Introduction
Vulvar cancer (VC) is one of the most common
malignancies of the female reproductive tract after cancer of the
uterine corpus, ovary and cervix (1). The most common histological type is
squamous cell carcinoma (SCC), which represents ~95% of all VC
cases (2). The incidence of vulvar
squamous cell carcinoma (VSCC) has rapidly increased during the
past few years (3,4). Currently, radical surgery is the major
treatment (5), yet postoperative
recurrence and metastasis are important factors that affect the
prognosis of patients with VSCC. Thus, it is important to increase
our understanding of the relevant molecular mechanisms of VSCC to
improve treatment and prevention.
There is growing interest related to the role of the
restructuring of the actin cytoskeleton in cancer progression and
metastasis. Among the many factors regulating actin cytoskeleton
reorganization, cofilin-1 (CFL1) is one of the most important
regulators. Wang et al observed that the protein and mRNA
expression levels of CFL1 were increased in esophageal squamous
cell carcinoma (ESCC), and CFL1 overexpression promoted the
development of ESCC (6). In
addition, a previous study found that the cofilin pathway was
closely associated with the ability of breast cancer cells to
invade and metastasize (7,8). However, CFL1 expression and its role
in VSCC is unclear; therefore, in the present study we aimed to
explore the influence of CFL1 on VSCC.
Materials and methods
Patient specimens
We investigated formalin-fixed and paraffin-embedded
VSCC tissues from 35 patients with VSCC (without preoperative
radiotherapy and chemotherapy) and formalin-fixed and
paraffin-embedded normal vulvar tissues from 20 patients that
underwent plastic surgery of the vulva. The archived tissues were
retrieved from the Department of Gynecological Pathology at China
Medical University, Shenyang, China, from June 2003 to June 2013.
We previously ensured that the tissues were diagnosed by two
experienced pathologists and de-identified prior to use. The
average age of the VSCC patients was 55.9±3.6 years (range, 32–75
years). According to the International Federation of Gynecology and
Obstetrics (FIGO) staging, 8 of the 35 patients presented with VSCC
stage I, 17 with stage II, and 10 with stage III; 19 cases
exhibited high differentiation, 10 cases had moderate
differentiation, and 6 cases had poor differentiation. Ten cases
suffered from lymph node metastasis and 25 cases were without lymph
node metastasis.
Immunohistochemistry
Xylene was used to deparaffinize consecutive
sections of the paraffin-embedded tissues, and different
concentrations of alcohol were used to rehydrate these sections.
For antigen retrieval, the sections were placed into 0.01 M citrate
buffer, then into a pressure cooker and heated to boiling for 2
min. The sections were immersed in 3% H2O2 at
37°C for 20 min to inactivate endogenous peroxidase. Non-specific
binding was prevented by adding 2% goat serum albumin for 30 min.
The sections were immersed in the CFL1 antibody (Proteintech Group,
Chicago, IL, USA) overnight at 4°C; and incubated with anti-mouse
IgG (1:5,000) at 37°C for 15 min and horseradish peroxidase-labeled
streptavidin solution (S-A/HRP) (both from Beijing Zhongshan Golden
Bridge Biotechnology Co., Ltd., Beijing, China) for another 15 min
the following day. After staining with 0.05% diaminobenzidine (DAB;
Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.), the
sections were counterstained using hematoxylin, then dehydrated,
cleared and mounted. For the negative control (NC),
phosphate-buffered saline (PBS) was utilized instead of the primary
antibody. Finally, we used the Metamorph system to measure the
optical density (OD) value.
Western blot analysis
The protein concentrations of 10 VSCC and 10 normal
vulvar tissues were determined via a protein assay kit (Beyotime
Biotechnology, Co., Ltd., Shanghai, China) according to the
manufacturer's protocol. Total denatured proteins were isolated by
10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE), then transferred to Hybond membranes (Amersham, Munich,
Germany) for at least 2 h, and blocked overnight in 5% fat-free
milk. The membranes were immersed in the primary antibody against
CFL1 for at least 2 h. Next, the membranes were rinsed three times
with TBST and incubated with the secondary antibody (anti-mouse IgG
antibodies; 1:5,000; Beijing Zhongshan Golden Bridge Biotechnology
Co., Ltd.) for 2 h. Finally, we analyzed specific bands using
ImageQuant LAS 4000 (Fujifilm, Tokyo, Japan) and ECL Plus detection
reagents (Beyotime Biotechnology, Co., Ltd.).
Cell culture and CFL1 knockdown and
inhibition
Vulvar carcinoma cell line SW962 was purchased from
Guangzhou Jennio Biotech Co., Ltd. (Guangzhou, China) and was grown
in RPMI-1640 medium containing 10% fetal bovine serum (FBS), 100
U/ml penicillin and 100 µg/ml streptomycin under a
humidified atmosphere of 5% CO2 at 37°C. The medium was
replaced every two days, and the cells were subcultured at ~80%
confluency. For CFL1 knockdown, we used CFL1 small interfering RNA
(siRNA) transfectants (Sigma-Aldrich, St. Louis, MO, USA). The
target CFL1 sequences were: 5′-GA AGGAGGAUCUGGUGUUUdTdT-3′ (sense)
and 5′-AAA CACCAGAUCCUCCUUCdTdT-3′ (antisense); the NC siRNA
sequences were: 5′-UUCUCCGAACGUGUCACGUTT-3′ (sense) and
5′-ACGUGACACGUUCGGAGAATT-3′ (anti-sense) (Sigma-Aldrich).
Periplocoside (Shanghai Nature Standard R&D and Biotech Co.,
Ltd., Shanghai, China), a CFL1 inhibitor, was dissolved in dimethyl
sulfoxide (DMSO) to different concentrations as described below
when required for the assays.
Cell proliferation assay
We used the MTT assay to explore the effect of CFL1
siRNA transfectants and periplocoside on SW962 cell growth.
Briefly, 5×103 cells/well were plated into 96-well
plates and treated with CFL1 siRNA and periplocoside at
concentrations of 0, 0.125, 0.25, 0.5, 1.0, 2.0 and 4.0 µM.
At time points 0, 24, 48 and 72 h, MTT was added into each well,
and the cells were incubated for 4 h; then DMSO (both from Beijing
Solarbio Science and Technology Co., Ltd., Beijing, China) was
added to each well and measured at a wavelength of 490 nm.
Flow cytometric apoptosis assay
Flow cytometry was carried out with cells stained
with propidium iodide (PI) and FITC-labeled Annexin V (BD
Biosciences, San Jose, CA, USA) in accordance with the
manufacturer's instructions. Briefly, following the incubation of
the cells treated with CFL1 siRNA and 0, 0.125, 0.25 and 0.5
µM of periplocoside for 48 h, the cells were rinsed twice
with PBS, and then resuspended in 100 µl 1X binding buffer
at a density of 1×105 cells/ml. Next, 5 µl of
FITC Annexin V and 5 µl PI were added to each sample. The
samples were gently vortexed and incubated for 15 min at 25°C in
the dark. Finally, 400 µl of 1X binding buffer was added,
and the cells were analyzed by flow cytometry within 1 h.
Cell cycle analysis
After treatment of the cells with CFL1 siRNA and 0,
0.125, 0.25 and 0.5 µM of periplocoside for 48 h at 37°C in
an atmosphere of 5% CO2, the cells were digested using
0.25% trypsin, harvested and washed twice with PBS at a density of
1×106 cells/ml. A total of 500 µl of ice-cold
ethanol was added to each tube equipped with 1 ml of cell
suspension for at least 2 h and stored at 4°C. Next, the cells were
washed twice with PBS, and PI containing RNase A (BD Biosciences)
was added, and the cells were cultivated at 4°C in the dark for 30
min. The cells were examined using flow cytometry at a wavelength
of 488 nm.
Wound healing assay
To investigate cell migration ability, we used a
wound-healing assay. The cells were plated at 1.0×106
cells/well in 6-well plates. At ~90% confluency, the cells were
wounded with a sterile 200-µl pipette tip, washed twice with
PBS, and grown in FBS-free medium. CFL1 siRNAs and different
concentrations of periplocoside were added to each well. Images of
cells were captured at 0, 24 and 48 h.
Cell invasion assay
To investigate cell invasiveness, we used a cell
invasion assay. A thin layer of Matrigel (Becton-Dickinson Labware,
Bedford, MA, USA) was spread on top of 6.5-mm Transwell chambers
(BD Biosciences). We placed the plates in a 37°C incubator for 4 h
to solidify the Matrigel. Culture medium was then added to the
bottom of the Transwell chambers. The cells resuspended in
serum-free RPMI-1640 were added to the top of the Transwell
chambers. Cells were treated with CFL1 siRNAs or 0, 0.125, 0.25 and
0.5 µM of periploco-side. After incubation for 48 h, the
cells that remained on the top of the chambers were removed by
cotton swabs, and the cells that had invaded the Matrigel and
reached the bottom of the filters were fixed in methyl alcohol. The
cells were then stained with 0.1% crystal violet. The number of
invaded cells was counted using a Olympus fluorescence microscope
(Tokyo, Japan).
Immunofluorescence
After placing coverslips in 6-well plates, we seeded
the cells at 100 cells/well. Subsequently, we discarded the medium
and exposed the cells to CFL1 siRNAs and different concentrations
of periplocoside for 48 h. We washed the cells twice with PBS, and
fixed the cells with 4% formaldehyde for 10 min. Then the cells
were permeabilized with 0.2% Triton X-100 for 10 min at 25°C. Next,
the cells were incubated overnight at 4°C with Alexa
Fluor® 594 phalloidin (Invitrogen Carlsbad, CA, USA) to
visualize the lamellipodia after being washed twice with PBS. The
following day, we added 1 µg/ml
4′,6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich) and incubated
the cells at 37°C for 15 min to stain the cell nuclei. Finally, the
coverslips were observed under a confocal laser microscope (Leica,
Solms, Germany).
Real-time reverse
transcription-polymerase chain reaction (RT-PCR)
According to the manufacturer's protocol, we used
TRIzol (Takara, Dalian, China) to extract RNA from the SW962 cells
transfected with CFL1 siRNAs and exposed to 0, 0.125, 0.25 and 0.5
µM of periplocoside for 48 h. Subsequently, we used a
Reverse Transcription System kit A5001 (Promega, Madison, WI, USA)
to reverse transcribe total RNA to complementary DNA (cDNA). The
PCR primers were from GenBank sequences (Table I). Augmentation of the cDNA was
carried out in 20-µl mixtures according to GoTaq qPCR Master
Mix A6001 (Promega), and the internal control was
glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
 | Table IPrimers for RT-PCR. |
Table I
Primers for RT-PCR.
| Gene | | Primer sequence | Annealing temperature
(°C) | Product size
(bp) | Extension time
(sec) |
|---|
| Cofilin-1 | F |
5′-GCCGCTATGCCCTCTA-3′ | | | |
| R |
5′-CAATTCATGCTTGATCCCT-3′ | 60 | 167 | 34 |
| 18s | F |
5′-ACGGACAGGATTGACAGATT-3′ | | | |
| R |
5′-GGCGTAGGGTAGGCACA-3′ | 60 | 288 | 34 |
Western blot analysis
The protein concentration of cells transfected with
CFL1 siRNAs and periplocoside (0, 0.125, 0.25 and 0.5 µM)
for 48 h was determined by a protein assay kit (Bio-Rad
Laboratories, Hercules, CA, USA) according to the manufacturer's
protocol. Total denatured proteins were separated by 10% SDS-PAGE
and transferred to Hybond membranes for at least 2 h, and then
blocked overnight in 5% fat-free milk. The membranes were incubated
for at least 2 h with antibodies against CFL1, cyclin A1, MMP2 and
MMP9 (Proteintech Group), Bax, Bcl-xL and STAT3 (Boster Biological
Technology, Ltd., Wuhan, China) for immunoblotting. Next, we washed
all of the membranes three times with TBST and incubated them with
secondary antibodies (anti-mouse or anti-rabbit IgG antibodies;
1:5,000; Dako, Carpinteria, CA, USA) for 2 h. Bands were observed
using ImageQuant LAS 4000 (Fujifilm) and ECL Plus detection
reagents (Beyotime Biotechnology, Co., Ltd.).
Statistical analysis
All experiments were repeated three or more times.
The evaluation of statistics was performed using the t-test to
analyze the measurement data and the Mann-Whitney U test to compare
the means of the different groups. A P-value of <0.05 was
considered to indicate a statistically significant result. SPSS
17.0 (SPSS, Inc., Chicago, IL, USA) was used to analyze all
data.
Results
Expression of CLF1 protein in the VSCC
tissues and the relationship of CFL1 expression to
clinicopathological features of VSCC
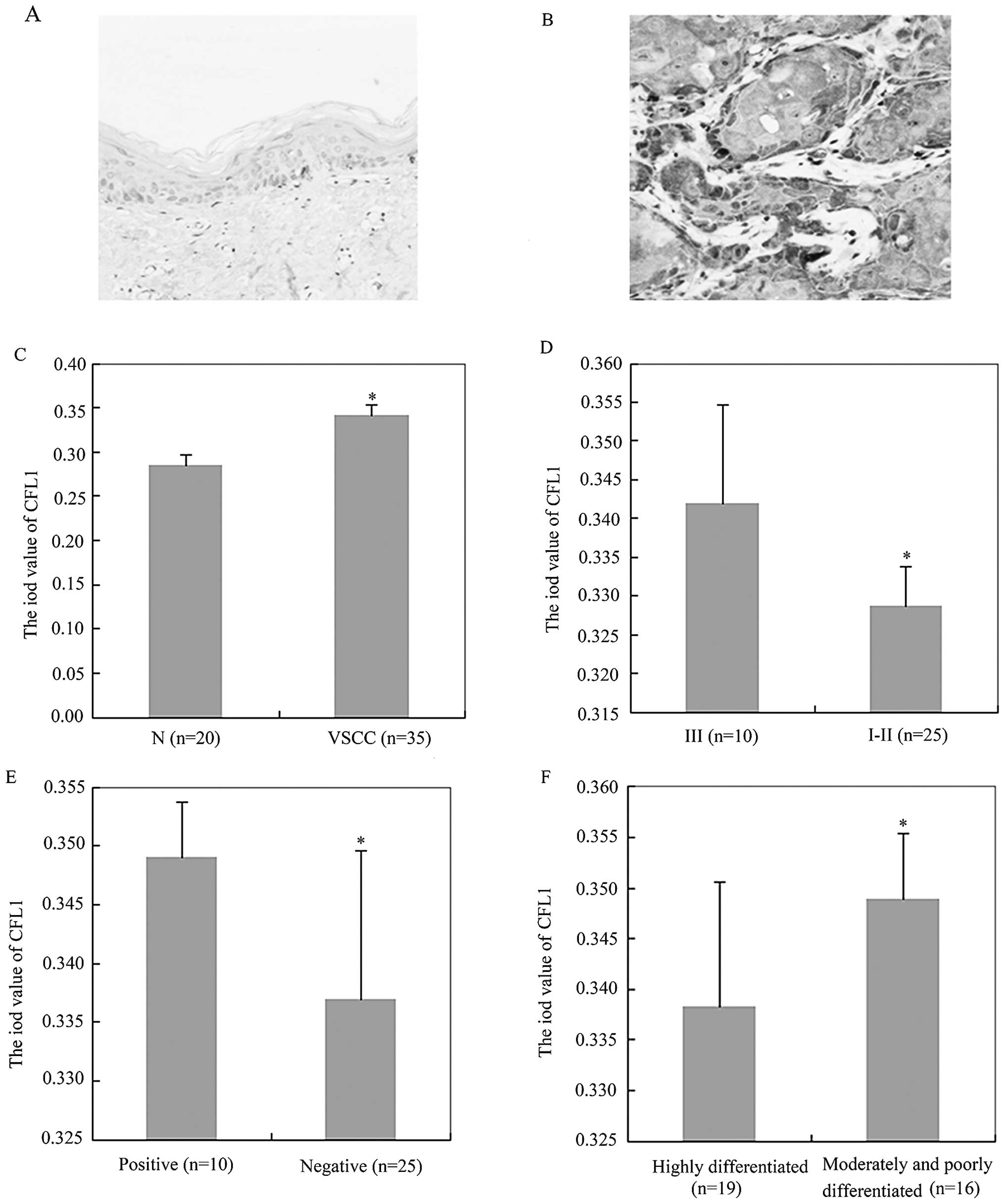
Immunohistochemistry indicated that CFL1 expression
was higher in the VSCC tissues (average OD value, 0.3410±0.0120)
than that in the normal vulvar tissues (average OD value,
0.2849±0.0116) (Fig. 1A–C;
P<0.05). Subsequently, we found that CFL1 expression was
positively associated with FIGO stage (Fig. 1D; P<0.05), lymphatic metastasis
(Fig. 1E; P<0.05) and
dedifferentiation (Fig. 1F;
P<0.05) by immunohistochemistry, while CFL1 expression was not
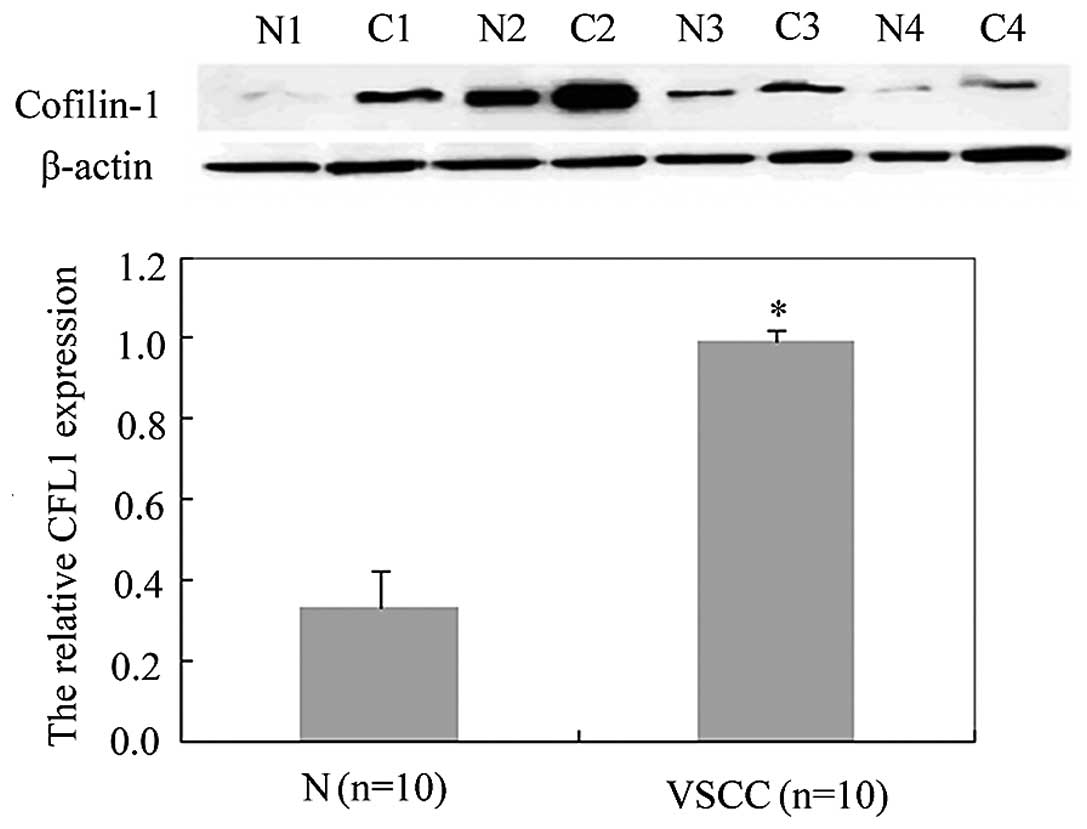
correlated with the age of the VSCC patients (P>0.05; Table II). In addition, western blot
analysis confirmed a similar result; the expression of CFL1 in
normal vulvar cells (relative CFL1 expression, 0.33±0.09) was
decreased compared with the expression in the VSCC cells (relative
CFL1 expression, 0.99±0.03) (Fig.
2; P<0.05).
| Table IICorrelation between cofilin-1
expression and the clinicopathological features of the VSCC
cases. |
Table II
Correlation between cofilin-1
expression and the clinicopathological features of the VSCC
cases.
| Clinical
features | Cases (n) | Mean optical
density | P-value |
|---|
| Age (years) | | | |
| <45 | 8 | 0.3363±0.0086 | 0.195 |
| ≥45 | 27 | 0.3426±0.0132 | |
| LN metastasis | | | |
| Positive | 10 | 0.3490±0.0047 | 0.005 |
| Negative | 25 | 0.3369±0.0127 | |
| FIGO stage | | | |
| III | 10 | 0.3420±0.0127 | 0.001 |
| I–II | 25 | 0.3287±0.0051 | |
|
Differentiation | | | |
| High | 19 | 0.3383±0.0123 | 0.001 |
| Moderate/poor | 16 | 0.3489±0.0065 | |
Effects of CFL1 knockdown on the
phenotype and expression of related molecules in the VSCC
cells
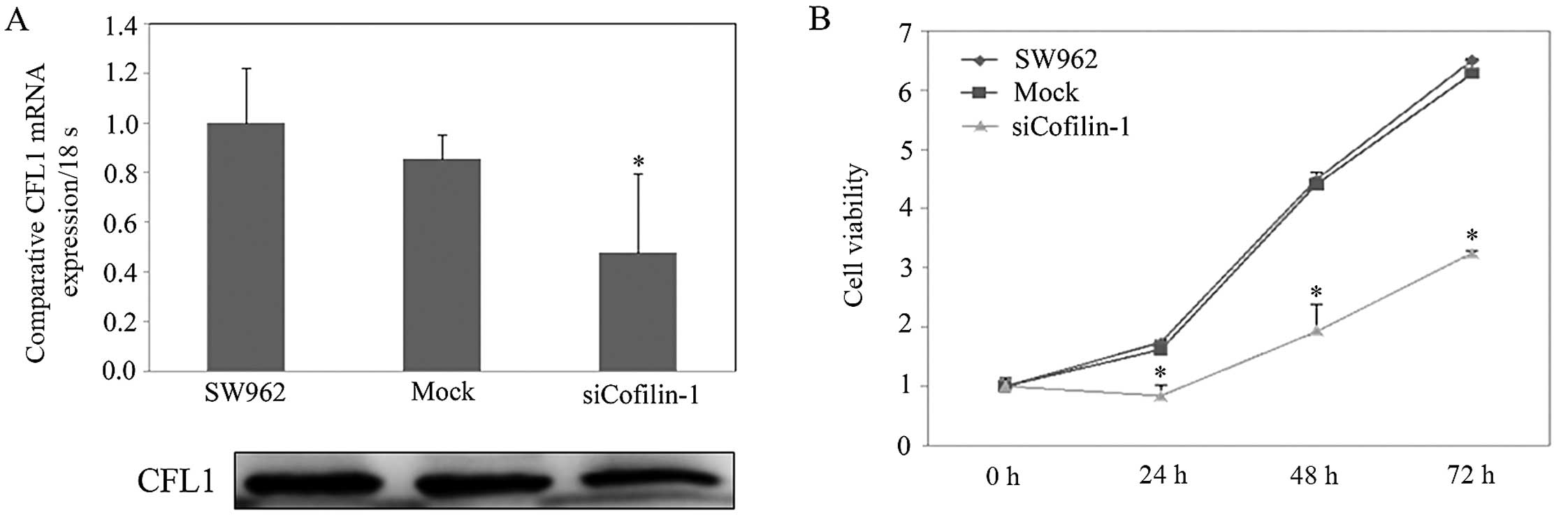
In Fig. 3A, we
showed that levels of CFL1 mRNA and protein expression were highest
in the SW962 cells, and CFL1 expression in the SW962 cells treated
with CFL1 siRNA transfectants was weaker when compared with levels
in the NC or mock-treated cells according to real-time PCR and
western blot assays (P<0.05). According to the MTT assay, we
found that cells transfected with CFL1 siRNA grew significantly
slower than the rate in the NC and mock-transfected cells (Fig. 3B; P<0.05). Cell transfected with
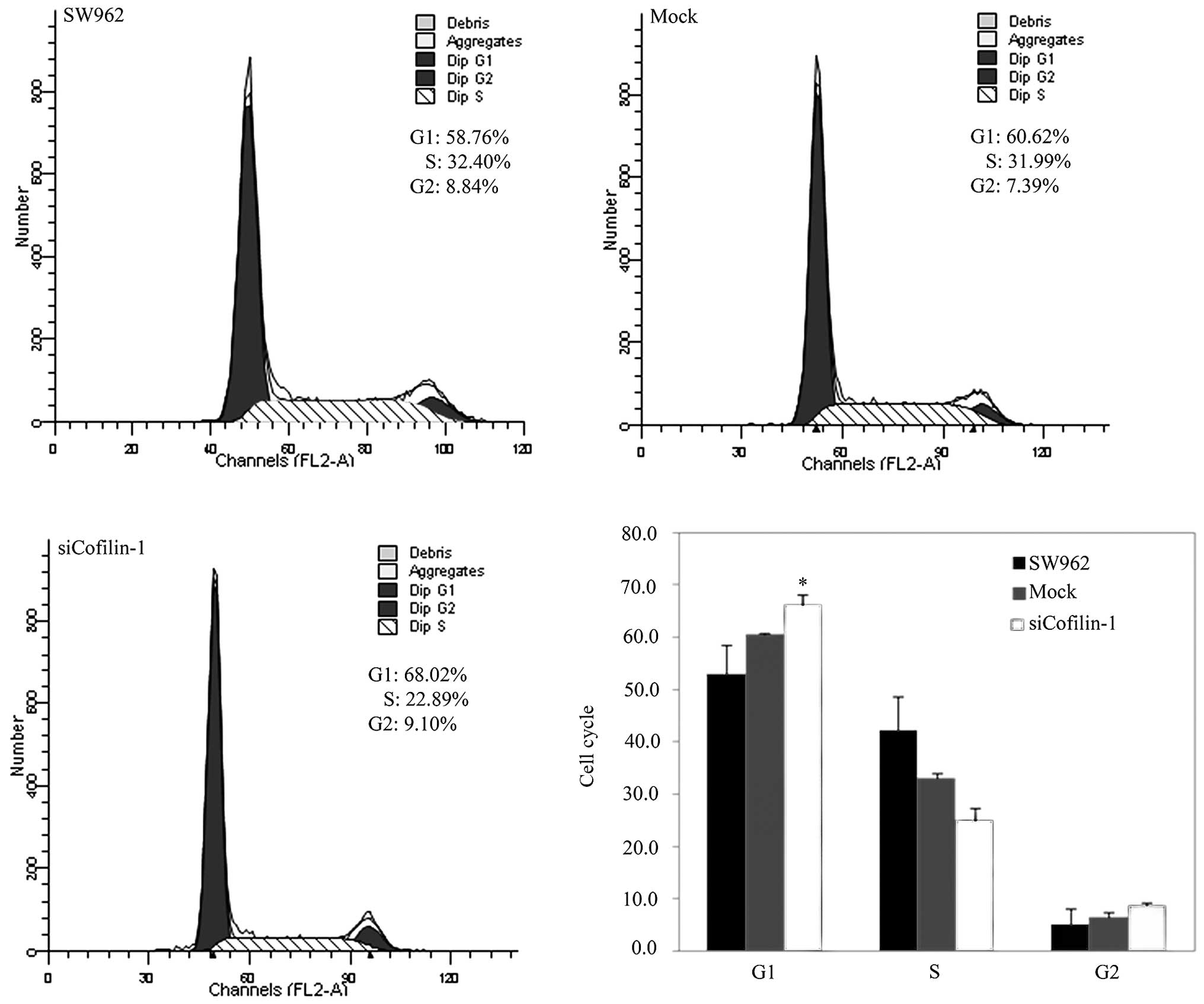
CFL1 siRNA exhibited G1 phase arrest according to PI staining and
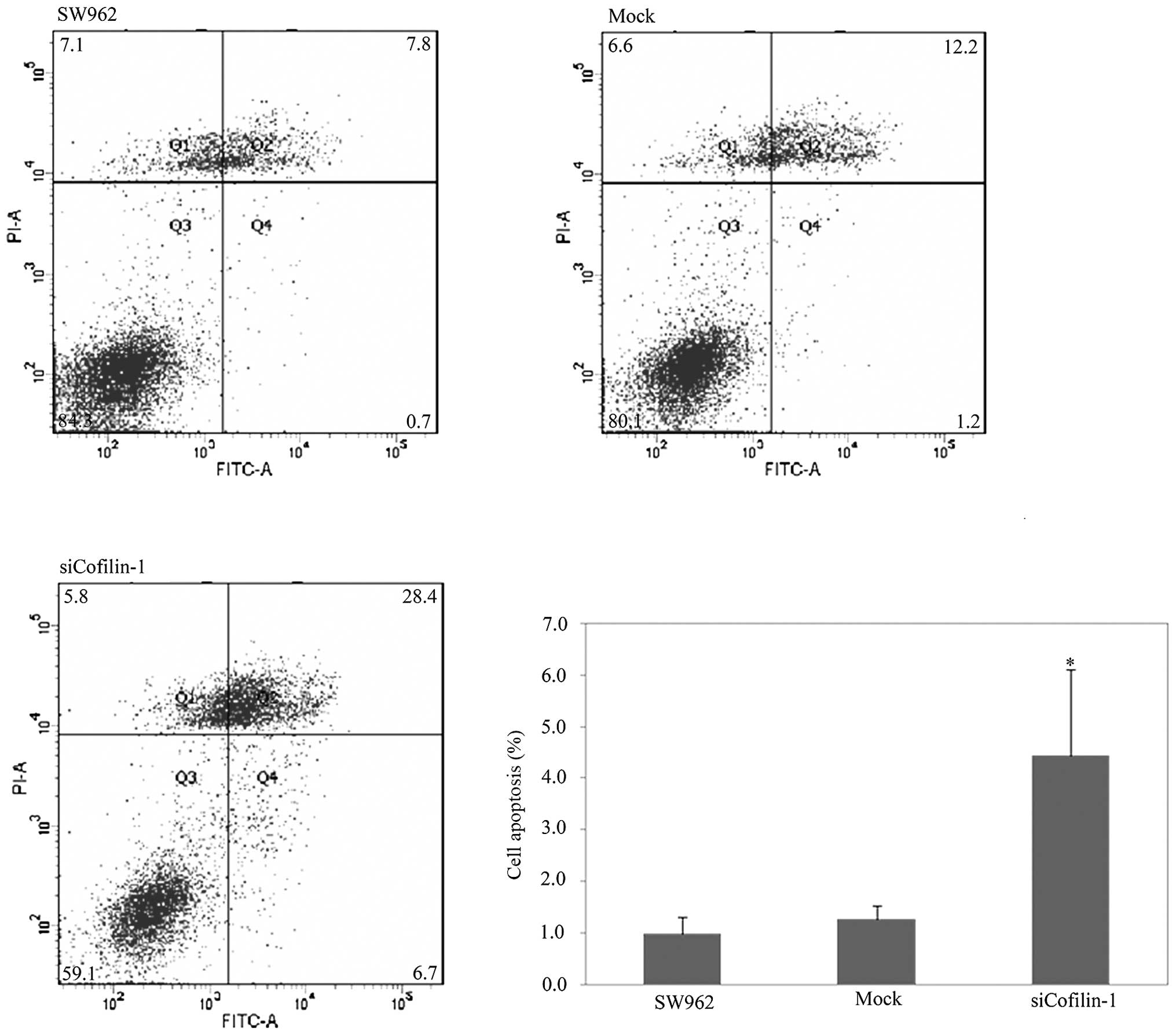
flow cytometry (Fig. 4; P<0.05),
and showed a significantly higher apoptosis rate by Annexin
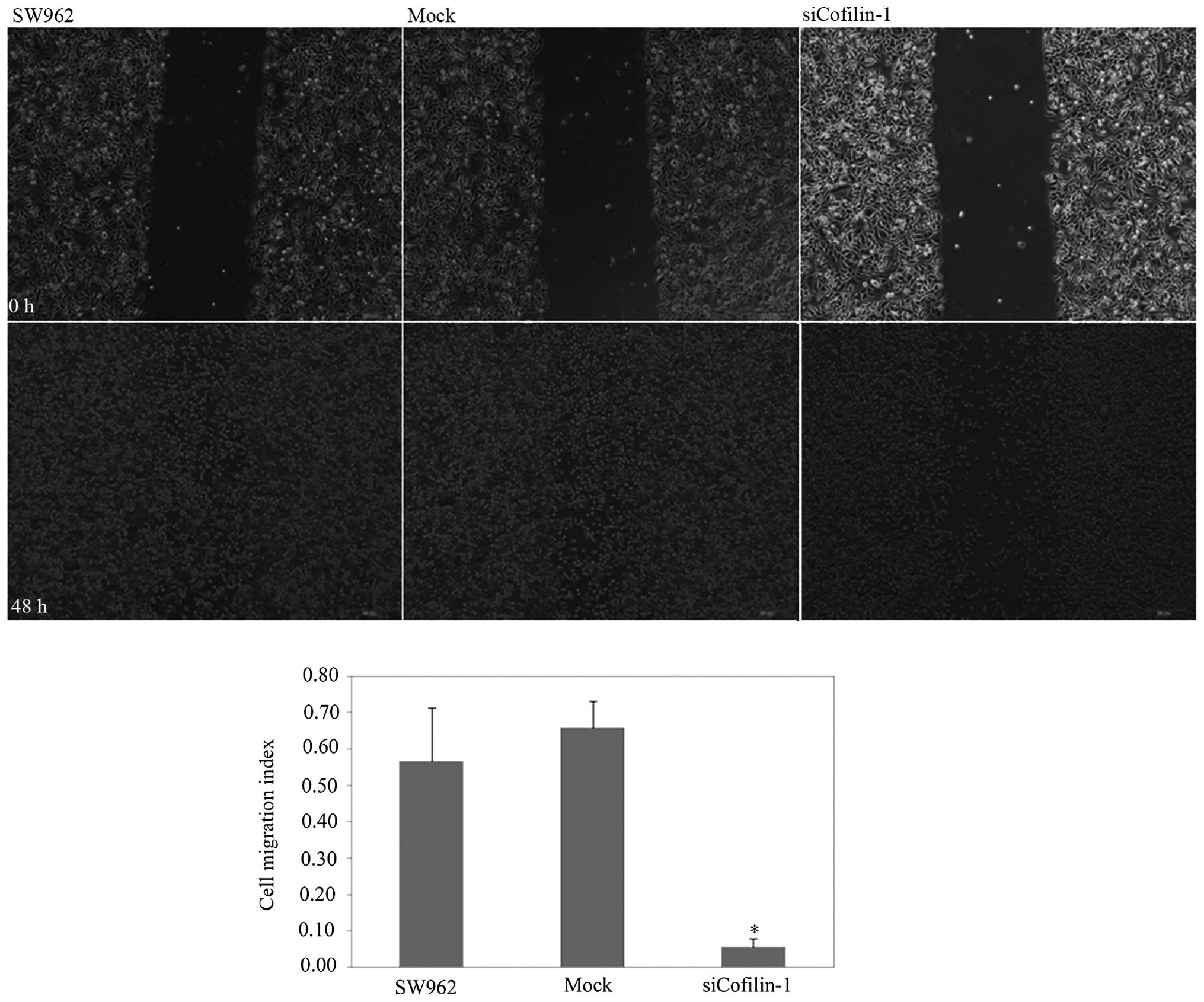
V-fluorescein isothiocyanate (FITC) staining (Fig. 5; P<0.05). Based on the scratch
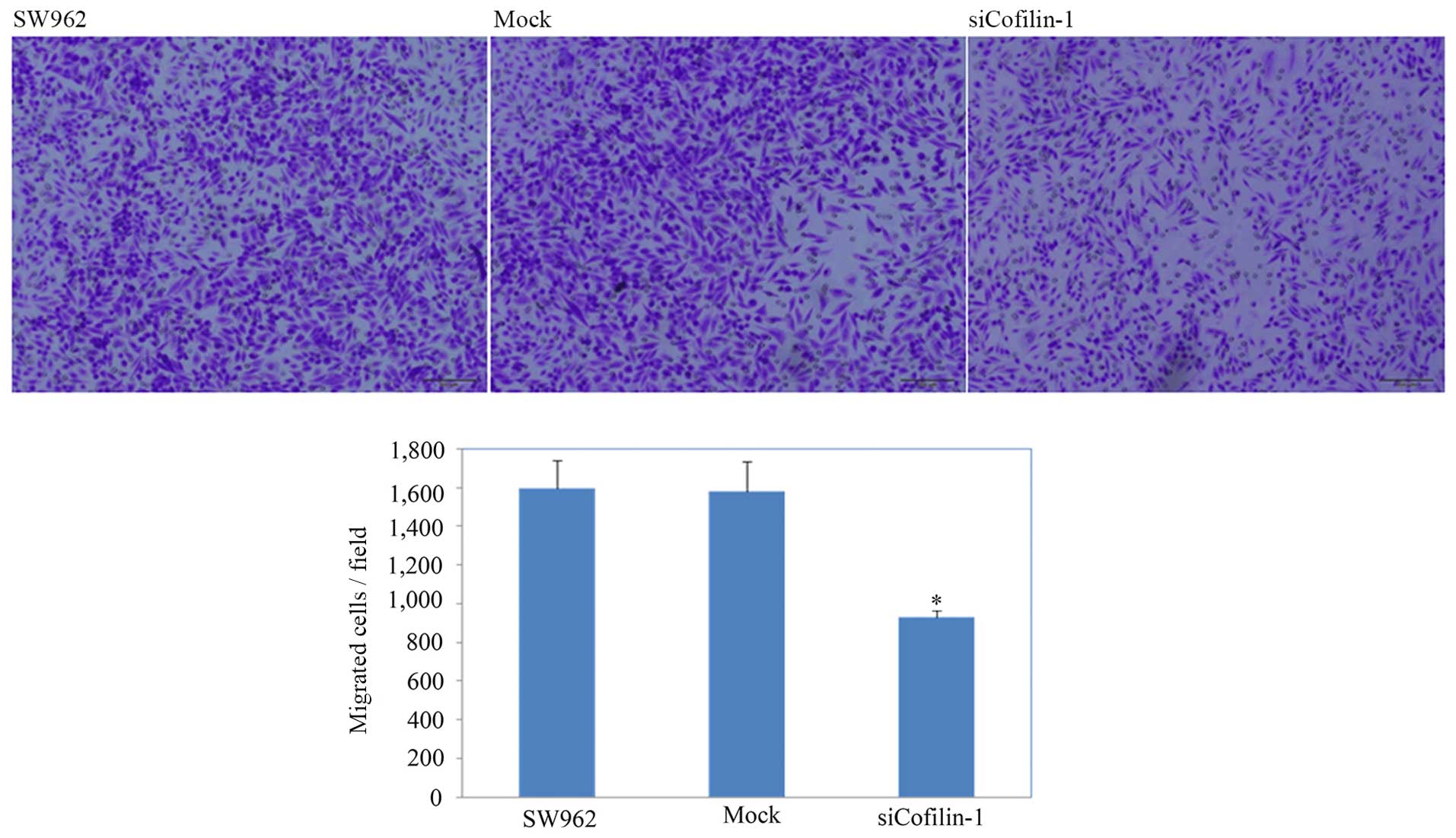
test and the Transwell invasion assay, CFL1 silencing suppressed
cell motility (Fig. 6; P<0.05)
and invasion (Fig. 7; P<0.05)
compared with the motility and invasion of the NC and
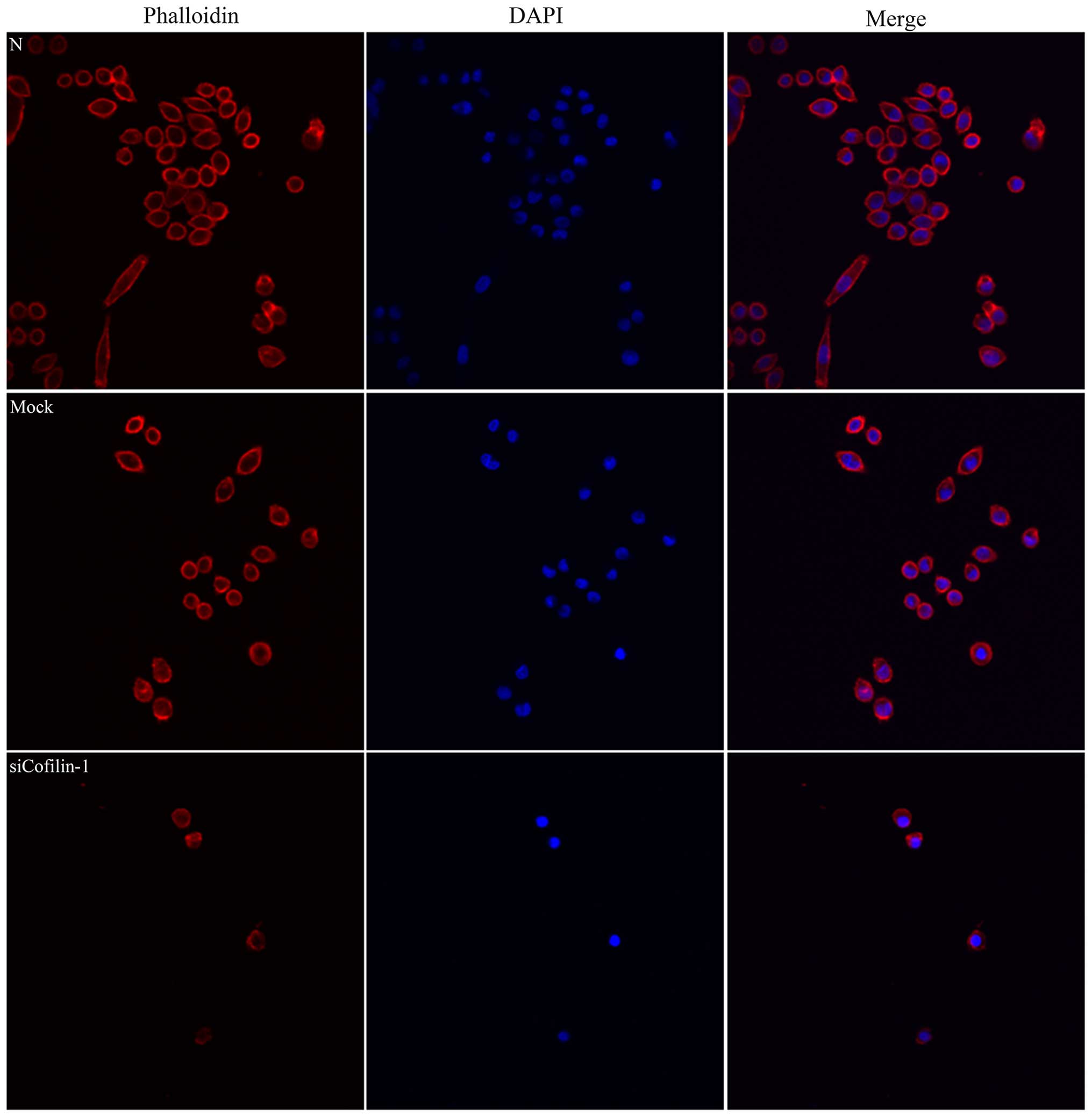
mock-transfected cells. In addition, cells with silenced CFL1 had
inhibited lamellipodium formation as indicated by the F-actin
structure (Fig. 8). In cells
transfected with the CFL1 siRNA transfectants, we found that the
protein expression levels of MMP2, MMP9, cyclin A1, STAT3 and
Bcl-xL were downregulated, while the Bax level was upregulated by
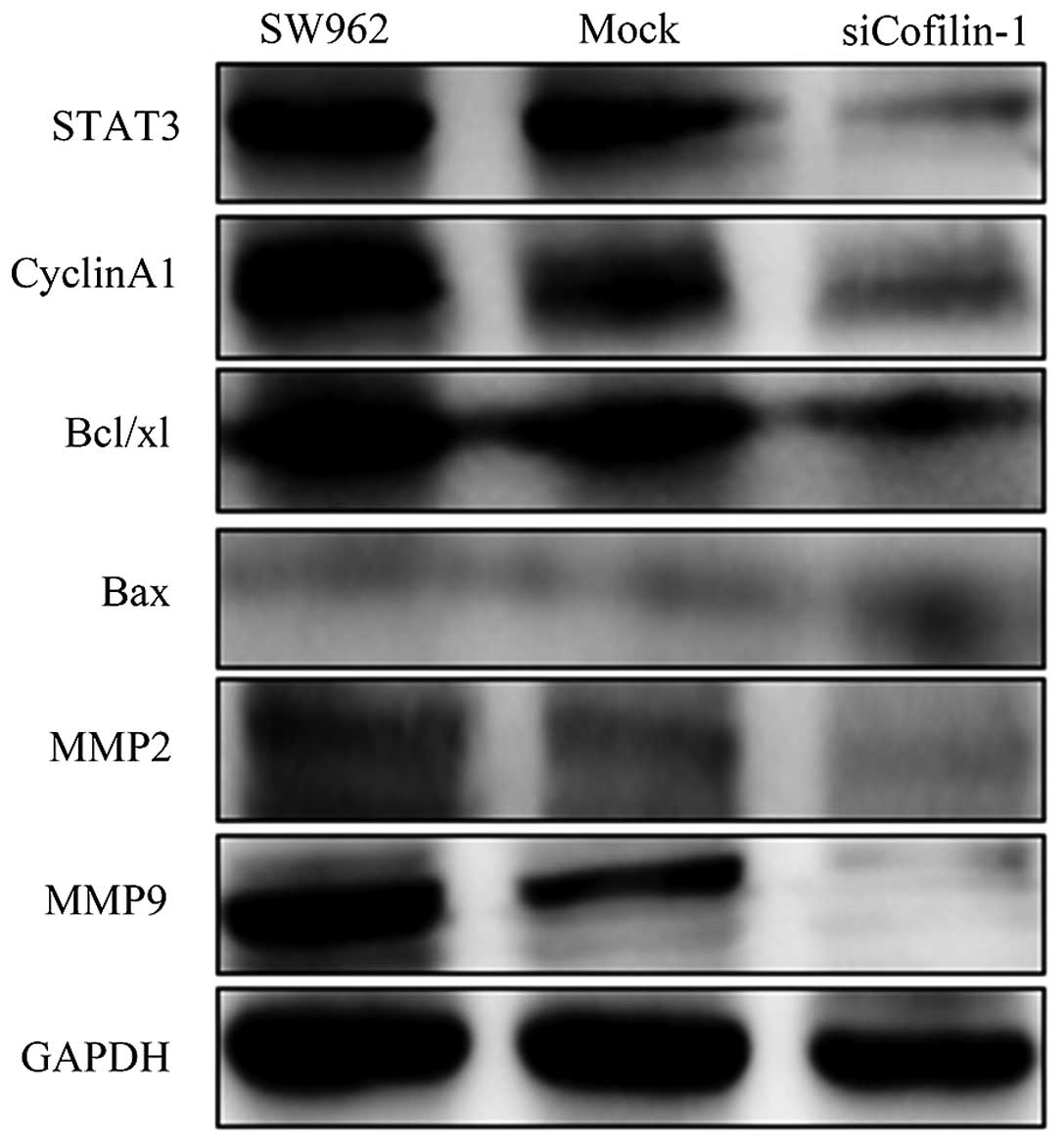
western blot assay (Fig. 9).
Effects of periplocoside on the phenotype
and expression of related molecules in the VSCC cells
We found that periplocoside decreased CFL1
expression in the SW962 cells. Additionally, SW962 cells treated
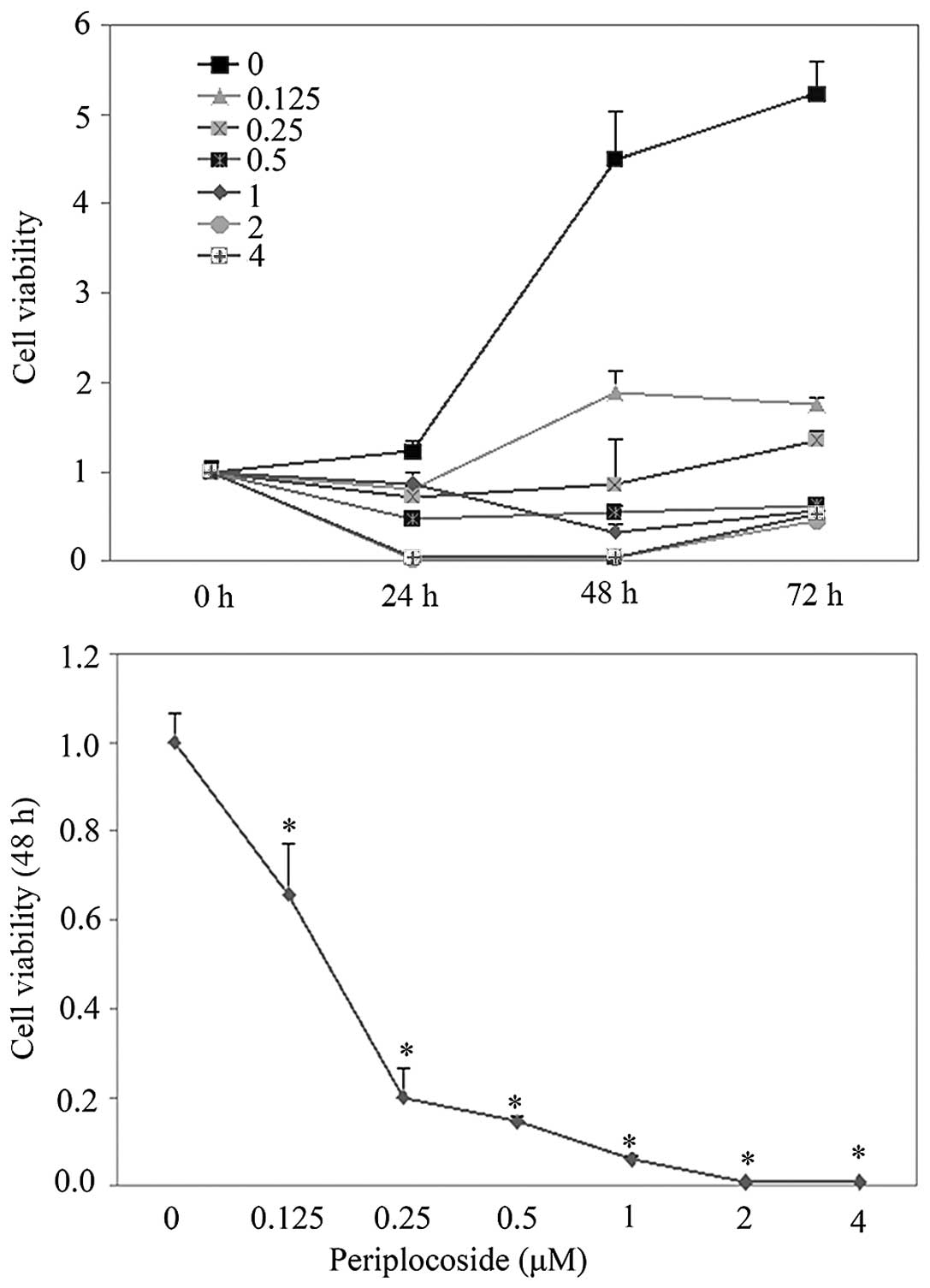
with periplocoside exhibited less proliferative capability compared
with that of the NC in a dose-dependent manner as determined the
MTT assay (Fig. 10; P<0.05).
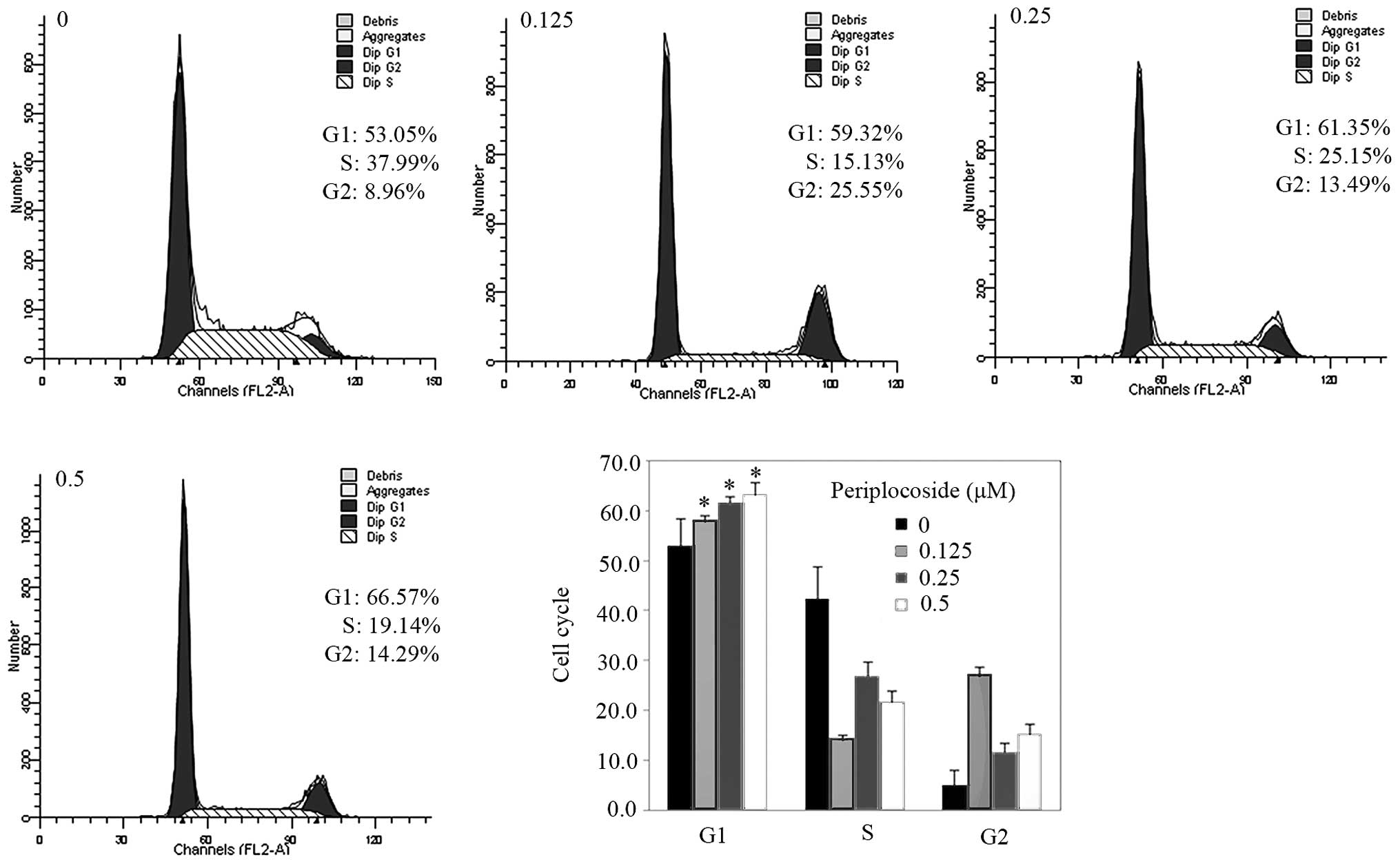
Periplocoside exposure induced G1 phase arrest (Fig. 11; P<0.05) by cell cycle assay,
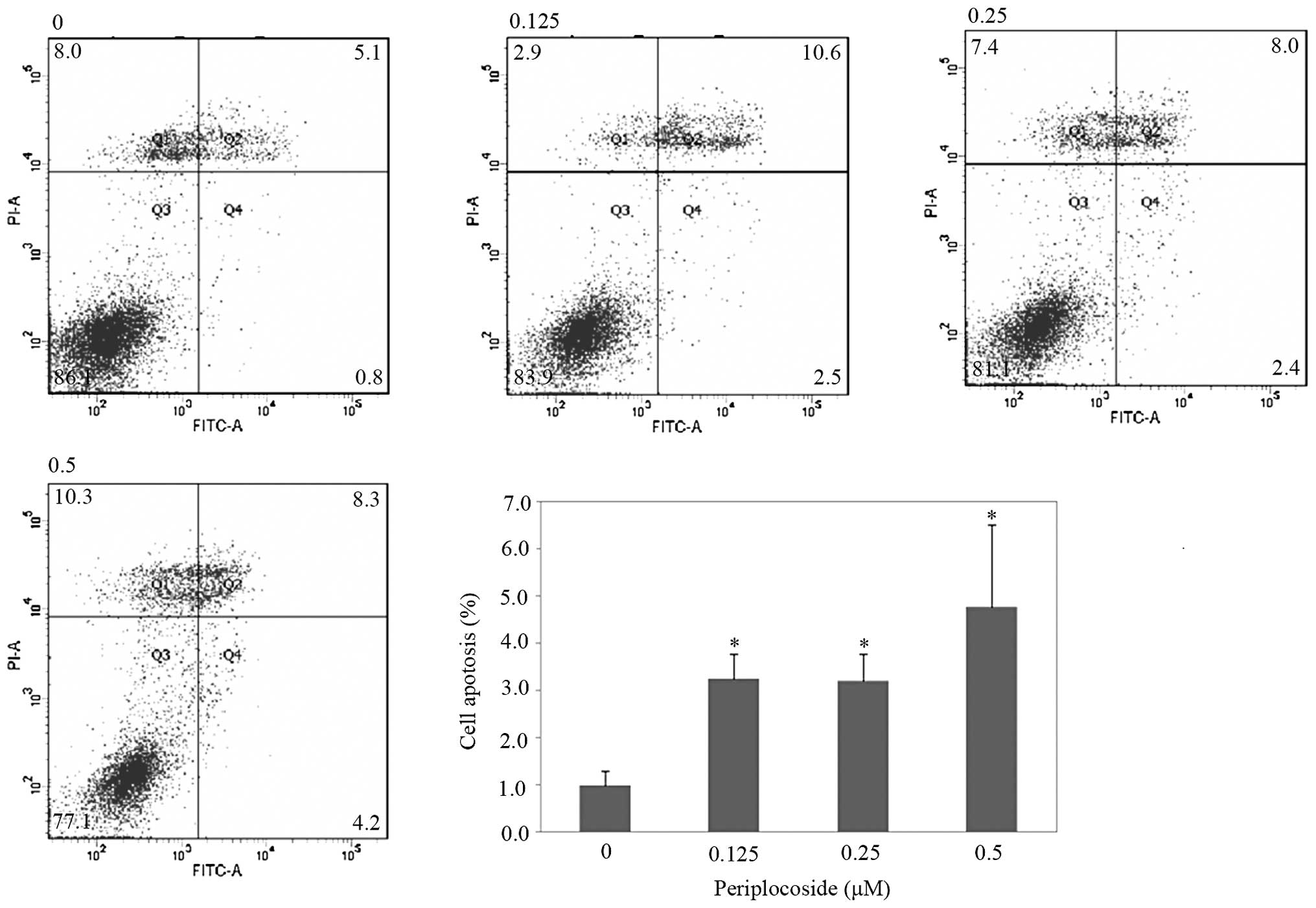
and resulted in the promotion of apoptosis in comparison with NC
(Fig. 12; P<0.05), as shown by
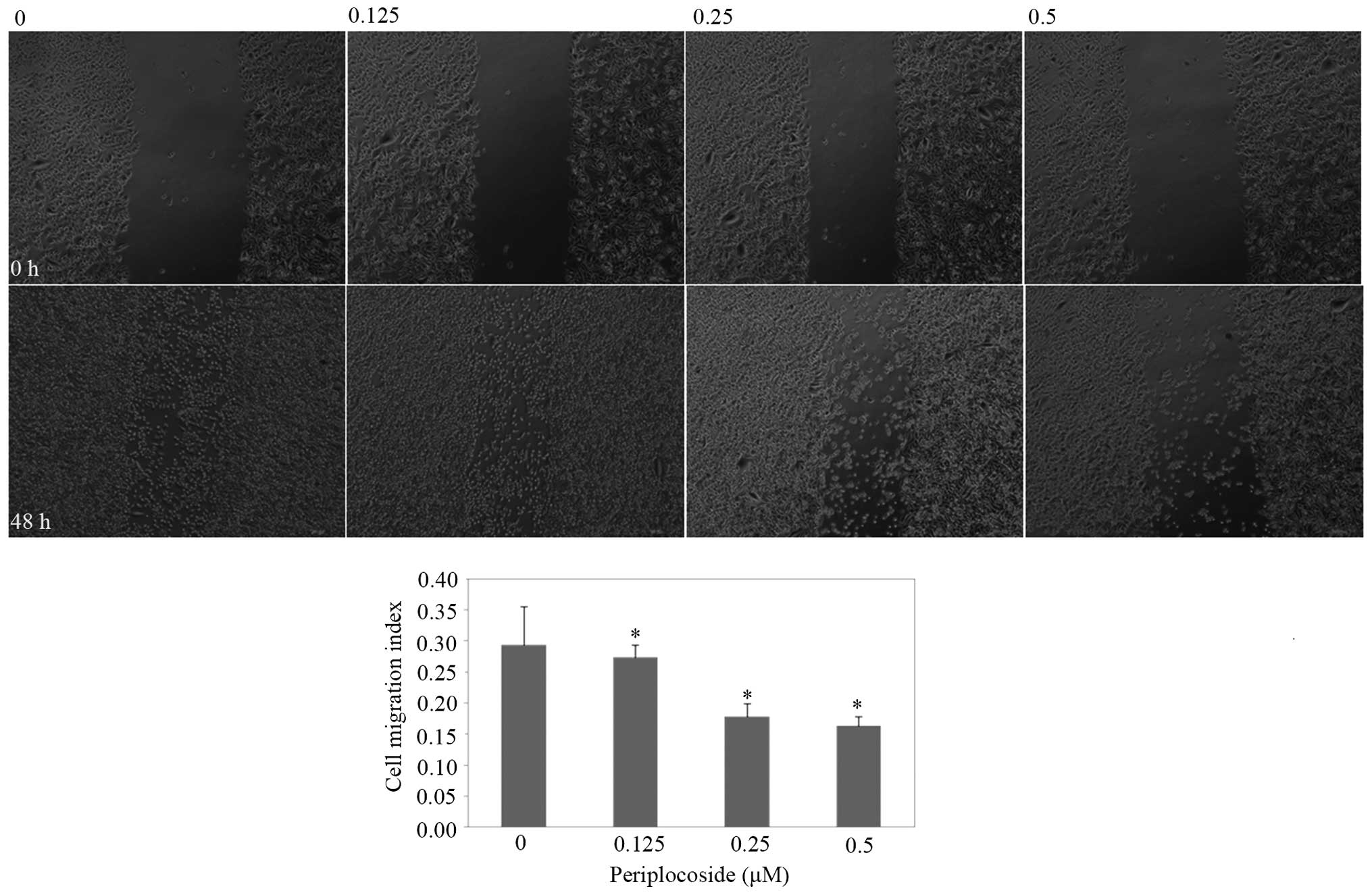
flow cytometric apoptosis analysis. Wound-healing and invasion
assays showed that periplocoside was able to reduce cell migration
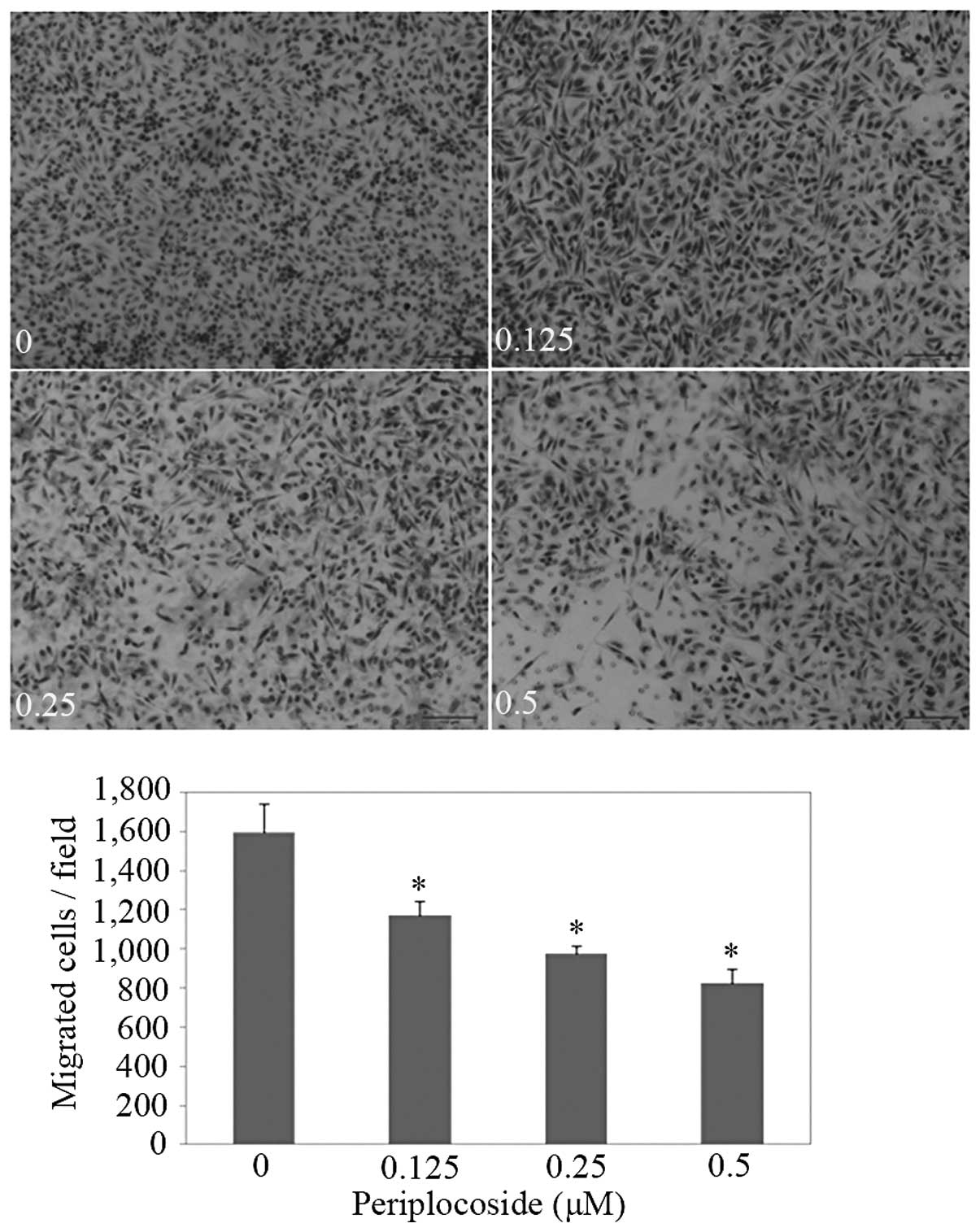
(Fig. 13; P<0.05) and invasion
(Fig. 14; P<0.05) abilities of
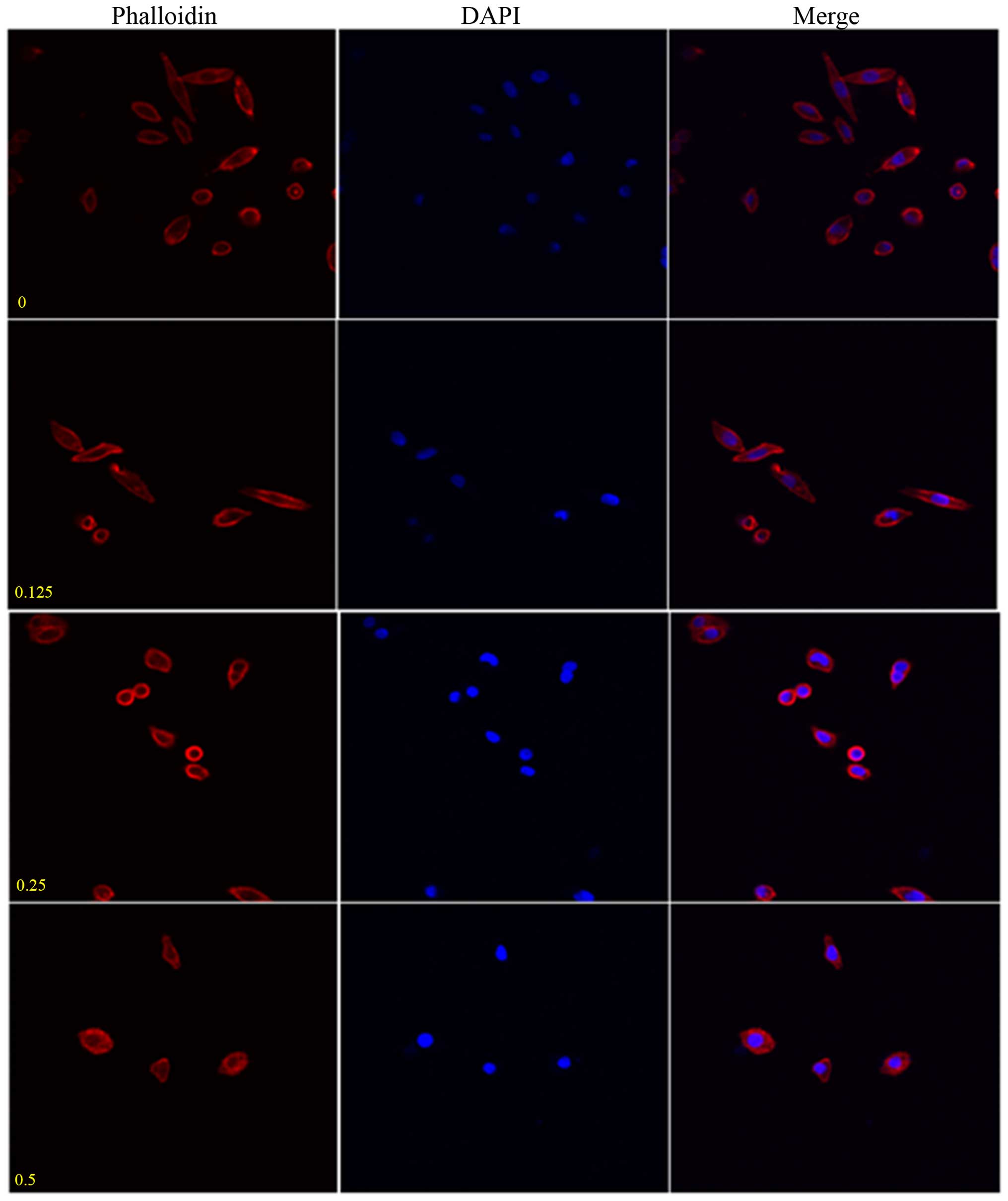
the SW962 cells, and immunofluorescence revealed that periplocoside
destroyed lamellipodium formation (Fig. 15). After treatment with
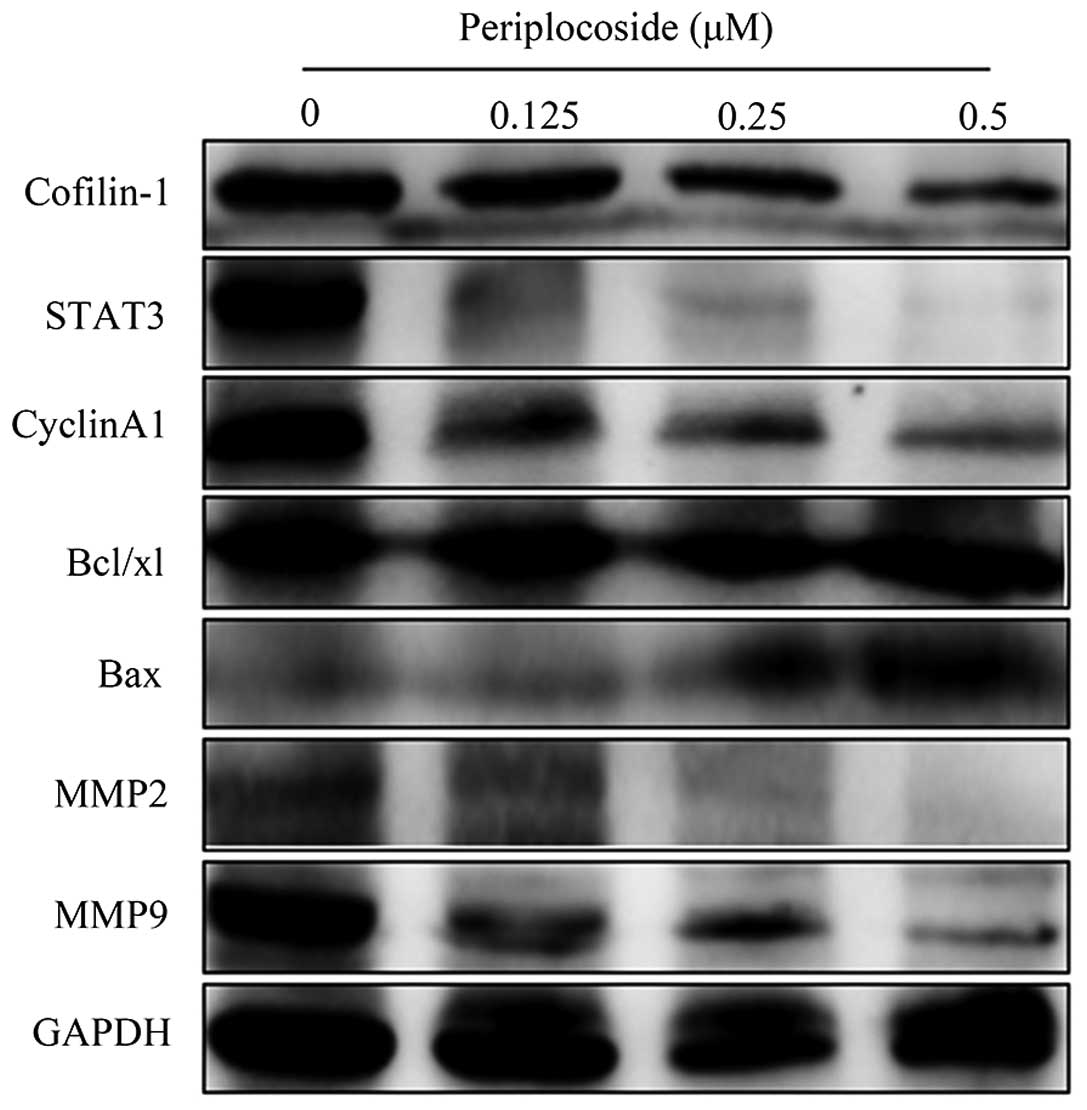
periplocoside, we utilized western blot analysis to assess the
levels of MMP2, MMP9, Bax, cyclin A1, STAT3 and Bcl-xL protein
expression in the SW962 cells and found that all proteins were
downregulated except for the level of Bax which was increased
(Fig. 16).
Discussion
CFL1 is the ubiquity of small proteins, ~19 kDa,
contributing to cytoskeletal dynamics through accelerating the
abscission and depolymerization of actin filaments. CFL1 plays an
important role in cytokinesis, cell motility and morphogenesis
(9). A number of studies have
reported that CFL1 is associated with cancer cell migration and
invasion capability, which are significant features of malignant
tumor cells in various solid tumor tissues (7,9,10). Lu
et al reported that CFL1 was upregulated and significantly
associated with the presence of lymph node metastasis, based on
immunohistochemistry (11). In
addition, a previous study reported that the expression of CFL1 in
ovarian carcinoma tissues was significantly higher than that in
borderline ovarian tumor and benign ovarian tissues by
immunohistochemistry, and that the expression of CFL1 was
positively associated with the degree of ovarian cancer
differentiation (12). In the
present study, we found that CFL1 expression was significantly
upregulated in VSCC cells compared with that in normal vulvar
tissues by immunohistochemistry and western blotting, and was
positively correlated with clinical stage (according to the FIGO
staging system), degree of differentiation and lymphatic
metastasis, which was consistent with previous findings. Taken
together, CFL1 expression is associated with VSCC aggressiveness
and tumor progression.
To determine the concrete biological function of the
CFL1 protein in VSCC, SW962 cells were subjected to CFL1 knockdown
using siRNA transfection. We found that CFL1 silencing induced G1
phase arrest as determined by flow cytometry, promoted apoptosis as
determined by cell apoptosis assay and reduced the formation of
lamellipodia, decreased cell migration by wound healing assay, and
inhibited the ability of cell invasion as determined by Transwell
assay, which indicated that CFL1 silencing may suppress the
aggressive phenotype of VSCC cells. Consistently, Polachini et
al found that knockdown of CFL1 expression by siRNA
significantly inhibited the invasive ability of oral cancer cells
in vitro as determined as determined by Boyden chamber assay
(13). In addition, a previous
study reported that silencing of CFL1 by shRNA delayed tumor
metastasis in mice with lung cancer (14). Additionally, we found that the
knockdown of CFL1 significantly downregulated Bcl-xL expression and
upregulated Bax expression, indicating that CFL1 knockdown promotes
cell apoptosis by mediating apoptosis-related proteins. CFL1
knockdown also suppressed the expression of cyclin A1 that
accelerates the transition of the G1/S phase (15), suggesting that CFL1 knockdown
inhibits the progression of tumor cells by inducing G1 arrest. CFL1
silencing reduced the protein expression of MMP2 and MMP9, which
implies that CFL1 silencing inhibits tumor invasion and metastasis
by suppressing the degradation of the extracellular matrix.
Furthermore, we observed that the silencing of CFL1 inhibited STAT3
expression. STAT3 is a critical member of the signal transducer and
activator of transcription family, and is activated by a wide
variety of cytokines and growth factors. STAT3 inhibition or
ablation has been shown repeatedly to reduce the growth of tumors,
induce apoptosis, and attenuate tumor cell invasion and metastasis
(16,17), which indicates that the antitumor
effects of CFL1 silencing are achieved by inhibiting the STAT3
pathway.
Periplocoside (18),
a traditional Chinese medicine, is derived from Cortex periplocae
and plays an important role in clinical treatment, including
anti-inflammation, enhancing bones and muscles, and nervous system
stimulation. Recently, some researchers have demonstrated that
periplocoside exhibits strong inhibitory effects on the
proliferation of many tumor cell lines in vitro, such as
SW480, SMC-7721, TE13, BGC-823, MCF-7 and PC3 (19,20).
Herein, we observed that periplocoside downregulated CFL1
expression in SW962 cells and inhibited cell growth in a
dose-dependent manner, which could be attributed to the inhibition
of proliferation, the induction of apoptosis, reduced migration and
low invasive ability. Meanwhile, we found that the protein
expression levels of MMP2, MMP9, cyclin A1, Bcl-xL and STAT3 were
reduced, while Bax expression was increased after treatment with
periplocoside, which suggests that periplocoside may inhibit the
expression of these proteins via the suppression of CFL1.
In conclusion, we demonstrated that abnormal CFL1
expression may affect vulvar carcinogenesis and subsequent
progression. CFL1 silencing by CFL1 siRNA significantly inhibited
VSCC cell progression, which suggests that CFL1 may be used as a
potential therapeutic target for vulvar cancer. In addition,
periplocoside, which was utilized in the present study for the
clinical treatment of vulvar cancer, showed strong antitumor
effects by suppressing CFL1 expression.
Acknowledgments
The present study was supported by the Natural
Scientific Foundation of China (no. 30973190).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sideri M, Jones RW, Wilkinson EJ, Preti M,
Heller DS, Scurry J, Haefner H and Neill S: Squamous vulvar
intraepithelial neoplasia: 2004 modified terminology, ISSVD Vulvar
Oncology Subcommittee. J Reprod Med. 50:807–810. 2005.
|
|
3
|
Buttmann-Schweiger N, Klug SJ, Luyten A,
Holleczek B, Heitz F, du Bois A and Kraywinkel K: Incidence
patterns and temporal trends of invasive nonmelanotic vulvar tumors
in Germany 1999–2011. A population-based cancer registry analysis.
PLoS One. 10:e01280732015. View Article : Google Scholar
|
|
4
|
Akhtar-Danesh N, Elit L and Lytwyn A:
Trends in incidence and survival of women with invasive vulvar
cancer in the United States and Canada: A population-based study.
Gynecol Oncol. 134:314–318. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marsden DE and Hacker NF: Contemporary
management of primary carcinoma of the vulva. Surg Clin North Am.
81:799–813. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang WS, Zhong HJ, Xiao DW, Huang X, Liao
LD, Xie ZF, Xu XE, Shen ZY, Xu LY and Li EM: The expression of CFL1
and N-WASP in esophageal squamous cell carcinoma and its
correlation with clinicopathological features. Dis Esophagus.
23:512–521. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang W, Mouneimne G, Sidani M, Wyckoff J,
Chen X, Makris A, Goswami S, Bresnick AR and Condeelis JS: The
activity status of cofilin is directly related to invasion,
intravasation, and metastasis of mammary tumors. J Cell Biol.
173:395–404. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang W, Eddy R and Condeelis J: The
cofilin pathway in breast cancer invasion and metastasis. Nat Rev
Cancer. 7:429–440. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hotulainen P, Paunola E, Vartiainen MK and
Lappalainen P: Actin-depolymerizing factor and cofilin-1 play
overlapping roles in promoting rapid F-actin depolymerization in
mammalian nonmuscle cells. Mol Biol Cell. 16:649–664. 2005.
View Article : Google Scholar :
|
|
10
|
van Rheenen J, Song X, van Roosmalen W,
Cammer M, Chen X, Desmarais V, Yip SC, Backer JM, Eddy RJ and
Condeelis JS: EGF-induced PIP2 hydrolysis releases and
activates cofilin locally in carcinoma cells. J Cell Biol.
179:1247–1259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu LI, Fu NI, Luo XU, Li XY and Li XP:
Overexpression of cofilin 1 in prostate cancer and the
corresponding clinical implications. Oncol Lett. 9:2757–2761.
2015.PubMed/NCBI
|
|
12
|
Zhou J, Wang Y, Fei J and Zhang W:
Expression of cofilin 1 is positively correlated with the
differentiation of human epithelial ovarian cancer. Oncol Lett.
4:1187–1190. 2012.PubMed/NCBI
|
|
13
|
Polachini GM, Sobral LM, Mercante AMC,
Paes-Leme AF, Xavier FC, Henrique T, Guimarães DM, Vidotto A,
Fukuyama EE, Góis-Filho JF, et al: Proteomic approaches identify
members of cofilin pathway involved in oral tumorigenesis. PLoS
One. 7:e505172012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng XC, Gong FM, Zhao YW, Zhou LX, Xie
YW, Liao HL, Lin HJ, Li ZY, Tang MH and Tong AP: Comparative
proteomic approach identifies PKM2 and cofilin-1 as potential
diagnostic, prognostic and therapeutic targets for pulmonary
adenocarcinoma. PLoS One. 6:e273092011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ji P, Agrawal S, Diederichs S, Bäumer N,
Becker A, Cauvet T, Kowski S, Beger C, Welte K, Berdel WE, et al:
Cyclin A1, the alternative A-type cyclin, contributes to G1/S cell
cycle progression in somatic cells. Oncogene. 24:2739–2744. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wake MS and Watson CJ: STAT3 the oncogene
- still eluding therapy? FEBS J. 282:2600–2611. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Katoh D, Nishizuka M, Osada S and Imagawa
M: Fad104, a positive regulator of adipocyte differentiation,
suppresses invasion and metastasis of melanoma cells by inhibition
of STAT3 activity. PLoS One. 10:e01171972015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Du YY, Liu X and Shan BE: Periplocin
extracted from cortex periplocae induces apoptosis of SW480 cells
through inhibiting the Wnt/beta-catenin signaling pathway. Ai
Zheng. 28:456–460. 2009.In Chinese. PubMed/NCBI
|
|
19
|
Zhao L, Shan B, Du Y, Wang M, Liu L and
Ren FZ: Periplocin from Cortex periplocae inhibits cell growth and
down-regulates survivin and c-myc expression in colon cancer in
vitro and in vivo via β-catenin/TCF signaling. Oncol Rep.
24:375–383. 2010.PubMed/NCBI
|
|
20
|
Lu Z, Song Q, Yang J, Zhao X, Zhang X,
Yang P and Kang J: Comparative proteomic analysis of anti-cancer
mechanism by periplocin treatment in lung cancer cells. Cell
Physiol Biochem. 33:859–868. 2014. View Article : Google Scholar : PubMed/NCBI
|