production of TGF-βs are often
observed in disease progression despite the involvement in the
maintenance of tissue homeostasis under both normal and dynamic
conditions (7,8). The TGF-β isoforms (TGF-β1-3) exert
their biological effects by binding to three receptors: type I
(TβRI), type II (TβRII) and type II (TβRIII). Once bound to TGF-β,
the constitutively active TβRII recruits, binds and activates TβRI,
thereby stimulating its protein kinase activity. The downstream
substrates for TβRI are members of the receptor-activated Smads
(R-Smads), namely Smad2 and Smad3. After inducing phosphorylation
by receptors, R-Smads form complexes with common mediated Smad4
(Co-Smad), and then translocate to the nucleus where regulation of
the transcription of certain genes occurs in cooperation with other
transcription factors (9).
TβRIII is the most abundantly expressed and
characterized TGF-β receptor, and is a proteoglycan comprised of
851 amino acids (10,11). As a TGF-β superfamily co-receptor,
the role of TβRIII in regulating TGF-β signaling is
context-dependent and complex (12,13).
The significant and indispensable role of TβRIII in mediating
signaling is exerted dependently through TβRII and TβRI as well as
potential signaling independently through TβRII and TβI.
Overexpression of TβRIII was found to restore TGF-β1 sensitivity in
CAL-27 human oral squamous cells (14). In MDA-MB-231 human breast cancer
cells, TβRIII expression inhibited Smad2/3 signaling dependent on
the TβRIII cytoplasmic domain, and the independent binding of
TβI/TβRII to TβRIII competed with TβI/TβRII signaling complex
formation, thus inhibiting TGF-β-mediated Smad signaling (15). Importantly, the alteration of TβRIII
expression level and the mediating roles have been observed in the
progression of several types of cancers including breast, prostate,
lung and ovarian cancer (16–19).
Furthermore, the anchorage-independent growth of human MCF-7 breast
cancer cells was inhibited when TβRIII expression was recovered
in vitro (20), and
suppression of tumorigenicity occurred in MDA-MB231 breast cancer
cells after increased TβRIII expression in athymic nude mice
(21). The expression level of
TβRIII and its regulatory role in HCC progression remain to be
clarified.
Therefore, in the present study, we investigated the
expression of TβRIII at both the mRNA and protein levels in HCC
patient tissues and cell lines. In vitro, we also studied
the regulatory roles of TβRIII in metastasis and invasion of HCC
and the related mechanisms.
Materials and methods
HCC patient tissues
Quantitative real-time PCR (qRT-PCR) and western
blotting were utilized to examine the expression of TβRIII at the
mRNA and protein level, respectively, in 10 HCC tissue specimens,
which were collected at the First Affiliated Hospital of Anhui
Medical University. All of the tissue specimens were pathologically
confirmed as primary hepatic carcinoma and were promptly placed
into sterile vials, and stored at −80°C until required. Tissue
collection was approved by the Ethics Committee of Anhui Medical
University. All procedures involving specimens obtained from human
subjects were performed following informed consent from the
patients.
Cell culture
Human HCC cell lines with stepwise metastatic
potential (HepG2, SMMC-7721, MHCC97H, HCCLM3) and the normal
immortalized liver cell line L02 were cultured in Dulbecco's
modified Eagle's medium (DMEM), supplemented with 10% (v/v) fetal
bovine serum (FBS) (both from Gibco, Life Technologies, USA) and
100 U/ml of penicillin and 100 µg/ml of streptomycin. Human
normal liver L02 cells, and human HCC cell lines HepG2 and
SMMC-7721 were purchased from Shanghai Cell Bank, Chinese Academy
of Sciences (Shanghai, China). MHCC97H and HCCLM3 cells were
purchased from Shanghai Institute of Biochemistry and Cell Biology,
Chinese Academy of Science (Shanghai, China). All of the cell lines
were maintained at 37°C in a 5% CO2 atmosphere.
Quantitative real-time PCR (qRT-PCR)
qRT-PCR was carried out to examine the mRNA
expression of TβRIII. Glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) served as an internal control. Total RNA was extracted
using TRIzol reagent (Invitrogen, Life Technologies, USA). Total
RNA (2 µg) was reverse transcribed using the RevertAid First
Strand cDNA Synthesis kit (Fermentas, Lithuania) according to the
manufacturer's instructions. The detection was carried out in a
real-time PCR detection system (ABI 7500) using the SYBR-GreenER
qPCR SuperMix Universal kit (Invitrogen, Life Technologies)
according to the manufacturer's instructions. The primer sequences
for each gene were: GAPDH forward primer,
5′-AGGTCGGAGTCAACGGATTTG-3′ and reverse primer,
5′-CCTGGAAGATGGTGATGGGAT-3′; TβRIII forward primer,
5′-ACCCCCAACTCTAACCC CTACA-3′ and reverse primer,
5′-GCCAATACTGTTAGGACAATAATTTTC-3′. The cycle threshold value was
defined as the PCR cycle number at which the reporter fluorescence
crosses the threshold. The cycle threshold value of each product
was determined and normalized against that of the internal control,
GAPDH.
TβRIII small interfering RNA (siRNA)
transfection
According to the TβRIII gene sequences, three siRNA
duplexes targeting the TβRIII gene were designed (GenePharma,
China): TβRIII-homo-2729 sense, 5′-GGGCCAUGAUGCAGAAUAATT-3′ and
antisense, 5′-UUAUUCUGCAUCAUGGCCCTT-3′; TβRIII-homo-2059 sense,
5′-GGUGUGGUCUACUAUAACUTT-3′ and antisense,
5′-AGUUAUAGUAGACCACACCTT-3′; TβRIII-homo-759 sense,
5′-GGUCACACUUCACCUGAAUTT-3′ and antisense,
5′-AUUCAGGUGAAGUGUGACCTT-3′. The transfection operation was
performed under guidance of the Lipofectamine 3000 protocol
(Invitrogen, Life Technologies). The transfection efficiency was
confirmed by western blotting.
Western blotting
Liver samples and human HCC cell lines were
collected and cultured respectively, according to the methods
described above. Liver tissue was homogenized in RIPA lysis buffer
and phenylmethylsulfonyl fluoride (PMSF) at a ratio of 99:1 (v/v).
The cells were washed twice with ice-cold phosphate-buffered saline
(PBS) after the corresponding treatment and lysed in cell lysis
buffer. The protein concentration was determined with the BCA
protein assay kit (Thermo Fisher Scientific, USA). A protein sample
was mixed with 5x sample buffer (4:1) (Bio-Rad, Hercules, CA, USA)
and heated in boiling water for 10 min. The proteins were separated
by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE), transferred to polyvinylidene fluoride (PVDF) membranes
(Millipore, Bedford, MA, USA), and incubated with blocking buffer
(0.05% Tween-20 PBS with 5% non-fat milk) for 2 h. Immunoblots were
incubated with the indicated primary antibodies overnight at 4°C,
followed by the appropriate horseradish peroxidase-conjugated
secondary antibody for 2 h at room temperature, and immunodetection
was visualized by enhanced chemiluminescence (Pierce, Rockford, IL,
USA) using hydrogen peroxide and luminol as substrates.
Autoradiographs were scanned using ImageQuant LAS 4000 mini (GE
Healthcare Bio-Sciences AB, Uppsala, Sweden). The density of the
specific bands was quantified using ImageJ software (National
Institutes of Health, Bethesda, MD, USA).
Wound healing assay
SMMC-7721 cells were seeded in a 6-well plate at
70–90% confluency and the cell monolayer was scraped in a straight
line to create a 'scratch' with a P200 pipette tip. The debris was
removed and the edge of the scratch was smoothed by washing the
cells twice with 1 ml PBS. Then, 2 ml serum-free DMEM was added
with different concentrations of TGF-β1 (0.2, 1, 5 and 25 ng/ml)
and images were captured using a computer-based microscopic imaging
system at the 0 h time point at a magnification of ×200. After 24
h, images of the wound were captured again under magnification of
×200.
Changes in the migratory ability of the human
SMMC-7721 cells after transfection with TβRIII siRNA were also
assessed by wound healing assay. SMMC-7721 cells were seeded to
70–90% confluency and starved for 12 h. TβRIII siRNA was
transfected into the SMMC-7721 cells under the guidance of the
Lipofectamine 3000 protocol. After 12 h, the wound was created
using the same method as described above. Images at 0 and 24 h were
captured by a computer-based microscopy imaging system under a
magnification of ×200, respectively. Cell motility was evaluated
according to the following formula: Cell motility ratio = (distance
24 h -distance 0 h)/distance 0 h.
Transwell assay
SMMC-7721 cells plated in 6-well plates previously
transfected with the TβRIII siRNA for 12 h were harvested by
trypsinization. The cells were washed once with PBS to remove the
influence of FBS and then the cell pellet was resuspended in
serum-free DMEM at 1×106 cells/ml. The cells
(25,000–50,000) were seeded in the upper chamber of a Transwell
filter, coated with Matrigel (BD Biosciences) in order to determine
the cell invasive ability. The cells were cultured for 48 h at 37°C
in a 5% CO2 incubator through Matrigel toward the lower
chamber, containing medium with 10% FBS as the chemoattractant.
After incubation, the medium in the upper chamber was discarded and
then the upper chamber was removed gently from the 24-well plates.
The filter was fixed in 90% alcohol for 30 min and hexamethyl
pararosaniline staining for 5 min. Images were captured of
representative fields of each filter, and the number of cells was
quantified.
Statistical analysis
All data are expressed as means ± SD. Statistical
analysis was performed using one-way ANOVA. Statistical
calculations were conducted using SPSS 10.0 soft ware. The results
were considered statistically significant for P-value <0.05.
Results
Decreased expression of TβRIII in HCC
patient tissues
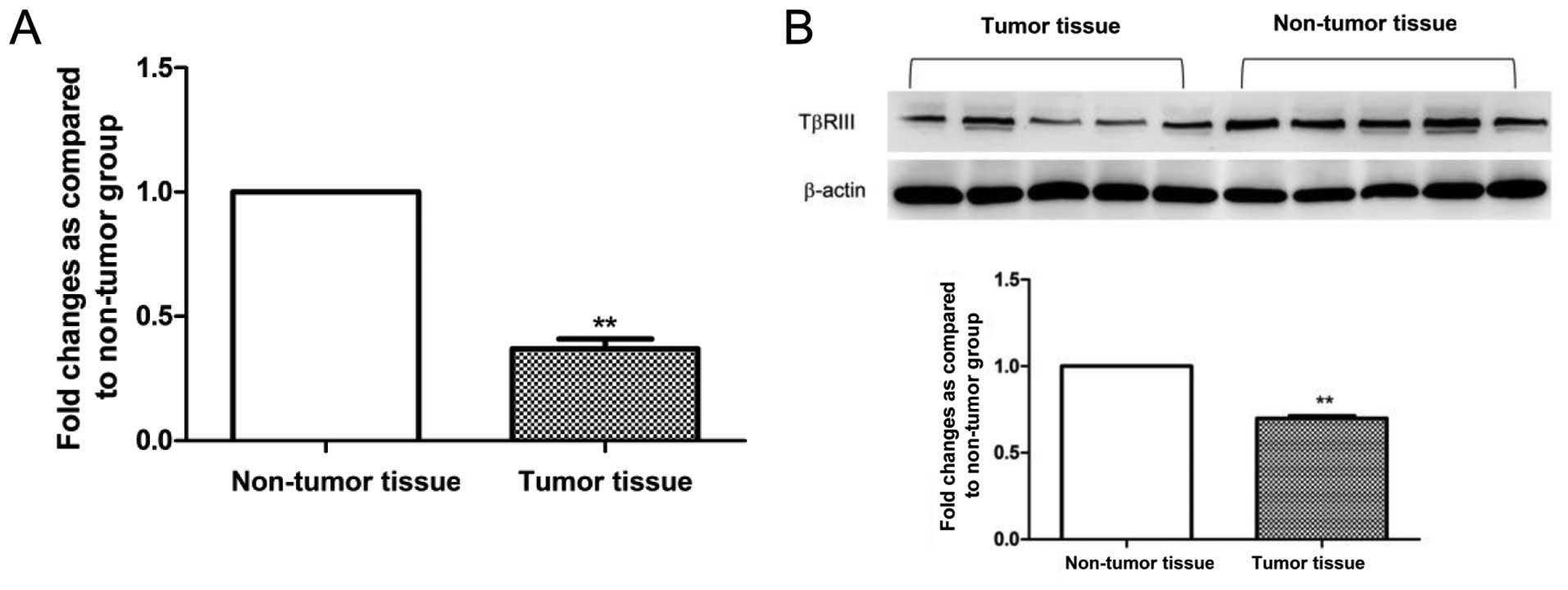
To investigate whether TβRIII expression is altered
in HCC patient specimens, we initially analyzed the mRNA expression
level in 10 human HCC tissue samples and compared the level with
the matched non-tumor tissue samples as normal controls by qRT-PCR.
From all the specimens, we observed a significant decrease in
TβRIII mRNA expression in tumor tissues compared with the matched
non-tumor tissue samples (Fig. 1A).
To confirm decreased expression of TβRIII in HCC patient tissues,
we examined TβRIII expression at the protein level by western blot
analysis. The result showed that the protein expression of TβRIII
was significantly decreased in the HCC tumor tissues compared with
the level in the matched non-tumor tissue samples (Fig. 1B). The results revealed the impaired
expression of TβRIII at both the mRNA and protein level.
Decreased expression of TβRIII in human
HCC cell lines along with increased metastatic potential
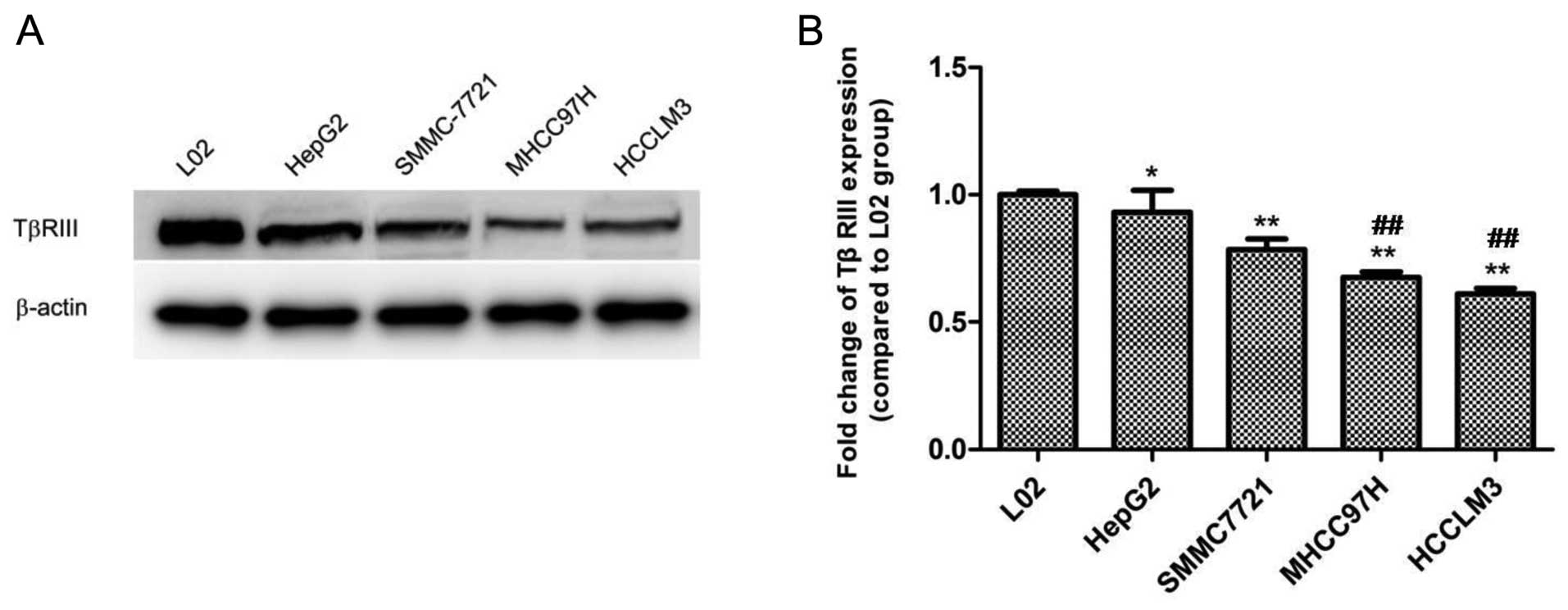
To further confirm the alteration in the expression
in TβRIII in HCC progression, human liver L02 cells and four HCC
cell lines with stepwise metastatic potential (HepG2, SMMC-7721,
MHCC97H and HCCLM3) were selected to observe the expression change
of TβRIII in vitro by western blotting. Western blot results
demonstrated the highest expression level in human liver L02 cells,
while the expression of TβRIII was gradually decreased with
increasing metastatic potential in the HCC cell lines. Moreover,
the expression of TβRIII in the HCC cell lines MHCC97H and HCCLM3
which possess high metastatic potential was markedly lower than
that in the HepG2 cells, an HCC cell line with low metastatic
potential (Fig. 2). These results
showed that a reduction in the expression of TβRIII occurred
simultaneously with the increasing metastatic potential of the HCC
cells.
TGF-β1 treatment enhances the migratory
ability of human HCC cell lines
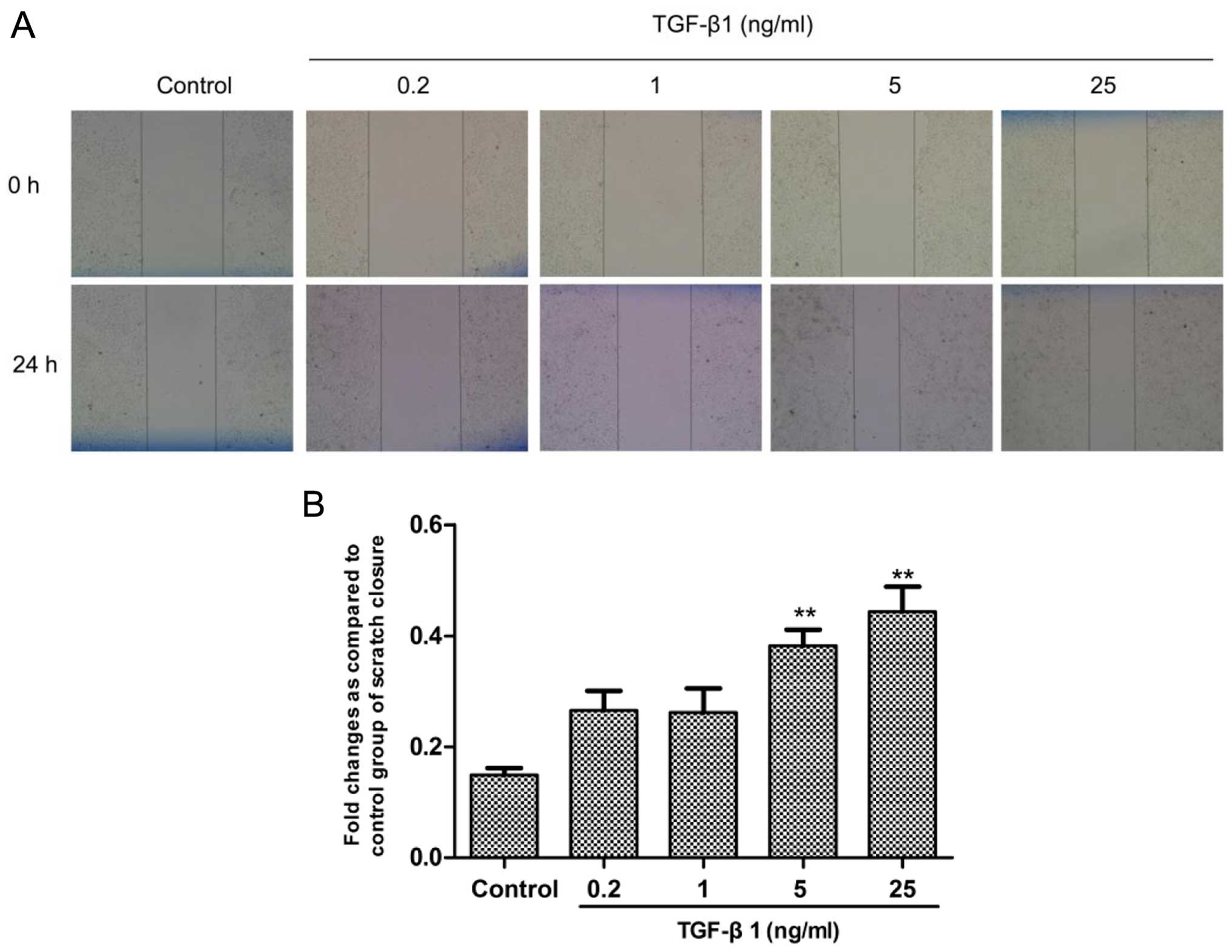
TGF-β1 is overexpressed and involved in the
regulation of the progression of various types of cancer, and we
investigated the effect of TGF-β1 on the migratory ability of human
HCC cells. Human SMMC-7721 cells were seeded in a 6-well plate
(5×105 cells/well) and starved for 12 h on the following
day. Then, SMMC-7721 cells were exposed to TGF-β1 for 24 h at
concentrations of 0.2, 1, 5 and 25 ng/ml. The wound healing assay
showed that TGF-β1 significantly elevated the migratory ability of
the SMMC-7721 cells in a concentration-dependent manner,
particularly in the 5 and 25 ng/ml treatment groups (Fig. 3). The results indicated that TGF-β1
promoted the metastasis of the HCC cells.
TGF-β1 stimulation reduces the expression
of TβRIII in human HCC cells
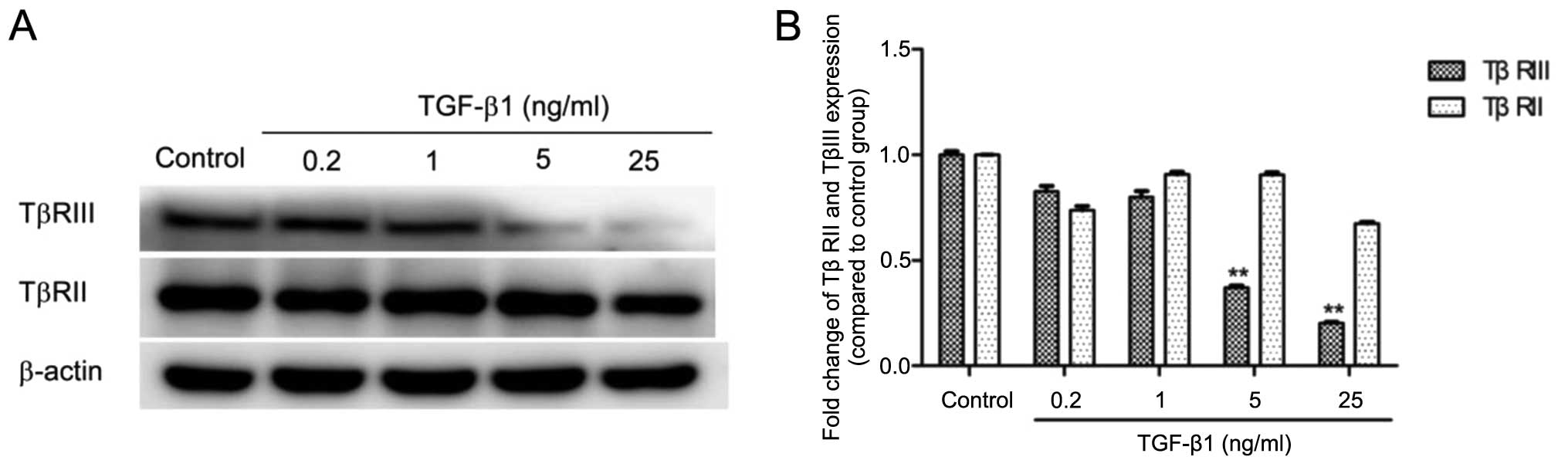
To further characterize the effect of TGF-β1 on the
expression of TβRIII, we treated human SMMC-7721 cells with TGF-β1
at stepwise concentrations (0.2, 1, 5 and 25 ng/ml) and assessed
the protein level of TβRIII using western blotting. Treatment with
TGF-β1 (5 and 25 ng/ml) resulted in a significantly decreased
expression level of the TβRIII protein (Fig. 4). To assess whether the effects of
TGF-β1 on TβRIII were specific, we analyzed the effect of TGF-β1 on
TβRII expression. Western blot results showed no significant
changes in the level of TβRII expression upon TGF-β1 treatment at
various concentrations (Fig. 4).
The results above indicated that it was TβRIII instead of TβRII
that was down-regulated by TGF-β1 in the human SMMC-7721 cells.
Decreased expression of TβRIII elevates
the migratory and invasive abilities of human HCC cells
We detected the gradually decreased expression of
TβRIII in human HCC cell lines along with increasing metastatic
potential. To investigate the roles of TβRIII in regulating the
migratory and invasive abilities of the HCC cells, we silenced the
expression of TβRIII by transfection of siRNA targeting the TβRIII
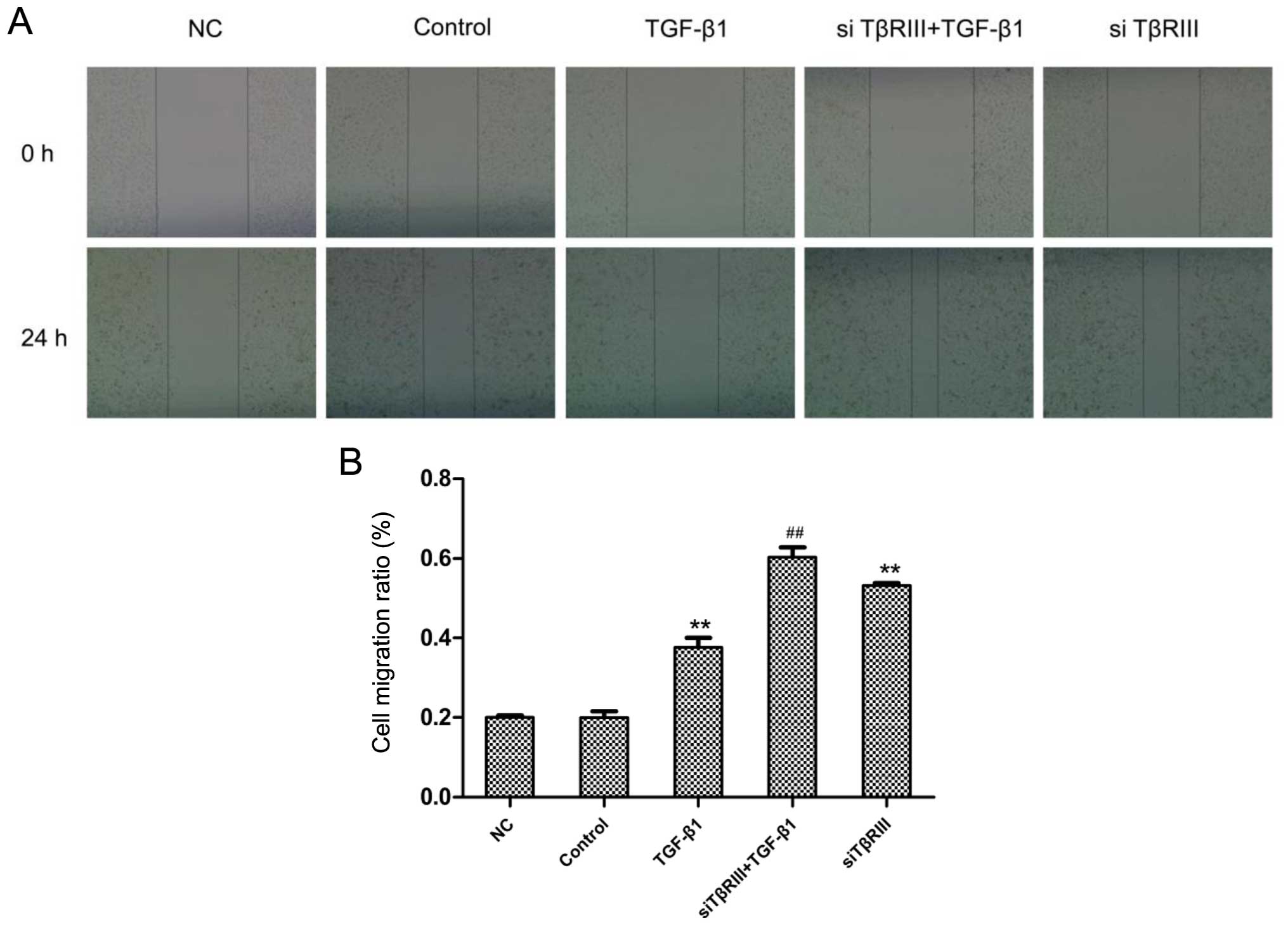
gene. The wound healing and Transwell assays were performed to
detect the migratory and invasive abilities of the SMMC-7721 cells
transfected with the siRNA targeting TβRIII, respectively. The
wound healing assay results revealed that the migratory ability of
the SMMC-7721 cells in the siRNA transfection groups was
significantly increased compared with the corresponding control
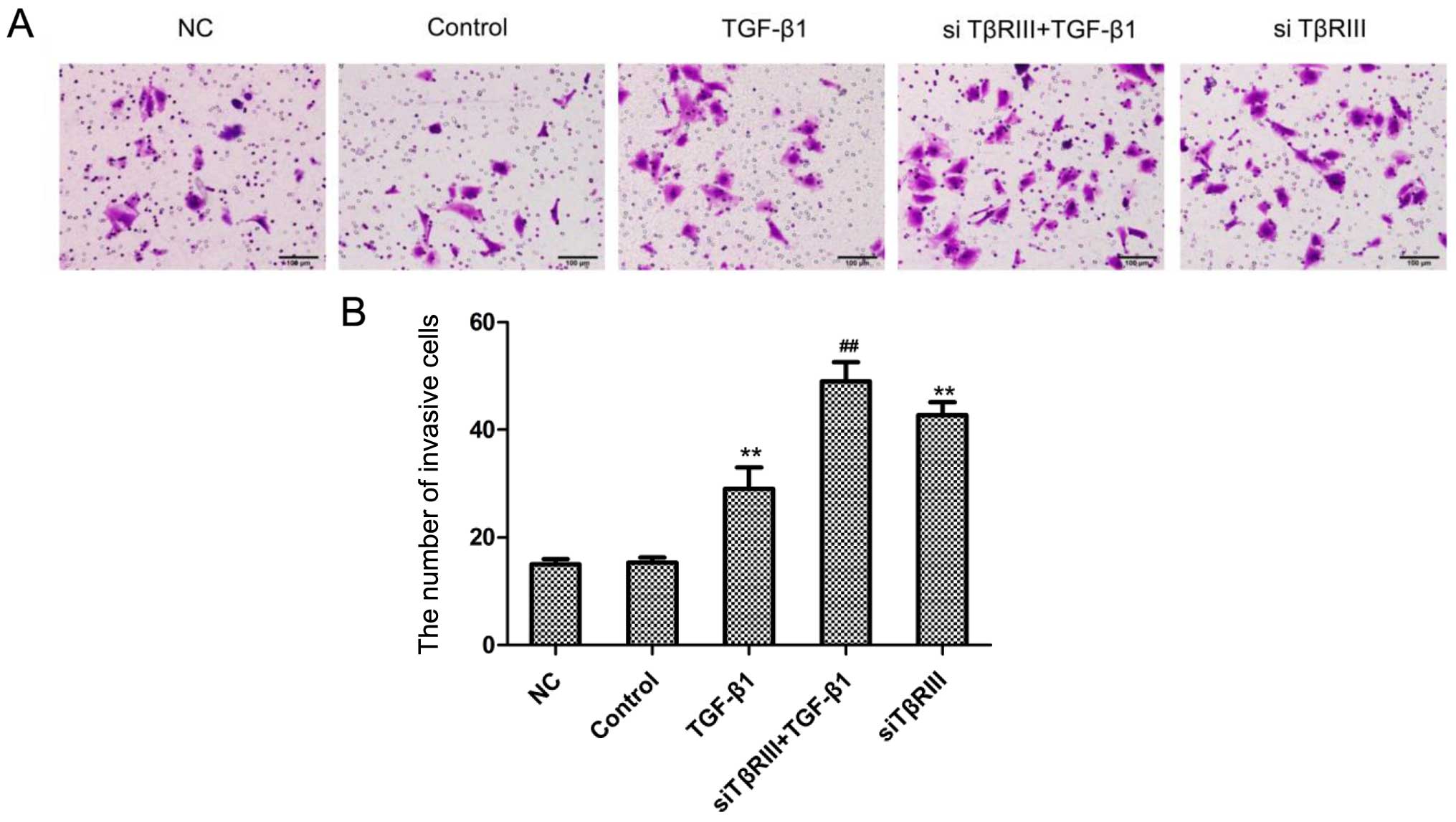
groups (Fig. 5). The Transwell
assay showed a significantly increased invasive ability of the
SMMC-7721 cells in the siRNA transfection groups compared with the
corresponding control groups (Fig.
6). Collectively, these results confirmed that the
downregulation of TβRIII expression promoted HCC cell migration and
invasion in vitro.
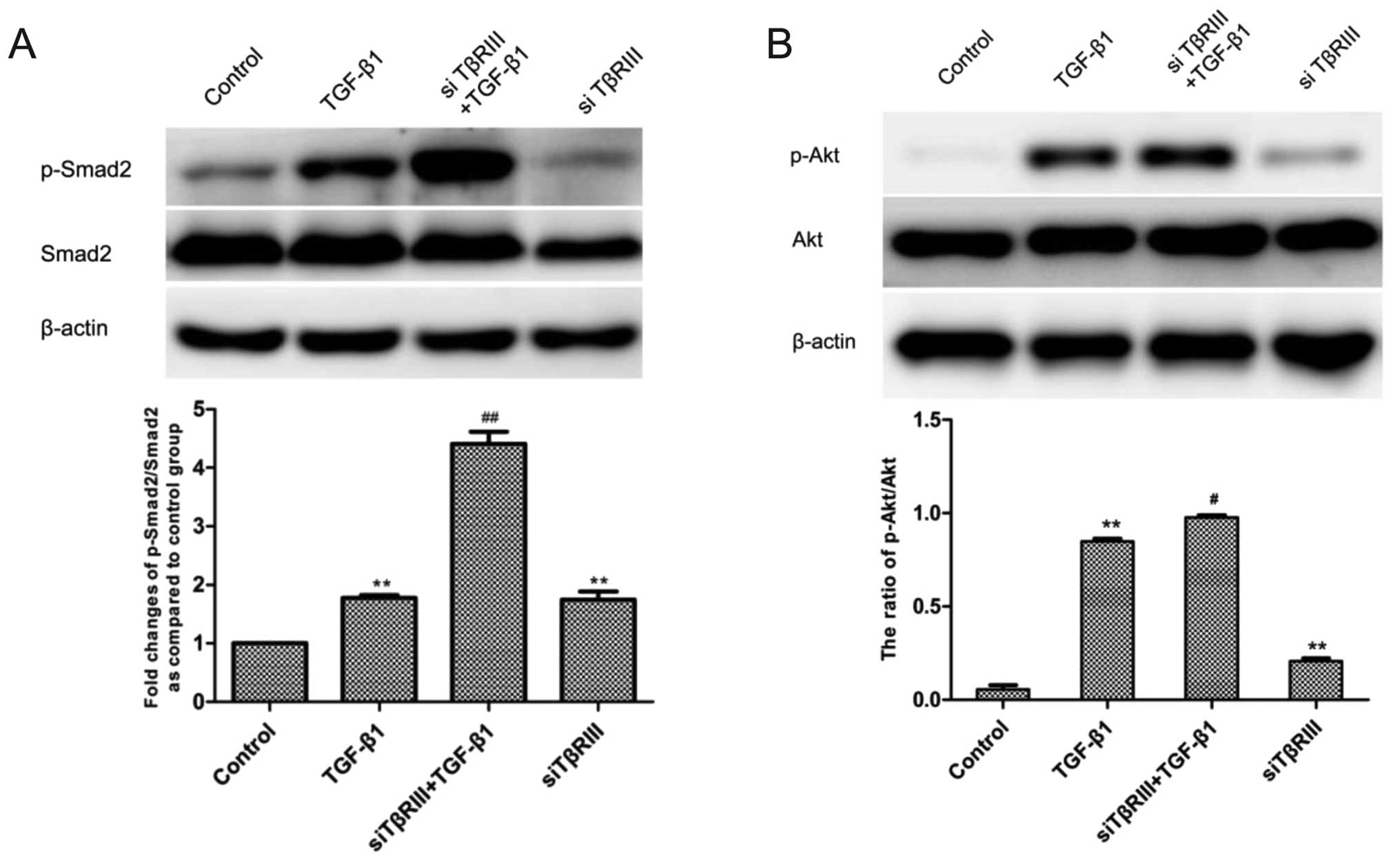
Silencing of TβRIII expression by siRNA
activates the Smad2 and Akt pathways in the SMMC-7721 cells
TGF-β1 was found to promote the migration of HCC
cells and reduced the expression of TβRIII, and furthermore,
silencing of the expression of TβRIII elevated the migration and
invasion abilities of the HCC cells. Thus, we hypothesized that the
inhibitory effects of TβRIII on migration and invasion of HCC cells
may be through regulation of the TGF-β downstream signaling
pathway. Therefore, to further confirm this hypothesis, we examined
Smad2 and Akt activation by western blotting after transfection of
TβRIII siRNA. The results revealed that silenced expression of
TβRIII in the siRNA transfection groups increased the expression
levels of p-Smad2 and p-Akt compared with the corresponding control
groups (Fig. 7).
Discussion
The three highly homologous isoforms of TGF-β in
humans (TGF-β1, TGF-β2 and TGF-β3) share a receptor complex and
signal in similar ways while their expression levels vary depending
on the tissue (22). TGF-β can
induce an epithelial-mesenchymal transition (EMT) of both
epithelial and endothelial cells, which contributes to disease
progression in both cancer and fibrosis (23). EMT promotes cellular migratory and
invasion properties, as cellular migration requires the loss of
cell-cell contacts and acquisition of fibroblastic characteristics.
Several studies have demonstrated the increased expression of
TGF-β1 in various types of cancers and the close association with
tumor metastasis, invasion and the poor prognosis of patients
(24,25). Although the increased expression of
TGF-β1 has been reported to be closely associated with a poor
prognosis and tumor angiogenesis in HCC (26–28),
the mechanism concerning the role of the promotion of tumorigenesis
remains unclear. The research based on the MDA-MB-231 breast cancer
and Ovca420 ovarian cancer cells in vitro revealed that
TGF-β1 reduced the expression of TβRIII at the mRNA and protein
levels in a dose- and time-dependent manner (29). In our previous study, we established
an HCC animal model by intraperitoneally injecting
diethylinitrosamine (DEN) in 14-day-old C57BL/6J mice. The results
revealed significantly increased expression of TGF-β1 and induced
activation of Smad2 signaling in a time-dependent manner after the
injection (30). In the present
study, we observed that TGF-β1 promoted the migratory ability and
induced decreased expression of TβRIII in HCC cells. These results
suggest that the TGF-β1 expression level may be a diagnostic
predictor of HCC progression, and its promotive effect on the
migration of HCC cells may be associated with the low expression of
TβRIII.
The negative regulation of TβRIII expression by
TGF-β1 may provide insight into the tumor-promoting effects of
elevated TGF-β1 and the role of decreased TβRIII expression in HCC
as well. The expression level of TGF-β1 was found to be elevated
and correlated with the migration and invasion in a spectrum of
human cancers (24,25). Nevertheless, the mechanism by which
the high level of TGF-β1 exerts tumor-promoting effects remains
unclear. A recent study found that TβRIII inhibited TGF-β-mediated
signaling through the independent binding to TβRI and TβRII and
competition with TβRI/TβRII complex formation (15). We found decreased expression of
TβRIII concomitant with the elevated level of TGF-β1, as well as
the role of TGF-β1 in promoting metastasis in the HCC cell lines.
Thus, the present study confirmed decreased TβRIII expression as a
possible mechanism for the tumor-promoting role of TGF-β1.
TGF-β binds to the constitutively active type II
TGF-β receptor (TβRII) along with the consequent activation of the
type I TGF-β receptor (TβRI), subsequently forming heteromeric
complexes to induce downstream signaling transduction (9,12).
Although lacking intrinsic enzymatic activity, TβRIII is the most
abundantly expressed TGF-β superfamily receptor and has a complex-
and context-dependent role in regulating TGF-β superfamily
signaling and cancer progression with mechanisms yet to be defined.
Recently, the decreased and even lost expression of TβRIII have
been reported in several types of cancers including breast,
prostate, lung and ovarian cancer (16–19).
Several studies have also demonstrated the aberrant expression of
TβRIII in HCC (27,31). However, the specific expression
condition and roles of TβRIII in HCC development and progression
require further research. The results of the present study showed
that the expression of TβRIII in human HCC tissues was
significantly decreased compared with the level in the matched
normal liver tissues, at the mRNA and protein levels. In our
previous study of an animal model, we also found that the
expression of TβRIII was decreased in DEN-induced HCC mice compared
with normal control mice along with increasing time after DEN
injection (30). These results
suggest that the decreased expression of TβRIII has a negative
regulatory role in HCC progression.
Notably, the regulatory roles of TβRIII in
inhibiting tumor invasion, angiogenesis, and metastasis have also
been defined. The restoration of TβRIII expression in murine
mammary cancer cells and prostate cancer cells inhibited tumor
invasion, angiogenesis, and metastasis both basally and in response
to TGF-β treatment in vivo and in vitro, respectively
(16,17,21,32),
and expression of TβRIII inhibited the motility and invasion of
NSCLC cells and knockdown of TβRIII expression increased its
invasion (18). To further
investigate the roles of TβRIII in the regulation of HCC migration
and invasion, we performed research using human HCC cell lines
in vitro. Consistently, the results showed that gradually
decreased expression of TβRIII was observed in HCC cell lines along
with increasing metastatic potential. Human normal liver L02 cells
showed the highest expression level of TβRIII protein, while the
MHCC97H and HCCLM3 cell lines with high metastatic potential
exhibited a markedly lower expression level than that in HepG2, an
HCC cell line with low metastatic potential. Based on the findings
of decreased TβRIII expression in HCC cell lines with increasing
metastatic potential, we hypothesized that TβRIII may be a negative
regulator of the migration and invasion of HCC. In addition, the
results of siRNA targeting of the TβRIII gene revealed that
knockdown of TβRIII led to the enhanced motility and invasiveness
of human SMC-7721 cells both basically and under TGF-β1
stimulation. Collectively, based on the results in HCC patient
tissues and in a previous HCC mouse model (30), we propose that low expression of
TβRIII is involved in the malignant progression of HCC, and TβRIII
plays a suppressive role in regulating the metastasis and invasion
of HCC.
TβRIII has been previously found to exert inhibitory
effects on the migration and invasion of HCC and on various types
of cancers; however, the related mechanisms remain to be defined.
TβRIII has been shown to have effects on both the Smad and non-Smad
signaling pathways in a ligand-dependent or -independent manner,
including the Smad, NF-κB and p38 signaling pathways (33–35).
In mouse lung fibroblasts, the expression of TβRIII was found to
reduce the phosphorylation of Smad2, Smad3 and Akt (36). Moreover, it has been reported that
restoration of TβRIII in breast cancer inhibited the expression
level of Smad2 or Smad3 in vivo and in vitro
(16). In our previous study in an
HCC mouse model, we detected that the expression of p-Smad2 was
elevated in a time-dependent manner, meanwhile the TGF-β1 level was
increased (30). In the present
study, we detected the expression of p-Smad2 and p-Akt in SMMC-7721
cells after transfection with TβRIII siRNA. The results revealed
that the expression levels of p-Smad2 and p-Akt were elevated in
the siRNA groups compared with the corresponding control groups.
These results suggest that TβRIII suppressed the progression of HCC
by inhibiting the activation of Smad2 and Akt pathways, and
subsequently decreasing the migration and invasion of HCC.
In conclusion, TβRIII displayed decreased expression
with HCC progression in vivo and in vitro, which may
be associated with the higher level of TGF-β1. An elevated level of
TGF-β1 and decreased expression of TβRIII may lead to the
activation of Smad2, and the mechanism between them is yet to be
defined. Furthermore, TβRIII acts as a suppressive factor in
regulating the migration and invasion of HCC, by inhibiting the
Smad2 and Akt pathways. The present study confimed that TβRIII may
be a candidate molecular target in HCC diagnosis and treatment.
Acknowledgments
The present study was financially supported by the
National Natural Science Foundation of China (nos. 81300332 and
81330081), the Specialized Research Fund for the Doctoral Program
of Higher Education of China (no. 20113420120002), and the Natural
Science Foundation of the Higher Education Institutions of Anhui
Province (no. KJ2012A153). The authors acknowledge the assistance
of the staff members of the Institute of Clinical Pharmacology,
Anhui Medical University, in conducting the present study.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buendia MA and Neuveut C: Hepatocellular
carcinoma. Cold Spring Harb Perspect Med. 5:a0214442015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Radeleff BA, Stampfl U, Sommer CM,
Bellemann N, Hoffmann K, Ganten T, Ehehalt R and Kauczor HU:
Transarterial ablation of hepatocellular carcinoma. Status and
developments. Radiologe. 52:44–55. 2012.In German. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lencioni R and Crocetti L: Local-regional
treatment of hepatocellular carcinoma. Radiology. 262:43–58. 2012.
View Article : Google Scholar
|
|
5
|
de Lope CR, Tremosini S, Forner A, Reig M
and Bruix J: Management of HCC. J Hepatol. 56(Suppl 1): S75–S87.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schmierer B and Hill CS: TGFbeta-SMAD
signal transduction: Molecular specificity and functional
flexibility. Nat Rev Mol Cell Biol. 8:970–982. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Akhurst RJ and Hata A: Targeting the TGFβ
signalling pathway in disease. Nat Rev Drug Discov. 11:790–811.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang S, Sun WY, Wu JJ and Wei W: TGF-β
signaling pathway as a pharmacological target in liver diseases.
Pharmacol Res. 85:15–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
López-Casillas F, Cheifetz S, Doody J,
Andres JL, Lane WS and Massagué J: Structure and expression of the
membrane proteoglycan betaglycan, a component of the TGF-beta
receptor system. Cell. 67:785–795. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang XF, Lin HY, Ng-Eaton E, Downward J,
Lodish HF and Weinberg RA: Expression cloning and characterization
of the TGF-beta type III receptor. Cell. 67:797–805. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thatcher JD: The TGF-beta signal
transduction pathway. Sci Signal. 3:tr42010.PubMed/NCBI
|
|
14
|
Li D, Xu D, Lu Z, Dong X and Wang X:
Overexpression of transforming growth factor type III receptor
restores TGF-β1 sensitivity in human tongue squamous cell carcinoma
cells. Biosci Rep. 35:352015. View Article : Google Scholar
|
|
15
|
Tazat K, Hector-Greene M, Blobe GC and
Henis YI: TβRIII independently binds type I and type II TGF-β
receptors to inhibit TGF-β signaling. Mol Biol Cell. 26:3535–3545.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dong M, How T, Kirkbride KC, Gordon KJ,
Lee JD, Hempel N, Kelly P, Moeller BJ, Marks JR and Blobe GC: The
type III TGF-beta receptor suppresses breast cancer progression. J
Clin Invest. 117:206–217. 2007. View
Article : Google Scholar
|
|
17
|
Turley RS, Finger EC, Hempel N, How T,
Fields TA and Blobe GC: The type III transforming growth
factor-beta receptor as a novel tumor suppressor gene in prostate
cancer. Cancer Res. 67:1090–1098. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Finger EC, Turley RS, Dong M, How T,
Fields TA and Blobe GC: TbetaRIII suppresses non-small cell lung
cancer invasiveness and tumorigenicity. Carcinogenesis. 29:528–535.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hempel N, How T, Dong M, Murphy SK, Fields
TA and Blobe GC: Loss of betaglycan expression in ovarian cancer:
Role in motility and invasion. Cancer Res. 67:5231–5238. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen C, Wang XF and Sun L: Expression of
transforming growth factor beta (TGFbeta) type III receptor
restores autocrine TGFbeta1 activity in human breast cancer MCF-7
cells. J Biol Chem. 272:12862–12867. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun L and Chen C: Expression of
transforming growth factor beta type III receptor suppresses
tumorigenicity of human breast cancer MDA-MB-231 cells. J Biol
Chem. 272:25367–25372. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Millan FA, Denhez F, Kondaiah P and
Akhurst RJ: Embryonic gene expression patterns of TGF beta 1, beta
2 and beta 3 suggest different developmental functions in vivo.
Development. 111:131–143. 1991.PubMed/NCBI
|
|
23
|
Derynck R and Akhurst RJ: Differentiation
plasticity regulated by TGF-beta family proteins in development and
disease. Nat Cell Biol. 9:1000–1004. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang AL, Liu SG, Qi WJ, Zhao YF, Li YM,
Lei B, Sheng WJ and Shen H: TGF-β1 protein expression in non-small
cell lung cancers is correlated with prognosis. Asian Pac J Cancer
Prev. 15:8143–8147. 2014. View Article : Google Scholar
|
|
25
|
Chen K, Wei H, Ling S and Yi C: Expression
and significance of transforming growth factor-β1 in epithelial
ovarian cancer and its extracellular matrix. Oncol Lett.
8:2171–2174. 2014.PubMed/NCBI
|
|
26
|
Ji GZ, Wang XH, Miao L, Liu Z, Zhang P,
Zhang FM and Yang JB: Role of transforming growth factor-beta1-smad
signal transduction pathway in patients with hepatocellular
carcinoma. World J Gastroenterol. 12:644–648. 2006.PubMed/NCBI
|
|
27
|
Abou-Shady M, Baer HU, Friess H, Berberat
P, Zimmermann A, Graber H, Gold LI, Korc M and Büchler MW:
Transforming growth factor betas and their signaling receptors in
human hepatocellular carcinoma. Am J Surg. 177:209–215. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsai JF, Chuang LY, Jeng JE, Yang ML,
Chang WY, Hsieh MY, Lin ZY and Tsai JH: Clinical relevance of
transforming growth factor-beta 1 in the urine of patients with
hepatocellular carcinoma. Medicine. 76:213–226. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hempel N, How T, Cooper SJ, Green TR, Dong
M, Copland JA, Wood CG and Blobe GC: Expression of the type III
TGF-beta receptor is negatively regulated by TGF-beta.
Carcinogenesis. 29:905–912. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang S, Sun WY, Gu YJ and Wei W: The
expression of type III TGF-β receptor in diethylnitrosamine-induced
liver cancer model. Acta Univ Medicinalis Anhui. 50:912–916.
2015.In Chinese.
|
|
31
|
Bae HJ, Eun JW, Noh JH, Kim JK, Jung KH,
Xie HJ, Park WS, Lee JY and Nam SW: Down-regulation of transforming
growth factor β receptor type III in hepatocellular carcinoma is
not directly associated with genetic alterations or loss of
heterozygosity. Oncol Rep. 22:475–480. 2009.PubMed/NCBI
|
|
32
|
Sharifi N, Hurt EM, Kawasaki BT and Farrar
WL: TGFBR3 loss and consequences in prostate cancer. Prostate.
67:301–311. 2007. View Article : Google Scholar
|
|
33
|
You HJ, How T and Blobe GC: The type III
transforming growth factor-beta receptor negatively regulates
nuclear factor kappa B signaling through its interaction with
beta-arrestin2. Carcinogenesis. 30:1281–1287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Criswell TL and Arteaga CL: Modulation of
NFkappaB activity and E-cadherin by the type III transforming
growth factor beta receptor regulates cell growth and motility. J
Biol Chem. 282:32491–32500. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Margulis V, Maity T, Zhang XY, Cooper SJ,
Copland JA and Wood CG: Type III transforming growth factor-beta
(TGF-beta) receptor mediates apoptosis in renal cell carcinoma
independent of the canonical TGF-beta signaling pathway. Clin
Cancer Res. 14:5722–5730. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ahn JY, Park S, Yun YS and Song JY:
Inhibition of type III TGF-β receptor aggravates lung fibrotic
process. Biomed Pharmacother. 64:472–476. 2010. View Article : Google Scholar : PubMed/NCBI
|