Introduction
Gastric cancer is the second leading cause of
cancer-related death worldwide (1).
For most patients, gastric cancer is only diagnosed at the advanced
stages with poor prognosis (2,3).
Gastric carcinogenesis is known as a multistep process involving a
series of epigenetic and genetic alterations (4–11). The
mechanism of gastric carcinogenesis has not yet been fully
elucidated and more studies are needed to search for novel
molecules which are involved in the process.
Long non-coding RNAs (lncRNAs) are RNA transcripts
which are >200 bp in length and do not encode for a protein
(12,13). Studies suggest that lncRNAs
constitute an important component of tumor biology (14–18).
Most lncRNAs play a functional role in gene expression by targeting
either genomically local or distant genes (19–21).
Evidence suggests that lncRNAs play essential roles in
tumorigenesis (14,22–24)
and cancer progression (15,25–27)
by acting as either oncogenes or tumor suppressors.
The regulation of HOX genes by lncRNAs is
gaining great interest in developmental biology research.
HOX genes are highly conserved at the genomic level. The
proteins which HOX genes encode are master regulators of
embryonic development and continue to be expressed throughout
adulthood in various tissues. HOXA transcript at the distal tip
(HOTTIP) is at the 5′ end of the HOXA cluster and
upregulates the expression of 5′ HOXA genes by binding the
adaptor protein WDR5 and targeting the WDR5/MLL complex (28). Evidence suggests that HOTTIP and
homeobox protein Hox-A13 (HOXA13) are both upregulated and
associated with progression and poor survival of hepatocellular
carcinoma (29). Moreover, the
expression of HOTTIP and HOXA13 showed a high correlation in
hepatocellular carcinoma (29). The
role of HOTTIP has also been investigated in pancreatic (30,31)
and lung cancer, and tongue squamous cell carcinoma (32,33).
For example, HOTTIP promoted disease progression and gemcitabine
resistance by regulating HOXA13 in pancreatic cancer (30). In addition, HOTTIP promoted tumor
growth and inhibited cell apoptosis in lung cancer (32). In addition, HOTTIP was found to be
highly expressed and correlated with the progression of tongue
squamous cell carcinoma (33).
However, the role of HOTTIP in gastric cancer has never been
reported.
In the present study, we investigated the expression
of HOTTIP in gastric tissues and the function of HOTTIP in gastric
cancer cells, with the aim of elucidating the mechanisms of gastric
carcinogenesis and progression.
Materials and methods
Cell culture
Human immortal gastric epithelial cell line GES-1
and human gastric cancer cell lines SGC7901, MKN28, MKN45 and
MGC803 were obtained from the Cell Resource Center, Shanghai
Institute of Biochemistry and Cell Biology at the Chinese Academy
of Sciences. Cells were cultured in RPMI-1640 medium supplemented
with 10% fetal bovine serum (FBS) (both from Gibco, Carlsbad, CA,
USA) at 37°C in a humidified incubator containing 5% carbon
dioxide.
Small interfering RNA (siRNA)
transfection
The siRNA oligonucleotides targeting HOTTIP, HOXA13
and the negative control were obtained from GenePharma Co., Ltd.
(Shanghai, China). Transfection of the oligonucleotides was
conducted with X-tremeGENE siRNA transfection reagent (Roche
Molecular Biochemicals, Indianapolis, IN, USA) according to the
manufacturer's instructions. The sequences of siRNAs used in the
present study are listed in Table
I.
 | Table IThe sequences of siRNAs used in the
present study. |
Table I
The sequences of siRNAs used in the
present study.
| siRNAs | Sense (5′-3′) | Antisense
(5′-3′) |
|---|
| NC |
UUCUCCGAACGUGUCACGUTT |
ACGUGACACGUUCGGAGAATT |
| siHOTTIP #1 |
GCUUUAGAGCCACAUACUUTT |
AAGUAUGUGGCUCUAAAGCTT |
| siHOTTIP #2 |
GAGACAGAGUAGGGUUCUATT |
UAGAACCCUACUCUGUCUCTT |
| siHOTTIP #3 |
GGCACUUUAUAUGCUGUAATT |
UUACAGCAUAUAAAGUGCCTT |
| siHOXA13 #1 |
GCCACGAAUAAAUUCAUUATT |
UAAUGAAUUUAUUCGUGGCTT |
| siHOXA13 #2 |
GCGGACAAGUACAUGGAUATT |
UAUCCAUGUACUUGUCCGCTT |
| siHOXA13 #3 |
GACGAGCUCAACAAGAACATT |
UGUUCUUGUUGAGCUCGUCTT |
RNA extraction and quantitative real-time
PCR
Total RNA was extracted from cells or tissues using
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer's instructions. The cDNA was synthesized using the
RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc., Rockford, IL, USA). Quantitative real-time PCR
was performed using a SYBR Premix Ex Taq™ II (Takara
Biotechnology Co., Ltd., Dalian, China) on a Bio-Rad CFX-96
Real-Time PCR system. GAPDH was used as an internal control. The
sequences of the primers are listed in Table II. All qRT-PCR reactions were
performed in triplicate.
| Table IIqRT-PCR primers used in the present
study. |
Table II
qRT-PCR primers used in the present
study.
| Genes | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| GAPDH |
GACTCATGACCACAGTCCATGC |
AGAGGCAGGGATGATGTTCTG |
| HOTTIP |
CCTAAAGCCACGCTTCTTTG |
TGCAGGCTGGAGATCCTACT |
| HOXA13 |
TGGAACGGCCAAATGTACTG |
TGGCGTATTCCCGTTCAAGT |
| HOXA11 |
GTACTTACTACGTCTCGGGTCCAG |
AGTCTCTGTGCACGAGCTCCT |
| HOXA10 |
GGGGACTTCTCTTCCAGTTTC |
GGGAGAATTGTGGTGTGCTT |
| HOXA9 |
CCACGCTTGACACTCACACT |
AGTTGGCTGCTGGGTTATTG |
Cell proliferation and colony formation
assays
Cell proliferation was measured by the Cell Counting
Kit-8 (CCK-8) assay (7Sea Biotech Co., Ltd., Shanghai, China).
Cells transfected with siRNA were seeded and cultured into 96-well
plates (3×103 cells/well) in 100 µl medium. At
different time points indicated in the figures, 10 µl CCK-8
solution was added into the medium and further incubated with the
cells for 3 h. The optical density (OD) was measured using a
microplate reader at 450 nm. The CCK-8 assays were performed in
triplicate.
For colony formation assay, cells transfected with
different siRNAs were seeded into 6-well plates at 300 cells/well.
After 14 days of incubation, cells were fixed with methyl alcohol
and stained with 0.5% crystal violet. The number of colonies (≥50
cells/colony) was counted. Each experiment was performed in
triplicate.
Cell migration and invasion assays
For migration assays, 5×104 cells were
plated in the top chamber with a non-coated membrane (24-well
insert; pore size, 8-µm; Corning, Corning, NY, USA). For
invasion assays, 1.5×105 cells were plated in the top
chamber with a Matrigel-coated membrane (24-well insert; pore size,
8-µm; Corning). Medium without serum was used in the top
chamber in both assays. Medium with 10% FBS was added to the lower
chamber. After incubation for 24 h (migration assay) or 48 h
(invasion assay), respectively, the cells that did not migrate or
invade through the pores were removed using a cotton swab. Cells on
the lower surface of the membrane were fixed with 4%
paraformaldehyde and stained with 0.1% crystal violet. The number
of migrated or invaded cells was counted. Each experiment was
performed in triplicate.
Western blot analysis
Cells were lysed with RIPA buffer containing
complete protease inhibitor mixture (Roche Molecular Biochemicals).
Proteins were separated by dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred to nitrocellulose
membranes (Pall Life Sciences, Ann Arbor, MI, USA). The membranes
were blocked in 5% non-fat milk and blotted with antibodies against
GAPDH (1:2,000) and HOXA13 (1:200) (both from Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), respectively. The
membranes were then incubated with horseradish
peroxidase-conjugated secondary antibodies and visualized with an
enhanced chemiluminescence reagent.
Tissue samples
A total of 50 paired gastric tissue samples (cancer
lesions and adjacent non-tumor mucosae) of gastric cancer patients
were obtained from the Department of General Surgery, The First
Affiliated Hospital of Xi'an Jiaotong University between June 2013
and February 2014. All patients did not receive chemotherapy or
radiotherapy prior to surgery. All samples were collected in the
same manner. The samples were immediately frozen in liquid nitrogen
and stored at −80°C until they were used. Informed consent was
obtained from each patient before the surgery. The present study
was approved by the Research Ethics Committee of Xi'an Jiaotong
University.
Statistical analysis
Statistical analysis was performed using IBM SPSS
Statistics software (IBM Corp., Armonk, NY, USA). Student's t-test
for parametric variables was used. Spearman test was used to
establish the correlation between HOTTIP and HOXA13. Data are
presented as mean ± SEM unless otherwise indicated. All P-values
were determined from two-sided tests, and statistical significance
was determined based on a P-value of 0.05.
Results
HOTTIP is upregulated in gastric cancer
cells and downregulation of HOTTIP inhibits cancer cell growth
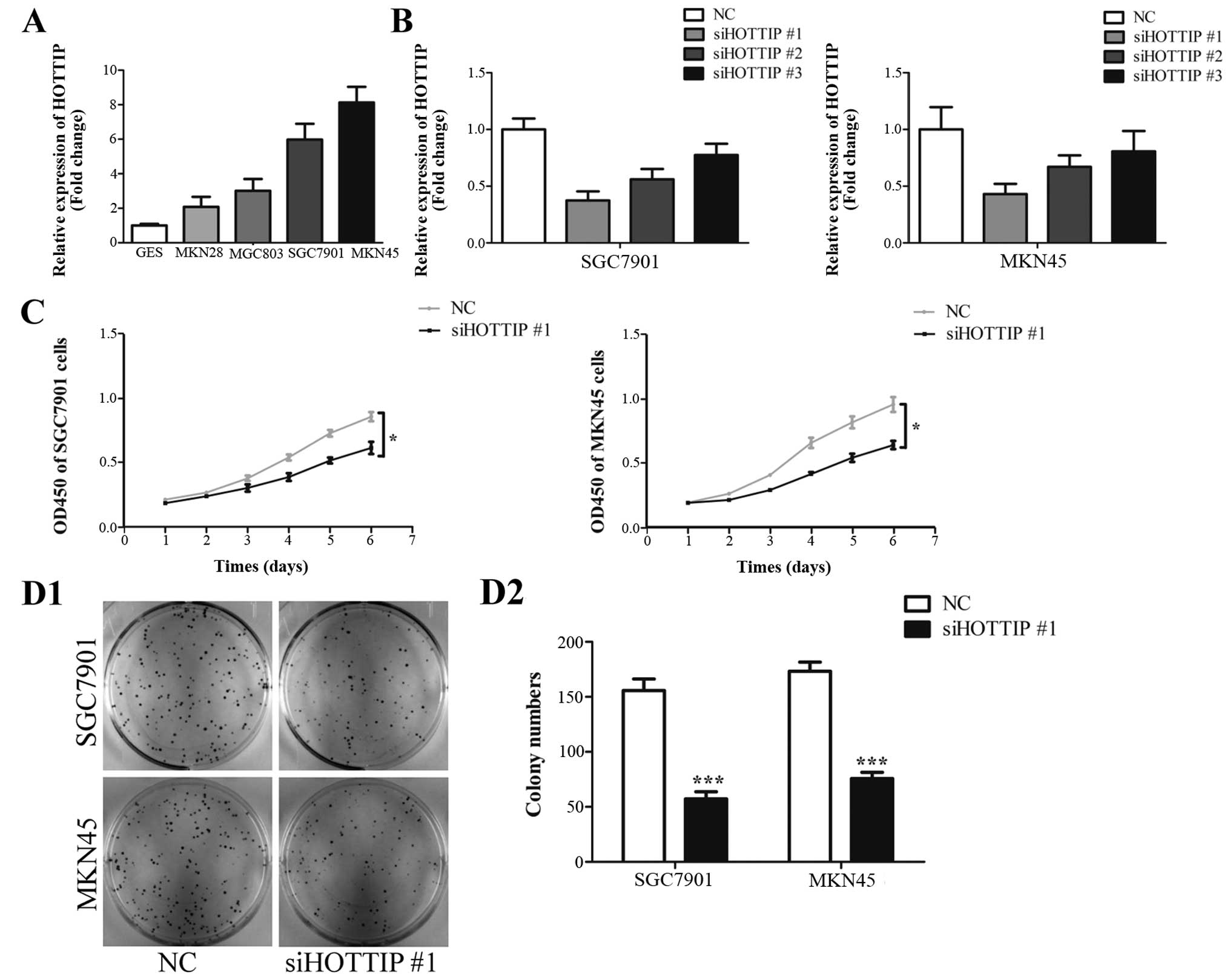
To determine the role of HOTTIP in gastric cancer,
we first investigated the expression of HOTTIP in the GES-1, MKN28,
MGC803, SGC7901 and MKN45 cell lines. In addition, we found that
HOTTIP was upregulated in gastric cancer cell lines compared with
that noted in the GES-1 cells (Fig.
1A). Then, we investigated the effect of HOTTIP on cell growth
by downregulating HOTTIP expression in the SGC7901 and MKN45 cells.
Efficiency of HOTTIP knockdown in the SGC7901 and MKN45 cells by
three specific siRNAs was confirmed by qRT-PCR and siHOTTIP #1 was
used in the following experiments (Fig.
1B). Knockdown of HOTTIP inhibited cell proliferation in the
SGC7901 and MKN45 cells (Fig. 1C).
The inhibition of cell growth by HOTTIP knockdown was further
confirmed by colony formation assay. Downregulation of HOTTIP
decreased colony numbers in the SGC7901 and MKN45 cells (Fig. 1D1 and D2). These results suggest
that HOTTIP plays a growth-promoting role in gastric cancer
cells.
Downregulation of HOTTIP inhibits cell
migration and invasion in gastric cancer
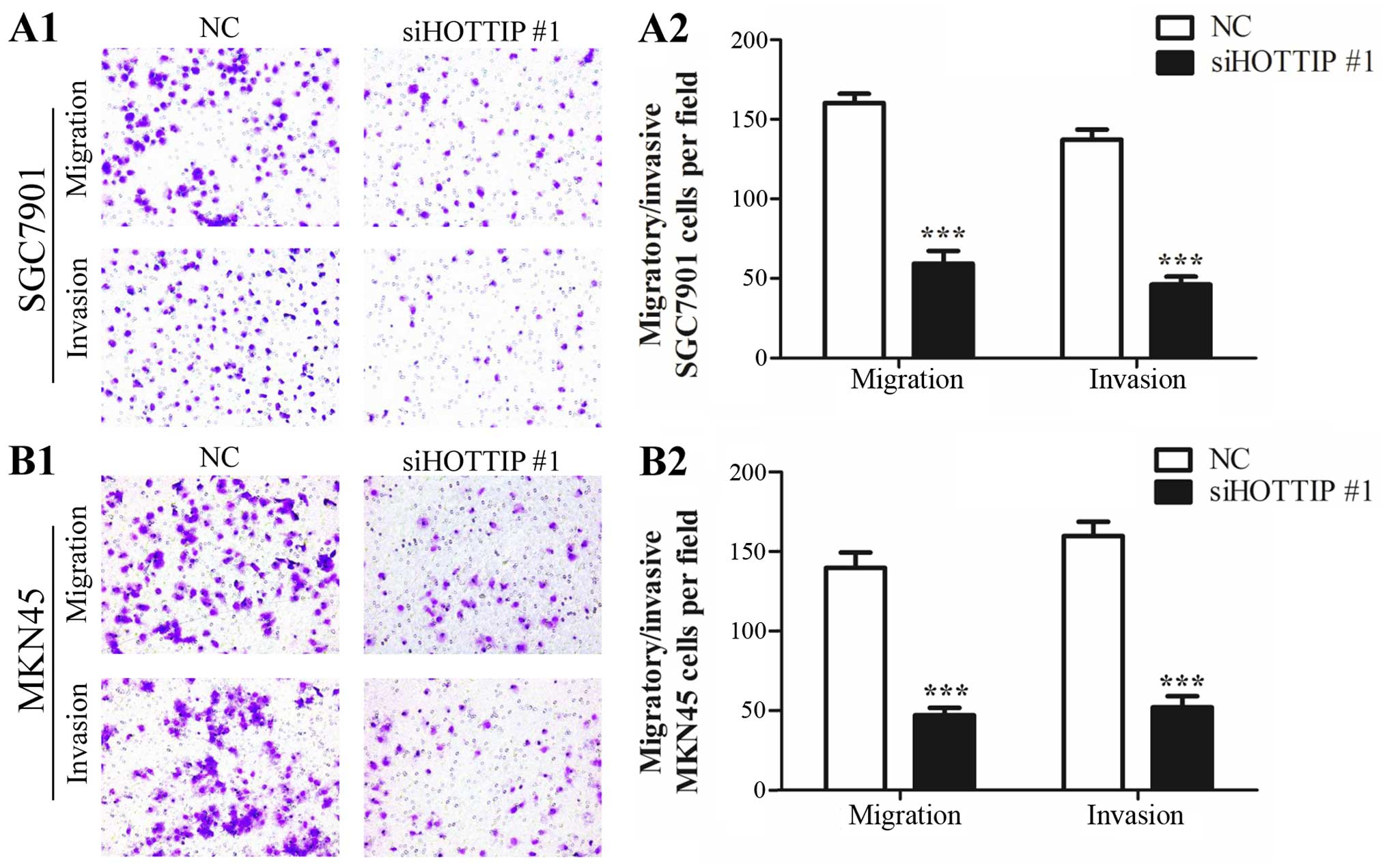
We next investigated the effect of HOTTIP on the
migration and invasion of SGC7901 and MKN45 cells. Downregulation
of HOTTIP led to a 2- to 3-fold reduction in the migratory and
invasive capabilities of the SGC7901 cells (Fig. 2A1 and A2). Similar results were
observed in the MKN45 cells with decreased expression of HOTTIP
(Fig. 2B1 and B2). These results
suggest that HOTTIP promotes both migration and invasion of gastric
cancer cells.
Downregulation of HOTTIP leads to
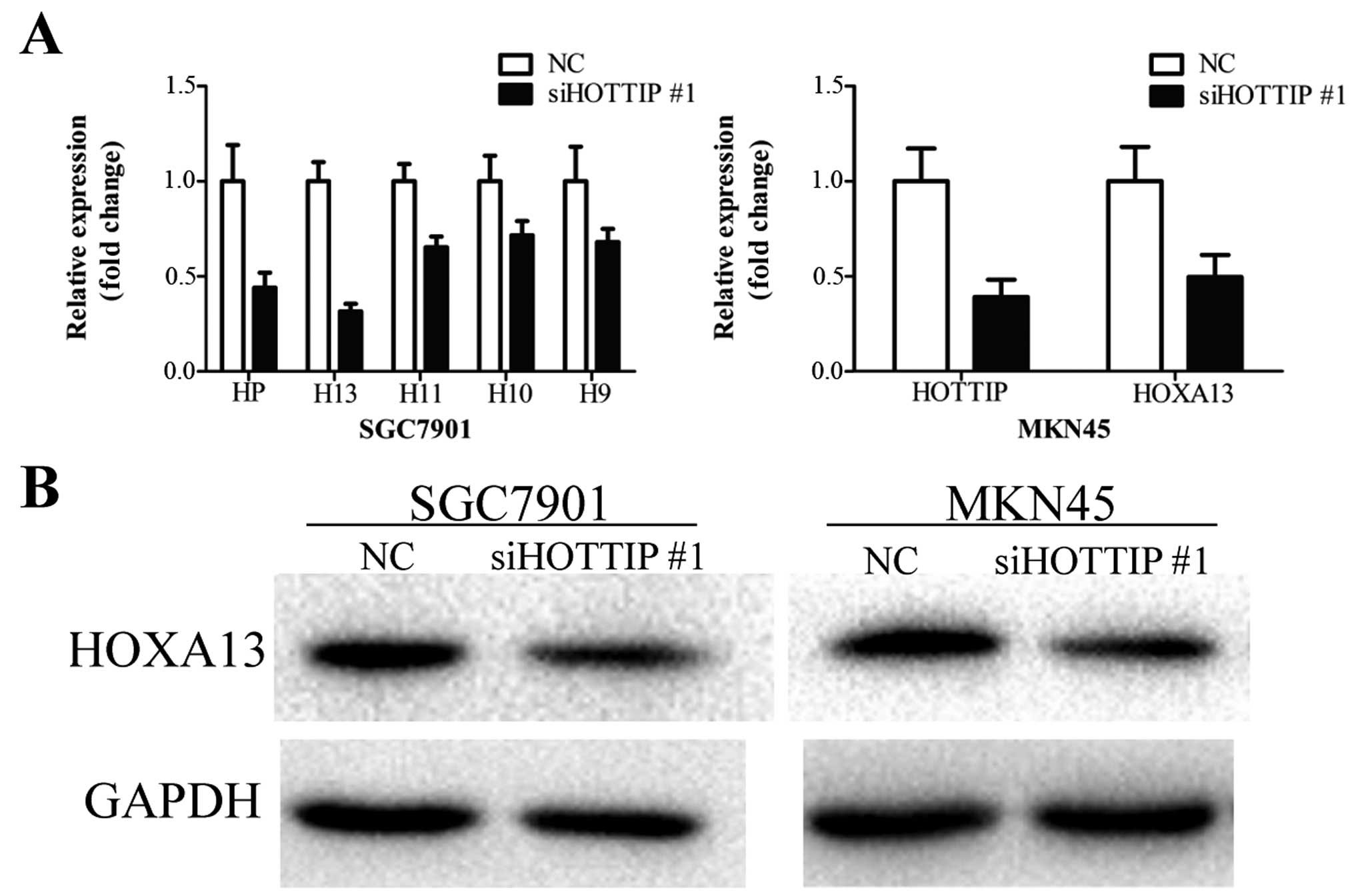
decreased HOXA13 expression in gastric cancer cells
HOTTIP knockdown was previously found to lead to a
reduction in HOXA gene expression in primary human
fibroblasts (28), hepatocellular
carcinoma (29) and pancreatic
cancer cells (30,31). To ascertain whether HOTTIP exhibits
a similar function in gastric cancer cells, we measured the
expression of several HOXA genes (HOXA13, HOXA11, HOXA10 and
HOXA9) in the SGC7901 cells treated with siHOTTIP #1.
Downregulation of HOTTIP led to different degrees of decrease in
the expression levels of these genes, among which HOXA13 expression
was decreased the most (Fig. 3A).
Downregulation of HOXA13 expression was further confirmed in MKN45
cells by qRT-PCR (Fig. 3A).
Knockdown of HOTTIP inhibited the HOXA13 protein level in the
SGC7901 and MKN45 cells (Fig. 3B).
These results suggest that HOTTIP regulates HOXA13 expression in
gastric cancer cells.
HOXA13 is involved in HOTTIP-induced
malignant phenotypes of gastric cancer cells
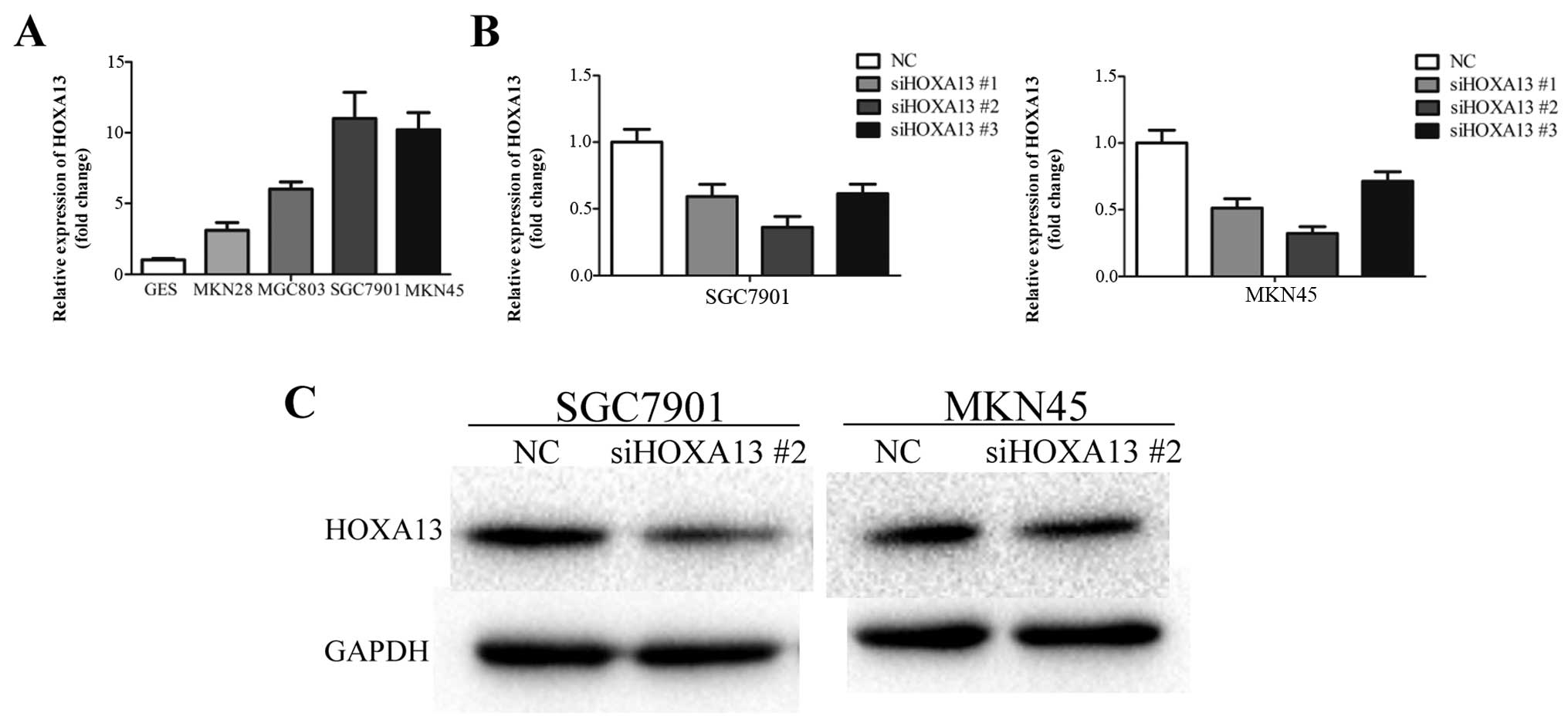
We investigated the expression of HOXA13 in the
GES-1 and gastric cancer cell lines. HOXA13 was upregulated in the
gastric cancer cell lines compared with GES-1, which was similar to
the HOTTIP expression pattern (Fig.
4A). To investigate the role of HOXA13 in gastric cancer, three
specific siRNAs against HOXA13 were used to inhibit HOXA13 mRNA
expression in the SGC7901 and MKN45 cells. siHOXA13 #2 showed most
significant knockdown efficiency and was used in the following
experiments (Fig. 4B). siHOXA13 #2
led to a clear reduction in the protein level of HOXA13 (Fig. 4C).
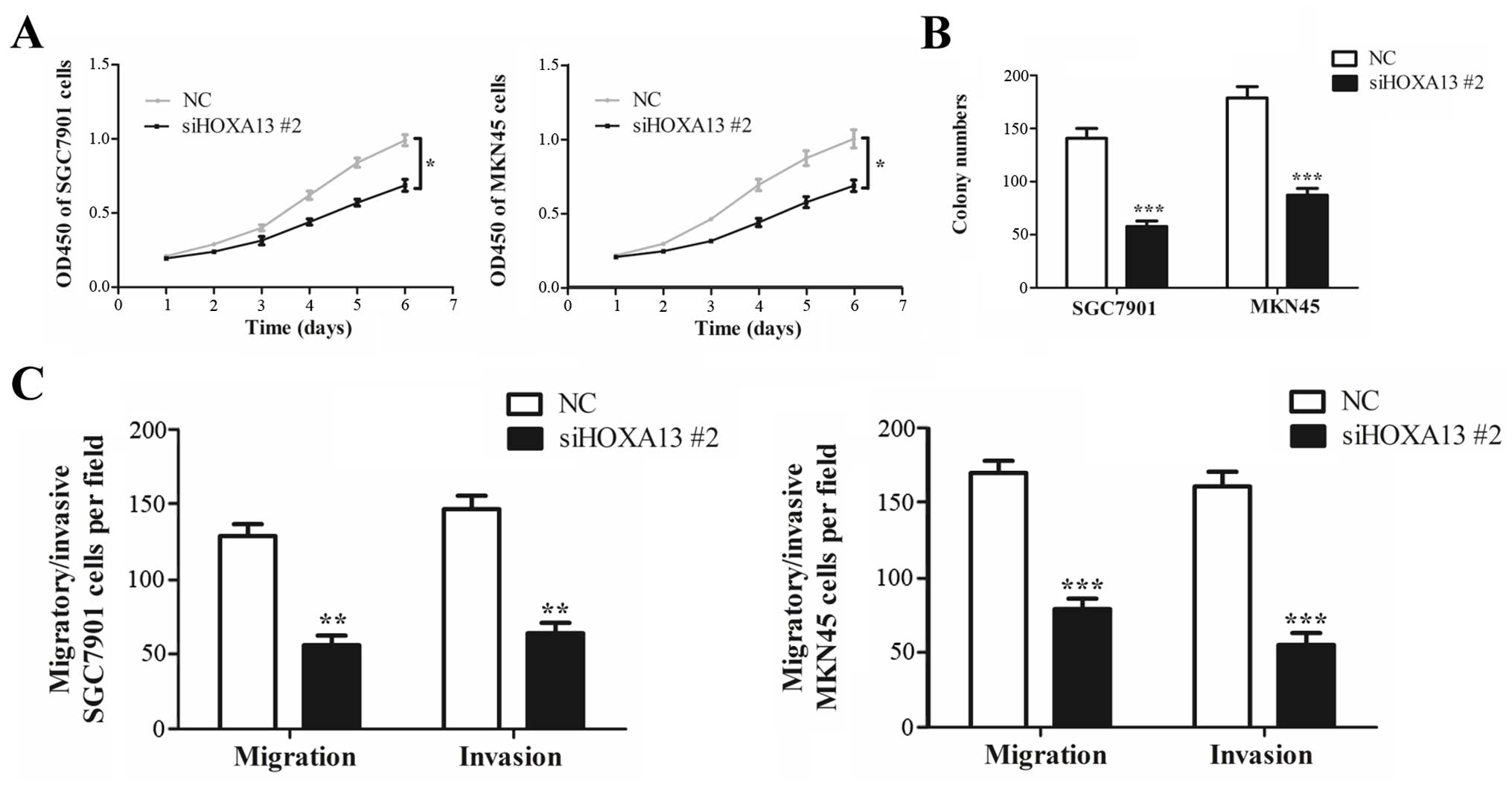
Knockdown of HOXA13 also inhibited cell growth
(Fig. 5A and B), migration and
invasion (Fig. 5C) in the SGC7901
and MKN45 cells, which resembled the inhibitory effects of HOTTIP
knockdown. These results indicate that HOXA13 was involved in
HOTTIP-induced malignant phenotypes of gastric cancer cells.
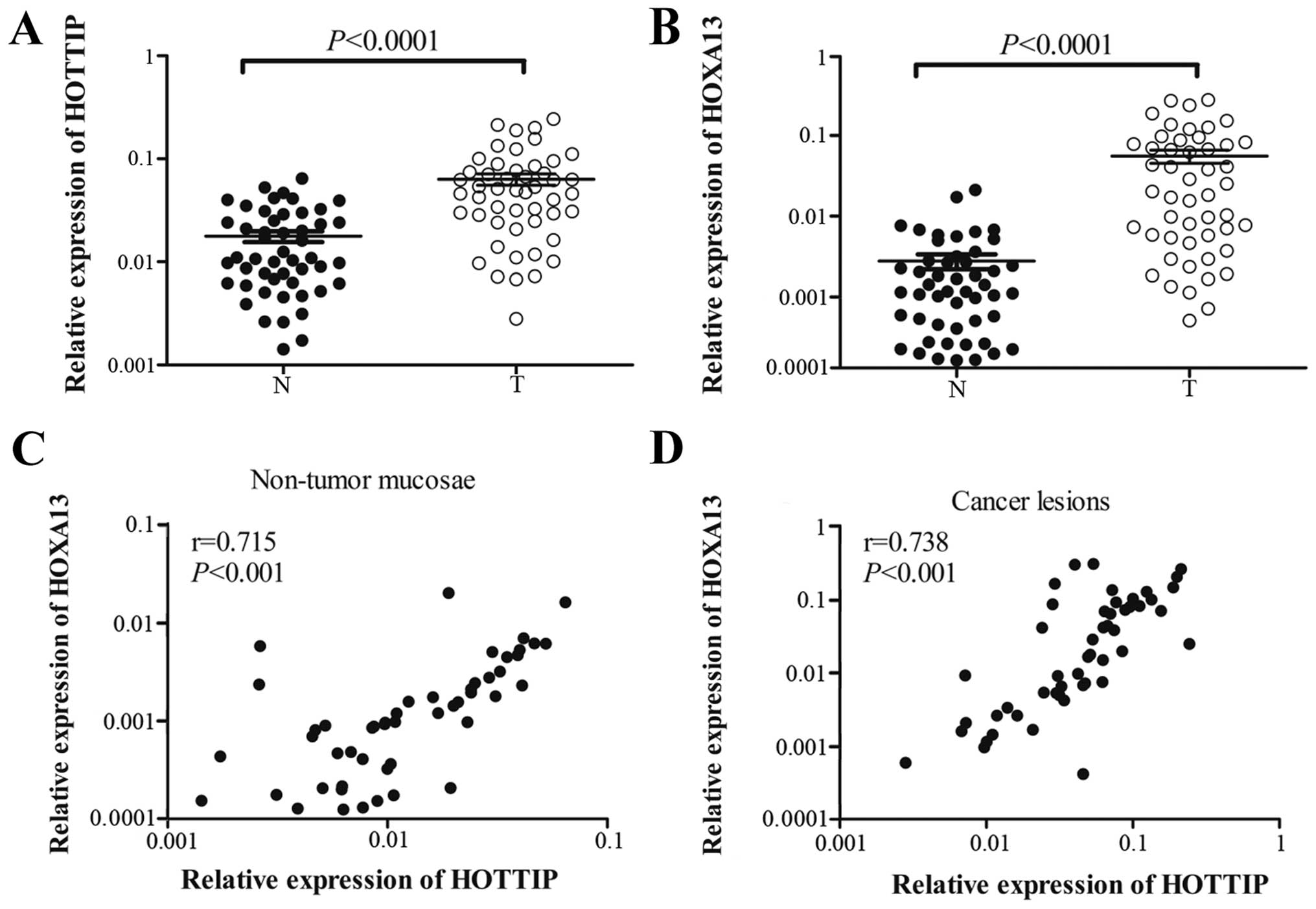
HOTTIP and HOXA13 are both upregulated in
gastric cancer
To further understand the relationship between
HOTTIP and HOXA13 in gastric cancer, we investigated HOTTIP and
HOXA13 expression levels in 50 pairs of primary gastric cancer
tissues and their counterpart non-tumorous tissues by qRT-PCR. The
results showed that HOTTIP and HOXA13 were both markedly
upregulated in the gastric cancer tissues when compared with these
levels in the non-tumorous tissues (Fig. 6A and B), which were consistent with
the expression patterns of HOTTIP and HOXA13 in the gastric cancer
cells. Correlations between the HOTTIP or HOXA13 expression levels
and clinicopathologic characteristics of gastric cancer are
summarized in Tables III or
IV, respectively. The data
revealed that expression levels of HOTTIP and HOXA13 were both
higher in gastric cancer which was poorly differentiated
(P<0.05), at advanced TNM stages (P<0.05) and showed lymph
node metastasis (P<0.01). Spearman analyses indicated that
HOTTIP and HOXA13 had a positive correlation both in non-tumor
mucosae (Fig. 6C) and cancer
lesions (Fig. 6D). These results
suggest that HOTTIP and HOXA13 are highly correlated and associated
with gastric cancer progression.
| Table IIIRelationship between HOTTIP
expression and clinicopathological parameters in the primary
gastric cancer cases. |
Table III
Relationship between HOTTIP
expression and clinicopathological parameters in the primary
gastric cancer cases.
| Variable | No. of cases | % | Relative expression
of HOTTIP | P-value |
|---|
| Age (years) | | | | 0.226 |
| ≥60 | 29 | 58 | 0.0716±0.0116 | |
| <60 | 21 | 42 | 0.0518±0.00995 | |
| Gender | | | | 0.384 |
| Male | 38 | 76 | 0.0672±0.0103 | |
| Female | 12 | 24 | 0.0508±0.00652 | |
| Tumor size
(cm) | | | | 0.362 |
| ≥5 | 28 | 56 | 0.0698±0.0111 | |
| <5 | 22 | 44 | 0.0550±0.0115 | |
| Degree of
differentiation | | | | 0.0250 |
| Well/moderate | 27 | 54 | 0.0469±0.00888 | |
| Poor | 23 | 46 | 0.0825±0.0130 | |
| TNM stage | | | | 0.00240 |
| I/II | 15 | 30 | 0.0274±0.00230 | |
| III/IV | 35 | 70 | 0.0787±0.0104 | |
| Lymph node
status | | | | 0.00950 |
| Metastasis | 38 | 76 | 0.0747±0.00971 | |
| No metastasis | 12 | 24 | 0.0272±0.00473 | |
| Table IVRelationship between HOXA13
expression and clinicopathological parameters in the primary
gastric cancer cases. |
Table IV
Relationship between HOXA13
expression and clinicopathological parameters in the primary
gastric cancer cases.
| Variable | No. of cases | % | Relative expression
of HOXA13 | P-value |
|---|
| Age (years) | | | | 0.520 |
| ≥60 | 29 | 58 | 0.0618±0.0152 | |
| <60 | 21 | 42 | 0.0476±0.0149 | |
| Gender | | | | 0.615 |
| Male | 38 | 76 | 0.0589±0.0131 | |
| Female | 12 | 24 | 0.0461±0.0175 | |
| Tumor size
(cm) | | | | 0.368 |
| ≥5 | 28 | 56 | 0.0645±0.0169 | |
| <5 | 22 | 44 | 0.0448±0.0116 | |
| Degree of
differentiation | | | | 0.0178 |
| Well/moderate | 27 | 54 | 0.0327±0.00832 | |
| Poor | 23 | 46 | 0.0831±0.0199 | |
| TNM stage | | | | 0.0192 |
| I/II | 15 | 30 | 0.0179±0.00674 | |
| III/IV | 35 | 70 | 0.0721±0.0143 | |
| Lymph node
status | | | | 0.00550 |
| Metastasis | 38 | 76 | 0.0722±0.0131 | |
| No metastasis | 12 | 24 |
0.00409±0.000987 | |
Discussion
In the present study, we found that both HOTTIP and
HOXA13 were upregulated in gastric cancer tissues compared with
levels in their counterpart non-tumorous tissues. In addition, the
expression levels of HOTTIP and HOXA13 were associated with poor
differentiation, advanced TNM stages and lymph node metastasis.
Moreover, HOTTIP and HOXA13 were highly correlated both in
non-tumor mucosae and cancer lesions. Downregulation of HOTTIP
inhibited cell growth and invasion. In addition, HOXA13 was
involved in HOTTIP-induced malignant phenotypes of gastric cancer
cells.
lncRNAs associated with human HOX gene loci
have been widely studied in recent years (21,28,34–36).
By characterizing the transcriptional landscape of the four human
HOX loci, researchers have identified 231 HOX lncRNAs
(21). HOTAIR, which was first
described in fibroblasts, was found to be located in the
HOXC cluster but regulated HOXD cluster genes
(21). HOTAIR was also found to
serve as a scaffold protein by binding polycomb repressive complex
2 (PRC) with its 5′ domain and the LSD1/CoREST/REST complex with
the 3′ domain (37). Unlike HOTAIR,
HOTTIP enhanced expression of neighboring HOXA genes
particularly HOXA13 (28).
Considering the vital role of HOX genes in development and
differentiation and their dysregulation-caused tumorigenesis and
tumor progression (38–42), it is important to understand the
mechanism of HOTTIP in the regulation of HOX gene
expression.
Upregulation of HOTTIP and HOXA13 has been reported
in various studies. HOTTIP and HOXA13 were both upregulated and
highly correlated in hepatocellular carcinoma (29) and pancreatic cancer (31,32). A
previous study demonstrated that HOTTIP was upregulated not only in
hepatocellular carcinoma tissues, but also in preneoplastic
diseases. However, the HOXA13 expression level was specifically
increased in hepatocellular carcinoma, indicating that upregulation
of HOTTIP preceded that of HOXA13 in hepatocellular carcinogenesis
during disease onset (29). HOTTIP
was also found to be upregulated in lung cancer (32) and tongue squamous cell carcinoma
(33), and involved in the tumor
progression in pancreatic cancer (30). The expression level of HOXA13 was
also increased and associated with tumor progression in
hepatocellular carcinoma (29),
pancreatic cancer (30), esophageal
squamous cell carcinoma (43) and
glioblastoma multiforme (44). A
recent study found that HOXA13 expression was higher in cancerous
tissues compared with that in their neighboring non-cancerous
tissues. Moreover, a higher expression level of HOXA13 was
significantly correlated with T and M stages, advanced UICC stage
and histological differentiation in gastric cancer based on
immunohistochemistry findings (45). In the present study, we also found
that HOTTIP and HOXA13 were upregulated in gastric cancer tissues
compared with level in their non-tumorous tissues. In addition, the
increase in the expression level of these two genes was correlated
with cancer tissue poor differentiation, advanced TNM stages and
lymph node metastasis. HOTTIP and HOXA13 were positively associated
in both non-tumor mucosae and cancer lesions. Our findings suggest
that HOTTIP and HOXA13 are likely involved in the tumorigenesis and
progression of gastric cancer.
Although we identified upregulation of HOTTIP and
HOXA13 in gastric cancer, the roles of HOTTIP and HOXA13 in gastric
cancer have never been fully understood. Downregulation of HOTTIP
and HOXA13 has been reported to inhibit cell proliferation in liver
cancer-derived cells (29). HOTTIP
and HOXA13 promoted cell proliferation, migration and invasion in
pancreatic cancer (30,31). Moreover, HOTTIP regulated the
expression of HOXA13 in hepatocellular carcinoma (29) and pancreatic cancer (30). However, Cheng et al showed
that HOTTIP regulated HOX genes including HOXA10, HOXA11,
HOXA9 and HOXA1, but not HOXA13 (31). HOXA13 was found to promote cell
growth of esophageal squamous cancer cells in vitro and
in vivo (43). HOXA13 also
promoted cell invasion in vitro and tumor growth in
vivo in glioblastoma multiforme (44). In the present study, we firstly
identified that HOTTIP and HOXA13 both promoted cell growth and
invasiveness in gastric cancer cells. In addition, downregulation
of HOTTIP led to decreased HOXA13 expression in gastric cancer
cells. The roles of HOTTIP and HOXA13 in gastric cancer cells in
vivo warrant further investigation. Taken together, these data
indicate that HOTTIP functions as an oncogene by regulating HOXA13
expression in gastric cancer.
In conclusion, our results showed that HOTTIP and
HOXA13 were upregulated and associated with poor differentiation,
advanced TNM stages and lymph node metastasis in gastric cancer.
HOTTIP and HOXA13 were highly correlated in both non-tumor mucosae
and cancer lesions. Downregulation of HOTTIP inhibited gastric
cancer cell growth and invasiveness through the regulation of
HOXA13. These results suggest that the molecular axis of HOTTIP and
HOXA13 contributes to gastric cancer progression. Our finding
provides a potential novel therapeutic target for gastric cancer
treatment.
Acknowledgments
The present study was supported in part by the
National Natural Science Foundation of China (no. 81472245).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takahashi T, Saikawa Y and Kitagawa Y:
Gastric cancer: Current status of diagnosis and treatment. Cancers.
5:48–63. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ajani JA, Bentrem DJ, Besh S, D'Amico TA,
Das P, Denlinger C, Fakih MG, Fuchs CS, Gerdes H, Glasgow RE, et al
National Comprehensive Cancer Network: Gastric cancer, version
2.2013: Featured updates to the NCCN Guidelines. J Natl Compr Canc
Netw. 11:531–546. 2013.PubMed/NCBI
|
|
4
|
Zheng L, Wang L, Ajani J and Xie K:
Molecular basis of gastric cancer development and progression.
Gastric Cancer. 7:61–77. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tamura G, Yin J, Wang S, Fleisher AS, Zou
T, Abraham JM, Kong D, Smolinski KN, Wilson KT, James SP, et al:
E-Cadherin gene promoter hypermethylation in primary human gastric
carcinomas. J Natl Cancer Inst. 92:569–573. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maekita T, Nakazawa K, Mihara M, Nakajima
T, Yanaoka K, Iguchi M, Arii K, Kaneda A, Tsukamoto T, Tatematsu M,
et al: High levels of aberrant DNA methylation in Helicobacter
pylori- infected gastric mucosae and its possible association with
gastric cancer risk. Clin Cancer Res. 12:989–995. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Niwa T, Tsukamoto T, Toyoda T, Mori A,
Tanaka H, Maekita T, Ichinose M, Tatematsu M and Ushijima T:
Inflammatory processes triggered by Helicobacter pylori infection
cause aberrant DNA methylation in gastric epithelial cells. Cancer
Res. 70:1430–1440. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
El-Omar EM, Carrington M, Chow WH, McColl
KE, Bream JH, Young HA, Herrera J, Lissowska J, Yuan CC, Rothman N,
et al: Interleukin-1 polymorphisms associated with increased risk
of gastric cancer. Nature. 404:398–402. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He C, Tu H, Sun L, Xu Q, Gong Y, Jing J,
Dong N and Yuan Y: SNP interactions of Helicobacter pylori-related
host genes PGC, PTPN11, IL1B, and TLR4 in susceptibility to gastric
carcinogenesis. Oncotarget. 6:19017–19026. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang TT, Ping YH, Wang AM, Ke CC, Fang
WL, Huang KH, Lee HC, Chi CW and Yeh TS: The reciprocal regulation
loop of Notch2 pathway and miR-23b in controlling gastric
carcinogenesis. Oncotarget. 6:18012–18026. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen J, Xiao Z, Wu WK, Wang MH, To KF,
Chen Y, Yang W, Li MS, Shin VY, Tong JH, et al: Epigenetic
silencing of miR-490-3p reactivates the chromatin remodeler SMARCD1
to promote Helicobacter pylori-induced gastric carcinogenesis.
Cancer Res. 75:754–765. 2015. View Article : Google Scholar
|
|
12
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Flynn RA and Chang HY: Long noncoding RNAs
in cell-fate programming and reprogramming. Cell Stem Cell.
14:752–761. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huarte M and Rinn JL: Large non-coding
RNAs: Missing links in cancer? Hum Mol Genet. 19:R152–R161. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Prensner JR, Iyer MK, Balbin OA,
Dhanasekaran SM, Cao Q, Brenner JC, Laxman B, Asangani IA, Grasso
CS, Kominsky HD, et al: Transcriptome sequencing across a prostate
cancer cohort identifies PCAT-1, an unannotated lincRNA implicated
in disease progression. Nat Biotechnol. 29:742–749. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Braconi C, Valeri N, Kogure T, Gasparini
P, Huang N, Nuovo GJ, Terracciano L, Croce CM and Patel T:
Expression and functional role of a transcribed noncoding RNA with
an ultraconserved element in hepatocellular carcinoma. Proc Natl
Acad Sci USA. 108:786–791. 2011. View Article : Google Scholar :
|
|
17
|
Pantoja C, de Los Ríos L, Matheu A,
Antequera F and Serrano M: Inactivation of imprinted genes induced
by cellular stress and tumorigenesis. Cancer Res. 65:26–33.
2005.PubMed/NCBI
|
|
18
|
Yap KL, Li S, Muñoz-Cabello AM, Raguz S,
Zeng L, Mujtaba S, Gil J, Walsh MJ and Zhou MM: Molecular interplay
of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by
polycomb CBX7 in transcriptional silencing of INK4a. Mol Cell.
38:662–674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brown CJ, Ballabio A, Rupert JL,
Lafreniere RG, Grompe M, Tonlorenzi R and Willard HF: A gene from
the region of the human X inactivation centre is expressed
exclusively from the inactive X chromosome. Nature. 349:38–44.
1991. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bartolomei MS, Zemel S and Tilghman SM:
Parental imprinting of the mouse H19 gene. Nature. 351:153–155.
1991. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E, et
al: Functional demarcation of active and silent chromatin domains
in human HOX loci by noncoding RNAs. Cell. 129:1311–1323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hou P, Zhao Y, Li Z, Yao R, Ma M, Gao Y,
Zhao L, Zhang Y, Huang B and Lu J: LincRNA-ROR induces
epithelial-to-mesen-chymal transition and contributes to breast
cancer tumorigenesis and metastasis. Cell Death Dis. 5:e12872014.
View Article : Google Scholar
|
|
23
|
Zhang H, Diab A, Fan H, Mani SK, Hullinger
R, Merle P and Andrisani O: PLK1 and HOTAIR accelerate proteasomal
degradation of SUZ12 and ZNF198 during hepatitis B virus-induced
liver carcinogenesis. Cancer Res. 75:2363–2374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan J, Xing Y, Wen X, Jia R, Ni H, He J,
Ding X, Pan H, Qian G, Ge S, et al: Long non-coding RNA ROR decoys
gene-specific histone methylation to promote tumorigenesis. Genome
Biol. 16:1392015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Barnhill LM, Williams RT, Cohen O, Kim Y,
Batova A, Mielke JA, Messer K, Pu M, Bao L, Yu AL, et al: High
expression of CAI2, a 9p21-embedded long noncoding RNA, contributes
to advanced-stage neuroblastoma. Cancer Res. 74:3753–3763. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hu Y, Wang J, Qian J, Kong X, Tang J, Wang
Y, Chen H, Hong J, Zou W, Chen Y, et al: Long noncoding RNA GAPLINC
regulates CD44-dependent cell invasiveness and associates with poor
prognosis of gastric cancer. Cancer Res. 74:6890–6902. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Gejman R, Mahta A, Zhong Y, Rice
KA, Zhou Y, Cheunsuchon P, Louis DN and Klibanski A: Maternally
expressed gene 3, an imprinted noncoding RNA gene, is associated
with meningioma pathogenesis and progression. Cancer Res.
70:2350–2358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang KC, Yang YW, Liu B, Sanyal A,
Corces-Zimmerman R, Chen Y, Lajoie BR, Protacio A, Flynn RA, Gupta
RA, et al: A long noncoding RNA maintains active chromatin to
coordinate homeotic gene expression. Nature. 472:120–124. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Quagliata L, Matter MS, Piscuoglio S,
Arabi L, Ruiz C, Procino A, Kovac M, Moretti F, Makowska Z,
Boldanova T, et al: Long noncoding RNA HOTTIP/HOXA13 expression is
associated with disease progression and predicts outcome in
hepatocellular carcinoma patients. Hepatology. 59:911–923. 2014.
View Article : Google Scholar :
|
|
30
|
Li Z, Zhao X, Zhou Y, Liu Y, Zhou Q, Ye H,
Wang Y, Zeng J, Song Y, Gao W, et al: The long non-coding RNA
HOTTIP promotes progression and gemcitabine resistance by
regulating HOXA13 in pancreatic cancer. J Transl Med. 13:842015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cheng Y, Jutooru I, Chadalapaka G, Corton
JC and Safe S: The long non-coding RNA HOTTIP enhances pancreatic
cancer cell proliferation, survival and migration. Oncotarget.
6:10840–10852. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Deng HP, Chen L, Fan T, Zhang B, Xu Y and
Geng Q: Long non-coding RNA HOTTIP promotes tumor growth and
inhibits cell apoptosis in lung cancer. Cell Mol Biol. 61:34–40.
2015.PubMed/NCBI
|
|
33
|
Zhang H, Zhao L, Wang YX, Xi M, Liu SL and
Luo LL: Long non-coding RNA HOTTIP is correlated with progression
and prognosis in tongue squamous cell carcinoma. Tumour Biol.
36:8805–8809. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maamar H, Cabili MN, Rinn J and Raj A:
linc-HOXA1 is a noncoding RNA that represses Hoxa1 transcription in
cis. Genes Dev. 27:1260–1271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang EB, Yin DD, Sun M, Kong R, Liu XH,
You LH, Han L, Xia R, Wang KM, Yang JS, et al: P53-regulated long
non-coding RNA TUG1 affects cell proliferation in human non-small
cell lung cancer, partly through epigenetically regulating HOXB7
expression. Cell Death Dis. 5:e12432014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang X, Lian Z, Padden C, Gerstein MB,
Rozowsky J, Snyder M, Gingeras TR, Kapranov P, Weissman SM and
Newburger PE: A myelopoiesis-associated regulatory intergenic
noncoding RNA transcript within the human HOXA cluster. Blood.
113:2526–2534. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liao WT, Jiang D, Yuan J, Cui YM, Shi XW,
Chen CM, Bian XW, Deng YJ and Ding YQ: HOXB7 as a prognostic factor
and mediator of colorectal cancer progression. Clin Cancer Res.
17:3569–3578. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cantile M, Pettinato G, Procino A,
Feliciello I, Cindolo L and Cillo C: In vivo expression of the
whole HOX gene network in human breast cancer. Eur J Cancer.
39:257–264. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Waltregny D, Alami Y, Clausse N, de Leval
J and Castronovo V: Overexpression of the homeobox gene HOXC 8 in
human prostate cancer correlates with loss of tumor
differentiation. Prostate. 50:162–169. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Costa BM, Smith JS, Chen Y, Chen J,
Phillips HS, Aldape KD, Zardo G, Nigro J, James CD, Fridlyand J, et
al: Reversing HOXA9 oncogene activation by PI3K inhibition:
Epigenetic mechanism and prognostic significance in human
glioblastoma. Cancer Res. 70:453–462. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Buske C, Feuring-Buske M, Abramovich C,
Spiekermann K, Eaves CJ, Coulombel L, Sauvageau G, Hogge DE and
Humphries RK: Deregulated expression of HOXB4 enhances the
primitive growth activity of human hematopoietic cells. Blood.
100:862–868. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gu ZD, Shen LY, Wang H, Chen XM, Li Y,
Ning T and Chen KN: HOXA13 promotes cancer cell growth and predicts
poor survival of patients with esophageal squamous cell carcinoma.
Cancer Res. 69:4969–4973. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Duan R, Han L, Wang Q, Wei J, Chen L,
Zhang J, Kang C and Wang L: HOXA13 is a potential GBM diagnostic
marker and promotes glioma invasion by activating the Wnt and TGF-β
pathways. Oncotarget. 6:27778–27793. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Han Y, Tu WW, Wen YG, Li DP, Qiu GQ, Tang
HM, Peng ZH and Zhou CZ: Identification and validation that
up-expression of HOXA13 is a novel independent prognostic marker of
a worse outcome in gastric cancer based on immunohistochemistry.
Med Oncol. 30:5642013. View Article : Google Scholar : PubMed/NCBI
|