Introduction
Methylation of several conserved lysine residues on
histone H3 and H4 tails by histone methyltransferases (HMTs) plays
a key role in chromatin structure and gene regulation. The role of
HMTs in promoting tumorigenesis and the progression of human
cancers has begun to emerge. Methylation of H3K9 is generally
associated with heterochromatin and gene repression and is a
docking site for HP1 (1–3). Histone lysine methylation is mediated
by HMT, many of which contain a conserved SET [Su(var)3–9, Enhancer
of zeste, Trithorax] domain, such as Suv39h1 (suppressor of
variegation 39h1) and G9a (4,5).
Suv39h1 belongs to a family of pericentromeric proteins and is
responsible for H3K9 trimethylation (6–9). G9a
is the major methylase responsible for mono- and di-methylation of
H3K9 and H3K27 euchromatic regions (10,11),
but it may also be present in heterochromatic regions (12). G9a plays an important role in the
silencing and subsequent de novo DNA methylation of
embryonic and germ-line genes during normal development and is
necessary for the maintenance of the DNA methylation profile of
mammalian cells (13,14). G9a and G9a-like protein (GLP) are
the primary HMTs responsible for histone H3 lysine 9 methylation in
euchromatic DNA (11).
Aberrant histone methylation has not been well
characterized in human disease, especially in gastric carcinoma.
The function of G9a is still not well known. In the present study,
we characterized the G9a, H3K9 me1 and H3K9 me2 expression in
gastric carcinoma tissue, evaluated the correlation between G9a,
H3K9 me2 and the clinicopathological features, in gastric
carcinoma. Furthermore, we applied the siRNA technique to deplete
G9a and measured consequent histone methylation in H3K9 me1, me2
me3 and H3K27 me1 and H3K27 me2 and histone acetylation of H3, cell
proliferation and apoptosis in MGC-803 cells to characterize the
function of G9a in gastric carcinoma.
Materials and methods
Collection of patient samples and
data
There were 175 gastric carcinoma patients who
underwent curative surgical resection at the Surgical Department of
Zhangzhou Affiliated Hospital of Fujian Medical University between
January 2001 and December 2011. None of these patients received
chemotherapy or radiotherapy before the operation. Tumor specimens
were collected in ten percent formalin and then embedded in
paraffin for histopathological analysis. All cases were
independently classified by two experienced pathologists as gastric
carcinoma according to classification of the World Health
Organization. The criteria of the TNM staging system were used to
classify clinical pathological factors and clinical stages of
gastric cancer (defined by the International Union Against Cancer
TNM classification of malignant tumors, seventh Edition, 2009). All
the patients gave signed informed consent. The gastric carcinoma
cases consisted of 133 men and 42 women with a mean age of 63
(range, 41–84) at the time of operation. A summary of patient
characteristics and pathological characteristics is presented in
Table I. Patient samples from
chronic superficial gastritis (n=30), chronic atrophic gastritis
(n=30) and moderate-severe atypical hyperplasia (n=30) were used as
control. The research was approved by the Ethics Committee of
Zhangzhou Affiliated Hospital of Fujian Medical University.
 | Table IClinical significance of G9a and H3K9
me2 expression in gastric carcinoma (n=175). |
Table I
Clinical significance of G9a and H3K9
me2 expression in gastric carcinoma (n=175).
| Group | n | G9a expression
| H3K9 me2
|
|---|
| Positive | Negative |
χ2/t | P-value | Positive | Negative |
χ2/t | P-value |
|---|
| Gender |
| Male | 133 | 113 | 20 | 2.2 | >0.05 | 120 | 13 | 0.06 | >0.05 |
| Female | 42 | 40 | 2 | | | 38 | 4 | | |
| Age (years) |
| <60 | 67 | 54 | 13 | 2.97 | >0.05 | 62 | 5 | 1.01 | >0.05 |
| ≥60 | 109 | 99 | 10 | | | 96 | 13 | | |
| Pathological
type |
| Papillary | 20 | 17 | 3 | 2.46 | >0.05 | 18 | 2 | 2.86 | >0.05 |
| Tubular | 92 | 81 | 11 | | | 85 | 7 | | |
|
Poor-differention | 28 | 24 | 4 | | | 25 | 3 | | |
| Signet-ring | 9 | 8 | 1 | | | 7 | 2 | | |
| Mucinous | 26 | 23 | 3 | | | 23 | 3 | | |
|
Differentiation |
|
Intermediate-high | 103 | 84 | 19 | 6.62 | <0.05 | 88 | 15 | 5.43 | <0.05 |
| Poor | 72 | 69 | 3 | | | 70 | 2 | | |
| Depth
infiltration |
| T1+T2 | 47 | 33 | 14 | 15.25 | <0.05 | 36 | 11 | 11.67 | <0.05 |
| T3+T4 | 128 | 120 | 8 | | | 122 | 6 | | |
| Lymph
metastasis |
| No | 26 | 18 | 8 | 7.35 | <0.05 | 20 | 6 | 4.56 | <0.05 |
| Yes | 149 | 135 | 14 | | | 138 | 11 | | |
| TNM stage |
| I+II | 81 | 66 | 15 | 3.89 | <0.05 | 68 | 13 | 4.33 | <0.05 |
| III+IV | 94 | 87 | 7 | | | 89 | 5 | | |
Tissue microarray construction and
immunohistochemistry
Immunohistochemical studies on G9a, H3K9 me1, and
H3K9 me2 were performed on formalin-fixed, paraffin-embedded tissue
sections obtained from the aforementioned patients with gastric
carcinoma and tissue microarrays. The primary antibodies were used
rabbit monoclonal G9a antibody (1:500 dilutions; Upstate
Biotechnology, Inc., Lake Placid, NY, USA), H3H9me1, H3H9me2
antibodies (1:500 dilutions; Upstate Biotechnology). A
representative tumor section paraffin block (donor block) was
collected from each case, and two tissue cores (2 mm in diameter)
were obtained using a trephine apparatus. Because gastric carcinoma
frequently shows histological heterogeneity, we sampled duplicate
tissue cores from separate areas in individual paraffin-embedded
gastric tumors for better representation of the tumor. Trephinated
paraffin tissue cores were then arranged in a new recipient
paraffin block (tissue array block). Cores containing tumor in
>50% of the area were considered adequate. Immunohistochemical
staining was performed using commercial polyclonal rabbit
anti-histone antibodies to HMT G9a, H3K9 me1 and H3K9 me2. Tissue
array blocks were section at a thickness of 4 µm and mounted on
precoated glass slides. The sections were deparaffinized and
hydrated prior to immunohistochemistry. The immunohistochemical S-P
method was performed according to the manufacturer's protocol.
Tissues positive for all the purchased antibodies were used as
positive controls, sections prepared with phosphate-buffer saline
instead of the primary antibody were used as negative controls.
Positive control exhibited brown color in the nuclei. The staging
was scored by two independent investigators without knowledge of
patient outcomes. The sections were evaluated at low magnification
(×100) to identify areas where G9a, H3K9 me1, H3K9 me2 were evenly
stained. Chevallier's semi-quantity system analysis was used in the
calculation of immunohistochemistry results. Results are presented
as the sum of scores presenting color density and the percentage of
stained cells. According to color density, non-stained cells were
scored as 0, slightly stained 1, intermediate-stained 2, strongly
stained as 3. When the number of positive cells was <25%,
25–50%, or 50–75%, or >75%, the immunoreactivity was scored as
1+, 2+, 3+, 4+, respectively. The two scores presenting color
density and the number of positive cells were added for each slide.
The score of 0 was negative, 1–2 was presented as +, 3–4 as ++, 5–6
as +++, and 7 as ++++. If the sum of the two scores was ≤2, it was
considered negative staining; >2 was considered positive
staining. The scores of each patient group were averaged.
Cell culture analyses
Cell culture studies were done using human gastric
carcinoma MGC-803 cells, which were purchased from the American
Type Culture Collection. Cells were maintained in 10% fetal bovine
serum, and RPMI-1640 containing L-glutamine under 37°C, saturated
humidity and 5% CO2. Cells were subcultured every 3–5
days. The subculture of cells was performed using 0.25% trypsin to
digest the attached cells for 2–3 min.
RNA interference
The approach by transient transfecting using
Lipofectamine™ 2000 was employed to deplete G9a gene in MGC-803
cell lines. G9a siRNA sense: 5′-UUCAGUCUCUACUAUGAUUTT-3′, antisense
5′-AAUCAUAGUAGAGACUGAATT-3′ were synthesized by Shanghai GenePharma
Co., Ltd. (Shanghai, China). Transfection with siRNA was performed
according to the Lipofectamine 2000 manufacturer's protocol
(Invitrogen, Carlsbad, CA, USA). Inducible MGC-803 cells
(1×105 cells/well) were seeded onto 24-well plates
(Corning Life Sciences, Acton, MA, USA) and transiently transfected
with 0, or 40, 80 or 120 nmol/l G9a siRNA. The efficiency of siG9a
and non-specific scrambled siRNA was tested using real-time PCR and
western blot analysis. All experiments were conducted in triplicate
using independent cultures. Both total RNA and protein were
extracted after 24-h transfection.
Real-time PCR for gene expression
Real-time PCR was done using RNA isolated from
MGC-803 cells, using TRIzol reagent (Invitrogen) and purified using
an RNeasy kit (Qiagen). The quantity and quality of RNA samples
were measured by absorbance at 260 and 280 nm. RNA samples with an
A260:A280 ratio 1.8:2.0 were stored at −80°C until use. cDNA
synthesis was performed using an Avian Myeloblastosis virus reverse
transcriptase kit, according to the manufacturer's protocol
(Promega, Madison, WI, USA). β-actin was used as internal control.
Primers used in PCR amplifications were: G9a forward,
5′-UUCAGUCUCUACUAUGAUUTT-3′ and reverse,
5′-AUCAUAGUAGAGACUGAATT-3′; β-actin forward,
5′-CTCCTTAATGTCACGCACGATTTC-3′ and reverse:
5′-CTACAATGAGCTGCGTGTGGC-3′. Amplicon size was 292 base pairs and
517 bp for G9a and β-actin, respectively. PCR reaction conditions
were: 94°C 45 sec, 56°C 1 min, 72°C 80 sec, and was repeated 30
cycles. PCR-amplified products were electrophoresed in 1.0% (w/v)
agarose gels with 1 µg/ml ethidium bromide. Experiments were
repeated twice.
Proliferation assay
After cells were transfected with G9a siRNA, the
proliferative activity was determined by the MTT method (0.5 mg/ml;
Sigma). According to the manufacturer's instructions, cells were
seeded into 96-well tissue culture plates at the density of
1×105/well in triplicate for 0, 24, 48, 72 and 96 h with
different concentrations (0, 40, 60, 80, 100 or 120 nmol/l) of G9a
siRNA. The spectrophotometric absorbance of the samples was
determined by using Ultra Multifunctional Microplate Reader (Tecan
US, Inc., Durham, NC, USA) at 492 and 630 nm for absorbance.
Suppression ratio was also calculated. The experiment was repeated
in triplicate.
Apoptosis detected by TUNEL assay
Cells at logarithmic phase were transfected with
negative control siRNA, and 40, 80 and 120 nmol/l G9a siRNA, and
were seeded in 6-well plates (1×106 cells/well; Costar
Life Sciences) with sterile coverglass placed at the bottom of each
well. After 24-h transfection, apoptotic cells were detected by
TUNEL, according to the manufacturer's protocol (DeadEnd™
Fluorometric TUNEL system; Promega).
Western blot analysis
Total protein lysates and western blot analysis were
performed as previously described (15). Briefly, MGC-803 cells were plated on
culture dishes and transfected by G9a siRNA with 40, 80 and 120
nmol/l for 24 h. Control cells were incubated in the medium with
Na2CO3 using the same time-points. After
incubation, total proteins were prepared from each culture
condition with a lysis buffer containing freshly prepared protease
inhibitors. Protein concentration was then measured using BCA
protein assay (Pierce, Rockford, IL, USA). Cell extracts were
subjected to 12% SDS-PAGE and electrophoretically transferred to
nitrocellulose membrane. Membranes were incubated with monoclonal
anti-mono-methyl-histone H3K9, anti-di-methyl-histone H3K9,
anti-tri-methyl-histone H3K9, anti-mono-methyl-histone H3K27,
anti-dimethyl-histone H3K27, anti-acetyl-histone H3, and G9a
(Upstate Biotechnology), BCL-2, Bax, pro-caspase-3, (Santa Cruz
Biotechnology, Santa Cruz, CA, USA). After being washed with TBS,
membrane was incubated with secondary antibody conjugated with
peroxidase. The signal was then detected using the chemiluminescent
detection system (Pierce) and analysed by a color image analysis
system (AlphaDigiDoc; Alpha Innotech, Corp., St. San Leandro, CA,
USA).
Statistical analysis
The Student's t-test for mean and standard
deviation, or the Mann-Whitney test for median and range were
performed for comparisons between the groups in regards to
continuous data. Comparisons among groups regarding categorical
data were analyzed by performing the Chi-square test. All data were
analyzed using the SPSS, version 16.0 computer program (SPSS, Inc.,
Chicago, IL, USA), and the significance of these differences was
defined as P<0.05.
Results
Overexpression of G9a, H3K9 me2 in
gastric carcinoma tissues
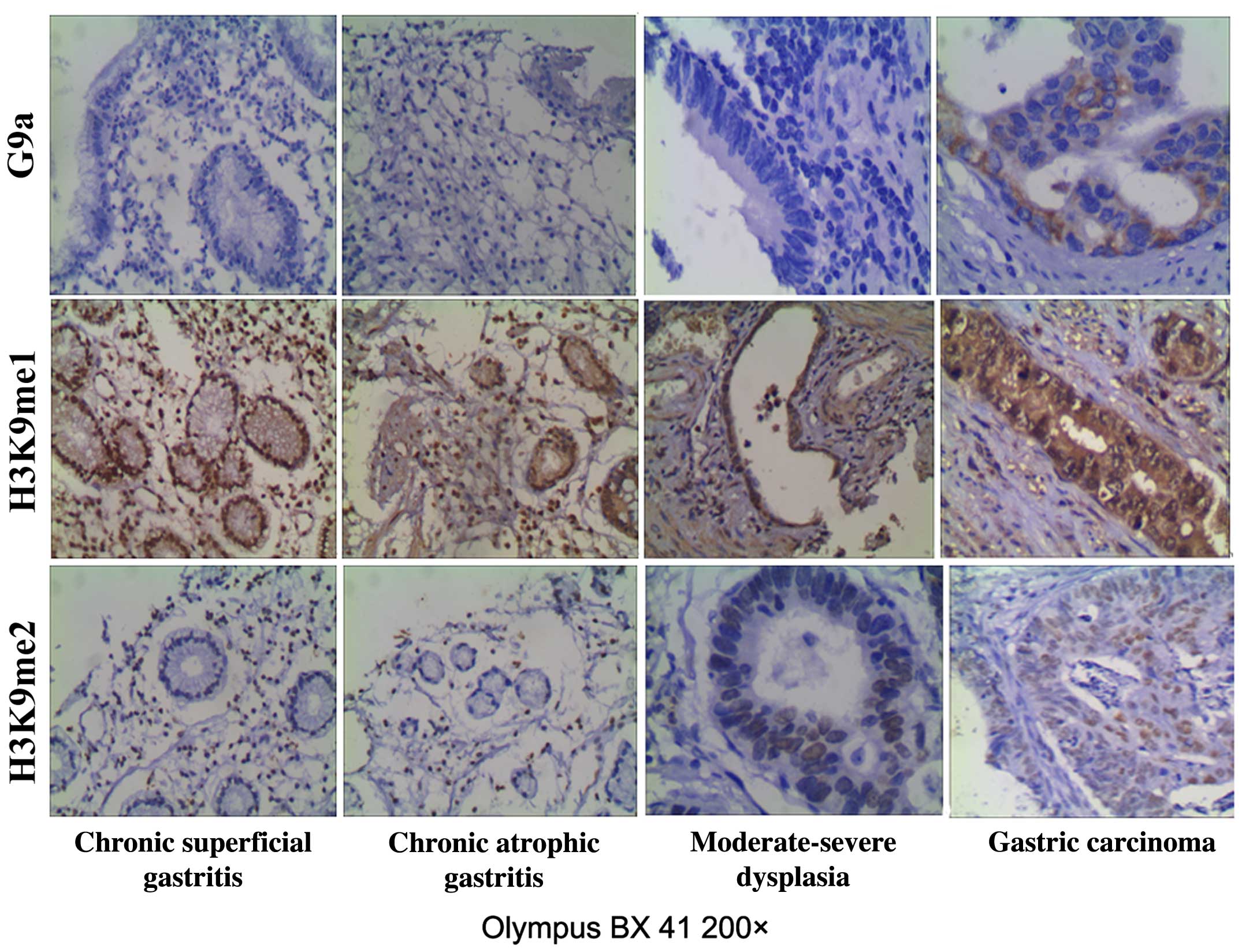
We found overexpression of G9a, and H3K9 me2 in
gastric carcinoma tissues. The expression of G9a in gastric
carcinoma was 87.43%, clearly higher than that in chronic
superficial gastritis (33.33%, P<0.05), chronic atrophic
gastritis (43.33%, P<0.05) and moderate-severe dysplasia
(46.67%, P<0.05). The histone H3K9 me2 in gastric carcinoma was
90.28%, higher than that in chronic superficial gastritis (36.67%,
P<0.05), chronic atrophic gastritis (40.00%, P<0.05), and
moderate-severe dysplasia (56.67%, P<0.05). The expression of
histone H3K9 me1 in gastric carcinoma was 50.86%, similar to
chronic superficial gastritis (53.33%, P>0.05), chronic atrophic
gastritis (50.00%, P>0.05) and moderate-severe dysplasia
(56.67%, P>0.05) (Fig. 1).
G9a and H3K9 me2 expression in gastric
carcinoma is associated with poor prognosis
The association of G9a, and H3K9 me2 expression with
clinical outcomes was analyzed separately. We divided the level of
staining scores into two groups: negative (score ≤2) and positive
(score >2). Among these specimens, High G9a and H3K9 me2
expression positively correlated with differentiation (P<0.05),
depth of infiltration (P<0.05), lymph node metastasis
(P<0.05), and TNM stage (P<0.05). They were not correlated
with age (P>0.05), gender (P>0.05) or pathological type
(P>0.05), respectively. The relationships between the levels of
G9a, and H3K9 me2 expression and the clinicopathplogical
characteristics of gastric cancer are summarized in Table I.
G9a-specific siRNA inhibition of G9a in
MGC-803 cells
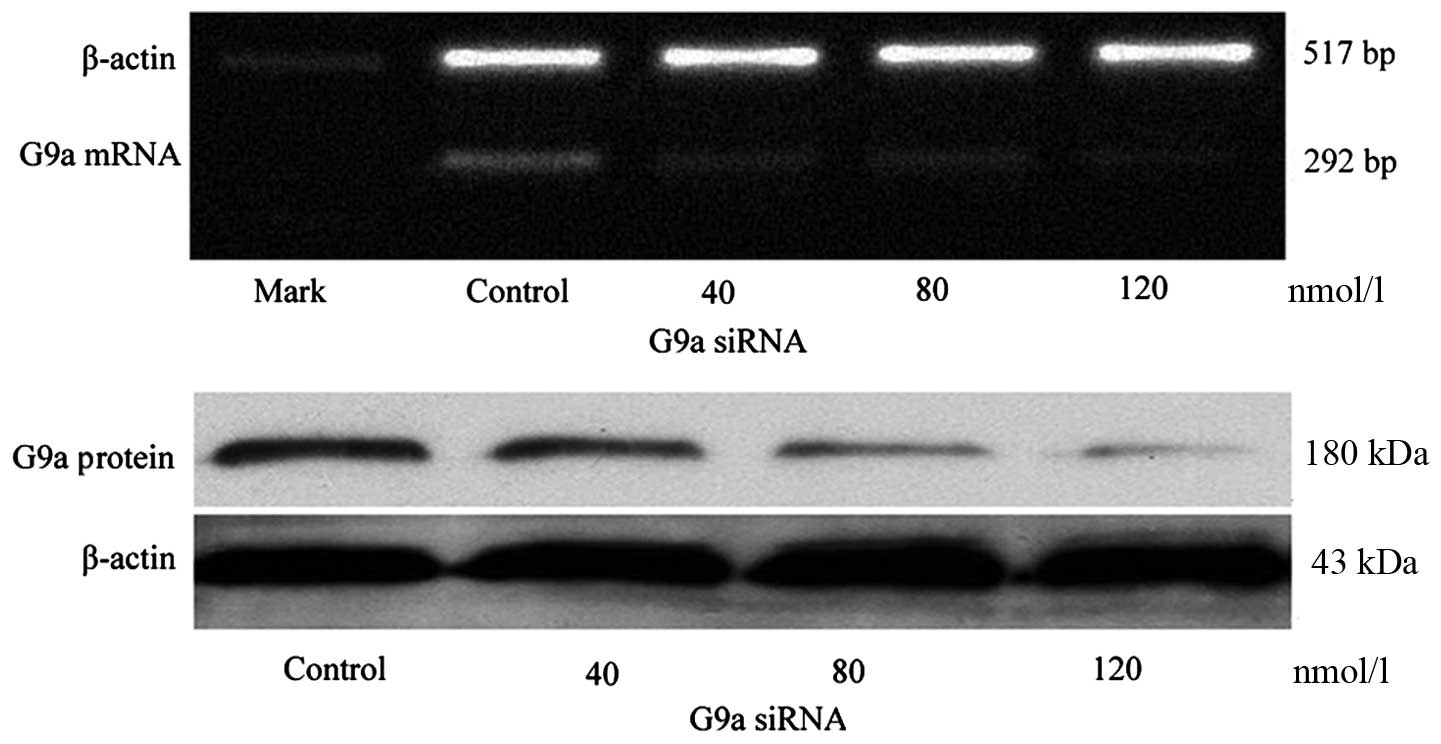
After 24 h transfection with G9a siRNA of 40, 80 and
120 nmol/l to MGC-803 cells, the amplification of G9a mRNA
attenuated concentration dependently. Gray value (to β-actin)
showed the amplification of G9a was 0.21±0.10 with 40 nmol/l,
0.11±0.04 with 80 nmol/l, and 0.08±0.02 with 120 nmol/l,
respectively, compared to the control (0.33±0.08),
χ2=17.43, P<0.05. The protein of G9a also decreased
concentration dependently (P<0.05; Fig. 2).
Depletion of G9a decreases cell
proliferation and promotes apoptosis in MGC-803 cells
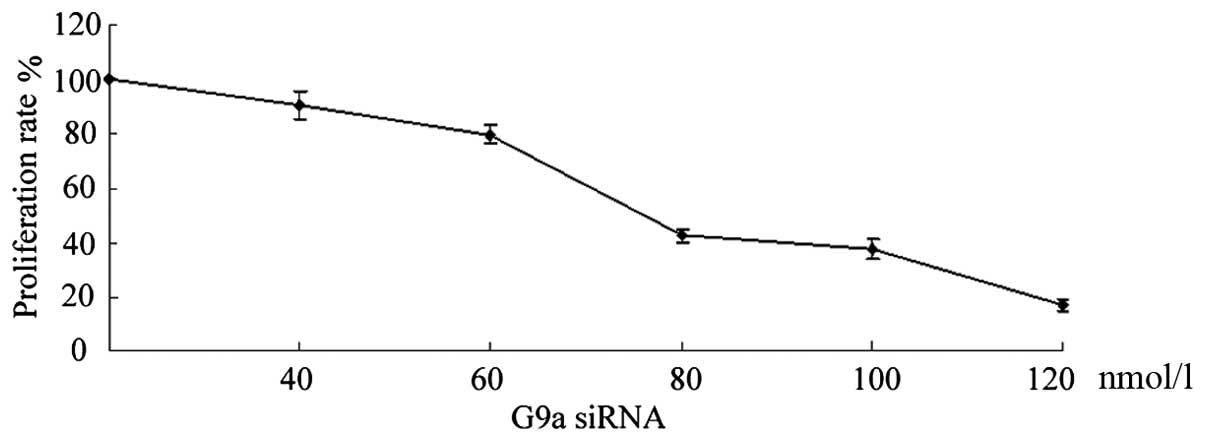
Depletion of G9a suppressed cell proliferation. To
elucidate the effect of G9a on proliferation and apoptosis, G9a was
knocked out by transfection of G9a siRNA in gastric cancer MGC-803
cells to exhibit cells biological changes. After 24-h transfection
with G9a siRNA, 82.36±4.46% reduction in cell density was seen with
40 nmol/l, 76.24±3.24% with 60 nmol/l, 42.53±3.25% with 80 nmol/l,
17.45±1.83% with 120 nmol/l (Fig.
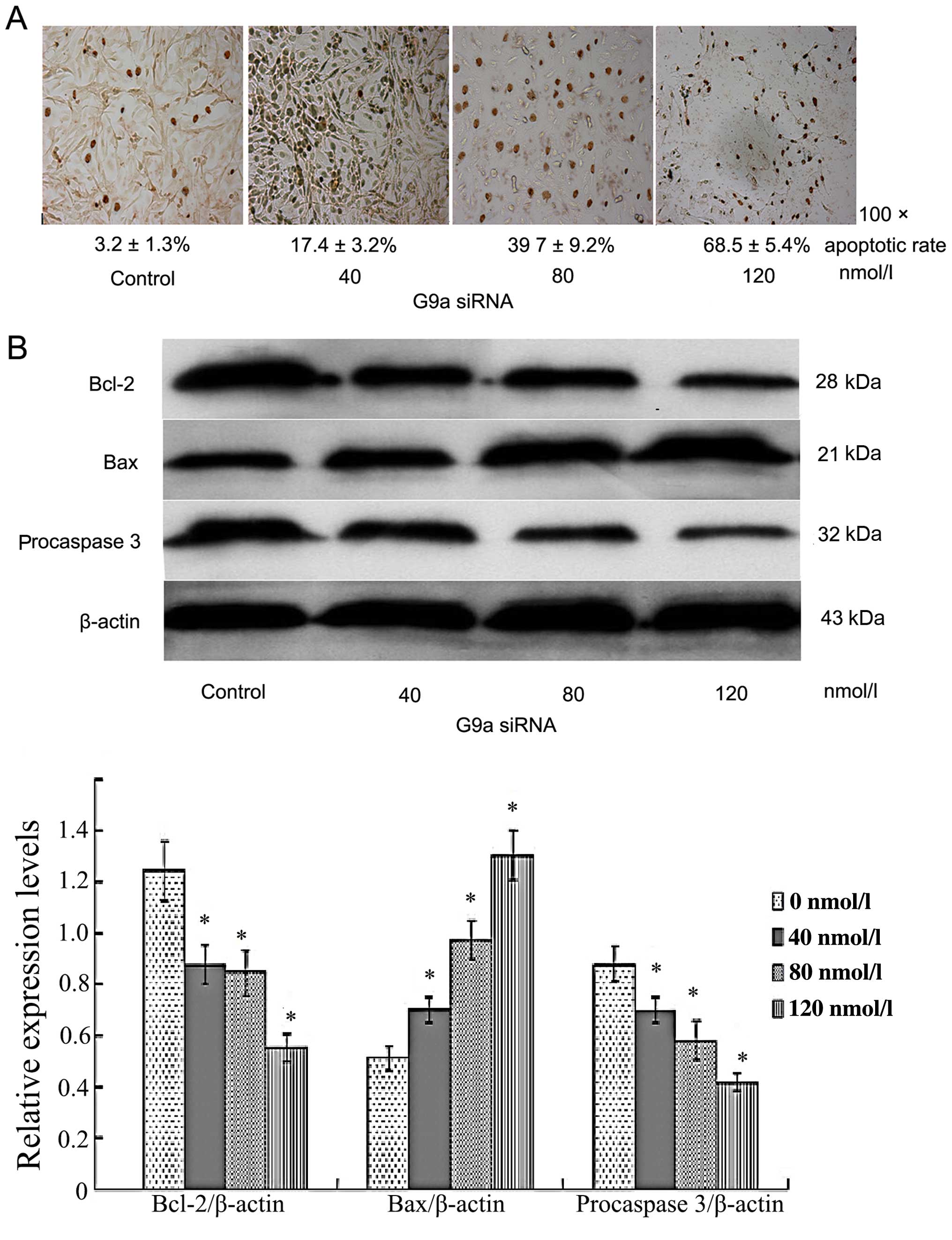
3). Apoptotic rate was 3.2±1.3, 17.4±3.2 and 39.7±9.2 and
68.5±5.4% after 24-h transfection of G9a siRNA with 0, 40, 80 and
120 nmol/l, respectively. The apoptotic rate was increased in a
concentration-dependent manner (P<0.05; Fig. 4A). We further investigated the
apoptosis-associated proteins BCL-2, pro-caspase-3 and Bax. It
demonstrated that G9a siRNA inhibited BCL-2, pro-caspase-3, and
promoted Bax significantly, thus promoting cell apoptosis (Fig. 4B).
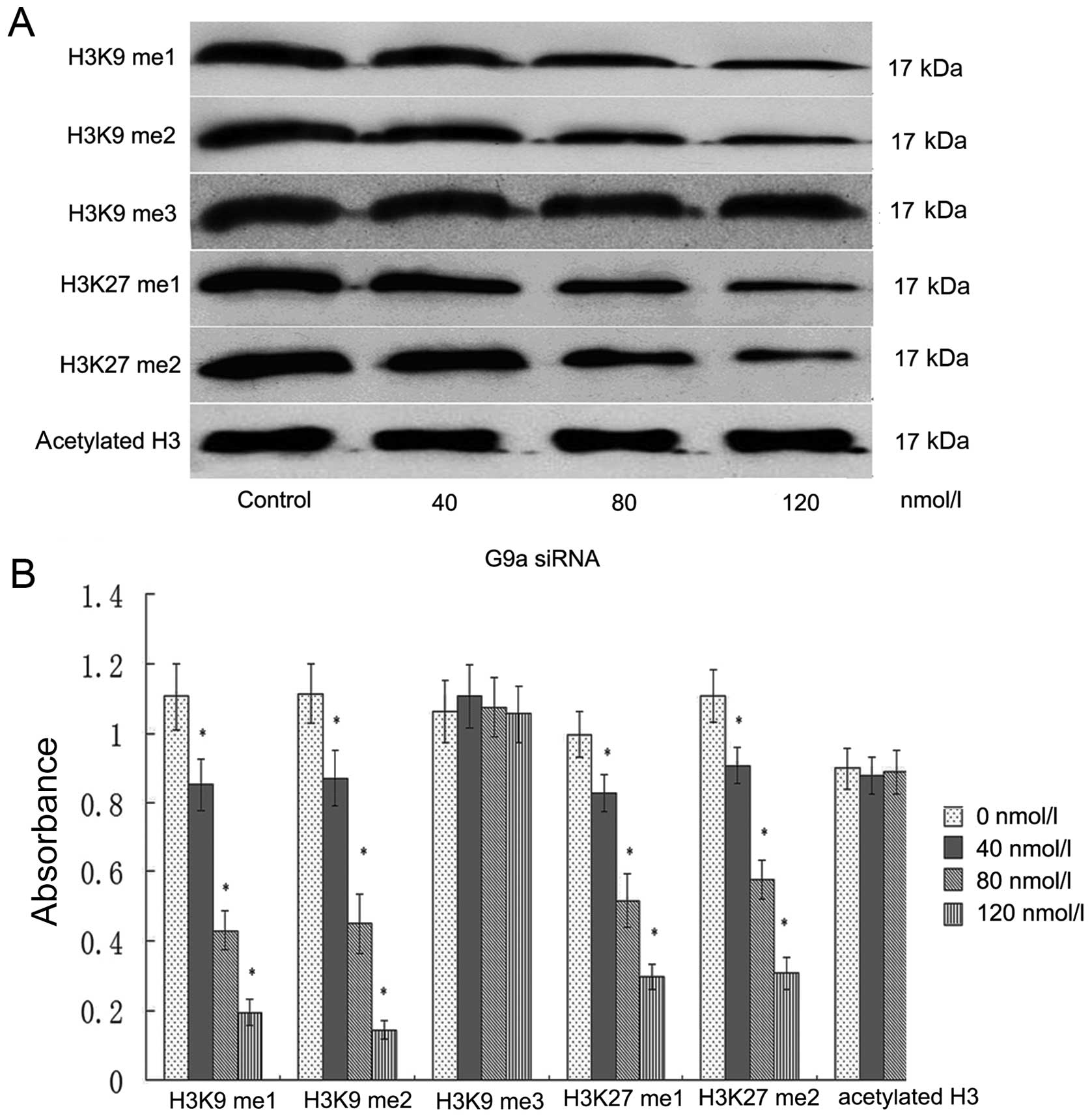
G9a depletion associates with reduction
of the H3K9 me1, H3K9 me2 and histone H3K27me and H3K27 me2 in
MGC-803 cells, but not with H3K9 me3, histone acetyl H3
Lysine residues can be mono, di, or trimethylated,
inducing different biological responses (16-18).
The present study was performed to identify the histone
modifications associated with the G9a, especially on histones
methylation of H3K9 and H3K27. To this end, we transfect of G9a
siRNA to gastric carcinoma MGC-803 cells and performed western blot
analysis to identify the effect of depletion of G9a on histone H3K9
and H3K27 and histone acetylation of H3. We demonstrated that
depletion of G9a for 24 h resulted in downregulation of histone
H3K9 me1, H3K9 me2 and H3K27 me1, H3K27 me2, compared with the
control. The relative (to β-actin), gray value showed H3K9 me1 was
1.21±0.19, 0.84±0.17, 0.42±0.14, 0.19±0.08, H3K9 me2 was 1.13±0.18,
0.86±0.15, 0.47±0.15, 0.16±0.04, H3K27 me was 1.01±0.12, 0.82±0.11,
0.51±0.14, 0.31±0.07; H3K27 me2 was 1.14±0.19, 0.91±0.14,
0.57±0.15, 0.33 ±0.05, in 0, 40, 80 and 120 nmol/l, respectively
(P<0.05). There was no change in histone H3K9 me3 and histone H3
acetylation. The expression of H3K9 me3 was 1.08±0.17, 1.11±0.16,
1.09±0.17, 1.07±0.16, Act-H3 was 0.93±0.15, 0.87±0.13, 0.91±0.16,
0.89±0.15 in 0, 40, 80 and 120 nmol/l, respectively (P>0.05)
(Fig. 5).
Discussion
Cancer cells are characterized by prominent
epigenetic dysregulation, including altered chromatin modification.
The histone methyltransferase G9a is overexpressed in a variety of
cancer types, such as pancreatic adenocarcinoma, lung cancer,
hepatocellular carcinoma (19–21),
and promotes tumor invasiveness and metastasis, although the
functional role of the HMTs overexpression in cancer remains
unclear. Previously we found the level of SUV39H1, another HMT, was
increased and associated with poor prognosis in gastric carcinoma
(22). In the present study, we
demonstrated that the expression of G9a, H3K9 me2 were aberrant in
gastric carcinoma. The expression of G9a, H3K9 me2 in gastric
carcinoma was 87.43 and 90.28%, respectively, which were higher
than that in the controls (P<0.05). Downregulation of H3K9 me1
in gastric carcinoma need to be investigated. Tissue invasion and
metastasis are the major causes of cancer-related death (23). The changes in global levels of
individual histone modifications are independently predictive of
the clinical outcome of prostate cancer, gastric adenocarcinomas,
as well as breast, ovarian and pancreatic cancers (24–26).
Some studies have found that HMTs specifically affect metastasis.
EZH2 (the polycomb group protein enhancer of zeste homolog 2) is
linked to cell proliferation and invasion in prostate and breast
cancer (27,28) and significantly associated with
distant metastases in gastric and prostate cancer (29,30).
We found that G9a and H3K9 me2 were positively correlated with
differentiation, depth of infiltration, lymph node metastasis and
TNM stage. These results support the hypothesis that aberrant
histone modification patterns are critically involved in the
tumorigenetic process and indicate that G9a seems to be required
for the maintenance of the malignant phenotype and could be valid
therapeutic targets in human neoplasia. The elevated levels of G9a
and H3K9 me2 predict poor prognosis. Chen et al (19) reported that G9a is endowed with
methyltransferase activity to concomitantly repress the downstream
effector Ep-CAM, thereby promoting the invasion step of the
invasion-metastasis cascade. The possible mechanisms of G9a
overexpression associated with cancer metastasis might be involved
in post-translational regulation. It is possible that high G9a
expressing cells may contain more cancer stem cell potential with
higher capability for metastasis.
The aberrant overexpression of G9a in gastric
carcinoma may make it a good candidate as a therapeutic molecular
target. We transfered G9a siRNA to MGC-803 cells and observed their
biological phenomenon. The evidence showed that depletion of G9a
reduced cell proliferation, and induced apoptosis in human gastric
cancer cell line MGC-803, along with downregulation of
pro-caspase-3, BCL-2, and significant upregulation of Bax, which
are related to apoptosis pathway. It was reported that G9a is
linked to aberrant silencing of several tumor-suppressor genes
through H3K9 methylation and DNA CpG methylation (31–33).
Current evidence suggests that H3K9 methylation and silencing of
the p16ink4a tumor suppressor gene can occur before CpG methylation
(34). G9a has been characterized
as transcriptional silencer by executing mono-, di- and
tri-methylation of euchromatic H3K9 as well as by promoting DNA
methylation (35). Reduction of G9a
expression coincided with the decrease in DNA methyltransferase
(DNMT) activity and indicated a functional impact of G9a on DNMT 1
operations. It was reported that RUNX3 was downregulated by hypoxia
in several gastric cancer cells and this suppression was on account
of histone deacetylation and methylation (36). This interaction enhanced the
catalytic activity of recombinant DNMT 1 in vitro, in the
absence of any other proteins (37). Regulation of DNA methylation by G9a
is independent of its catalytic activity, implying that its
physical presence, but not H3K9 methylation or methylation of
DNMT1, is relevant (14).
Lysine residues can be mono, di, or trimethylated,
inducing different biological responses (16,18,20).
Thus, for example, highly condensed heterochromatic regions show a
high degree of H3K9 me3, whereas euchromatic regions are
preferentially enriched in mono- and dimethylated H3K9 (38). G9a and GLP are homologous HMT and
mediate histones H3K9 me1 and H3K9 me2 and methylation at H3K27
(10). Studies in knockout mice for
Suv39h1/2 and G9a revealed that G9a is mainly responsible for
monomethylation and dimethylation of H3K9, whereas Suv39h1 and
Suv39h2 direct trimethylation of H3K9 (39). We found that depletion of G9a
decreased H3K9 me1, me2, and H3K27 me1, me2, and could not change
H3K9 me3 and histone acetylation of H3. Previously studied showed
that silencing Suv39H1 downregulated histone acetylated H3, H3K9
me3, and enhanced P15 in MGC-803. It did not affect H3K27, H3K14
and H4 (22,40).
In conclusion, our data show that G9a and H3K9 me2
are highly expressed in gastric cancer, and associate with poor
prognosis. Depletion of G9a reduces H3K9 me1, H3K9 me2, H3K27 me1,
H3K27 me2, and producse tumor cell apoptosis and inhibits cell
proliferation. The results from this study highlight the potential
of G9a, a methyltransferase, that could be a therapeutic target for
gastric carcinoma.
Acknowledgments
The present study was partly supported by
grant-in-aid from the Foundation of Science and Technology of
Fujian Medical University, Fujian, China (no. FZS13004Z), the Key
Medical Innovations Project Science Research Foundation of Health
Bureau of Fujian Provincial Health (2012-CX-32), the Science
Foundation of Fujian Province (2012J01420), and the Introductive
Major Project of Science Research Foundation of Fujian Province
(201212004).
Abbreviations:
|
H3K9 me1
|
monomethylated histone H3 lysine 9
|
|
H3K9 me2
|
dimethylated histone H3 lysine 9
|
|
H3K9 me3
|
trimethylation of H3K9
|
|
H3K27 me1
|
monomethylated histone H3 lysine
27
|
|
H3K27 me2
|
dimethylated histone H3 lysine 27
|
|
TNM
|
tumor-node-metastasis
|
|
HMT
|
histone methyltransferases
|
|
GLP
|
G9a-like protein
|
|
DNMT
|
DNA methyltransferase
|
References
|
1
|
Bannister AJ, Zegerman P, Partridge JF,
Miska EA, Thomas JO, Allshire RC and Kouzarides T: Selective
recognition of methylated lysine 9 on histone H3 by the HP1 chromo
domain. Nature. 410:120–124. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jacobs SA, Taverna SD, Zhang Y, Briggs SD,
Li J, Eissenberg JC, Allis CD and Khorasanizadeh S: Specificity of
the HP1 chromo domain for the methylated N-terminus of histone H3.
EMBO J. 20:5232–5241. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lachner M, O'Carroll D, Rea S, Mechtler K
and Jenuwein T: Methylation of histone H3 lysine 9 creates a
binding site for HP1 proteins. Nature. 410:116–120. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kouzarides T: Chromatin modifications and
their function. Cell. 128:693–705. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schneider R, Bannister AJ and Kouzarides
T: Unsafe SETs: Histone lysine methyltransferases and cancer.
Trends Biochem Sci. 27:396–402. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aagaard L, Schmid M, Warburton P and
Jenuwein T: Mitotic phosphorylation of SUV39H1, a novel component
of active centromeres, coincides with transient accumulation at
mammalian centromeres. J Cell Sci. 113:817–829. 2000.PubMed/NCBI
|
|
7
|
Aagaard L, Laible G, Selenko P, Schmid M,
Dorn R, Schotta G, Kuhfittig S, Wolf A, Lebersorger A, Singh PB, et
al: Functional mammalian homologues of the Drosophila PEV-modifier
Su(var)3-9 encode centromere-associated proteins which complex with
the heterochromatin component M31. EMBO J. 18:1923–1938. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Melcher M, Schmid M, Aagaard L, Selenko P,
Laible G and Jenuwein T: Structure-function analysis of SUV39H1
reveals a dominant role in heterochromatin organization, chromosome
segregation, and mitotic progression. Mol Cell Biol. 20:3728–3741.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
O'Carroll D, Scherthan H, Peters AH,
Opravil S, Haynes AR, Laible G, Rea S, Schmid M, Lebersorger A,
Jerratsch M, et al: Isolation and characterization of Suv39h2, a
second histone H3 methyltransferase gene that displays
testis-specific expression. Mol Cell Biol. 20:9423–9433. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tachibana M, Sugimoto K, Fukushima T and
Shinkai Y: Set domain-containing protein, G9a, is a novel
lysine-preferring mammalian histone methyltransferase with
hyperactivity and specific selectivity to lysines 9 and 27 of
histone H3. J Biol Chem. 276:25309–25317. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tachibana M, Sugimoto K, Nozaki M, Ueda J,
Ohta T, Ohki M, Fukuda M, Takeda N, Niida H, Kato H, et al: G9a
histone methyltransferase plays a dominant role in euchromatic
histone H3 lysine 9 methylation and is essential for early
embryogenesis. Genes Dev. 16:1779–1791. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Estève PO, Patnaik D, Chin HG, Benner J,
Teitell MA and Pradhan S: Functional analysis of the N- and
C-terminus of mammalian G9a histone H3 methyltransferase. Nucleic
Acids Res. 33:3211–3223. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong KB, Maksakova IA, Mohn F, Leung D,
Appanah R, Lee S, Yang HW, Lam LL, Mager DL, Schübeler D, et al:
DNA methylation in ES cells requires the lysine methyltransferase
G9a but not its catalytic activity. EMBO J. 27:2691–2701. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ikegami K, Iwatani M, Suzuki M, Tachibana
M, Shinkai Y, Tanaka S, Greally JM, Yagi S, Hattori N and Shiota K:
Genome-wide and locus-specific DNA hypomethylation in G9a deficient
mouse embryonic stem cells. Genes Cells. 12:1–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma X, Fang Y, Beklemisheva A, Dai W, Feng
J, Ahmed T, Liu D and Chiao JW: Phenylhexyl isothiocyanate inhibits
histone deacetylases and remodels chromatins to induce growth
arrest in human leukemia cells. Int J Oncol. 28:1287–1293.
2006.PubMed/NCBI
|
|
16
|
Lachner M, Sengupta R, Schotta G and
Jenuwein T: Trilogies of histone lysine methylation as epigenetic
landmarks of the eukaryotic genome. Cold Spring Harb Symp Quant
Biol. 69:209–218. 2004. View Article : Google Scholar
|
|
17
|
Jenuwein T: The epigenetic magic of
histone lysine methylation. FEBS J. 273:3121–3135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Santos-Rosa H, Schneider R, Bannister AJ,
Sherriff J, Bernstein BE, Emre NC, Schreiber SL, Mellor J and
Kouzarides T: Active genes are tri-methylated at K4 of histone H3.
Nature. 419:407–411. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen MW, Hua KT, Kao HJ, Chi CC, Wei LH,
Johansson G, Shiah SG, Chen PS, Jeng YM, Cheng TY, et al: H3K9
histone methyltransferase G9a promotes lung cancer invasion and
metastasis by silencing the cell adhesion molecule Ep-CAM. Cancer
Res. 70:7830–7840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan Y, Tang AJ, Castoreno AB, Kuo SY,
Wang Q, Kuballa P, Xavier R, Shamji AF, Schreiber SL and Wagner BK:
Gossypol and an HMT G9a inhibitor act in synergy to induce cell
death in pancreatic cancer cells. Cell Death Dis. 4:e6902013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kondo Y, Shen L, Suzuki S, Kurokawa T,
Masuko K, Tanaka Y, Kato H, Mizuno Y, Yokoe M, Sugauchi F, et al:
Alterations of DNA methylation and histone modifications contribute
to gene silencing in hepatocellular carcinomas. Hepatol Res.
37:974–983. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cai L, Ma X, Huang Y, Zou Y and Chen X:
Aberrant histone methylation and the effect of Suv39H1 siRNA on
gastric carcinoma. Oncol Rep. 31:2593–2600. 2014.PubMed/NCBI
|
|
23
|
Steeg PS: Metastasis suppressors alter the
signal transduction of cancer cells. Nat Rev Cancer. 3:55–63. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seligson DB, Horvath S, Shi T, Yu H, Tze
S, Grunstein M and Kurdistani SK: Global histone modification
patterns predict risk of prostate cancer recurrence. Nature.
435:1262–1266. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park YS, Jin MY, Kim YJ, Yook JH, Kim BS
and Jang SJ: The global histone modification pattern correlates
with cancer recurrence and overall survival in gastric
adenocarcinoma. Ann Surg Oncol. 15:1968–1976. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei Y, Xia W, Zhang Z, Liu J, Wang H,
Adsay NV, Albarracin C, Yu D, Abbruzzese JL, Mills GB, et al: Loss
of trimethylation at lysine 27 of histone H3 is a predictor of poor
outcome in breast, ovarian, and pancreatic cancers. Mol Carcinog.
47:701–706. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ding L, Erdmann C, Chinnaiyan AM, Merajver
SD and Kleer CG: Identification of EZH2 as a molecular marker for a
precancerous state in morphologically normal breast tissues. Cancer
Res. 66:4095–4099. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bryant RJ, Cross NA, Eaton CL, Hamdy FC
and Cunliffe VT: EZH2 promotes proliferation and invasiveness of
prostate cancer cells. Prostate. 67:547–556. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi JH, Song YS, Yoon JS, Song KW and Lee
YY: Enhancer of zeste homolog 2 expression is associated with tumor
cell proliferation and metastasis in gastric cancer. APMIS.
118:196–202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Min J, Zaslavsky A, Fedele G, McLaughlin
SK, Reczek EE, De Raedt T, Guney I, Strochlic DE, Macconaill LE,
Beroukhim R, et al: An oncogene-tumor suppressor cascade drives
metastatic prostate cancer by coordinately activating Ras and
nuclear factor-kappaB. Nat Med. 16:286–294. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Watanabe H, Soejima K, Yasuda H, Kawada I,
Nakachi I, Yoda S, Naoki K and Ishizaka A: Deregulation of histone
lysine methyl-transferases contributes to oncogenic transformation
of human bronchoepithelial cells. Cancer Cell Int. 8:152008.
View Article : Google Scholar
|
|
32
|
Wozniak RJ, Klimecki WT, Lau SS, Feinstein
Y and Futscher BW: 5-Aza-2′-deoxycytidine-mediated reductions in
G9A histone methyltransferase and histone H3 K9 di-methylation
levels are linked to tumor suppressor gene reactivation. Oncogene.
26:77–90. 2007. View Article : Google Scholar
|
|
33
|
Xin ZH, Ma HL, Wu SY and Dai CH:
Determination of the inhibitory activity of angiotensin-converting
enzyme inhibitor captopril by high performance capillary
electrophoresis. Yao Xue Xue Bao. 38:843–845. 2003.In Chinese.
|
|
34
|
Bachman KE, Park BH, Rhee I, Rajagopalan
H, Herman JG, Baylin SB, Kinzler KW and Vogelstein B: Histone
modifications and silencing prior to DNA methylation of a tumor
suppressor gene. Cancer Cell. 3:89–95. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Collins R and Cheng X: A case study in
cross-talk: The histone lysine methyltransferases G9a and GLP.
Nucleic Acids Res. 38:3503–3511. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee SH, Kim J, Kim WH and Lee YM: Hypoxic
silencing of tumor suppressor RUNX3 by histone modification in
gastric cancer cells. Oncogene. 28:184–194. 2009. View Article : Google Scholar
|
|
37
|
Estève PO, Chin HG, Smallwood A, Feehery
GR, Gangisetty O, Karpf AR, Carey MF and Pradhan S: Direct
interaction between DNMT1 and G9a coordinates DNA and histone
methylation during replication. Genes Dev. 20:3089–3103. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martin C and Zhang Y: The diverse
functions of histone lysine methylation. Nat Rev Mol Cell Biol.
6:838–849. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kondo Y, Shen L, Ahmed S, Boumber Y,
Sekido Y, Haddad BR and Issa JP: Downregulation of histone H3
lysine 9 methyltransferase G9a induces centrosome disruption and
chromosome instability in cancer cells. PLoS One. 3:e20372008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Czermin B, Schotta G, Hülsmann BB, Brehm
A, Becker PB, Reuter G and Imhof A: Physical and functional
association of SU(VAR)3–9 and HDAC1 in Drosophila. EMBO Rep.
2:915–919. 2001. View Article : Google Scholar : PubMed/NCBI
|