Introduction
Colorectal cancer (CRC), for which distant
metastasis accounts for the leading cause of cancer mortality, is
one of the most malignant gastrointestinal carcinomas worldwide
(1). The pathogenesis of CRC is a
complex process and involves environmental influences, genetic
alterations, and the host immune system and their interactions. The
formation of an inflammatory microenvironment also plays a pivotal
role in the development of CRC (2).
The host microenvironment is comprised of numerous infiltrating
immune cell types and resident tumor cells. Among the immune cells,
macrophages play an indispensable role. For instance, macrophage
infiltration in colon tissue has been observed in both inflammatory
bowel disease (IBD) and CRC (4),
and the cytokines secreted by macrophages under certain conditions
have directly or indirectly stimulated inflammation and tumor
progression (5,6). Although great progress has been made,
the interactions and the molecular mechanisms between CRC and the
inflammatory microenvironment remain unknown.
S100A8 (calgranulin A or S100 calcium-binding
protein A8) is a member of the low-molecular calcium binding S100
protein family. It also belongs to the family of damage-associated
molecular patterns (DAMPs). Studies have shown that S100A8 plays an
important role in regulating tissue inflammation as a ligand of
toll-like receptor 4 (TLR4) and/or receptor for advanced glycation
end products (RAGE) (7,8). Aberrant expression of S100A8 has been
found in different types of tumors including CRC (9) and the accumulation of S100A8 positive
cells is always located in the invasive margin of colorectal
carcinoma (10) which plays a
critical role in the tumor microenvironment. Interestingly,
mounting evidence indicates that the apical surface of the
intestinal epithelium is exposed to lipopolysaccharides (LPS) from
the lumen due to intestinal mucosal permeability and bacterial
translocation (11). LPS could
cause a cascade of inflammatory responses mediated by diverse
inflammatory cells that secrete various chemokines and cytokines
favoring CRC growth and migration. Nevertheless, little research
has been conducted to elucidate the effects of S100A8 on modulating
the inflammatory response triggered by LPS in the tumor
microenvironment.
MicroRNAs (miRNAs) are small non-coding RNAs which
function in the post-transcriptional regulation of genes and
cellular homeostasis (12).
Deregulated miRNA expression has been observed in diverse disorders
such as autoimmune diseases, chronic inflammatory pathologies and
cancers, including CRC (13).
miR-155, an oncogenic miRNA, is identified as a link between
inflammation and cancer (14).
Previous studies have found that miR-155 expression is induced by
proinflammatory mediators, such as TNF-α and LPS (15), and overexpression of miR-155 has
been found to elevate inflammatory cytokine production in
intestinal myofibroblasts (16) and
monocyte/macrophages (17,18). In addition, some studies indicate
that the promoter region of pri-miR-155 contains putative
NF-κB-binding sites (19,20) and miR-155 upregulation and
subsequent molecular events are associated with inflammatory
processes such as activation of NF-κB (21,22).
However, the role of S100A8 in the modulation of miR-155 expression
in macrophages in the inflammatory microenvironment and its
contribution to the development of CRC have not yet been
investigated.
In the present study, we mimicked the intestinal
inflammatory environment using LPS to investigate the role of
S100A8 in the tumor microenvironment. We found that S100A8 markedly
promoted miR-155 expression and inflammatory cytokine secretion in
the macrophages, which was involved in the activation of the NF-κB
signaling pathway. The results indicate that S100A8 facilitated the
migration but not the viability of CRC cells in the tumor
microenvironment by conditioned medium (CM) culture or co-culture
manner. Our study highlights the significance of S100A8 in the
progression of CRC, and may provide an opportunity for the
development of targeted therapy for CRC.
Materials and methods
Reagents and antibodies
Recombinant plasmids pGST-moluc, pGST-moluc-hS100A8
and pNF-κB-luc were kindly provided by Professor T.C. He (The
University of Chicago Medical Center, Chicago, IL, USA). Competent
bacteria E.coli BL21 were retained in our laboratory. LPS,
inhibitor of NF-κB (Bay 11-7082) and phorbol myristate acetate
(PMA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Radio
immunoprecipitation assay (RIPA) buffer was obtained from Beyotime
(Shanghai, China). Phosphatase inhibitor and protease inhibitor
were purchased from Roche Diagnostics GmbH (Mannheim, Germany).
Nuclear-Cytosol extraction kit was obtained from KeyGen Biotech
Co., Ltd. (Nanjing, China). Polyvinylidene difluoride (PVDF)
membranes and enhanced chemiluminescence (ECL) kit were both
purchased from Millipore Corporation (Billerica, MA, USA).
3-(4,5-Dimethylthiazol-2-yl)-2, 5-diphenyltrazolium bromide (MTT)
reagent was obtained from Sigma-Aldrich. Lipofectamine™ 2000 and
TRIzol reagents were both obtained from Invitrogen Life
Technologies (Carlsbad, CA, USA). Real-time PCR kit and SYBR Premix
Ex Taq™ II were both procured from Takara Bio, Inc. (Dalian,
China). Single Luciferase Reporter Assay system was obtained from
New England Biolabs, Inc. (NEB; Ipswich, MA, USA). Human IL-1β
ELISA kit and human TNF-α ELISA kit were both acquired from ExCell
Biology (Shanghai, China). Crystal violet was purchased from
Sigma-Aldrich. All antibodies used in this study were as follows:
mouse anti-GST monoclonal antibody was obtained from Zoonbio Tech
Co., Ltd. (Nanjing, China), mouse anti-S100A8 monoclonal antibody
was purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA) and rabbit anti-CD284 (TLR4) polyclonal antibody was acquired
from ImmunoWay Biotechnology, Co. (Newark, DE, USA). Rabbit
anti-phospho-NF-κB p65 (Ser536) (93H1) antibody was procured from
Cell Signaling Technology, Inc. (Boston, MA, USA) and rabbit
anti-histone H3.1 monoclonal antibody was obtained from Abmart Inc.
(Shanghai, China). Mouse anti-β-actin monoclonal antibody, goat
anti-rabbit IgG and goat anti-mouse IgG were all purchased from
Zhongshan Golden Bridge Biotechnology Co., Ltd. (Beijing, China).
Alexa Fluor 488 conjugated goat anti-rabbit IgG was acquired from
Beyotime.
Cell lines, cell culture and
differentiation
Human monocyte/macrophage (THP-1) cells and human
colorectal carcinoma cell lines HCT116 and SW480 were purchased
from the American Type Culture Collection (ATCC; Manassas, VA,
USA). THP-1 cells were cultured in Roswell Park Memorial Institute
(RPMI)-1640 medium and HCT116 and SW480 cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) in a controlled
atmosphere of 5% Co2 and 37°C. All the media were
supplemented with 10% fetal bovine serum (FBS) (Gibco Life
Technologies, Carlsbad, CA, USA), 100 U/ml penicillin and 100
µg/ml streptomycin (both obtained from HyClone, Logan, UT,
USA).
Differentiation was achieved by THP-1 cells with PMA
(50 ng/ml). The cells were resuspended at 1×106 cells/ml
and then incubated at 37°C for 24 h. The PMA-containing medium was
removed and the cells were incubated in fresh medium for another 24
h to eliminate the effect of PMA, and used for the following
experiments.
Preparation and identification of
recombinant protein
The preparation of recombinant GST and GST-hS100A8
proteins used in this study was previously described (23). In brief, the pGST-moluc and
pGST-moluc-hS100A8 were transformed into BL21 bacteria by calcium
chloride transformation. Isopropylthio-β-D-galactoside was used to
induce the expression of GST and GST-hS100A8 proteins. Then the
bacteria were collected and sonicated on ice and the supernatant
was spun and incubated with glutathione-sepharose 4B beads
overnight. Next, GST and GST-hS100A8 on the beads were eluted by
elution buffer containing reduced glutathione. Finally, the two
proteins were filtered via a 0.22-µm membrane and stored at
−80°C.
The recombinant GST and GST-hS100A8 proteins were
subjected to polyacrylamide gel electrophoresis. After
electrophoresis, the gel was incubated in Coomassie Brilliant Blue
staining solution for 1 h. Then the Coomassie Brilliant Blue
elution buffer was used to de-stain the gel until the background
turned transparent. All steps were processed on a rotary shaker
with a slow speed of revolution.
Western blot analysis
Differentiated THP-1 macrophages treated with and
without LPS, GST and GST-hS100A8 or pretreated with Bay 11-7082 (50
ng/ml) were collected and washed with ice-cold PBS three times, and
then lysed in ice-cold RIPA buffer containing phosphatase inhibitor
and protease inhibitor. Nuclear and cytoplasmic proteins were
extracted using a nuclear-cytosol extraction kit according to the
manufacturer's instructions. The concentration of proteins was
determined by bicinchoninic acid (BCA) assay and denatured by
boiling water bath. The proteins were separated using 10% sodium
dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and
blotted onto the PVDF membranes. Then the membranes were blocked
with 5% bovine serum albumin at 37°C for 2 h and incubated with
anti-GST monoclonal antibody (1:1,000 dilution), anti-S100A8
monoclonal antibody (1:1,000 dilution), anti-CD284 (TLR4)
polyclonal antibody (1:1,000 dilution), anti-phospho-NF-κB p65
(Ser536) (93H1) antibody (1:1,000 dilution), anti-histone H3.1
monoclonal antibody (1:5,000 dilution), or anti-β-actin monoclonal
antibody (1:1,000 dilution) and then with a relevant secondary
antibody conjugated with horseradish peroxidase (1:5,000 dilution).
The proteins of interest on the membranes were detected using an
ECL kit. The results were recorded using the Bio-Rad
Electrophoresis Documentation (Gel Doc 1000) and Quantity One
version 4.5.0 software (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Preparation of macrophage conditioned
medium
Differentiated THP-1 macrophages in 100 mm dishes
were treated without and with LPS, GST and GST-hS100A8 for 24 h and
the medium was collected, filtered and used as macrophage CM.
Cell viability assay
An MTT assay was used to evaluate the cell
viability. The cells were seeded in 96-well plates and exposed to
the different treatments. After the indicated incubation time, 10
µl of MTT reagent was added into the wells respectively,
followed by another 4 h of incubation at 37°C. Dimethyl sulfoxide
(100 ml) was added to dissolve the formazan product for 10 min at
room temperature. Finally, the absorbance was measured daily for
the following five days at 492 nm using a microplate reader. Each
treatment was performed in quintuplicate, and the overall
experiment was repeated thrice.
Immunofluorescence staining
Differentiated THP-1 macrophages were seeded on
sterile glass coverslips in 24-well culture plates and treated
without and with LPS, GST and GST-hS100A8 in serum-free RPMI-1640
medium for the indicated time. The cells were washed with PBS,
fixed with methanol at room temperature, and permeabilized with
0.25% Triton X-100 at 37°C for 15 min. After being blocked with
goat serum for 30 min at 37°C, the cells were incubated with
anti-CD284 (TLR4) polyclonal antibody or anti-phospho-NF-κB p65
(Ser536) (93H1) antibody (1:200 dilution in PBS) at 4°C overnight.
Subsequently, the cells were rinsed with PBS to remove the primary
antibody and incubated with Alexa Fluor 488-conjugated goat
anti-rabbit IgG at a 1:200 dilution for 1 h at 37°C in the dark.
Next, the cells were washed with PBS, counterstained with DAPI for
5 min, washed again with PBS and then the coverslips were mounted
using antifade mounting medium. The fluorescence of the different
groups was viewed and photographed with an inverted fluorescence
microscope (Nikon Eclipse 80i; Nikon Corporation, Tokyo,
Japan).
Transient transfection
Differentiated THP-1 macrophages were transfected
with the miR-155 inhibitor or miR-NC at a final concentration of
100 nmol/l using Lipofectamine™ 2000 according to the
manufacturer's instructions. Then the cells were treated without
and with LPS and GST-hS100A8 for 24 h and total RNA and the
supernatant was collected to measure IL-1β and TNF-α by
semi-quantitative RT-PCR (semi-PCR) and ELISA.
Total RNA isolation, semi-PCR and
quantitative real-time PCR (qPCR)
Total RNA from the treated cells was extracted using
TRIzol reagents, cDNA samples synthesized using random primers for
IL-1β, TNF-α and GAPDH with real-time (RT)-PCR kit according to the
manufacturer's instructions were used as templates for semi-PCR.
Semi-PCR products were identified by electrophoresis with 2%
agarose gels and recorded using the Gel Doc 1000 imaging system.
cDNA samples synthesized using specific primers for miRNAs and U6
(GenScript Co., Ltd., Nanjing, China) were used as templates for
the detection of miRNAs by quantitative qPCR. qPCR was performed on
the CFX96 real-time PCR detection system from Bio-Rad using SYBR
Premix Ex Taq™ II. Data were collected and analyzed by the
comparative 2−ΔΔCt method with U6 as the control. The
primers in this study are shown in Table I.
 | Table IThe primers used in this study. |
Table I
The primers used in this study.
| Gene | | Sequences |
|---|
| GAPDH | Forward
primer: |
5′-CAGCGACACCCACTCCTC-3′ |
| Reverse
primer: |
5′-TGAGGTCCACCACCCTGT-3′ |
| IL-1β | Forward
primer: |
5′-GCCCTAAACAGATGAAGTGCTC-3′ |
| Reverse
primer: |
5′-GCCCTAAACAGATGAAGTGCTC-3′ |
| TNF-α | Forward
primer: |
5′-CAGCCTCTTCTCCTTCCTGA-3′ |
| Reverse
primer: |
5′-GGAAGACCCCTCCCAGATAGA-3′ |
| hsa-miR-155-5p | RT primer: |
5′-CTCAACTGGTGTCGTGGGTCGGCAATTCAGTTGAGACCCCTAT-3′ |
| Forward
primer: |
5′-ACACTCCAGCTGGGTTAATGCTAATCGTGAT-3′ |
| Reverse
primer: |
5′-TGGTGTCGTGGAGTCG-3′ |
| U6 | RT primer: |
5′-AACGCTTCACGAATTTGCGT-3′ |
| Forward
primer: |
5′-AACGCTTCACGAATTTGCGT-3′ |
| Reverse
primer: |
5′-AACGCTTCACGAATTTGCGT-3′ |
Gaussia luciferase activity assay
Gaussia luciferase reporter assay was carried
out as previously described (24).
Differentiated THP-1 macrophages were evenly seeded in a 24-well
plate and transfected with pNF-κB-luc plasmid using Lipofectamine™
2000 according to the manufacturer's instructions. Then the cells
were then treated without and with LPS, GST and GST-hS100A8 for 24
h or pretreated with Bay 11-7082 for a period of 30 min prior to
the treatment. Gaussia luciferase activity was measured
using a Single Luciferase Reporter Assay system according to the
manufacturer's instructions.
ELISA
Supernatant of differentiated THP-1 macrophages in
different groups was collected, centrifuged at 1,000 × g for 20 min
and stored at −80°C. Specific ELISA kits were used for measuring
IL-1β and TNF-α according to the manufacturer's instructions.
Flow cytometric analysis
HCT116 cells were co-cultured with differentiated
THP-1 macrophages which were treated without and with LPS, GST and
GST-hS100A8 or pretreated with Bay 11-7082. After being treated for
24 h, the cells were harvested by trypsinization and fixed with 70%
ice-cold ethanol for at 4°C overnight. Cell cycle distribution was
analyzed by using a FACSVantage SE flow cytometer
(Becton-Dickinson, San Jose, CA, USA).
Hoechst staining assay
The Hoechst staining assay was performed according
to the manufacturer's instructions using Hoechst 33258. The HCT116
cells seeded in 24-well plates were co-cultured with the
differentiated THP-1 macrophages which were treated without and
with LPS, GST and GST-hS100A8 or pretreated with Bay 11-7082 for 24
h. Then the cells were fixed with 4% paraformaldehyde for 10 min
and stained with Hoechst staining solution (1:1,000 dilution) for
30 min at room temperature in the dark. Finally, the cells were
viewed under an inverted fluorescence microscope. Each experiment
was performed three times.
Wound healing assay
HCT116 and SW480 cells were seeded into 6-well
culture plates and incubated at 37°C until the cells grew to 95%
confluency. A wound was made in the center of the cell monolayers
with a 10-µl sterile plastic pipette tip, and the well was
washed with PBS three times and incubated in serum-free DMEM.
Differentiated THP-1 macrophages seeded into the upper inserts
(Millipore Corporation) and treated without and with LPS, GST,
GST-hS100A8 or pretreated with Bay 11-7082 were co-cultured with
the above cells, respectively. Wound areas were observed and
photographed at ×100 magnification at 0, 24, 48 and 72 h.
Transwell migration assay
The Transwell migration assay was conducted using
24-well Transwell cell culture chambers (Millipore Corporation).
Briefly, HCT116 and SW480 cells were harvested after 24 h of
co-culture with the differentiated THP-1 macrophages which were
treated without and with LPS, GST and GST-hS100A8 or pretreated
with Bay 11-7082. Then, 1×105 CRC cells were resuspended
in 200 µl of DMEM supplemented with 10% FBS, and added to
the upper inserts. DMEM (500 µl) with 10% FBS was added to
the lower chamber. After the CRC cells attached, the medium in the
upper inserts was replaced with serum-free DMEM, and the medium in
the lower chambers was replaced with 20% FBS DMEM. Twenty-four
hours later, the migrated cells were fixed with methanol for 20 min
and stained with 0.05% crystal violet for 30 min. Cells on the
upper surface of the insert membrane were removed with cotton
swabs. The migrated cells were counted at ×100 magnification in 10
different fields for each insert. The experiments were repeated
three times.
Statistical analysis
All data are presented as mean ± SEM (standard error
of the mean). Differences were analyzed using the two-tailed
Student's t-test for comparison between two groups or one-way ANOVA
followed by Tukey's multiple comparison test. All the statistical
analyses were performed using GraphPad Prism 5 (GraphPad Software,
Inc., La Jolla, CA, USA). Significant probability values are
indicated as P<0.05.
Results
Effects of LPS and recombinant S100A8
protein on the viability of the differentiated THP-1
macrophages
In this study, we first prepared GST and GST-hS100A8
proteins, which were identified by SDS-PAGE and western blot
analysis (Fig. 1A and B). Their
purities were confirmed to be >90% by Quantity One software
after SDS-PAGE and Coomassie Brilliant Blue staining (Fig. 1A). The purified proteins were used
to treat cells in our subsequent experiments.
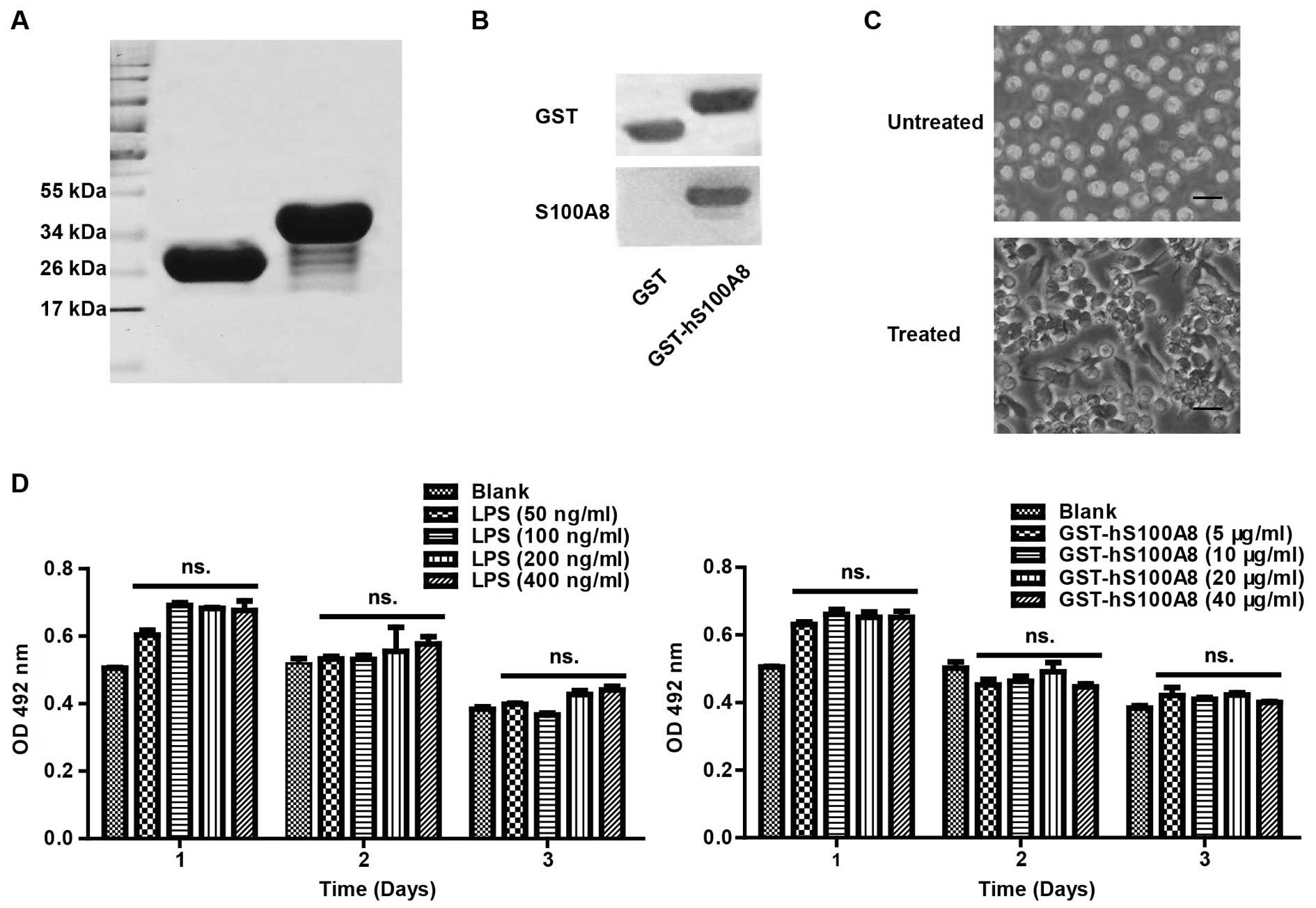 | Figure 1Characterization of the human THP-1
cell line. (A) Recombinant protein GST-hS100A8 was found to be ~37
kDa, and GST was ~26 kDa; their purities were >90% (by Quantity
One software). (B) GST-hS100A8 was recognized by anti-GST and
anti-S100A8 antibodies by western blot analysis. Left lane, GST
protein; right lane, GST-hS100A8 protein. (C) Morphological changes
of THP-1 cells with differentiation. Representative differential
contrast images of THP-1 cells treated with or without PMA. Black
scale bars, 10 µm. (D) MTT assay. The viability of the
differentiated THP-1 cells treated with different concentrations of
LPS or GST-S100A8 was detected by MTT assay at 24, 48 and 72 h, as
described in Materials and methods. The absorbance was measured at
492 nm using a microplate reader. The results represent the mean
absorbance ± SEM of 5 independent experiments. ns., P>0.05, as
compared with each group. PMA, Phorbol myristate acetate; LPS,
lipopolysccharides. |
THP-1 is a human monocytic leukemia cell line, and
it can be induced by PMA to differentiate into macrophage-like
cells, which can mimic native monocyte-derived macrophages. After
treatment with PMA, the cells were characterized by reduced
proliferation, increased adherence and extended pseudopodia
(Fig. 1C), indicating that the
THP-1 cells were induced to differentiate into macrophages.
Secondly, the effects of LPS and S100A8 on the
viability of the macrophages were assayed by the MTT assay. After
the cells were treated with LPS at 0, 50, 100, 200 and 400 ng/ml or
GST-hS100A8 at 0, 5, 10, 20 and 40 µg/ml for 3 successive
days, we found that there were no significant effects on cell
viability compared to the control group (Fig. 1D).
S100A8 activates the TLR4/NF-κB pathway
in the macrophages in an inflammatory microenvironment
Previous studies have shown that TLR4 is the
dominant receptor mediating S100A8 signaling in macrophages, and
the NF-κB pathway is consistently activated by the ligands of TLR4
(25,26). We examined the effects of S100A8 on
the activation of TLR4 and NF-κB in the macrophages by
immunofluorescence staining. The results showed that there was a
more obvious activation on TLR4 following the treatment of the
combination LPS and S100A8 as well as LPS or S100A8 respectively
(Fig. 2A).
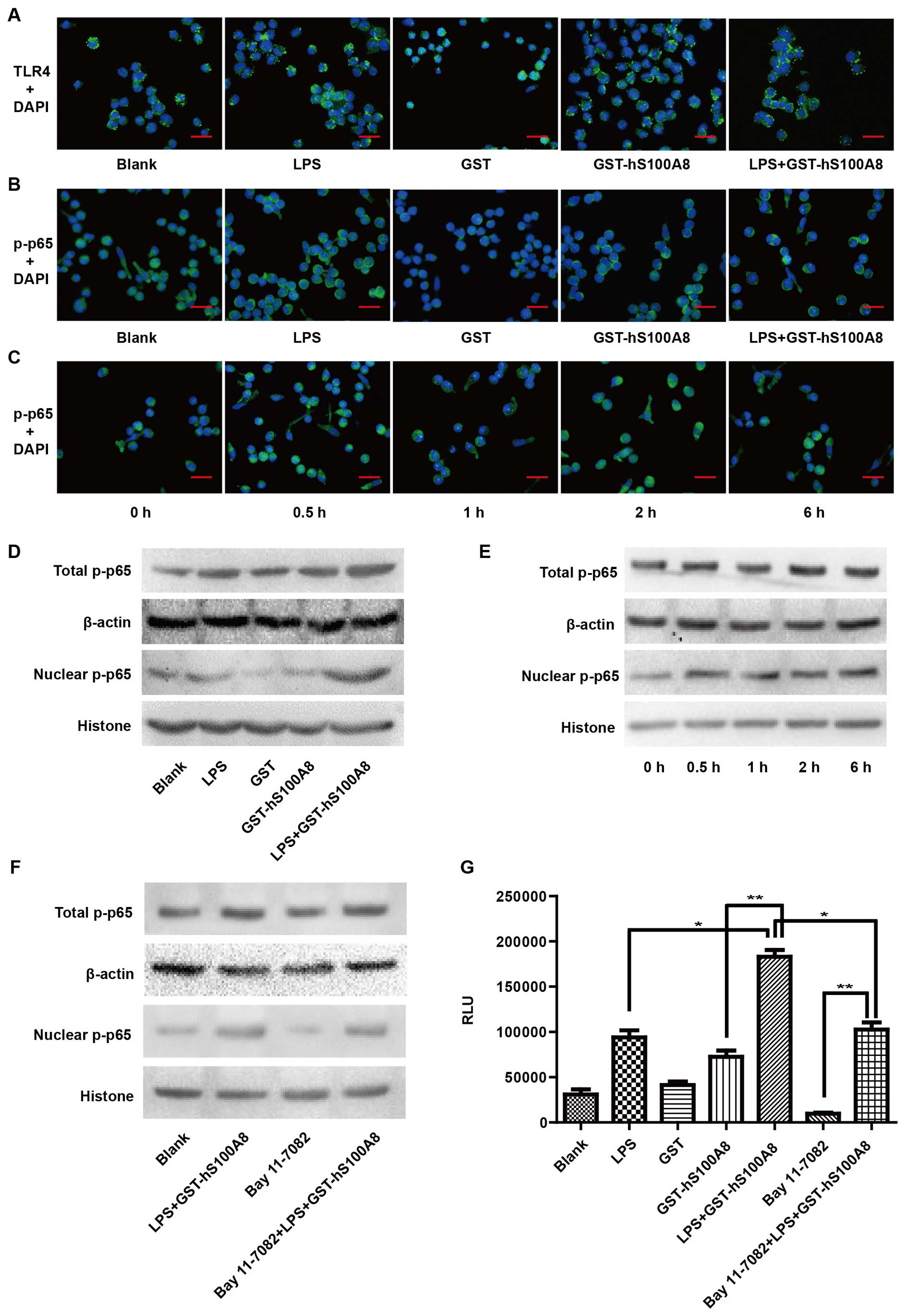 | Figure 2Recombinant S100A8 protein induces
the activation of TLR4 and NF-κB in macrophages in an inflammatory
microenvironment. (A) Macrophages were not treated or treated with
LPS (100 ng/ml), GST (10 µg/ml), GST-hS100A8 (10
µg/ml) and LPS+ GST-hS100A8 for 24 h, and the expression of
TLR4 was determined by immunofluorescence staining. (B) Macrophages
were not treated or treated with LPS (100 ng/ml), GST (10
µg/ml), GST-hS100A8 (10 µg/ml) and LPS+ GST-hS100A8
for 1 h or (C) treated with LPS (100 ng/ml) and GST-hS100A8 (10
µg/ml) for 0, 0.5, 1, 2, 6 and 12 h, and nuclear
translocation of p-NF-κB p65 was determined by immunofluorescence
staining. The representative images are shown. Red scale bars, 10
µm. (D and E) Macrophages were treated as above and total
and nuclear p-NF-κB p65 proteins were assessed by western blot
analysis. (F) Macrophages were pretreated with an inhibitor of
NF-κB (Bay 11-7082, 50 ng/ml) for a period of 30 min prior to
treatment for 24 h without or with LPS (100 ng/ml) and GST-hS100A8
(10 µg/ml). Total and nuclear p-NF-κB p65 proteins were
assessed by western blot analysis. β-actin and histone were used as
internal reference controls. (G) Macrophages were transfected with
the pNF-κB-luc plasmid, then pretreated with Bay 11-7082 for a
period of 30 min prior to treatment for 24 h without or with LPS
(100 ng/ml), GST (10 µg/ml), GST-hS100A8 (10 µg/ml),
LPS+ GST-hS100A8 and a combination of Bay 11-7082+LPS+GST-hS100A8.
The luciferase activity of NF-κB was detected in the supernatant.
*P≤0.05, **P≤0.01 compared with the control
group. TLR4, toll-like receptor 4; LPS, lipopolysaccharides. |
Next, we examined whether the treatment of LPS and
S100A8 can transport p-NF-κB p65 to the nucleus. Our results
confirmed that S100A8 promoted p-NF-κB p65 nuclear translocation
and was more significant in the inflammatory microenvironment
(Fig. 2B). Moreover, as shown in
Fig. 2C, the treatment of LPS and
S100A8 significantly promoted p-NF-κB p65 nuclear translocation and
reached a peak value 1 h after treatment
To further confirm that S100A8 facilitates p-NF-κB
p65 nuclear translocation in the inflammatory microenvironment,
western blot analysis was performed to detect the distribution of
p-NF-κB p65 in macrophages. We found that the levels of total and
nuclear p-NF-κB p65 were elevated much more after the treatment of
LPS and S100A8 together than these levels in the control group
(Fig. 2D). Moreover, the levels of
nuclear p-NF-κB p65 were increased after the treatment of LPS and
S100A8 with time, but the levels of total p-NF-κB p65 were not
significantly altered (Fig. 2E).
The treatment of LPS and S100A8 together partially reversed the
suppressive effect of Bay 11-7082 on the macrophages (Fig. 2F). Furthermore, a luciferase
activity assay was used to analyze the activation of the NF-κB
pathway. S100A8 significantly increased luciferase activity of
NF-κB in the inflammatory microenvironment and partially reversed
the suppressive effect of Bay 11-7082 (Fig. 2G). These findings indicated that the
recombinant S100A8 protein could distinctly activate the TLR4/NF-κB
signaling pathway of macrophages in an inflammatory
microenvironment.
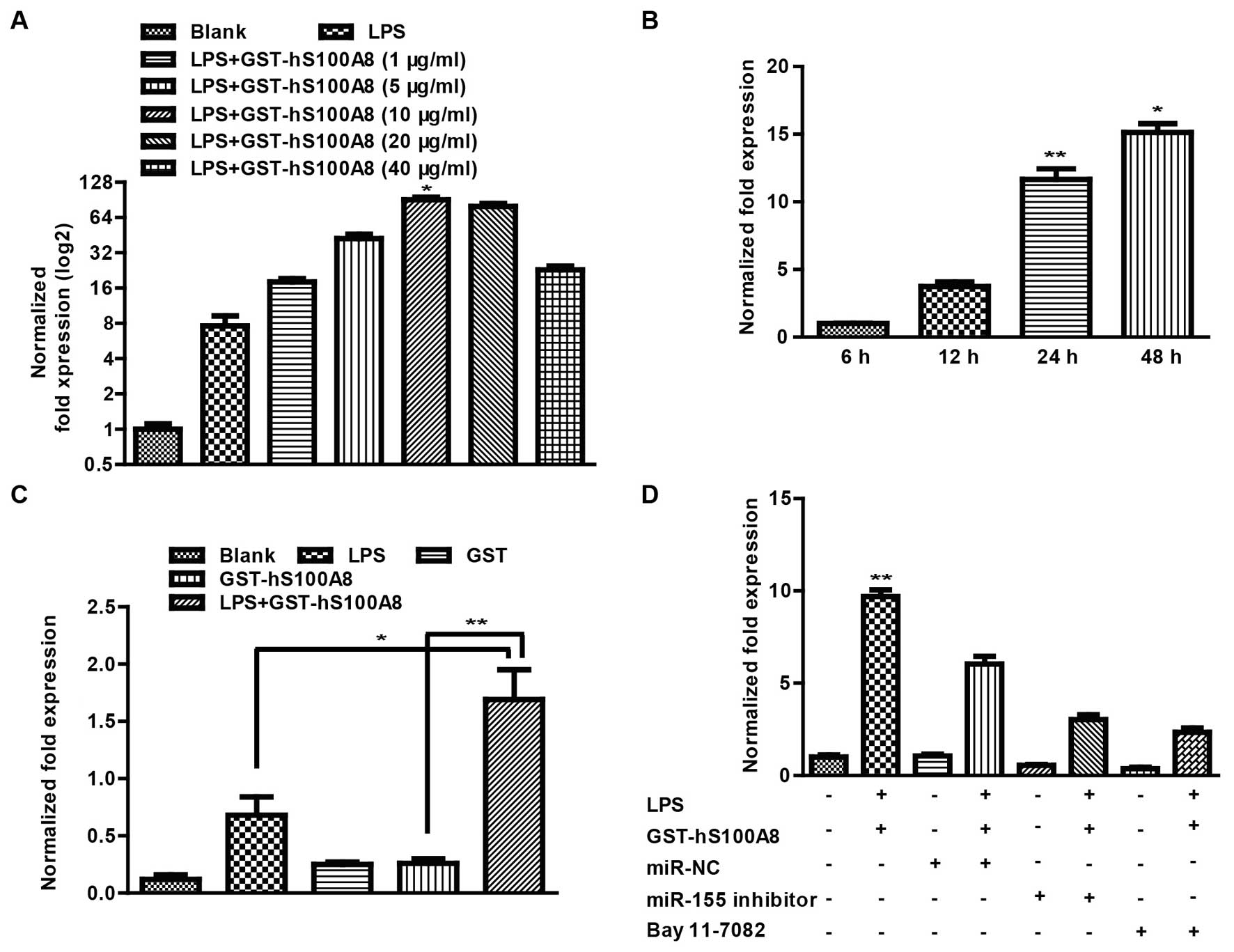
S100A8 promotes miR-155 expression
through activation of the NF-κB pathway
It is well known that miR-155 is an oncogenic RNA
that has been shown to be critical in the crosstalk between
inflammation and cancer (27). To
test whether S100A8 can regulate the expression of miR-155 in an
inflammatory microenvironment, we first investigated the influence
of S100A8 on the expression of miR-155 at different protein
concentrations and treatment times. After the macrophages were
treated without and with LPS (100 ng/ml) and GST-hS100A8 at 0, 5,
10, 20, and 40 µg/ml for 24 h, we found that LPS (100 ng/ml)
and GST-hS100A8 at 10 µg/ml had a more significant effect on
expression of miR-155 (Fig. 3A).
After the macrophages were treated with a combination of LPS (100
ng/ml) and GST-hS100A8 (10 µg/ml) for 6, 12, 24 and 48 h, we
found that the expression of miR-155 was increased with time
(Fig. 3B). It should be noted that
S100A8 upregulated the expression of miR-155 in a time-dependent
manner. In addition, after the macrophages were treated without and
with LPS (100 ng/ml), GST, GST-hS100A8 at 10 µg/ml for 24 h,
we found an obvious increase in the expression of miR-155 following
the treatment of a combination of LPS and S100A8 compared with that
of the other groups (Fig. 3C) and
treatment of LPS and S100A8 partially reversed the suppressive
effect of the miR-155 inhibitor (Fig.
3D). However, following pretreatment with Bay 11-7082 for a
period of 30 min prior to the treatment of LPS and S100A8 for 24 h,
the expression of miR-155 was decreased compared with that of the
treatment of the combination of LPS and S100A8 (Fig. 3D).
Collectively, these results suggested that S100A8
induced the upregulation of miR-155 expression and depended on the
activation of the NF-κB pathway.
S100A8-induced upregulation of miR-155
enhances the secretion of inflammatory cytokines in the macrophages
in the inflammatory microenvironment
It has been previously reported that inflammatory
cytokines are mainly derived from immune cells such as
monocyte/macrophages and dendritic cells (28) and miR-155 regulates inflammatory
cytokine production in tumor-associated macrophages (29). To determine whether miR-155
expression is involved in the secretion of inflammatory cytokines
in macrophages, we detected the mRNA of IL-1β and TNF-α and their
protein levels in supernatant by RT-PCR and ELISA respectively. The
results showed that treatment with the combination of LPS and
S100A8 had a more significant effect compared to other groups
(Fig. 4A), and that the treatment
with LPS and S100A8 increased the mRNA levels with time and
reaching a peak value at 24 h (Fig.
4B). Similarly, after treatment of LPS and S100A8 for 24 h, we
found a significant increase in the levels of IL-1β and TNF-α in
the supernatant compared to the other groups (Fig. 4C). Further analysis showed that the
levels of IL-1β and TNF-α mRNA in the macrophages were reduced
after transfection with the miR-155 inhibitor or pretreatment with
Bay 11-7082 (Fig. 4D) and the
secretion of IL-1β and TNF-α in the supernatant was markedly
decreased after transfection with the miR-155 inhibitor or
pretreatment with Bay 11-7082 (Fig.
4E). Analyzing the above results, it was determined that
recombinant S100A8 protein enhanced the secretion of inflammatory
cytokines in macrophages in the inflammatory microenvironment and
this effect was through activation of NF-κB signaling and aberrant
expression of miR-155.
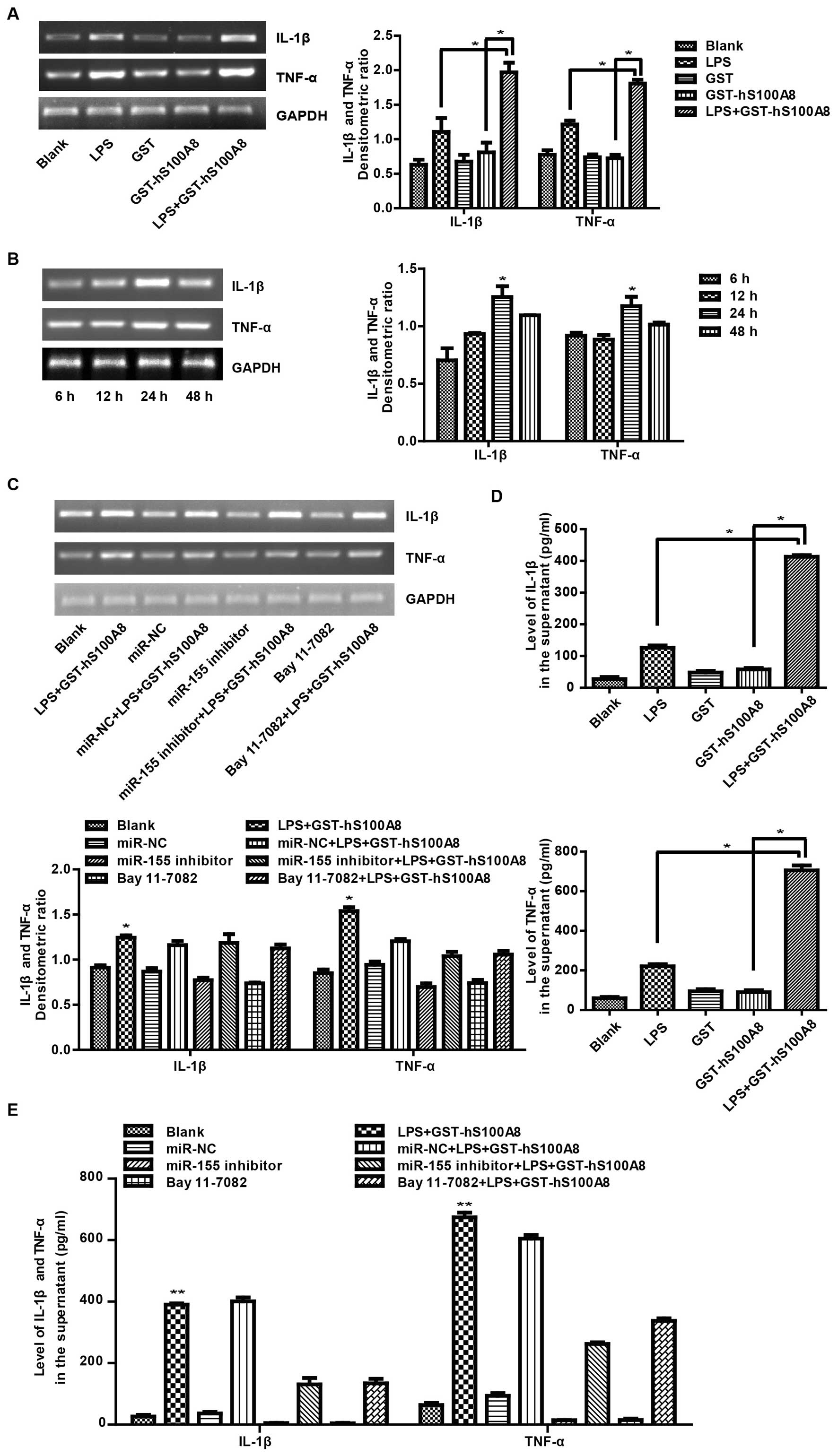 | Figure 4Recombinant S100A8 protein
strengthens the secretion of IL-1β and TNF-α in macrophages in an
inflammatory microenvironment. (A) Macrophages were not treated or
treated with LPS (100 ng/ml), GST (10 µg/ml), GST-hS100A8
(10 µg/ml) or LPS+GST-hS100A8 for 24 h or (B) not treated or
treated with LPS (100 ng/ml), GST-hS100A8 (10 µg/ml) for 6,
12, 24 or 48 h and the mRNA of IL-1β and TNF-α were determined by
RT-PCR. GAPDH was used as an internal reference control. (C)
Macrophages were transfected with miR-155 inhibitor or miR-NC or
pretreated with the inhibitor of NF-κB for a period of 30 min prior
to treatment for 24 h without or with LPS (100 ng/ml), GST-hS100A8
(10 µg/ml) or LPS+GST-hS100A8. Then, mRNA levels of IL-1β
and TNF-α were determined by RT-PCR. GAPDH was used as an internal
reference control. (D) Macrophages were not treated or treated with
LPS (100 ng/ml), GST (10 µg/ml) or GST-hS100A8 (10
µg/ml) for 24 h and IL-1β and TNF-α in the supernatant were
analyzed by ELISA. (E) Macrophages were transfected with miR-155
inhibitor or miR-NC or pretreated with the inhibitor of NF-κB for a
period of 30 min prior to treatment for 24 h without or with LPS
(100 ng/ml), GST-hS100A8 (10 µg/ml) or LPS+GST-hS100A8 and
IL-1β and TNF-α in the supernatant were analyzed by ELISA.
*P≤0.05, **P≤0.01 compared to the control
group. LPS, lipopolysaccharides. |
S100A8 facilitates the migration but not
the viability of CRC cells co-cultured with macrophages through the
activation of NF-κB in the inflammatory microenvironment
Compelling evidence from human and experimental
tumors indicated that tumor-associated macrophages (TAM) release a
broad array of cytokines, growth factors and enzymes to promote the
proliferation and metastasis of cancer cells (30,31),
particularly in spontaneous intestinal tumorigenesis, which has
proven the importance of inflammatory cytokines in facilitating
tumor promotion and progression (5). To investigate the effects of S100A8 on
the viability and migration of CRC cells co-cultured with
macrophages in an inflammatory microenvironment, an MTT assay was
first conducted to measure the proliferation of HCT116 and SW480
cells after treatment with different macrophage CM (Fig. 5A and B). FCM was used to analyze the
cell cycle and Hoechst 33258 fluorescent staining was performed to
detect the cell apoptosis of HCT116 cells co-cultured with
macrophages after the different treatments (data not shown). The
above experimental methods were described in Materials and methods.
It was established that there was no significant difference in
viability among the six groups. Furthermore, a wound healing assay
was used to detect the migration of SW480 cells co-cultured with
macrophages after different treatments. There was a significant
increase in the wound closure rate after being co-cultured with
macrophages which were treated with LPS and S100A8 together
compared with that of the other groups, and the effects were
impaired by Bay 11-7082 (Fig. 5C).
Similar results were observed in HCT116 cells (data not shown). The
effect on cell migration by LPS and S100A8 co-treatment was further
confirmed by Transwell migration assay, which showed that the
number of trans-membrane migrated HCT116 cells after treatment with
LPS and S100A8 was markedly increased compared with that of the
other groups, but its effects were impaired by Bay 11-7082
(Fig. 5D). Similar results were
observed in SW480 cells (data not shown). It is suggested that
inflammatory cytokines produced in excess after the treatment of
LPS and S100A8 in macrophages facilitated the migration of CRC
cells, but there was no significant difference in terms of
viability of CRC cells.
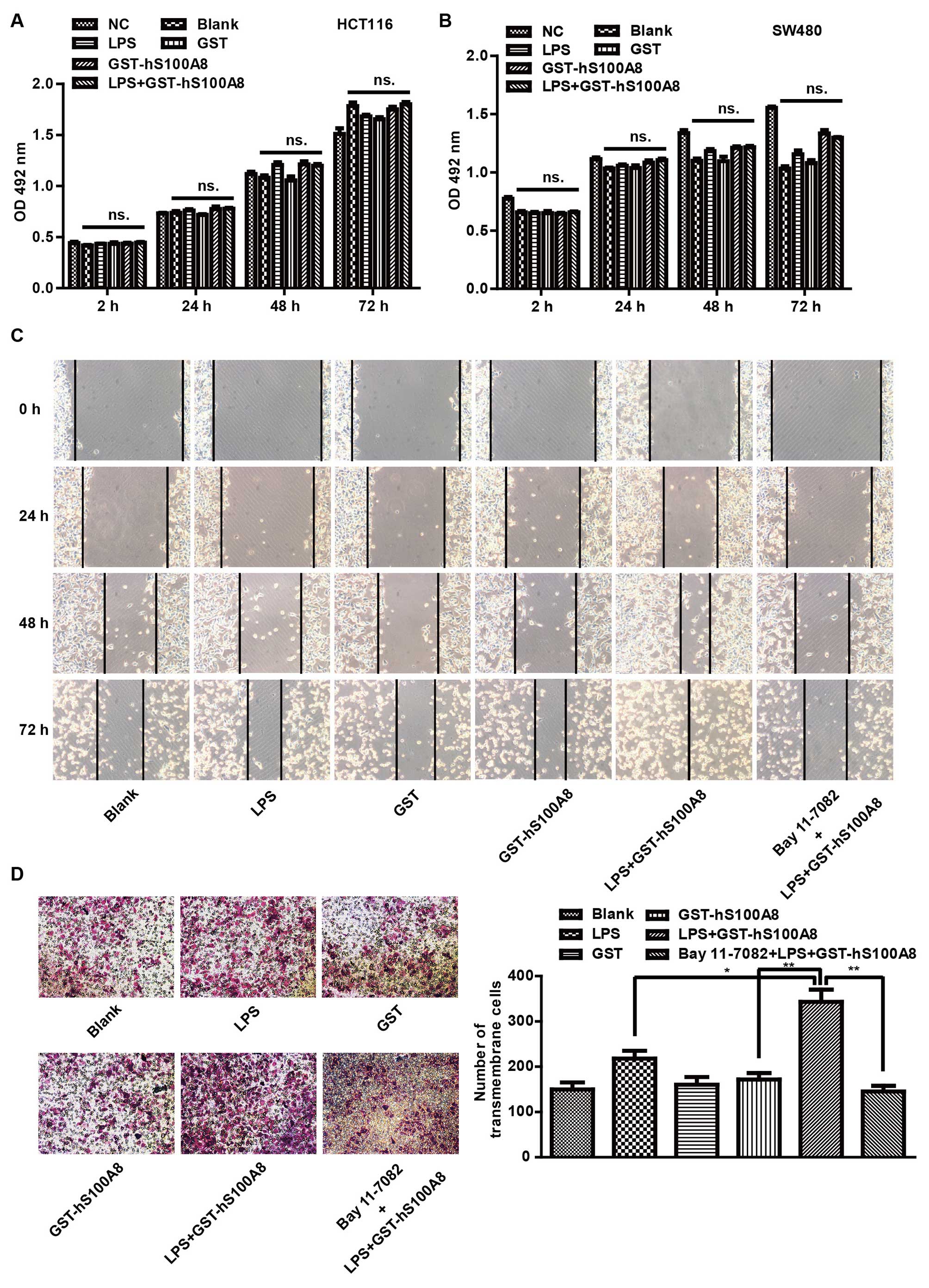 | Figure 5Recombinant S100A8 protein
facilitates the migration of CRC cells co-cultured with macrophages
through activation of NF-κB in an inflammatory microenvironment.
(A) HCT116 and (B) SW480 cells were treated with the different
macrophage conditioned media for 2, 24, 48 and 72 h, and the
viability was measured using the MTT assay. Results are expressed
as the mean absorbances ± SEM of 5 independent experiments. (C)
Wound healing assay for analyzing the effects of S100A8 on the
migration of SW480 cells co-cultured with macrophages for 0, 24, 48
and 72 h which were not treated or treated with LPS (100 ng/ml),
GST and GST-hS100A8 (10 µg/ml) and LPS+GST-hS100A8. The
incision width of different sites was measured. (D) Transwell
migration assay for analyzing the migration of HCT116 cells
co-cultured with macrophages for 24 h which were not treated or
treated with LPS (100 ng/ml), GST or GST-hS100A8 (10 µg/ml)
and LPS+GST-hS100A8. Representative images of transmembrane cells
are shown. The mean of transmembrane cells ± SEM per microscopic
field of 3 independent experiments was calculated. Magnification,
×100. ns., P>0.05; *P≤0.05, **P≤0.01
compared with the control group. CRC colorectal cancer; LPS,
lipopolysaccharides. |
Discussion
Increasing evidence indicates that the expression of
S100A8 is increased in CRC tissues and the elevated expression of
S100A8 is associated not only with histological differentiation,
but also with Dukes' stage and lymph node metastasis in CRC
(9). In the present study, we found
that S100A8 promoted the expression of miR-155 and the secretion of
IL-1β and TNF-α through activation of the NF-κB signaling pathway
in macrophages, eventually facilitating the migration of CRC cells,
which occurred in the inflammation environment mimicked by LPS.
These findings suggest that S100A8 may be an important
proinflammatory factor involved in CRC progression. Aberrant
expression of miR-155 and secretion of inflammatory cytokines may
be involved in the oncogenic properties of S100A8.
The pivotal role of the tumor inflammatory
microenvironment in tumor progression has been generally accepted,
and CRC represents a paradigm, as in CRC there is clear evidence
that persistent inflammation is linked to increasing cancer risk
and tumor development (32). The
tumor inflammatory microenvironment has been shown to be composed
of tumor cells, infiltrating immune cells and various inflammatory
cytokines. The immune cells and macrophages play a crucial role
(4) in the tumor inflammatory
microenvironment. Macrophages possess a high degree of functional
plasticity, and they can alter their functional profiles time and
again in response to environmental changes anywhere between the two
extremes of their phenotypical programs, M1 and M2 polarization
(33). It has been shown that
macrophages are polarized to M1 macrophages when exposed to LPS and
IFn-γ and exhibit an antitumor effect, when exposed to Th2
cytokines (such as IL-4 and IL-13), they are polarized to M2
macrophages and cause protumoral progression (34,35).
Although previous studies indicated that TAMs, which are derived
from circulating monocytes recruited at the tumor site by
chemotactic factors, had several M2-associated protumoral functions
including promotion of tumor cell growth, metastasis and
suppression of adaptive immunity (35,36),
their role regarding tumor progression is still controversial.
THP-1 macrophages were used as a macrophage model because
PMA-treated THP-1 macrophages have an M2 functional profile
(35,37). Hence, in this study, we used PMA to
induce THP-1 monocytes to differentiate into macrophages with M2
functional profiles and investigated the effect of S100A8 in the
modulation of macrophages and CRC cells in the inflammatory
microenvironment.
Several studies have reported that CRC cells may be
exposed to increased levels of LPS because of bacterial
translocation (38) or systemic
endogenous LPS without infections (39). It is suggested that LPS is an
important component in the inflammatory microenvironment. Although
LPS causes significant systemic inflammation and increases hepatic
recruitment and liver metastasis by CRC cells through TLR4
signaling (10), the impact of
S100A8 in the modulation of macrophages in an LPS mimicking
inflammatory microenvironment is very limited. Previous studies
have shown that proliferation of differentiated THP-1 macrophages
is stopped and adherence is not very firm, thus after the cells
were treated with LPS and S100A8 for 3 successive days, the
viability of the macrophages was decreased. The results suggested
that different concentrations of LPS and S100A8 may have no
significant effect on the viability of differentiated THP-1
macrophages.
It is well known that the NF-κB signaling pathway
plays a crucial role in immune-regulation and inflammatory response
via the induction of inflammatory cytokines (such as IL-6, IL-1β
and TNF-α), cyclooxygenase-2 (COX-2) and inducible nitric oxide
synthase (iNOS) (40). For
instance, transcriptional activity of NF-κB may be most critical in
S100A8/A9-induced expression of proinflammatory cytokines in
monocytes/macrophages (41). S100A9
promoted human embryo lung fibroblasts to secrete proinflammatory
cytokines through activation of the NF-κB pathways (42). Additionally, S100A4 induced an
inflammatory response partially mediated by activation of NF-κB in
mononuclear cells from patients with RA (43). On the contrary, bergamot juice can
inhibit LPS-induced proinflammatory cytokines involved in the
inhibition of NF-κB activation (40). In accordance with these studies, our
data showed that S100A8 significantly activated the NF-κB pathway
in macrophages in the inflammatory microenvironment and promoted
both gene expression and secretion of inflammatory cytokines (IL-1β
and TNF-α). Moreover, an inhibitor of NF-κB partially reversed the
activation of NF-κB and expression of inflammatory cytokines
induced by S100A8 in the inflammatory microenvironment. Several
studies have reported that S100A8 activated NF-κB in a
TLR4-dependent manner (25,44). Although we found that LPS and/or
S100A8 treatment could elevate the activation of TLR4, to
substantiate this idea, neutralization of TLR4 should be used to
block TLR4 signaling and elucidate the mechanisms of S100A8 in CRC
progression through the TLR4/NF-κB signaling pathway. This will be
investigated in our subsequent studies.
miR-155 is considered as an important oncomir and
was first found within the third exon of B-cell integration cluster
(BIC) gene (45) and is highly
expressed in a variety of solid tumors, such as breast, lung and
colon cancer (46–48). Recent research demonstrated that
miR-155 was found to be induced by diverse TLR ligands through the
NF-κB signaling pathways (49). In
our study, we found that S100A8 upregulated the expression of
miR-155 in a time-dependent manner in the inflammatory
microenvironment. This effect was dependent on the activation of
the NF-κB pathway. This confirmed the theory that the promoter
region of pri-miR-155 contains putative NF-κB-binding sites and
NF-κB may potentially modulate the miR-155 expression under certain
circumstances (19–22). In addition, we found that aberrant
miR-155 expression controlled gene expression of IL-1β and TNF-α
and secretions of IL-1β and TNF-α in macrophages. It is possible
that miR-155 post-transcriptionally modulates the expression of
multiple target genes, including suppressor of cytokine signaling 1
(SOCS1) (16), forkhead box O3
(FOXO3a) (50), transcription
factor CCAAT/enhancer binding protein β (C/EBPβ) (51), which were proven to regulate the
expression of various inflammatory cytokines. Moreover, some
studies have reported that TNF-α can significantly increase miR-155
expression in macrophages (52). It
is consistent with the idea that increasing miR-155 production
which is subjected to some proinflammatory mediators may be
attributed to an autocrine loop (53).
Previous studies have reported that TAMs could
directly activate tumor-promoting genes [such as COX-2, epidermal
growth factor receptor (EGFR) and matrix metalloproteinase (MMP)]
in cancer cells (54). Extensive
evidence has confirmed that the inflammatory cytokines, secreted by
macrophages, have a complicated role in the pathogenesis of IBD and
CRC (5). Interestingly,
inflammatory factors were not in equivalent levels in the
intratumoral and peritumoral sites. IL-1β, GM-CSF, G-CSF, TnF-α,
and IL-6 were significantly increased in tumor tissues compared
with adjacent non-tumor tissues (55,56).
IL-6 and IL-8 released from myofibroblasts were found to upregulate
S100A8/A9 in tumor-infiltrated myeloid cells (57). However, our aforementioned data
revealed that S100A8 enhanced the secretion of IL-1β and TNF-α in
macrophages in the inflammatory microenvironment. In subsequent
studies, we tested the the effects of S100A8 on the viability and
migration of HCT116 and SW480 cells with macrophage CM or
co-cultured with macrophages in the inflammatory microenvironment.
Macrophage CM had no significant difference on proliferation in
both cell lines as well as cell cycle and apoptosis, whereas
migration was significantly increased in cells co-cultured with the
macrophages stimulated by LPS and S100A8. These results are
consistent with the notion that tumor-induced systemic inflammation
contributes to metastases without influencing primary tumor
progression (58). In the future,
it is necessary to ascertain the effect of S100A8 on the
development of CRC in xenograft tumor growth and lung metastasis in
animal models.
Collectively, our data showed that S100A8
upregulated the expression of miR-155 through the activation of
NF-κB by differentiating THP-1 macrophages in the inflammatory
microenvironment, leading to the accumulation of the inflammatory
cytokines IL-1β and TNF-α, and ultimately facilitating the
migration of colorectal cancer cells.
Acknowledgments
This study was supported by Chongqing Graduate
Student Research Innovation Project Funding (no. CYS15133). We
would like to thank T.C He (Medical Center, The University of
Chicago) for the kind provision of the recombinant pGST-moluc,
pGST-moluc-hS100A8 and pNF-κB-luc plasmid.
References
|
1
|
Wang D, Fu L, Sun H, Guo L and Dubois RN:
Prostaglandin E2 promotes colorectal cancer stem cell expansion and
metastasis in mice. Gastroenterology. 149:1884–1895.e4. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Van den Eynden GG, Majeed AW, Illemann M,
Vermeulen PB, Bird NC, Høyer-Hansen G, Eefsen RL, Reynolds AR and
Brodt P: The multifaceted role of the microenvironment in liver
metastasis: biology and clinical implications. Cancer Res.
73:2031–2043. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang W, Li X, Zheng D, Zhang D, Huang S,
Zhang X, Ai F, Wang X, Ma J, Xiong W, et al: Dynamic changes of
peritoneal macrophages and subpopulations during ulcerative colitis
to metastasis of colorectal carcinoma in a mouse model. Inflamm
Res. 62:669–680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morales C, Rachidi S, Hong F, Sun S,
Ouyang X, Wallace C, Zhang Y, Garret-Mayer E, Wu J, Liu B, et al:
Immune chaperone gp96 drives the contributions of macrophages to
inflammatory colon tumorigenesis. Cancer Res. 4:446–459. 2014.
View Article : Google Scholar
|
|
5
|
De Simone V, Franzè E, Ronchetti G,
Colantoni A, Fantini MC, Di Fusco D, Sica GS, Sileri P, MacDonald
TT, Pallone F, et al: Th17-type cytokines, IL-6 and TNF-α
synergistically activate STAT3 and NF-κB to promote colorectal
cancer cell growth. Oncogene. 34:3493–3503. 2015. View Article : Google Scholar
|
|
6
|
Jia XH, Feng GW, Wang ZL, Du Y, Shen C,
Hui H, Peng D, Li ZJ, Kong DL and Tian J: Activation of mesenchymal
stem cells by macrophages promotes tumor progression through immune
suppressive effects. Oncotarget. Mar 14–2016.Epub ahead of print.
View Article : Google Scholar
|
|
7
|
Vrakas CN, O'Sullivan RM, Evans SE, Ingram
DA, Jones CB, Phuong T and Kurt RA: The Measure of DAMPs and a role
for S100A8 in recruiting suppressor cells in breast cancer lung
metastasis. Immunol Invest. 44:174–188. 2015. View Article : Google Scholar
|
|
8
|
Chen B, Miller AL, Rebelatto M, Brewah Y,
Rowe DC, Clarke L, Czapiga M, Rosenthal K, Imamichi T, Chen Y, et
al: S100A9 induced inflammatory responses are mediated by distinct
damage associated molecular patterns (DAMP) receptors in vitro and
in vivo. PLoS One. 10:e01158282015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duan L, Wu R, Ye L, Wang H, Yang X, Zhang
Y, Chen X, Zuo G, Zhang Y, Weng Y, et al: S100A8 and S100A9 are
associated with colorectal carcinoma progression and contribute to
colorectal carcinoma cell survival and migration via Wnt/β-catenin
pathway. PLoS One. 8:e620922013. View Article : Google Scholar
|
|
10
|
Stulík J, Osterreicher J, Koupilová K,
Knízek, Macela A, Bures J, Jandík P, Langr F, Dedic K and Jungblut
PR: The analysis of S100A9 and S100A8 expression in matched sets of
macroscopically normal colon mucosa and colorectal carcinoma: the
S100A9 and S100A8 positive cells underlie and invade tumor mass.
Electrophoresis. 20:1047–1054. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang P and Li Z, Li H, Lu Y, Wu H and Li
Z: Pyruvate kinase M2 accelerates pro-inflammatory cytokine
secretion and cell proliferation induced by lipopolysaccharide in
colorectal cancer. Cell Signal. 27:1525–1532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lujambio A and Lowe SW: The microcosmos of
cancer. Nature. 482:347–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elton TS, Selemon H, Elton SM and
Parinandi NL: Regulation of the MIR155 host gene in physiological
and pathological processes. Gene. 532:1–12. 2013. View Article : Google Scholar
|
|
16
|
Pathak S, Grillo AR, Scarpa M, Brun P,
D'Incà R, Nai L, Banerjee A, Cavallo D, Barzon L, Palù G, et al:
MiR-155 modulates the inflammatory phenotype of intestinal
myofibroblasts by targeting SOCS1 in ulcerative colitis. Exp Mol
Med. 47:e1642015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mashima R: Physiological roles of miR-155.
Immunology. 145:323–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jin HM, Kim TJ, Choi JH, Kim MJ, Cho YN,
Nam KI, Kee SJ, Moon JB, Choi SY, Park DJ, et al: MicroRNA-155 as a
proinflammatory regulator via SHIP-1 down-regulation in acute gouty
arthritis. Arthritis Res Ther. 16:R882014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thompson RC, Vardinogiannis I and Gilmore
TD: Identification of an NF-κB p50/p65-responsive site in the human
MIR155HG promoter. BMC Mol Biol. 14:242013. View Article : Google Scholar
|
|
20
|
Quinn SR, Mangan NE, Caffrey BE, Gantier
MP, Williams BR, Hertzog PJ, McCoy CE and O'Neill LA: The role of
Ets2 transcription factor in the induction of microRNA-155
(miR-155) by lipopolysaccharide and its targeting by
interleukin-10. J Biol Chem. 289:4316–4325. 2014. View Article : Google Scholar :
|
|
21
|
Wang B, Majumder S, Nuovo G, Kutay H,
Volinia S, Patel T, Schmittgen TD, Croce C, Ghoshal K and Jacob ST:
Role of microRNA-155 at early stages of hepatocarcinogenesis
induced by choline-deficient and amino acid-defined diet in C57BL/6
mice. Hepatology. 50:1152–1161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Curtis AM, Fagundes CT, Yang G,
Palsson-McDermott EM, Wochal P, McGettrick AF, Foley NH, Early JO,
Chen L, Zhang H, et al: Circadian control of innate immunity in
macrophages by miR-155 targeting Bmal1. Proc Natl Acad Sci USA.
112:7231–7236. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
You L, Xu LL, Guo YY, Zou ZY, Li YY, Sun
SS, Luo JY and Zhou L: Prokaryotic expression, purification and
identification of GST-human S100A9 fusion protein. Chin J Biochem
Pharm. 32:253–256. 2011.
|
|
24
|
Zhao C, Wu N, Deng F, Zhang H, Wang N,
Zhang W, Chen X, Wen S, Zhang J, Yin L, et al: Adenovirus-mediated
gene transfer in mesenchymal stem cells can be significantly
enhanced by the cationic polymer polybrene. PLoS One. 9:e929082014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deguchi A, Tomita T, Ohto U, Takemura K,
Kitao A, Akashi-Takamura S, Miyake K and Maru Y: Eritoran inhibits
S100A8-mediated TLR4/MD-2 activation and tumor growth by changing
the immune microenvironment. Oncogene. 35:1445–1456. 2016.
View Article : Google Scholar
|
|
26
|
Hiratsuka S, Watanabe A, Sakurai Y,
Akashi-Takamura S, Ishibashi S, Miyake K, Shibuya M, Akira S,
Aburatani H and Maru Y: The S100A8-serum amyloid A3-TLR4 paracrine
cascade establishes a pre-metastatic phase. Nat Cell Biol.
10:1349–1355. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tili E, Croce CM and Michaille JJ:
miR-155: On the crosstalk between inflammation and cancer. Int Rev
Immunol. 28:264–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moldoveanu AC, Diculescu M and Braticevici
CF: Cytokines in inflammatory bowel disease. Rom J Intern Med.
53:118–127. 2015.PubMed/NCBI
|
|
29
|
Kurowska-Stolarska M, Alivernini S,
Ballantine LE, Asquith DL, Millar NL, Gilchrist DS, Reilly J, Ierna
M, Fraser AR, Stolarski B, et al: MicroRNA-155 as a proinflammatory
regulator in clinical and experimental arthritis. Proc Natl Acad
Sci USA. 108:11193–11198. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pollard JW: Tumour-educated macrophages
promote tumour progression and metastasis. Nat Rev Cancer. 4:71–78.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mantovani A, Romero P, Palucka AK and
Marincola FM: Tumour immunity: effector response to tumour and role
of the microenvironment. Lancet. 371:771–783. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Erreni M, Mantovani A and Allavena P:
Tumor-associated Macrophages (TAM) and Inflammation in Colorectal
Cancer. Cancer Microenviron. 4:141–154. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Watkins SK, Egilmez NK, Suttles J and
Stout RD: IL-12 rapidly alters the functional profile of
tumor-associated and tumor-infiltrating macrophages in vitro and in
vivo. J Immunol. 178:1357–1362. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Edin S, Wikberg ML, Dahlin AM, Rutegård J,
Öberg Å, Oldenborg PA and Palmqvist R: The distribution of
macrophages with a M1 or M2 phenotype in relation to prognosis and
the molecular characteristics of colorectal cancer. PLoS One.
7:e470452012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tjiu JW, Chen JS, Shun CT, Lin SJ, Liao
YH, Chu CY, Tsai TF, Chiu HC, Dai YS, Inoue H, et al:
Tumor-associated macrophage-induced invasion and angiogenesis of
human basal cell carcinoma cells by cyclooxygenase-2 induction. J
Invest Dermatol. 129:1016–1025. 2009. View Article : Google Scholar
|
|
36
|
Balkwill F, Charles KA and Mantovani A:
Smoldering and polarized inflammation in the initiation and
promotion of malignant disease. Cancer Cell. 7:211–217. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Daigneault M, Preston JA, Marriott HM,
Whyte MK and Dockrell DH: The identification of markers of
macrophage differentiation in PMA-stimulated THP-1 cells and
monocyte-derived macrophages. PLoS One. 5:e86682010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Konstantinov SR, Kuipers EJ and
Peppelenbosch MP: Functional genomic analyses of the gut microbiota
for CRC screening. Nat Rev Gastroenterol Hepatol. 10:741–745. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Maddocks OD, Short AJ, Donnenberg MS,
Bader S and Harrison DJ: Attaching and effacing Escherichia coli
down-regulate DNA mismatch repair protein in vitro and are
associated with colorectal adenocarcinomas in humans. PLoS One.
4:e55172009. View Article : Google Scholar
|
|
40
|
Risitano R, Currò M, Cirmi S, Ferlazzo N,
Campiglia P, Caccamo D, Ientile R and Navarra M: Flavonoid fraction
of bergamot juice reduces LPS-induced inflammatory response through
SIRT1-mediated NF-κB inhibition in THP-1 monocytes. PLoS One.
9:e1074312014. View Article : Google Scholar
|
|
41
|
Sunahori K, Yamamura M, Yamana J, Takasugi
K, Kawashima M, Yamamoto H, Chazin WJ, Nakatani Y, Yui S and Makino
H: The S100A8/A9 heterodimer amplifies proinflammatory cytokine
production by macrophages via activation of nuclear factor kappa B
and p38 mitogen-activated protein kinase in rheumatoid arthritis.
Arthritis Res Ther. 8:R692006. View
Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu X, Chen H, Zhu X, Ma Y, Liu Q, Xue Y,
Chu H, Wu W, Wang J and Zou H: S100A9 promotes human lung
fibroblast cells activation through receptor for advanced glycation
end-product-mediated extracellular-regulated kinase 1/2,
mitogen-activated protein-kinase and nuclear factor-κB-dependent
pathways. Clin Exp Immunol. 173:523–535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cerezo LA, Remáková M, Tomčik M, Gay S,
Neidhart M, Lukanidin E, Pavelka K, Grigorian M, Vencovský J and
Šenolt L: The metastasis-associated protein S100A4 promotes the
inflammatory response of mononuclear cells via the TLR4 signalling
pathway in rheumatoid arthritis. Rheumatology (Oxford).
53:1520–1526. 2014. View Article : Google Scholar
|
|
44
|
Austermann J, Friesenhagen J, Fassl SK,
Petersen B, Ortkras T, Burgmann J, Barczyk-Kahlert K, Faist E,
Zedler S, Pirr S, et al: Alarmins MRP8 and MRP14 induce stress
tolerance in phagocytes under sterile inflammatory conditions. Cell
Reports. 9:2112–2123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
O'Connell RM, Taganov KD, Boldin MP, Cheng
G and Baltimore D: MicroRNA-155 is induced during the macrophage
inflammatory response. Proc Natl Acad Sci USA. 104:1604–1609. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bertoli G, Cava C and Castiglioni I:
MicroRNAs: new biomarkers for diagnosis, prognosis, therapy
prediction and therapeutic tools for breast cancer. Theranostics.
5:1122–1143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xie K, Ma H, Liang C, Wang C, Qin N, Shen
W, Gu Y, Yan C, Zhang K, Dai N, et al: A functional variant in
miR-155 regulation region contributes to lung cancer risk and
survival. Oncotarget. 6:42781–42792. 2015.PubMed/NCBI
|
|
48
|
Qu YL, Wang HF, Sun ZQ, Tang Y, Han XN, Yu
XB and Liu K: Up-regulated miR-155-5p promotes cell proliferation,
invasion and metastasis in colorectal carcinoma. Int J Clin Exp
Pathol. 8:6988–6994. 2015.PubMed/NCBI
|
|
49
|
Koch M, Mollenkopf HJ, Klemm U and Meyer
TF: Induction of microRNA-155 is TLR-and type IV secretion
system-dependent in macrophages and inhibits DNA-damage induced
apoptosis. Proc Natl Acad Sci USA. 109:E1153–E1162. 2012.
View Article : Google Scholar
|
|
50
|
Min M, Peng L, Yang Y, Guo M, Wang W and
Sun G: MicroRNA-155 is involved in the pathogenesis of ulcerative
colitis by targeting FOXO3a. Inflamm Bowel Dis. 20:652–659. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Worm J, Stenvang J, Petri A, Frederiksen
KS, Obad S, Elmén J, Hedtjärn M, Straarup EM, Hansen JB and
Kauppinen S: Silencing of microRNA-155 in mice during acute
inflammatory response leads to derepression of c/ebp Beta and
down-regulation of G-CSF. Nucleic Acids Res. 37:5784–5792. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang RN, Zheng B, Li LM, Zhang J, Zhang
XH and Wen JK: Tongxinluo inhibits vascular inflammation and
neointimal hyperplasia through blockade of the positive feedback
loop between miR-155 and TNF-α. Am J Physiol Heart Circ Physiol.
307:H552–H562. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bala S, Marcos M, Kodys K, Csak T,
Catalano D, Mandrekar P and Szabo G: Up-regulation of microRNA-155
in macrophages contributes to increased tumor necrosis factor α
(TNF-α) production via increased mRNA half-life in alcoholic liver
disease. J Biol Chem. 286:1436–1444. 2011. View Article : Google Scholar
|
|
54
|
Cardoso AP, Pinto ML, Pinto AT, Oliveira
MI, Pinto MT, Gonçalves R, Relvas JB, Figueiredo C, Seruca R,
Mantovani A, et al: Macrophages stimulate gastric and colorectal
cancer invasion through EGFR Y1086, c-Src, Erk1/2 and Akt
phosphorylation and small GTPase activity. Oncogene. 33:2123–2133.
2014. View Article : Google Scholar
|
|
55
|
He G, Zhang H, Zhou J, Wang B, Chen Y,
Kong Y, Xie X, Wang X, Fei R, Wei L, et al: Peritumoural
neutrophils negatively regulate adaptive immunity via the
PD-L1/PD-1 signalling pathway in hepatocellular carcinoma. J Exp
Clin Cancer Res. 34:1412015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li X, Liu C, Ip BC, Hu KQ, Smith DE,
Greenberg AS and Wang XD: Tumor progression locus 2 ablation
suppressed hepatocellular carcinoma development by inhibiting
hepatic inflammation and steatosis in mice. J Exp Clin Cancer Res.
34:1382015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kim JH, Oh SH, Kim EJ, Park SJ, Hong SP,
Cheon JH, Kim TI and Kim WH: The role of myofibroblasts in
upregulation of S100A8 and S100A9 and the differentiation of
myeloid cells in the colorectal cancer microenvironment. Biochem
Biophys Res Commun. 423:60–66. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Coffelt SB, Kersten K, Doornebal CW,
Weiden J, Vrijland K, Hau CS, Verstegen NJ, Ciampricotti M,
Hawinkels LJ, Jonkers J, et al: IL-17-producing γδ T cells and
neutrophils conspire to promote breast cancer metastasis. Nature.
522:345–348. 2015. View Article : Google Scholar : PubMed/NCBI
|