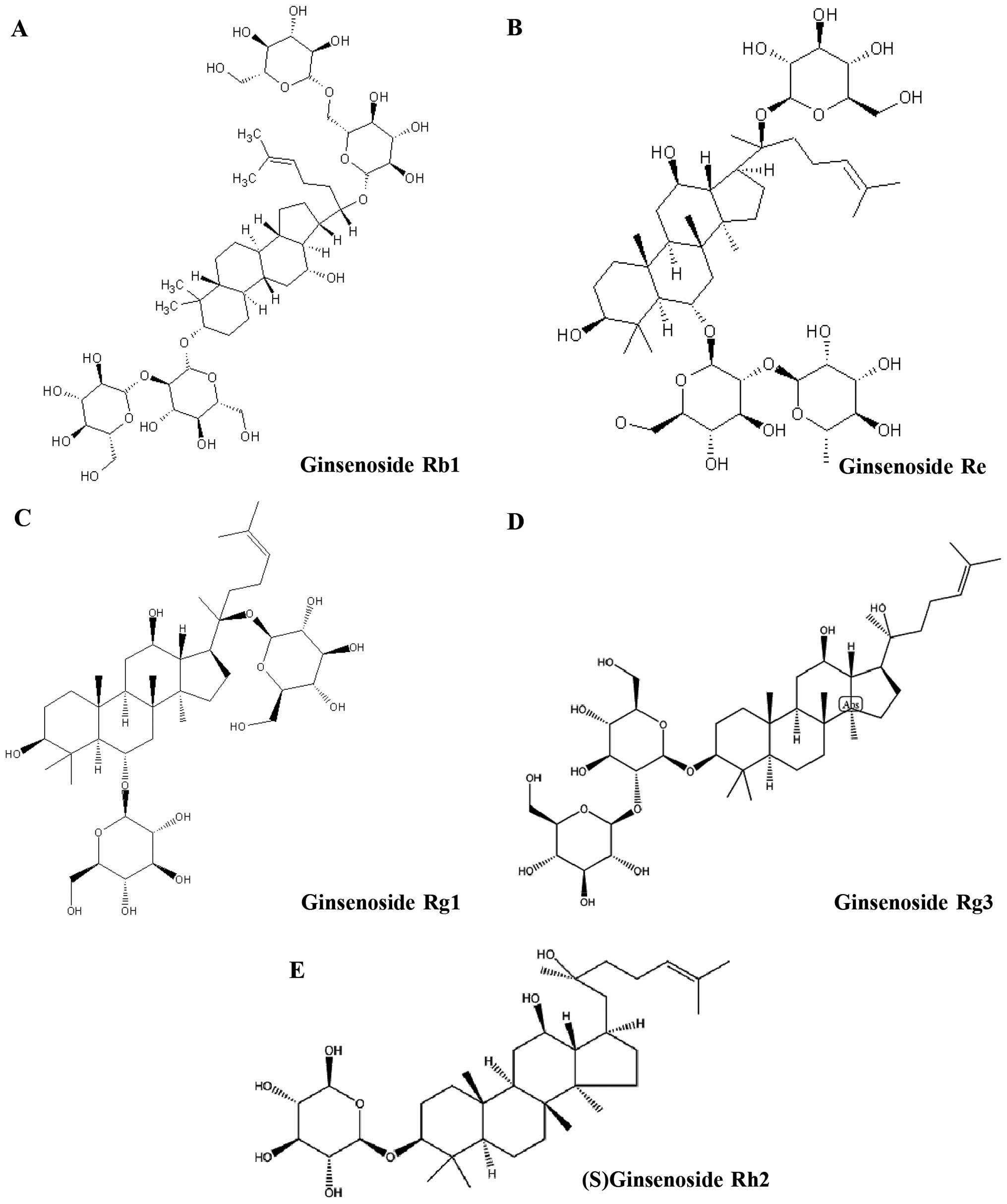
Introduction
Leukemia is a type of hematological malignancy of
the blood and the bone marrow characterized by an abnormal increase
in immature hemamegba called 'blasts' (1). Chemotherapy drugs kill cancer cells as
well as normal cells, leading to significant side effects (2). As a step in overcoming this limitation
in chemotherapy, medicines prepared from natural traditional
Chinese medicine (TCM) are currently being considered as anticancer
agents (3,4). In the past decades, a series of
studies have demonstrated that many extracted products from natural
plants can kill cancer cells including leukemia cells in
vitro (5).
Ginsenosides (total saponins of Panax
ginseng, TSPG), are important components extracted from the
root of Panax ginseng. TSPG as an extract of a medicinal
plant is used in a wide range of ailments and has been reported to
have various potent biological functions, including improvement of
physical and mental capacity, reduction of fatigue, regulation of
the central nervous system and antitumor effects (6–10).
TSPG includes many monomers, such as ginsenoside Rb1 (Rb1),
ginsenoside Re (Re), ginsenoside Rg1 (Rg1), ginsenoside Rg3 (Rg3),
20(S)-ginsenoside Rh2 [(S)Rh2] and compound K. TSPG
shows enhancement of proliferation and differentiation of bone
marrow cells (9–12). To date, TSPG and its monomers have
not been compared in regard to their effects on cell viability,
thus research on the effects of TSPG and other monomers on
proliferation inhibition in KG-1a cells is warranted.
(S)Rh2 belongs to the protopanaxadiol family
and has attracted attention in the research concerning
chemoprevention and chemotherapy (13–15).
Recently, researchers have found that (S)Rh2 exhibits
anticancer functions and inhibits the growth and induces the
apoptosis of various cancer cell lines, including leukemia cells
(10,16–20).
Evidence has proven that (S)Rh2 displays a marked inhibitory
effect on leukemia cells, although its specific molecular mechanism
is not well understood. Moreover, the Wnt pathway is apparently
related to tumor progression. Therefore, we hypothesized that there
may be some correlation between (S)Rh2 and the Wnt
pathway.
The Wnt signaling pathway plays an important role in
embryogenesis and shows little or no activity in fully
differentiated cells (21).
Self-renewal, proliferation and differentiation are tightly
controlled during normal hematopoiesis to ensure lifelong
hematopoietic stem cell homeostasis and blood production (22). Deregulation of these processes
results in hematologic dysplasia, deficiency, myeloproliferation or
leukemia. The unnatural activity of the Wnt pathway is involved in
the development of several types of cancer (23). For acute myeloid leukemia (AML), it
has been confirmed that Wnt genes are overexpressed and activated
(24). The importance of the Wnt
pathway for AML cell survival has been demonstrated by the fact
that inhibition of the pathway results in the decline of cell
growth (25). (S)Rh2 can
obviously inhibit proliferation, differentiation and promote
apoptosis of leukemia cells, yet its mechanisms are not fully
clear. For the purpose of exploiting new strategies for tumor
treatment, it is important to illuminate the concrete mechanism of
(S)Rh2 in regards to its effect on leukemia, which is vital
to clinical experimental research.
Materials and methods
Reagents and cell culture
TSPG, Rb1, Re, Rg1, Rg3 and (S)Rh2 were
purchased from Nanjing Ze Lang Pharmaceutical Technology (Nanjing,
China). Structures of Rb1, Re, Rg1, Rg3 and (S)Rh2 are shown
(Fig. 1). These agents were
dissolved in dimethyl sulfoxide (DMSO) at a concentration of 100 mM
and stored at −20°C. Human leukemia KG-1a cells were cryo-preserved
in our laboratory, and cultured in Roswell Park Memorial Institute
(RPMI)-1640 medium containing 10% fetal bovine serum (HyClone,
Waltham, MA, USA) at 37°C in air 5% CO2 incubator at
constant humidity, and maintained for logarithmic growth by
passaging cells every 48 h.
CCK-8 assay
The CCK-8 assay was employed for determination of
cell viability. Briefly, 1.0×104 cells/well were planted
in 96-well plates and cultured for different times. At the end of
the time, 10 µl CCK-8 working solution was added to each
well and then cultured at 37°C for 2 h. Then plates were detected
at 450 nm on a spectrophotometric plate reader. The drug
concentration resulting in 50% inhibition of growth
(IC50) was determined.
Transmission electron microscopic
observation
For the ultra-structural characteristic observation
assay, cells were harvested and fixed at 1.0×107 with
2.5% glutaraldehyde for 6 h at 4°C and then with 1% osmium
tetroxide for 2 h prior to dehydration with ethyl alcohol.
Ultra-thin sections (60 nm) were prepared and placed on grids,
stained with 2% uranyl acetate solution and 0.2% lead citrate in
0.1 M NaOH. The cells were observed by a H-600 transmission
electron microscope (Hitachi, Japan).
Cell cycle assay
The KG-1a cells were plated at a concentration of
2.0×105 cells/ml and incubated with (S)Rh2 or
TSPG for 48 h. The cells were collected by centrifugation at 1,000
× g for 3 min, added to 70% ethanol and then washed once with PBS.
The cells were then resuspended in 1 ml of PBS containing 2.5
µg/ml ribonuclease and 50 µg/ml propidium iodide
(Beyotime Institute of Biotechnology, Shanghai, China), incubated
in the dark for 30 min at room temperature and analyzed using flow
cytometry (FCM).
Cell apoptosis assay
Briefly, for the Annexin V assay, the cells were
seeded at a concentration of 2.0×105 cells/ml and
incubated with (S)Rh2 or TSPG for 48 h. Samples were
prepared based on the instructions provided together with the
Annexin V apoptosis kit. Briefly, after treatment for the indicated
times, the cells were collected and washed twice with binding
buffer containing 10 mM HEPES, pH 7.4, 140 mM NaCl, 2.5 mM
CaCl2. Next, 1.0×105 cells were resuspended
in 100 µl of binding buffer, and then 5 µl of Annexin
V-FITC and 10 µl of propidium iodide (50 µg/ml, stock
concentration) were added to the cell suspension. After gently
mixing, the cells were incubated for 15 min at room temperature,
and then 400 µl of binding buffer was added to get the
sample ready. Quantification of cell death was analyzed with a BD
FACScan.
Immunofluorescence staining
In brief, KG-1a cells cultured in 6-well plates were
treated with DMSO or (S)Rh2 (60 µM) for 24 h. The
cells were washed with ice-cold phosphate buffered saline (PBS)
three times and fixed with 4% paraformaldehyde. The cell membrane
was ruptured by 0.3% tristone, and closed with mountain goat serum
(HyClone, Waltham, MA, USA). Antibodies against β-catenin (1:100),
TCF4 (1:100), cyclin D1 (1:100), NF-κBp65 (1:100) (Santa Cruz
Biotechnology Inc., Santa Cruz, USA) were added respectively.
Anti-rabbit secondary fluorescent antibody was added and incubated
overnight for 1 h, and stained with PI (Beyotime Institute of
Biotechnology, Shanghai, China). Then, 50% glycerol was used for
mounting, and imaging was carried out under a fluorescence
microscope (Olympus, Tokyo, Japan).
Quantitative real-time PCR (qRT-PCR)
Following treatment, the cells were harvested and
total RNA was immediately extracted using TRIzol reagent
(Invitrogen) according to the manufacturer's instructions. For
expression analysis of β-catenin, TCF4, cyclin D1 and NF-κBp65
genes, 2 µg of total RNA was used to synthesize first-strand
DNA with reverse transcriptase according to the manufacturer's
instructions (Promega, Madison, WI, USA). Quantitative RT-PCR
(qRT-PCR) was performed with a Green PCR Master Mix kit (Shanghai,
Shine Co, China). Briefly, one microliter of first-strand cDNA and
gene-specific primers were used along with Hotstart Fluo-PCR Mix in
a 20-µl reaction under the following conditions:
pre-denaturation at 95°C for 5 min, 35 cycles of denaturation at
95°C for 10 sec, annealing at 57°C for 15 sec, and extension at
72°C for 20 sec. Each sample was performed in triplicate and was
quantified based on the formula 2−ΔCt. The primer pairs
for qRT-PCR are listed in Table
I.
 | Table IPrimer pairs for qRT-PCR. |
Table I
Primer pairs for qRT-PCR.
| Gene name | Sequence
(5′-3′) |
|---|
| β-catenin | F:
CAAAGCCTCAGGTCATAAACA |
| R:
GTGGGATGGTGGGTGTAAGA |
| TCF4 | F:
TGAGGTCCTGATGCGGTTGG |
| R:
TCGCCTTTGTTCTCCTTGATGC |
| Cyclin D1 | F:
AGGCTGGCTTCATCCACT |
| R:
CACCAAGGGTTAATTCTTCA |
| NF-κBp65 | F:
CCCCACGAGCTTGTAGGAAAG |
| R:
CCAGGTTCTGGAAACTGTGGAT |
| GAPDH | F:
CATCAAGAAGGTGGTGAAGCA |
| R:
CGTCAAAGGTGGAGGAGTGG |
Western blotting
Following treatment with (S)Rh2 for 48 h, the
cells were lysed in ice-cold RIPA lysis buffer. The lysates were
centrifuged at 15,000 × g for 10 min at 4°C to obtain the proteins.
The protein content of the cell extracts was determined by
bicinchonininc acid (BCA). A total of 30–40 µg of protein
was electrophoresed on 10–15% SDS-PAGE gels and transferred to PDVF
membranes. The membranes were blocked, incubated with the primary
Abs at the appropriate concentration, and subsequently incubated
with horseradish peroxidase-conjugated goat anti-rabbit IgG or goat
anti-mouse IgG (1:2,000 dilutions). Labeled bands were detected by
Immun-Star TMHRP Chemiluminescent kit, and images were captured and
the intensity of the bands was quantified by the Bio-Rad VersaDoc™
image system (Bio-Rad, Regents Park, Australia).
Chromatin immunoprecipitation (ChIP)
assays
For the ChIP assay, KG-1a cells were treated with
DMSO (control), or 60 µM (S)Rh2 for 48 h. Treated
cells were cross-linked with 1% formaldehyde for 15 min at room
temperature. The cross-linked cells were then resuspended in 0.3 ml
of lysis buffer (50 mM Tris-HCl, pH 8.1/1% SDS/10 mM EDTA protease
inhibitors) and subjected three times for 10 sec followed by
centrifugation for 10 min. The average size of the sheared fragment
was expected to be 300–1,000 bp. Immunoprecipitates were collected
three times with 1% SDS/0.1 M NaHCO3. Eluates were
pooled and heated at 65°C for 6 h to reverse the formaldehyde
cross-linking. DNA fragments were purified with a QIAquick spin kit
(Qiagen, Chatsworth, CA, USA). For PCR, 1 µl from a 50
µl DNA extraction and 38 cycles of amplification were used
with the following promoter-specific primers: c-myc forward,
5-GCTTGGCGGGAAAAAGAA GGG-3 and reverse, 5-AGAGCTGCCTTCTTAGGTCG-3;
cyclin D1 forward, 5-GTTCCTGGAAGGGCGACTAA-3 and reverse,
5-GGGGTGGGATCTGAGATTTG-3.
Statistical analysis
The intensity of the immunoreactive band was
determined by a densitometer (Bio-Rad, Hercules, CA, USA). Data are
expressed as the mean ± SEM of three independent experiments, and
one-way ANOVA analysis was conducted using the statistical software
SPSS 22.0. Each treatment group was compared with the control group
with Dunnett's t-test, and P-value <0.05 was considered to
indicate a statistically significant result.
Results
TSPG, Rb1, Re, Rg1, Rg3 and (S)Rh2
inhibit the growth of KG-1a cells
We assessed the effects of TSPG, Rb1, Re, Rg1, Rg3
and (S)Rh2 on the cell viability of KG-1a cells using Cell
Counting Kit-8 assays. (S)Rh2 was found to have a marked
inhibitory effect on KG-1a cells at a lower concentration, compared
with Rb1, Re, Rg1 and Rg3. (S)Rh2 had the lowest
IC50 values (in the mid-µM range) compared with
the other ginsenosides tested. Rb1, Re, Rg1, Rg3 did not
significantly decrease cell viability even at the high-µM
range. These results showed that (S)Rh2 decreased the
viability of the KG-1a cells. TSPG had a lesser effect on viability
than the other members of the ginsenosides in the KG-1a cells
(Fig. 2).
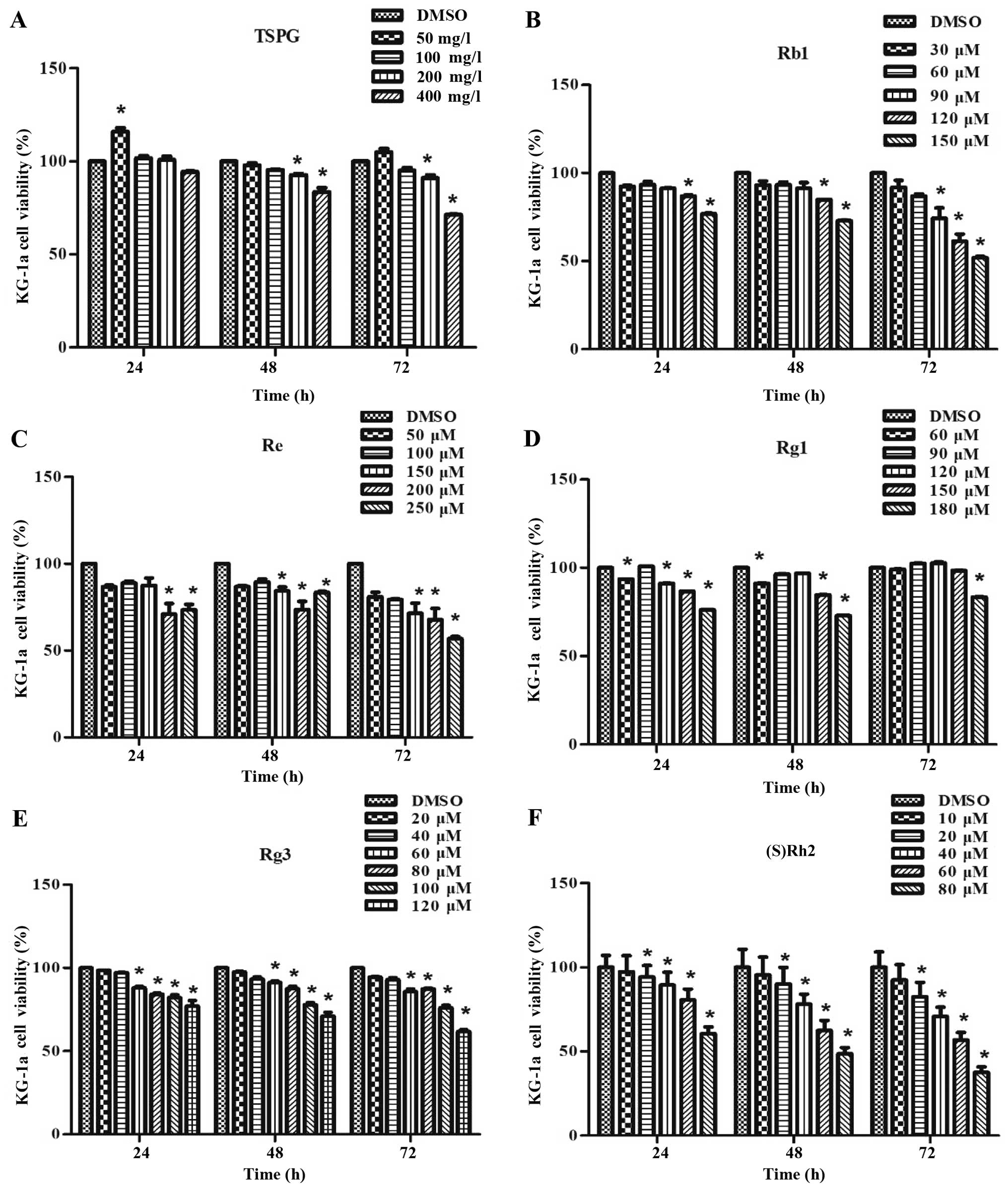 | Figure 2Effect of TSPG, Rb1, Re, Rg1, Rg3 and
(S)Rh2 on the cell viability of KG-1a cells as determined by
the CCK-8 assay. KG-1a cells were incubated with or without various
concentrations of TSPG, Rb1, Re, Rg1, Rg3 and (S)Rh2 for 24,
48 and 72 h. Data are presented as mean ± SEM. (n=3) for each
group. *p<0.05, a significant difference compared with the
control by one-way ANOVA. Similar results were observed in
replicate experiments. |
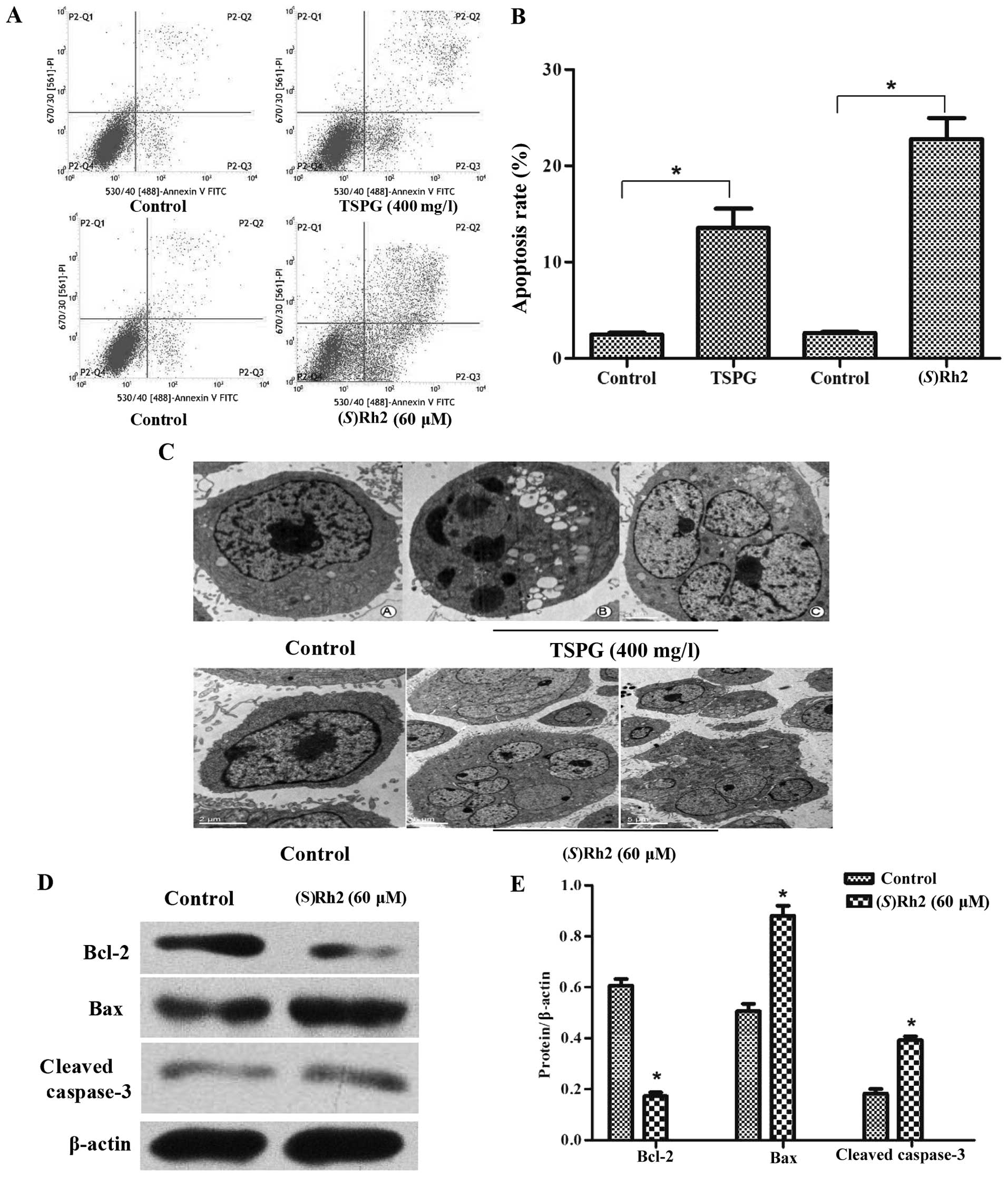
TSPG and (S)Rh2 induce the apoptosis of
KG-1a cells
Further experiments were performed using KG-1a cells
to evaluate the effect of TSPG and (S)Rh2 on apoptosis and
the mechanism involved. To determine whether the cell growth
inhibitory effect of TSPG and (S)Rh2 is associated with the
induction of cell apoptosis, we used Annexin V/7-AAD double
staining. Annexin V+ and 7-AAD− cells were
designated as early apoptotic and Annexin V+ and
7-AAD+ cells were designated as necrotic. A higher
number of apoptotic cells was observed in the TSPG (400 mg/l) and
(S)Rh2 (60 µM) treatment groups than that in the
control group (Fig. 3A). Analysis
of the cell population revealed distinct sets of populations.
Following treatment with TSPG (400 mg/l) and (S)Rh2 (60
µM) for 24 h, the percentage of Annexin V+ and
7-AAD− apoptotic cells increased gradually from 3.10 to
8.63 and 19.53%, while the percentage of
FITC+/7-AAD+ necrotic cells increased from
2.17 to 6.68 and 16.52% (Fig. 3B).
These results revealed that (S)Rh2 was significantly more
potent at inducing apoptosis than TSPG. To further characterize the
TSPG- and (S)Rh2-induced apoptosis, we observed
ultra-microstructures in the KG-1a cells following treatment with
TSPG (400 mg/l) or (S)Rh2 (60 µM) for 48 h. These
cells had different levels of apoptosis, chromatin occured
margination, nucleoli decreased or disappeared, and apoptotic
bodies increased (Fig. 3C). In
addition to its anti-proliferative effects, we also noted that
(S)Rh2 caused an increase in apoptosis in the KG-1a cells.
Thus, we assessed the expression of several apoptosis-related
proteins, including Bcl-2, Bax and cleaved caspase-3. The effect on
apoptosis was consistent with the cell apoptosis assay.
(S)Rh2 increased the expression of Bcl-2 and cleaved
caspase-3 (Fig. 3D and E). Cleaved
caspase-3 was activated. We demonstrated that (S)Rh2 induced
apoptosis via both the intrinsic and extrinsic pathways.
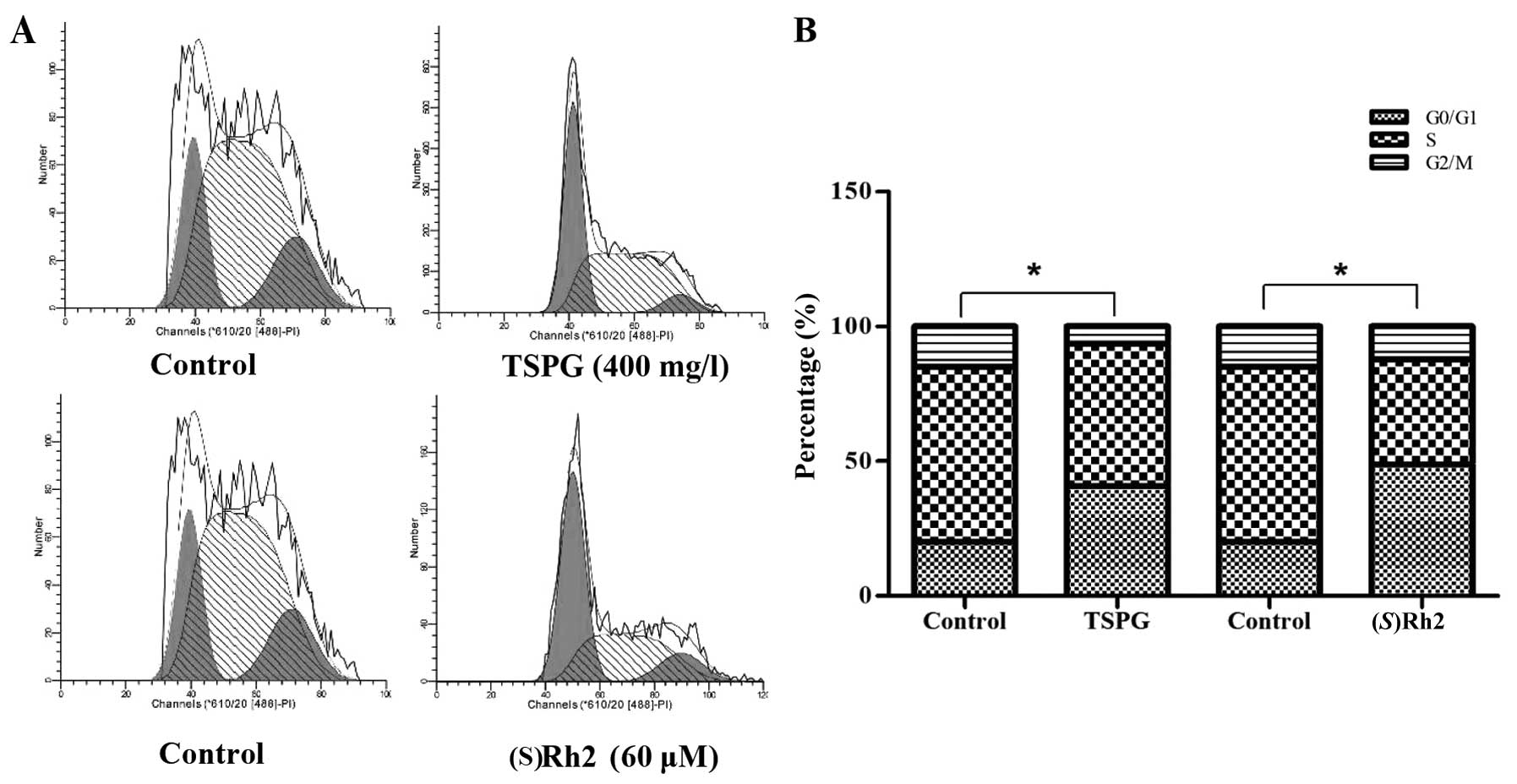
(S)Rh2 induces cell cycle arrest at the
G0/G1 phase in the KG-1a cells
CCK-8 results showed that TSPG and (S)Rh2
inhibited the proliferation of the KG-1a cells. To determine
whether the cell growth inhibitory effect of (S)Rh2 is
associated with cell cycle arrest, changes in cell cycle
distribution were detected by flow cytometry. Compared with the
percentage in the control group, the KG-1a cells after treatment
with (S)Rh2 (60 µM) showed an increased percentage of
cells in the G0/G1 phase from 20.08±3.12 to 48.76±4.22%; the
percentage in the S phase decreased from 64.87±0.52 to 39.22±0.86%;
the percentage in the G2/M phase decreased from 15.05±3.92 to
12.02±2.81% (Fig. 4A and B), and
the difference was statistically significant (p<0.05).
(S)Rh2 induced KG-1a cell cycle arrest at the G0/G1 phase.
TSPG also had an effect on cell cycle arrest (data not shown). To
further explore the underlying mechanisms, we studied the
expression of cell cycle-associated proteins. We found that
(S)Rh2 decreased the level of cyclin D1. The decreased
expression of cyclin D1 may be associated with the G0/G1 cell cycle
arrest induced by (S)Rh2. The results shown that
(S)Rh2 induced KG-1a cell cycle arrest at the G0/G1
phase.
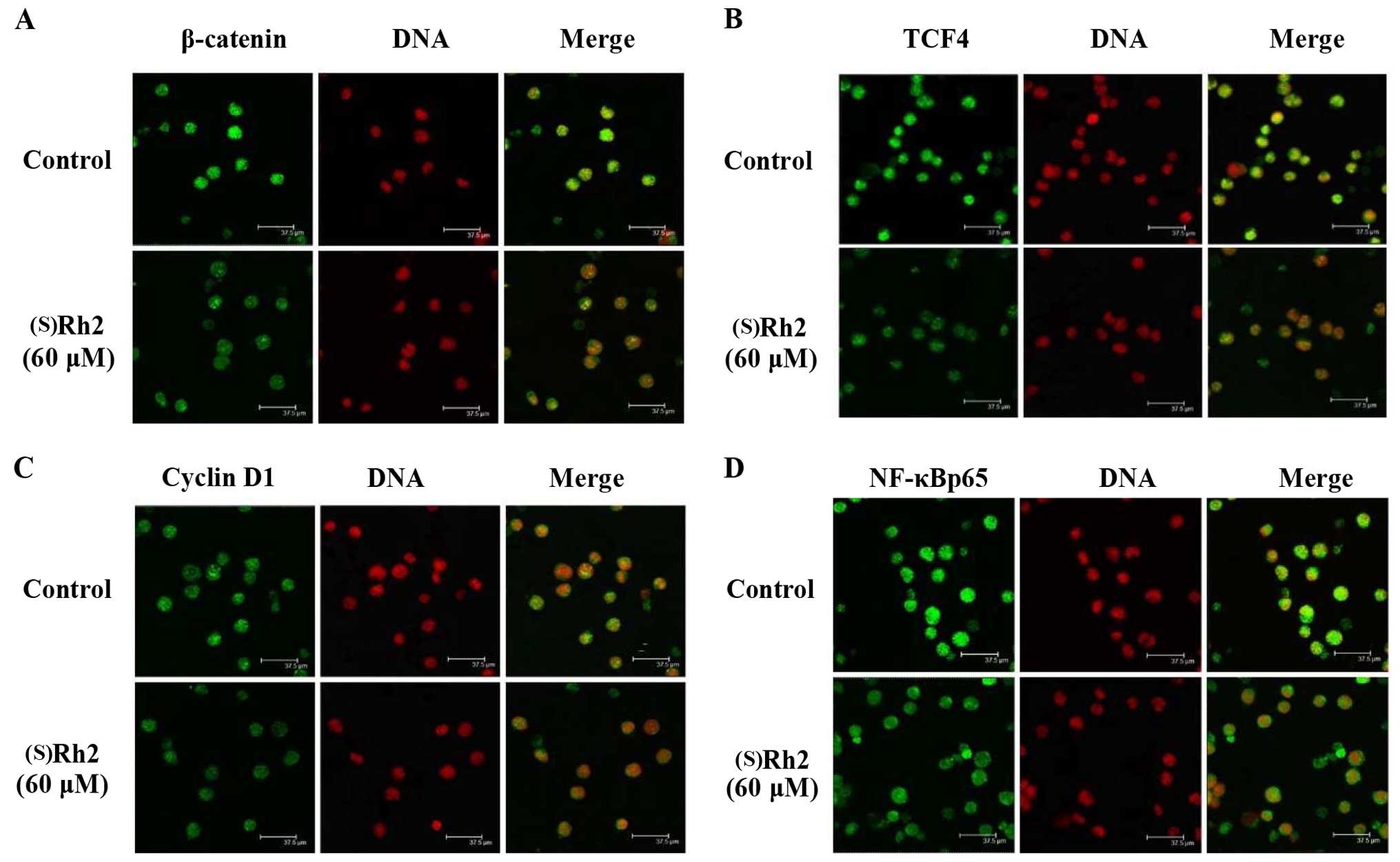
(S)Rh2 affects the localization and
expression of β-catenin, NF-κBp65, TCF-4, and cyclin D1 protein in
the KG-1a cells
Since nuclear β-catenin is a hallmark of the
activated Wnt/β-catenin signaling, we performed immunofluorescence
to determine the localization of β-catenin to further validate the
activation of Wnt/β-catenin signaling. As shown in Fig. 5A, treated cells exhibited strong
green fluorescence in the cytoplasm, and a small amount of green
fluorescence could be visible in the nucleus. Compared with the
control group, β-catenin was obvious altered in the cytoplasm. TCF4
was expressed mainly in the nucleus and the expression of TCF4 in
the nucleus, was significantly decreased after (S)Rh2
treatment for 48 h (Fig. 5B).
Cyclin D1 was expressed in the cytoplasm or nucleus. The expression
level was significantly reduced after (S)Rh2 treatment for
48 h (Fig. 5C). NF-κBp65 is mainly
in the nucleus or cytoplasm, and its expression in the nucleus in
the cells following treatment with (S)Rh2 for 48 h was
significantly reduced (Fig. 5D).
Taken together, these data suggest that (S)Rh2 induced KG-1a
cell cycle arrest at the G0/G1 phase, and may be responsible
for the downregulation of the Wnt/β-catenin signaling pathway
enhancing the cell cycle arrest of KG-1a cells.
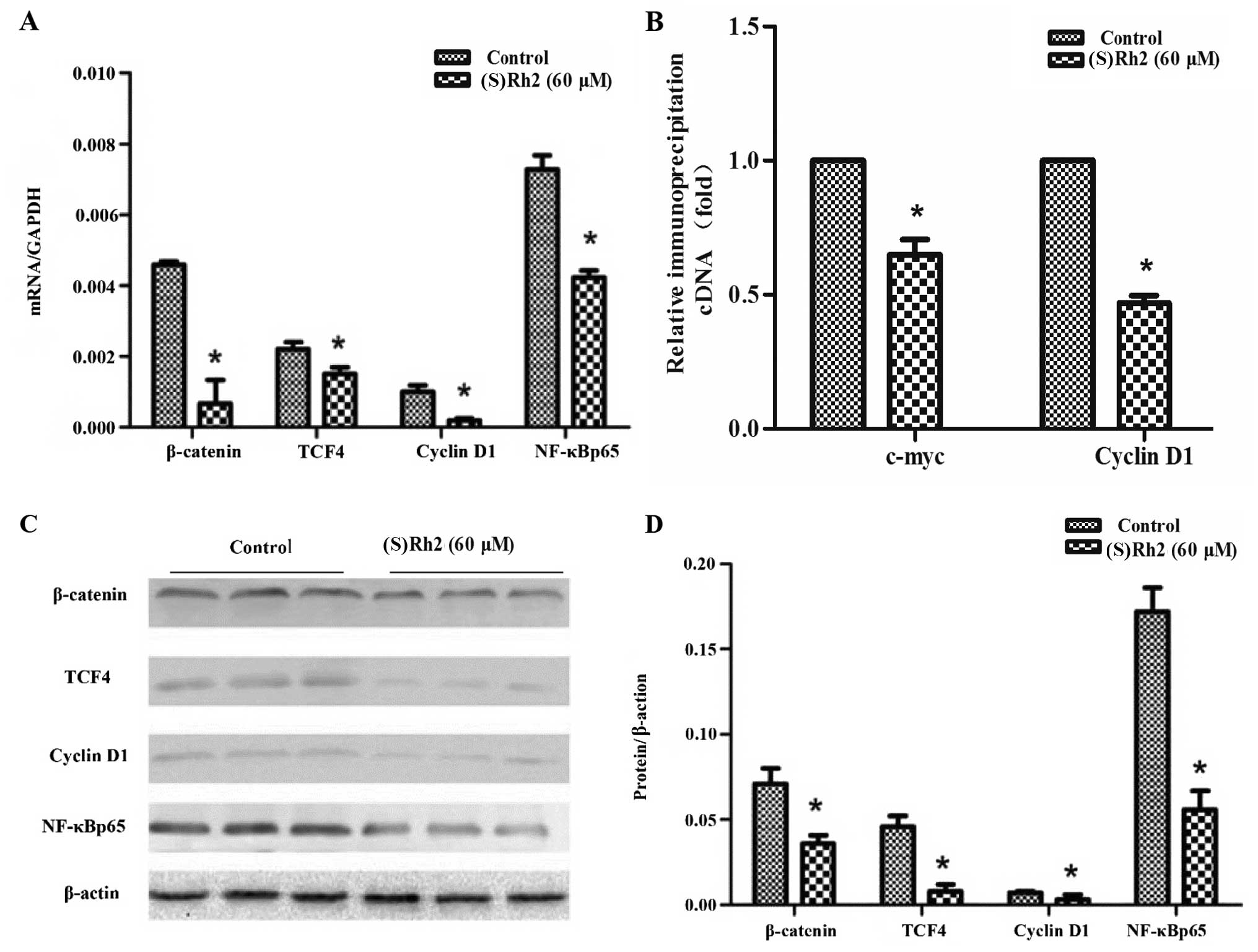
(S)Rh2 affects the mRNA level and protein
expression of β-catenin, TCF-4, cyclin D1 and NF-κBp65 in the KG-1a
cells
KG-1a cells were treated with (S)Rh2 (60
µM) for 48 h. We assessed the mRNA level and protein
expression of several Wnt/β-catenin pathway-related and cell
cycle-related proteins, including β-catenin, TCF-4, cyclin D1 and
NF-κBp65 (Fig. 6). Treatment with
(S)Rh2 (60 µM) for 48 h caused significant
downregulation of the mRNA expression of β-catenin, TCF-4, NF-κBp65
and cyclin D1 in the KG-1a cells, compared with the control group
(Fig. 6A). We also found that in
the (S)Rh2 treatment group reduced protein expression of
β-catenin, TCF4, NF-κBp65 and cyclin D1 was noted in the KG-1a
cells (Fig. 6C and D).
(S)Rh2 affected the gene transcription of c-myc and cyclin
D1 in the KG-1a cells. The ChIP-PCR assay shown that (S)Rh2
decreased the β-catenin/TCF4 target gene transcription, such as
c-myc and cyclin D1. These genes are involved in apoptosis and
proliferation, The ChIP-PCR assay was performed using TCF4 IgG
(Fig. 6B). The downstream genes of
the Wnt/β-catenin pathway including β-catenin, NF-κB and cyclin D1
were also downregulated, further suggesting that (S)Rh2
induced KG-1a cell cycle arrest at the G0/G1 phase and apoptosis by
the Wnt/β-catenin pathway.
Discussion
While cytotoxic agents are often used as traditional
chemotherapy drug, there is an ongoing search for more effective
specific agents that spare normal host tissues. Numerous natural
TCM products have anticancer properties. These compounds generally
have low toxicity and have protective effects against a variety of
cancers (26). On the basis of
research in vitro, we and other researchers have
demonstrated that a structure-activity relationship exists among
the ginsenosides, with the panaxadiols generally being more active
than the panaxatriols in the killing of cancer cells, and the
glycerogelatin compounds being less active as the number of sugar
moieties increases (Fig. 1). To our
knowledge, (S)Rh2 is one of the most active ginsenosides to
be evaluated. (S)Rh2 demonstrated strong effects, and was
more effective at lower doses than TSPG, Rb1, Re, Rg1, Rg3 in KG-1a
cells. The obvious cancer-specific effects of these compounds can
be distinguished from the chemical structure. These may be ascribed
to their differential chemical structure. Anticancer activities
increase with a decrease in the number of sugar moieties in a
ginsenoside molecule. Ginsenosides with four or more sugar
molecules, for example Rb1, showed no significant
anti-proliferative effects; while Re with three sugar residues
weakly inhibited the growth of KG-1a cells; Rg3 and Rg1 (with two
sugar residues), and (S)Rh2 (with one sugar residue) have
been found to inhibit various types of cancer cells and also
enhance the efficacy of chemotherapy agents when treated in
combination (27). (S)Rh2
showed a 2- to 14-fold relatively stronger anti-proliferative
effect than TSPG, Rb1, Re, Rg1, Rg3. (S)Rh2 inhibited the
proliferation and induced the apoptosis of KG-1a cells although the
possible molecular mechanisms require further study.
(S)Rh2 demonstrated anti-proliferative, and
proapoptotic effects and regulation of cell cycle progression.
Although TSPG may be marketed as an anticancer agent, it did not
exert any appreciable effect on KG-1a cells. (S)Rh2
demonstrated a more potent effect and was more effective at lower
doses than the other compounds. (S)Rh2 exhibited decreases
in survival and proliferation, increases in apoptosis and cell
cycle arrest at the G1 phase in KG-1a cells (Fig. 4). Apoptosis is an evolutionarily
conserved form of cell suicide and is characterized by distinctive
morphological changes. We observed that TSPG (400 mg/l) and
(S)Rh2 (60 µM) increased condensed apoptotic nuclei
(Fig. 3C), as evidenced by
chromosomal condensation and formation of apoptotic bodies. TSPG
and (S)Rh2 increased the percentage of Annexin
V+/7-AAD− KG-1a cells as evidence of apoptosis (Fig. 3A and B). Our findings also revealed
that (S)Rh2 can induce apoptosis in KG-1a cells more
efficiently than TSPG.
The Wnt/β-catenin signaling pathway is an
importantly drug discovery target (28). β-catenin is an important factor
involved in the regulation of apoptosis and proliferation in the
Wnt signaling pathway. The Wnt/β-catenin signaling pathway may have
abnormal activation in acute leukemia cells (29). One study has demonstrated that the
accumulation of β-catenin is prompted by increased β-catenin/TCF4
transcriptional activity in the cytoplasm and nucleus, and nuclear
β-catenin accumulation implicates Wnt signaling pathway activation
(30). Therefore, Wnt signaling
pathway inhibitors can induce apoptosis and inhibit proliferation
in acute leukemia (31). The
immunofluorescence experiments confirmed that (S)Rh2 reduced
the β-catenin protein transfer from the cytoplasm to the nucleus in
KG-1a cells. Real-time PCR, immunofluorescence and western blotting
confirmed that (S)Rh2 significantly reduced β-catenin mRNA
and protein expression compared with the control group in the KG-1a
cells. Thus, the Wnt/β-catenin pathway is likely to be an
(S)Rh2 target for the treatment of acute leukemia.
TCF4 is a nuclear transcription factor and has a
molecule 'switch' dual function (32). The Wnt/β-catenin pathway depends on
the presence of different Wnt molecules that bind to the frizzled
receptor. When Wnt/β-catenin pathway proteins are activated,
several proteins that are important for the phosphorylation of
β-catenin require the receptor. Binding of these proteins enables
β-catenin to accumulate in the cytoplasm and translocate into the
nucleus, where it interacts with members of the TCF4 transcription
factor family to induce target gene expression (33,34).
When Wnt/β-catenin pathway proteins are inactivated, several
proteins are released from the frizzled receptor and TCF4 family to
restraint target gene expression (35,36).
Our experiment demonstrated that (S)Rh2 decreased the
protein and mRNA expression of TCF4 in KG-1a cells.
Immunofluorescence results showed that TCF4 is mainly located in
the nucleus, and (S)Rh2 significantly reduced TCF4
expression. ChIP-PCR assays also showed that (S)Rh2
downregulated the transcription of β-catenin/TCF4 target
genes, such as cyclin D1 and c-myc. β-catenin/TCF4 associated cell
proliferation is provided by cyclin D1 and c-myc which have been
reported to be regulated in the wnt pathway (37–39).
Therefore, we speculate that (S)Rh2 can reduce nuclear
β-catenin expression and restrain activation of β-catenin
downstream genes in KG-1a cells.
Cyclin D1 is a Wnt/β-catenin downstream target gene
(40,41). Wnt/β-catenin pathway activated
cyclin D1, disrupting the cell cycle leading to cell abnormal
proliferation in AML (42).
TCF4/β-catenin complex can directly stimulate the activity of CDKs
promoting cell cycle by upregulating cyclin D1. Cyclin D1 is a cell
cycle-related oncogene, which significantly reduces the cells in
the G1 phase by inactivation of (S)Rh2, suppressing
malignant cell proliferation. The experiment showed that
(S)Rh2 arrested KG-1a cells in the G0/G1 phase.
Immunofluorescence and western blotting showed that cyclin D1
protein was significantly reduced. It can be seen that
(S)Rh2 reduced cyclin D1 gene transcription and protein
expression levels. ChIP-PCR also showed that (S)Rh2
downregulated β-catenin/TCF4 target gene transcription, such as
cyclin D1 which is a downstream gene of the Wnt/β-catenin signaling
pathways. Thus, these results indicate that (S)Rh2 induced
cycle arrest at the G0/G1 phase, inhibited proliferation and
promoted apoptosis in the KG-1a cells.
NF-κB is an important member of the Rel gene family
that causes many hematologic malignancies and interacts with
members of the TCF transcription factor family to induce target
gene expression-induced apoptosis and cell cycle arrest (43,44).
This study showed that the expression of NF-κBp65 was reduced and
shifted to the cell cytoplasm from the nucleus following gradual
treatment with (S)Rh2 in the KG-1a cells. The NF-κB and Wnt
signaling pathways can regulate the same target gene transcription
which is mainly associated with cell proliferation, cell cycle
regulation and apoptosis (45). The
irregular activation of gene promoter through NF-κB and TCF
targeted activation of cyclin D1 and c-myc resulting in abnormal
proliferation of tumor cells (46).
(S)Rh2 inhibited proliferation and induced apoptosis
possibly through inhibition of the Wnt and NF-κB pathways in the
KG-1a cells.
In the present study, we found that (S)Rh2
played an antitumor role accompanied by β-catenin protein
expression. When chromatin is in a state of relaxation, β-catenin
can combine with TCF4 in the nucleus. Here, TCF4 competes with
β-catenin to form a complex and is recruited to target promoters to
suppress their expression, such as c-myc, Bcl-2, cyclin D1, thus
inhibiting proliferation and promoting apoptosis in human leukemic
cells (47). We detected that Bax,
Bcl-2, cyclin D1 and cleaved caspase-3 protein was increased in the
(S)Rh2-treated group. At the same time, (S)Rh2
inhibited proliferation and apoptosis through the Wnt/β-catenin
signaling pathway in human leukemic KG-1a cells.
In conclusion, we initially considered that
(S)Rh2 more efficiently inhibits the proliferation and
induces the apoptosis than other monomers in KG-1a cells. The
mechanism may be through the abnormal Wnt pathway by inhibiting the
activation of β-catenin protein, thereby inhibiting its downstream
protein TCF4 and cyclin D1 expression. Meanwhile, inhibition of the
Wnt pathway in crosstalk also inhibits the NF-κB pathway.
Therefore, we believe that (S)Rh2 induced proliferation
inhibition and apoptosis through the Wnt/β-catenin signaling
pathway in human leukemic KG-1a cells. Therefore, (S)Rh2
could be a potential and powerful chemopreventive agent to treat
human leukemia, although further research must be carried out to
fully investigate the mechanisms of action.
Acknowledgments
This study was supported by the Science and
Technology Foundation of Chongqing Municipal Education Commission
(nos. KJ110328 and KJ110308) and the National Natural Science
Foundation of China (no. 81171929). The authors would like to
acknowledge College of Life Science for the technical support.
References
|
1
|
Pui C-H, Robison LL and Look AT: Acute
lymphoblastic leukaemia. Lancet. 371:1030–1043. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jia WD, Sun HC, Zhang JB, Xu Y, Qian YB,
Pang JZ, Wang L, Qin LX, Liu YK and Tang ZY: A novel peptide that
selectively binds highly metastatic hepatocellular carcinoma cell
surface is related to invasion and metastasis. Cancer Lett.
247:234–242. 2007. View Article : Google Scholar
|
|
3
|
Wang S, Wu X, Tan M, Gong J, Tan W, Bian
B, Chen M and Wang Y: Fighting fire with fire: poisonous Chinese
herbal medicine for cancer therapy. J Ethnopharmacol. 140:33–45.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kitaoka F, Kakiuchi N, Long C, Itoga M,
Mitsue A, Mouri C and Mikage M: Molecular characterization of
akebia plants and the derived traditional herbal medicine. Biol
Pharm Bull. 32:665–670. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xia T, Wang JC, Xu W, Xu LH, Lao CH, Ye QX
and Fang JP: 20S-ginsenoside Rh2 induces apoptosis in human
leukaemia Reh cells through mitochondrial signaling pathways. Biol
Pharm Bull. 37:248–254. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Toh DF, Patel DN, Chan EC, Teo A, Neo SY
and Koh HL: Anti-proliferative effects of raw and steamed extracts
of Panax notoginseng and its ginsenoside constituents on human
liver cancer cells. Chin Med. 6:42011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shergis JL, Zhang AL, Zhou W and Xue CC:
Panax ginseng in randomised controlled trials: a systematic review.
Phytother Res. 27:949–965. 2013. View
Article : Google Scholar
|
|
8
|
Lee HS, Kim MR, Park Y, Park HJ, Chang UJ,
Kim SY and Suh HJ: Fermenting red ginseng enhances its safety and
efficacy as a novel skin care anti-aging ingredient: in vitro and
animal study. J Med Food. 15:1015–1023. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang JWLR, Wang YP, et al: The role of
total saponins of panax ginseng in vitro induced CD34 hematopoietic
stem/progenitor cell proliferation. Chin J Anat. 29:430–432.
2006.
|
|
10
|
Zhang C, Yu H and Hou J: Effects of 20 (S)
-ginsenoside Rh2 and 20 (R)-ginsenoside Rh2 on proliferation and
apoptosis of human lung adenocarcinoma A549 cells. Zhongguo Zhong
Yao Za Zhi. 36:1670–1674. 2011.In Chinese. PubMed/NCBI
|
|
11
|
Shi Q, Li J, Feng Z, Zhao L, Luo L, You Z,
Li D, Xia J, Zuo G and Chen D: Effect of ginsenoside Rh2 on the
migratory ability of HepG2 liver carcinoma cells: recruiting
histone deacetylase and inhibiting activator protein 1
transcription factors. Mol Med Rep. 10:1779–1785. 2014.PubMed/NCBI
|
|
12
|
Guo XX, Li Y, Sun C, Jiang D, Lin YJ, Jin
FX, Lee SK and Jin YH: p53-dependent Fas expression is critical for
ginsenoside Rh2 triggered caspase-8 activation in HeLa cells.
Protein Cell. 5:224–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang JH, Han SJ, Ryu JH, Jang IS and Kim
DH: Ginsenoside Rh2 ameliorates scopolamine-induced learning
deficit in mice. Biol Pharm Bull. 32:1710–1715. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bae EA, Han MJ, Shin YW and Kim DH:
Inhibitory effects of Korean red ginseng and its genuine
constituents ginsenosides Rg3, Rf, and Rh2 in mouse passive
cutaneous anaphylaxis reaction and contact dermatitis models. Biol
Pharm Bull. 29:1862–1867. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nam MH, Kim SI, Liu JR, Yang DC, Lim YP,
Kwon KH, Yoo JS and Park YM: Proteomic analysis of Korean ginseng
(Panax ginseng C.A. Meyer). J Chromatogr B Analyt Technol Biomed
Life Sci. 815:147–155. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kitts DD, Popovich DG and Hu C:
Characterizing the mechanism for ginsenoside-induced cytotoxicity
in cultured leukemia (THP-1) cells. Can J Physiol Pharmacol.
85:1173–1183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dunn IF and Black PM: The neurosurgeon as
local oncologist: cellular and molecular neurosurgery in malignant
glioma therapy. Neurosurgery. 52:1411–1422. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu N, Wu GC, Hu R, Li M and Feng H:
Ginsenoside Rh2 inhibits glioma cell proliferation by targeting
microRNA-128. Acta Pharmacol Sin. 32:345–353. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qu X, Qu S, Yu X, Xu H, Chen Y, Ma X and
Sui D: Pseudo-G-Rh2 induces mitochondrial-mediated apoptosis in
SGC-7901 human gastric cancer cells. Oncol Rep. 26:1441–1446.
2011.PubMed/NCBI
|
|
20
|
Kim MJ, Yun H, Kim DH, Kang I, Choe W, Kim
SS and Ha J: AMP-activated protein kinase determines apoptotic
sensitivity of cancer cells to ginsenoside-Rh2. J Ginseng Res.
38:16–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cadigan KM and Nusse R: Wnt signaling: a
common theme in animal development. Genes Dev. 11:3286–3305. 1997.
View Article : Google Scholar
|
|
22
|
Neppl RL and Wang DZ: The myriad essential
roles of microRNAs in cardiovascular homeostasis and disease. Genes
Dis. 1:18–39. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Polakis P: Drugging Wnt signalling in
cancer. EMBO J. 31:2737–2746. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu D, Zhao Y, Tawatao R, Cottam HB, Sen M,
Leoni LM, Kipps TJ, Corr M and Carson DA: Activation of the Wnt
signaling pathway in chronic lymphocytic leukemia. Proc Natl Acad
Sci USA. 101:3118–3123. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu D, Liu JX, Endo T, Zhou H, Yao S,
Willert K, Schmidt-Wolf IG, Kipps TJ and Carson DA: Ethacrynic acid
exhibits selective toxicity to chronic lymphocytic leukemia cells
by inhibition of the Wnt/beta-catenin pathway. PLoS One.
4:e82942009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang W, Wang H, Rayburn ER, Zhao Y, Hill
DL and Zhang R: 20(S)-25-methoxyl-dammarane-3beta, 12beta,
20-triol, a novel natural product for prostate cancer therapy:
activity in vitro and in vivo and mechanisms of action. Br J
Cancer. 98:792–802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nag SA, Qin JJ, Wang W, Wang MH, Wang H
and Zhang R: Ginsenosides as anticancer agents: in vitro and in
vivo activities, structure-activity relationships, and molecular
mechanisms of action. Front Pharmacol. 3:252012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Takahashi-Yanaga F and Sasaguri T: The
Wnt/beta-catenin signaling pathway as a target in drug discovery. J
Pharmacol Sci. 104:293–302. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mai YJ, Qiu LG, Li ZJ, Yu Z, Li CH, Wang
YF, Wang GR and Li Q: The expression of beta-catenin and its
significance in leukemia cells. Zhonghua Xue Ye Xue Za Zhi.
28:541–544. 2007.In Chinese. PubMed/NCBI
|
|
30
|
Saldanha G, Ghura V, Potter L and Fletcher
A: Nuclear beta-catenin in basal cell carcinoma correlates with
increased proliferation. Br J Dermatol. 151:157–164. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Minke KS, Staib P, Puetter A, Gehrke I,
Gandhirajan RK, Schlösser A, Schmitt EK, Hallek M and Kreuzer KA:
Small molecule inhibitors of WNT signaling effectively induce
apoptosis in acute myeloid leukemia cells. Eur J Haematol.
82:165–175. 2009. View Article : Google Scholar
|
|
32
|
in't Hout FE, van der Reijden BA,
Monteferrario D, Jansen JH and Huls G: High expression of
transcription factor 4 (TCF4) is an independent adverse prognostic
factor in acute myeloid leukemia that could guide treatment
decisions. Haematologica. 99:e257–e259. 2014. View Article : Google Scholar
|
|
33
|
Chang HR, Cheng TL, Liu TZ, Hu HS, Hsu LS,
Tseng WC, Chen CH and Tsao DA: Genetic and cellular
characterizations of human TCF4 with microsatellite instability in
colon cancer and leukemia cell lines. Cancer Lett. 233:165–171.
2006. View Article : Google Scholar
|
|
34
|
Tian W, Xu Y, Han X, Duggineni S, Han X,
Huang Z and An J: Development of a novel fluorescence
polarization-based assay for studying the β-catenin/Tcf4
interaction. J Biomol Screen. 17:530–534. 2012. View Article : Google Scholar
|
|
35
|
Cadigan KM: Wnt-beta-catenin signaling.
Curr Biol. 18:R943–R947. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lento W, Congdon K, Voermans C, Kritzik M
and Reya T: Wnt signaling in normal and malignant hematopoiesis.
Cold Spring Harb Perspect Biol. 5:pii: a008011. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dai WB, Ren ZP, Chen WL, Du J, Shi Z and
Tang DY: Expression and significance of APC, beta-catenin, C-myc,
and cyclin D1 proteins in colorectal carcinoma. Ai Zheng. 26:963–6.
2007.In Chinese. PubMed/NCBI
|
|
38
|
Baek SH, Kioussi C, Briata P, Wang D,
Nguyen HD, Ohgi KA, Glass CK, Wynshaw-Boris A, Rose DW and
Rosenfeld MG: Regulated subset of G1 growth-control genes in
response to derepression by the Wnt pathway. Proc Natl Acad Sci
USA. 100:3245–3250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
He TC, Sparks AB, Rago C, Hermeking H,
Zawel L, da Costa LT, Morin PJ, Vogelstein B and Kinzler KW:
Identification of c-MYC as a target of the APC pathway. Science.
281:1509–1512. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gandhirajan RK, Poll-Wolbeck SJ, Gehrke I
and Kreuzer KA: Wnt/β-catenin/LEF-1 signaling in chronic
lymphocytic leukemia (CLL): a target for current and potential
therapeutic options. Curr Cancer Drug Targets. 10:716–727. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tung JN, Chiang CC, Tsai YY, Chou YY, Yeh
KT, Lee H and Cheng YW: CyclinD1 protein expressed in pterygia is
associated with β-catenin protein localization. Mol Vis.
16:2733–2738. 2010.PubMed/NCBI
|
|
42
|
Wang YX, Zhang JH and Gu ZW: Beta-catenin
and cyclin D1 mRNA levels in newly diagnosed patients with acute
myeloid leukemia and their significance. Zhongguo Shi Yan Xue Ye
Xue Za Zhi. 17:304–308. 2009.In Chinese. PubMed/NCBI
|
|
43
|
Fabre C, Carvalho G, Tasdemir E, Braun T,
Adès L, Grosjean J, Boehrer S, Métivier D, Souquère S, Pierron G,
et al: NF-kappaB inhibition sensitizes to starvation-induced cell
death in high-risk myelodysplastic syndrome and acute myeloid
leukemia. Oncogene. 26:4071–4083. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Frelin C, Imbert V, Griessinger E, Peyron
AC, Rochet N, Philip P, Dageville C, Sirvent A, Hummelsberger M,
Bérard E, et al: Targeting NF-kappaB activation via pharmacologic
inhibition of IKK2-induced apoptosis of human acute myeloid
leukemia cells. Blood. 105:804–811. 2005. View Article : Google Scholar
|
|
45
|
Li K, Hu C, Mei C, Ren Z, Vera JC, Zhuang
Z, Jin J and Tong H: Sequential combination of decitabine and
idarubicin synergistically enhances anti-leukemia effect followed
by demethylating Wnt pathway inhibitor promoters and downregulating
Wnt pathway nuclear target. J Transl Med. 12:1672014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chun KS and Surh YJ: Signal transduction
pathways regulating cyclooxygenase-2 expression: potential
molecular targets for chemoprevention. Biochem Pharmacol.
68:1089–1100. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yeh CT1, Yao CJ, Yan JL, Chuang SE, Lee
LM, Chen CM, Yeh CF, Li CH and Lai GM: Apoptotic cell death and
inhibition of Wnt/beta-catenin signaling pathway in human colon
cancer cells by an active fraction (hs7) from Taiwanofungus
camphoratus. Evid Based Complement Alternat Med. 2011:7502302011.
View Article : Google Scholar
|