Introduction
Normal physiology depends on an adequate blood
supply. Angiogenesis is the formation of new vessels from
pre-existing vessels. It is crucial to the development of embryos
as well as several physiologic and pathologic processes including
the female reproductive cycle, wound healing and the growth of
solid tumors (1,2). Endothelial cells (ECs) play an
important role in angiogenesis and maintain vascular homeostasis
(3). Compelling evidence has shown
that vascular endothelial growth factor (VEGF) promotes endothelial
cell proliferation and is deregulated in most vascular diseases and
cancers (4,5). However, the precise mechanism of the
regulation of VEGF in human diseases particularly tumors remains
not fully understood.
MicroRNAs (miRNAs) are highly conserved,
single-stranded non-coding small RNAs, which regulate gene
expression at the post-transcriptional level by inhibiting protein
translation from mRNAs or by promoting degradation of mRNAs
(6,7). Numerous miRNAs are aberrantly
expressed in cancer tissues and play a critical role in
tumorigenesis and anticancer drug resistance (8,9).
miRNAs function as either tumor suppressors or oncogenes
(oncomiRs). miRNA-7 (miR-7) was found to display tumor-suppressive
function in colon cancer by targeting oncogenic YY1 (10), and in hepatocarcinoma via
suppression of CCNE1 expression (11). Recently, a lentiviral-based miRNA
library screening indicated that miR-7 is an anti-angiogenic miRNA,
which was further validated in an animal model (12), suggesting that miR-7 is a novel
anti-angiogenic therapeutic target. However, the underlying
molecular mechanism of the suppression of angiogenesis by miR-7 is
largely unknown.
Krüppel-like factors (KLFs) are a family of
evolutionarily conserved zinc finger-containing transcription
factors that bind 'CACCC' or 'GT-box' sites through a DNA-binding
domain consisting of three zinc fingers positioned at their
carboxyl-terminal end (13).
Krüppel-like factor 4 (KLF4), Oct3/4, Sox2 and c-Myc are the four
defined critical factors inducing differentiated cells into
pluripotent stem cells (14).
Mounting evidence indicates that KLF4 is highly expressed in ECs
and vascular smooth muscle cells (VSMCs) and acts as a critical
regulator of vascular homeostasis (15). Diverse vasoprotective stimuli,
including laminar shear stress and simvastatin, induce the
expression of KLF4 in ECs (16,17).
Considering the overexpression of KLF4 in ECs and the
anti-angiogenic function of miR-7, we hypothesized that
downregulation of miR-7 may promote EC angiogenesis by targeting
KLF4.
In the present study, we knocked down miR-7 by
miR-7-specific antagomir in HUVECs and assessed angiogenesis. We
found that miR-7 silencing resulted in enhanced angiogenesis,
accompanied by upregulation of KLF4 and VEGF, revealing a novel
angiogenesis-related miR-7-KLF4-VEGF signaling axis.
Materials and methods
Cell culture and transfection
HUVECs were obtained from the American Type Culture
Collection (ATCC; Manassas, VA, USA) and cultured in Endothelial
Cell Growth Medium-2 (EGM-2; Lonza, Portsmouth, NH, USA) at 37°C in
a humidified atmosphere of 5% CO2 as previously
described (18). The HUVECs were
transfected with oligonucleotides in 6-well plates using
Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's instructions. Briefly, 100 pmol of
miR-7 mimics, miR-7 inhibitors (miR-ASO) or small interfering RNAs
(siRNAs) against KLF4 (GenePharma, Shanghai, China) were formulated
with 500 µl serum-free RPMI-1640 medium (Invitrogen). The
transfection complex was directly added to the cells and replaced
with fresh complete medium 6 h later. Analyses of the effects of
miRNAs or siRNAs on recipient cells were performed 48 h after
transfection as previously described (11,19).
Plasmid construction and luciferase
report assay
The 3′-untranslated region (3′-UTR) of KLF4 was
amplified from human genomic DNA and cloned into the pGL3-control
vector (Promega, Madison, WI, USA) (19). Mutation of the miR-7 binding core
sequence on the 3′-UTR region of the KLF4 gene was achieved using
overlap extension PCR methods using primers as previously reported
(19). HUVECs were seeded in
24-well plates and then transfected with 500 ng of wild-type or
mutant reporter plasmid, 25 ng of Renilla luciferase
expression vector, along with miR-7 mimics (10 nM) or the negative
control (10 nM) using Lipofectamine 2000 reagent. After 48 h, the
cell extract was used to determine firefly and Renilla
luciferase activities with the Dual-Luciferase Reporter Assay
system (Promega).
Quantitative real-time PCR
Total RNA was isolated from the cells using TRIzol
reagent (Invitrogen) according to the manufacturer's instructions.
RNAs were reverse transcribed into cDNA using miScript Reverse
Transcription kit (Qiagen, Valencia, CA, USA). All PCR conditions
were optimized to produce a single product in the exponential
range. Quantitative real-time PCR was performed using SYBR-Green
Master Mix and analyzed using an ABI Step One Real-Time PCR system
(Applied Biosystems, Foster City, CA, USA). GAPDH was used as an
internal loading control. Quantitative RT-PCR for KLF4, VEGF and
GAPDH was performed using the following primers: KLF4 forward,
5′-CCCACATGAAGCGACTTCCC-3′ and reverse 5′-CAGGTCCAGGAGATCGTTGAA-3′;
VEGF forward, 5′-AGGGCAGAATCATCACGAAGT-3′ and reverse
5′-AGGGTCTCGATTGGATGGCA-3′); GAPDH forward,
5′-TCACCAGGGCTGCTTTTAAC-3′ and reverse,
5′-GACAAGCTTCCCGTTCTCAG-3′.
After placing the PCR tubes into the thermal cycler,
the process entailed an initial denaturation step at 95°C for 10
min followed by a second step at 95°C for 15 sec and 60°C for 30
sec for 40 cycles with a melting curve analysis. The level of
target mRNA was normalized to the level of GAPDH and was compared
with the control. For the detection of miR-7, miRNA from cultured
cells was extracted by mirVana miRNA Isolation kit (Ambion,
Carlsbad, CA, USA) according to the manufacturer's instructions.
TaqMan MicroRNA Reverse Transcription kit was used to produce cDNA
for TaqMan MicroRNA assay (Applied Biosystems). Human miR-7
expression was quantified in real-time with FAM dye-labeled
MGB-probes and normalized to snU6 (Applied Biosystems). Data were
analyzed using the ΔΔCT method.
Tube formation assay
HUVECs were seeded in a 6-well plate
(8×104 cells/well) and transfected with 50 nM miRNA
mimics with Lipofectamine 2000 reagent in serum-free medium. After
4 h, the medium was replaced with complete EGM-2 medium. A 96-well
plate was coated with 50 µl Matrigel™ (BD Biosciences, San
Jose, CA, USA) and incubated for 30 min at 37°C. After incubation
for 48 h, the transfected HUVECs were counted, prepared in a cell
suspension in EGM-2 medium and seeded on top of the Matrigel™
(7,500 cells/well). After incubation for 17 h, the wells were
imaged with a Nikon TE2000 microscope at a magnification of ×10.
Tubule formation was quantified by counting the number of branch
points in at least three independent wells under similar conditions
using NIH ImageJ software.
ELISA detection of VEGF levels
The cell growth-conditioned medium from the above
experiments was collected and analyzed for VEGF levels with
immunosorbent assay (ELISA) kits (ACEL, Kanagawa, Japan) according
to the manufacturer's instructions. The data are presented as mean
± SEM.
Chromatin immunoprecipitation assay
(ChIP)
ChIP was performed as previously described (20). Briefly, HUVECs were grown and then
washed twice with phosphate-buffered saline (PBS) and cross-linked
using 1% formaldehyde for 10 min. Following lysis of the cells and
sonication, DNA-protein complexes in the lysates were subjected to
immunoprecipitation using anti-KLF4 (HPA002926; Sigma, St. Louis,
MO, USA) or control normal IgG. After precipitation of the
immunocomplex with protein G-agarose, isolated DNA was used as a
template in PCR with specific primers spanning the target region of
the VEGF promoter. Primers used for amplification of the human VEGF
promoter were as follows: VEGF-1 (−0.6 kb) forward,
5′-GCAGACGGCAGTCACTAGG-3′ and reverse, 5′-CACTCTCGAAGACGCTGCT-3′;
VEGF-2 (−3.1 kb) forward, 5′-GAACATTTGGGAAATCTCTTGC-3′ and reverse
5′-TGTGGCTCTGAAGGCAGTT-3′; VEGF-3 (−3.2 kb) forward,
5′-GCCTTGGCCCAATCACTAC-3′ and reverse,
5′-GTGGAGAAAAGTGCGAAAGG-3′.
Lentiviral packaging
The cDNA of human transcription factor KLF4
(GenBank: BC030811/029923) was amplified by PCR, cloned into the
vector pMD-18T (Takara, Dalian, China) and confirmed by DNA
sequencing (20,21). The KLF4 open reading frame (ORF) was
then subcloned into the lentiviral vector pLenti-6.3 (Invitrogen).
Viral packaging and infection were performed according to the
standard protocols as recommended by the manufacturer
(Invitrogen).
Western blotting
Total protein was extracted from the cells using
lysis buffer containing 20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 5 mM
EDTA, 1% Triton X-100, 1% DTT and 1% protease inhibitor cocktail
(Roche). Equal amounts of protein extracts (40 µg) were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene
fluoride (PVDF) membrane. Membranes were blocked with 5% w/v
non-fat dry milk dissolved in Tris-buffered saline plus Tween-20
(TBS-T; 0.1% Tween-20; pH 8.3) at room temperature for 1 h, then
incubated with primary antibodies at 4°C overnight. The primary
antibodies used were rabbit anti-KLF4 and anti-β-actin (Sigma).
After washing with TBS-T, the membranes were incubated with
horseradish peroxidase (HRP)-labeled secondary antibodies (Sigma)
for 30-45 min at room temperature. Immunobands were visualized
using enhanced chemiluminescence (ECL) kit (GE Healthcare,
Waukesha, WI, USA) according to the manufacturer's instructions and
exposed to X-ray film. β-actin was used as a loading control.
Statistical analysis
All statistical analyses were performed using the
GraphPad Prism software, version 4 (GraphPad Software, San Diego
CA, USA). Data are expressed as mean ± SEM. Comparison of the means
was performed using two-sided Student's t-test and a P<0.05
value was considered to indicate a statistically significant
result.
Results
Downregulation of miR-7 promotes the
angiogenesis of HUVECs and induces the expression of VEGF and
KLF4
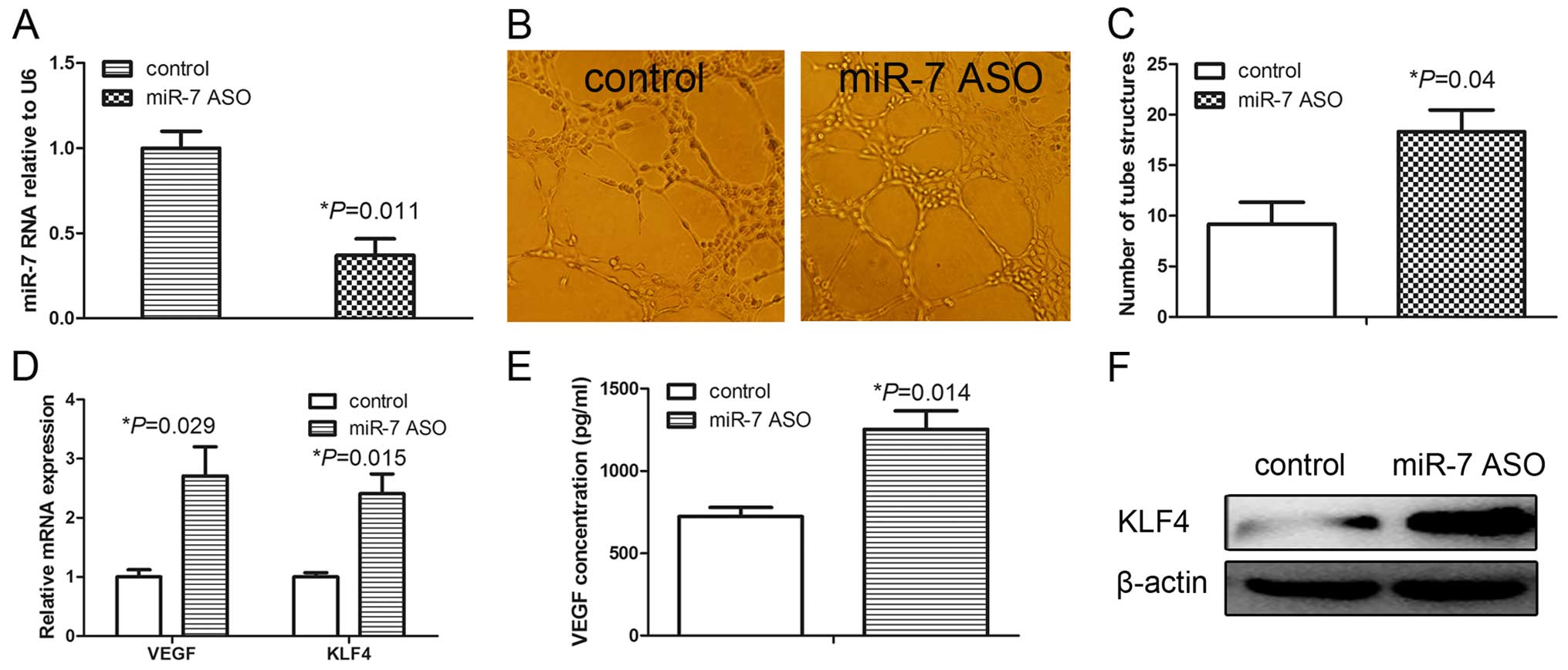
We assessed the role of miR-7 in the angiogenesis of
HUVECs by first transfecting HUVECs with a miR-7 inhibitor (miR-7
ASO). As shown in Fig. 1A,
transient transfection of miR-7 ASO significantly decreased the
miR-7 expression compared with that of the control. The tube
formation assay was performed to evaluate the effect of miR-7 on
the angiogenic potential of HUVECs. Knockdown of miR-7
significantly induced the tube formation of HUVECs (Fig. 1B and C) suggesting that miR-7
inhibition resulted in enhanced angio-genesis in the HUVECs.
VEGF plays a key role in endothelial cell migration
and proliferation, and angiogenesis. We consequently assessed the
VEGF expression following miR-7 inhibition. qRT-PCR analyses showed
that VEGF mRNA levels were significantly increased in the HUVECs
treated with miR-7 ASO (Fig. 1D).
ELISA analyses further demonstrated significantly upregulated VEGF
protein levels in the cell culture medium from the HUVECs treated
with miR-7 ASO (Fig. 1E). Previous
studies indicated that KLF4 is overexpressed in ECs and plays an
important role in angiogenesis (15,16).
We further assessed the KLF4 expression following miR-7 inhibition
in the HUVECs and found that the mRNA levels of KLF4 were
significantly induced by miR-7 ASO (Fig. 1D). Similarly, the KLF4 protein level
was markedly upregulated in the HUVECs treated with miR-7 ASO
(Fig. 1F). These results
demonstrated that downregulation of miR-7 promoted angiogenesis of
HUVECs, accompanied by the upregulation of the expression of VEGF
and KLF4.
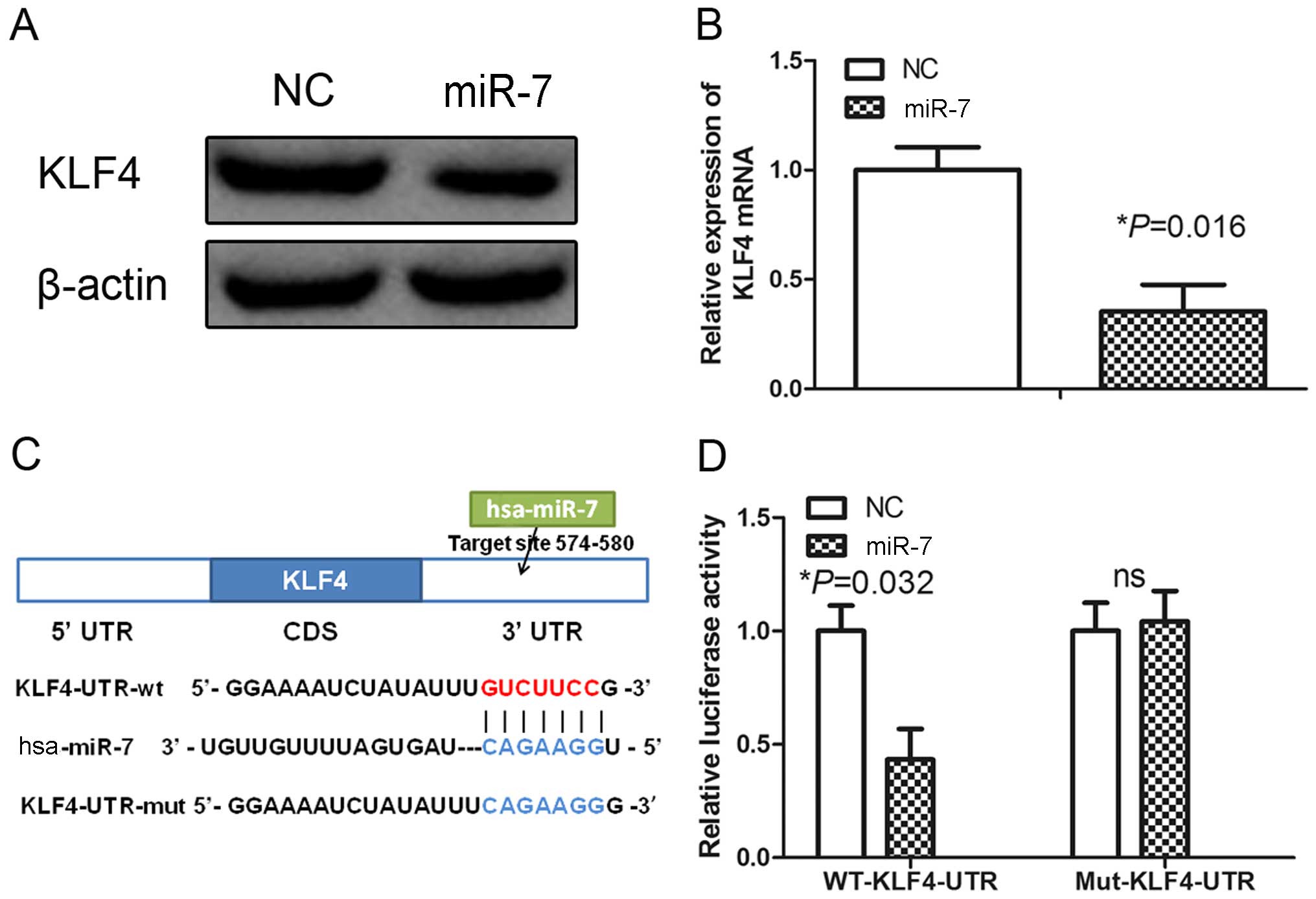
miR-7 targets KLF4 in HUVECs
To explore the mechanism associated with the
downregulation of miR-7 in angiogenesis, we searched for potential
miR-7 target genes using miRNA target prediction tools, including
TargetScan, PicTar and miRanda (22). We found that KLF4 is a potential
miR-7 target. Previous studies have shown that KLF4 is a central
regulator of sprouting angiogenesis via the Notch signaling pathway
(20), and promoted epithelial cell
transformation via targeting KLF4. These data suggested that miR-7
may inhibit angiogenesis by targeting KLF4 in HUVECs. To test this
hypothesis, we analyzed the expression of KLF4 in HUVECs
transfected with control or miR-7 mimics. Western blot analysis
showed that miR-7 mimics decreased expression of KLF4 (Fig. 2A). Furthermore, qRT-PCR analysis
revealed that the levels of KLF4 mRNA were obviously decreased in
the HUVECs transfected with the miR-7 mimics (Fig. 2B). These results suggest that miR-7
may negatively regulate KLF4 expression by disrupting the stability
of KLF4 mRNA.
To confirm that KLF4 is a direct target of miR-7, we
constructed luciferase reporters with the KLF4 3′-UTR containing a
miR-7 binding site (Fig. 2C). The
constructs were transiently transfected into HUVECs along with
miR-7 or NC mimics and the relative luciferase activity was
measured at 48 h. The relative luciferase activity of the construct
with wild-type 3′-UTR was significantly repressed by 60% following
miR-7 mimic transfection (Fig. 2D).
However, site-directed mutagenesis of the miR-7 binding site within
the 3′-UTR of KLF4 completely abolished this suppression (Fig. 2D), indicating that miR-7 directly
binds to this site in HUVECs.
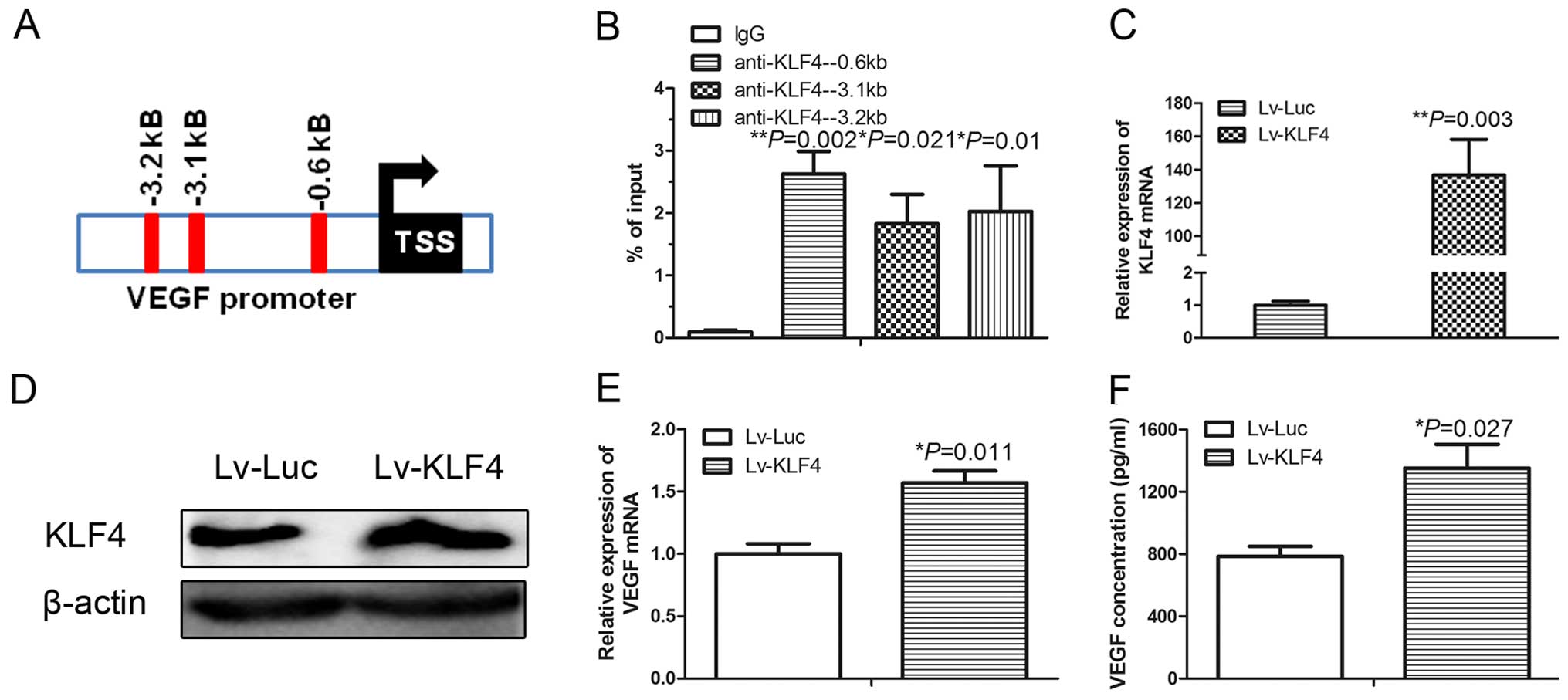
KLF4 activates VEGF transcription in
HUVECs by directly interacting with the VEGF promoter
KLF4 is a transcription factor. The simultaneous
promotion of the expression of KLF4 and VEGF in HUVECs after miR-7
inhibition suggests that KLF4 may promote VEGF transcription.
Indeed, bioinformatic analysis identified the presence of multiple
DNA-binding elements of the KLF4 transcription factor in the VEGF
promoter (Fig. 3A). We then
performed a ChIP and found that the KLF4 antibody specifically
pulled down the VEGF promoter in the HUVECs (Fig. 3B). We next overexpressed KLF4 by
infecting HUVECs with a lentivirus cloned with KLF4 (Lv-KLF4).
Compared with the control lentivirus (Lv-Luc), infection of Lv-KLF4
significantly increased the mRNA (Fig.
3C) and protein level (Fig. 3D)
of KLF4 in the HUVECs. As expected, ectopic overexpression of KLF4
significantly induced the VEGF mRNA levels in the HUVECs treated
with miR-7 ASO (Fig. 3E), and the
concentration of VEGF protein in the cell culture medium from
HUVECs treated with miR-7 ASO (Fig.
3F). These results indicate that KLF4 playsed a direct role in
the transcriptional regulation of VEGF.
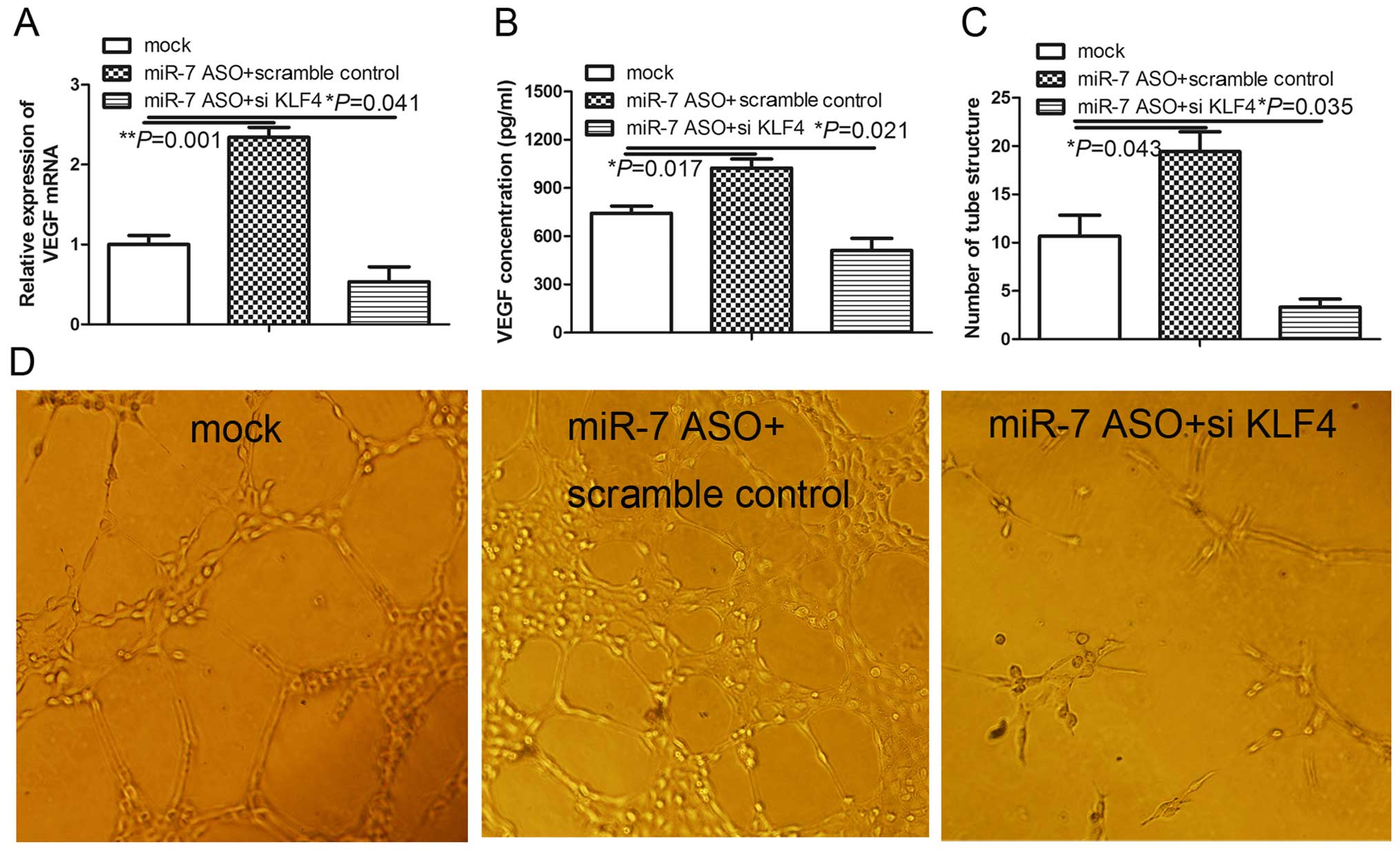
KLF4 knockdown inhibits
miR-7-knockdown-induced angiogenesis in HUVECs
Our observations showed that KLF4 is a functional
target of miR-7 and transcriptional activator of VEGF, which
promotes angiogenesis, suggesting that knockdown of KLF4 may
attenuate angiogenesis induced by miR-7 inhibition. We tested this
hypothesis by transfecting exogenous KLF4 short hairpin RNA into
the miR-7-transfected HUVECs using a previously described transient
transfection system (11). KLF4
silencing significantly abolished the induction of VEGF mRNA in the
HUVECs treated with miR-7 ASO (Fig.
4A) and the concentration of VEGF protein in the cell culture
medium from HUVECs treated with miR-7 ASO (Fig. 4B). Consistent with the key function
of VEGF in angiogenesis, KLF4 knockdown significantly prevented the
induction of angiogenesis by the miR-7 inhibitor in the HUVECs
(Fig. 4C and D). Taken together,
the results indicate that KLF4 knockdown abrogates
miR-7-knockdown-induced angiogenesis, suggesting that KLF4 is a
functional mediator of miR-7 in the regulation of angiogenesis in
HUVECs.
Discussion
In the present study, we demonstrated that miR-7
downregulation promoted the angiogenesis of HUVECs, accompanied by
the induction of VEGF and KLF4 expression. Furthermore, we found
that miR-7 directly targeted KLF4 and KLF4 promoted the gene
transcription of VEGF by directly interacting with the VEGF
promoter in the HUVECs. Consequently, KLF4 silencing inhibited
miR-7-knockdown-induced angiogenesis in HUVECs. These findings
indicate that the miR-7-KLF4-VEGF signaling axis plays an important
role in the regulation of angiogenesis in HUVECs, suggesting that
the miR-7-KLF4 signaling pathway is a potential target for
development of anti-angiogenic therapeutics in vascular diseases as
well as solid tumors.
Growing evidence has shown that aberrant expression
of miRNAs is directly associated with the development and
progression of several types of cancers (23). Deregulation of miR-7 has been found
in cervical (24), breast (25,26),
colorectal (27) and lung cancer
(28). It was recently reported
that miR-7 overexpression inhibited tumor angiogenesis and the
growth of murine xenograft glioblastoma (12). Consistent with the present study, we
found that downregulation of miR-7 led to enhanced tube formation
in HUVECs. Furthermore, our results showed that the miR-7 inhibitor
increased VEGF by targeting KLF4. These data suggest that miR-7 is
an angio-genesis inhibitor and a potential agent for the
development of anti-angiogenic therapeutics.
VEGF is the most potent pro-angiogenesis factor and
plays a crucial role in tumor angiogenesis and metastatic spread of
cancer cells. Thus, targeting VEGF is a very promising strategy for
cancer treatment, with numerous anti-VEGF small molecules in
preclinical and clinical trials (29). However, the precise molecular
mechanism of VEGF regulation in HUVECs remains to be determined.
Zheng et al found that overexpression of KLF4 in ECs
significantly impaired tube formation (30). It was reported that defective
angiogenesis led to reduced tumor growth independent of endothelial
cell proliferation (31). KLF4 is
an upstream regulator of Notch signaling and sustained expression
of KLF4 leads to defective angiogenesis, resulting in attenuation
of tumor growth (20). By contrast,
Hale et al found that sustained expression of KLF4 promoted
sprouting angiogenesis in the HUVECs (20). Consistent with the present study, we
found that KLF4 positively regulated the expression of VEGF in
HUVECs. Therefore, further animal models are needed to clarify the
functions of KLF4 in the regulation of angiogenesis in solid
tumors.
It is well documented that a single miRNA can
post-transcriptionally suppress multiple targets. It is, therefore,
quite possible that miR-7 may also target other
angiogenesis-promoting genes in addition to KLF4 simultaneously, to
promote angiogenesis. For example, a recent study indicated that
miR-7-5p was frequently downregulated in glioblastoma
microvasculature, and miR-7-5p overexpression in HUVECs inhibited
vascular endothelial cell proliferation by targeting RAF1 (32). Babae et al found that miR-7
is a novel anti-angiogenic therapeutic miRNA that was systemically
delivered into both ECs and tumor cells, suggesting a potential
therapeutic role for miR-7 as a novel antitumor agent (12). Similarly, we observed that miR-7
knockdown promoted HUVEC angiogenesis, accompanied by increased
levels of both VEGF and KLF4, explaining the promotion of
angiogenesis by miR-7 inhibitors. It should be noted that VEGF is
not a direct target gene of miR-7. Our findings suggest that KLF4
is a mediator between miR-7 and VEGF in the regulation of
angiogenesis.
In summary, we found that KLF4 is a direct target of
miR-7 and transcription activator of VEGF in HUVECs, revealing an
additional mechanism underlying the inhibition of miR-7 in
angiogenesis. VEGF plays a key role in endothelial cell migration,
proliferation and angiogenesis (33,34).
Our findings suggest that miR-7 downregulation may contribute to
sustained angiogenesis in cancer.
Acknowledgments
The present study was supported by the Science and
Technology Development Program of Shaanxi Province of China
(2013KJXX-91).
References
|
1
|
Nakatsu MN, Sainson RC, Aoto JN, Taylor
KL, Aitkenhead M, Pérez-del-Pulgar S, Carpenter PM and Hughes CC:
Angiogenic sprouting and capillary lumen formation modeled by human
umbilical vein endothelial cells (HUVEC) in fibrin gels: The role
of fibroblasts and Angiopoietin-1. Microvasc Res. 66:102–112. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Conway EM, Collen D and Carmeliet P:
Molecular mechanisms of blood vessel growth. Cardiovasc Res.
49:507–521. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu X, Zhao R, Lin S, Bai X, Zhang L, Yuan
S and Sun L: CXCL16 induces angiogenesis in autocrine signaling
pathway involving hypoxia-inducible factor 1α in human umbilical
vein endothelial cells. Oncol Rep. 35:1557–1565. 2016.
|
|
4
|
Wu Z, Cai X, Huang C, Xu J and Liu A:
miR-497 suppresses angiogenesis in breast carcinoma by targeting
HIF-1α. Oncol Rep. 35:1696–1702. 2016.PubMed/NCBI
|
|
5
|
Yoshikawa N, Shimizu N, Ojima H, Kobayashi
H, Hosono O and Tanaka H: Down-regulation of hypoxia-inducible
factor-1 alpha and vascular endothelial growth factor by HEXIM1
attenuates myocardial angiogenesis in hypoxic mice. Biochem Biophys
Res Commun. 453:600–605. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dong W, Li B, Wang J, Song Y, Zhang Z, Fu
C and Zhang P: Diagnostic and predictive significance of serum
microRNA-7 in esophageal squamous cell carcinoma. Oncol Rep.
35:1449–1456. 2016.
|
|
7
|
Zhang H, Qu Y, Duan J, Deng T, Liu R,
Zhang L, Bai M, Li J, Zhou L, Ning T, et al: Integrated analysis of
the miRNA, gene and pathway regulatory network in gastric cancer.
Oncol Rep. 35:1135–1146. 2016.PubMed/NCBI
|
|
8
|
Chakrabarti M and Ray SK: Antitumor
activities of luteolin and silibinin in glioblastoma cells:
Overexpression of miR-7-1-3p augmented luteolin and silibinin to
inhibit autophagy and induce apoptosis in glioblastoma in vivo.
Apoptosis. 21:312–328. 2016. View Article : Google Scholar
|
|
9
|
Zhao JG, Men WF and Tang J: MicroRNA-7
enhances cytotoxicity induced by gefitinib in non-small cell lung
cancer via inhibiting the EGFR and IGF1R signalling pathways.
Contemp Oncol. 19:201–206. 2015.
|
|
10
|
Zhang N, Li X, Wu CW, Dong Y, Cai M, Mok
MT, Wang H, Chen J, Ng SS, Chen M, et al: microRNA-7 is a novel
inhibitor of YY1 contributing to colorectal tumorigenesis.
Oncogene. 32:5078–5088. 2013. View Article : Google Scholar
|
|
11
|
Zhang X, Hu S, Zhang X, Wang L, Zhang X,
Yan B, Zhao J, Yang A and Zhang R: MicroRNA-7 arrests cell cycle in
G1 phase by directly targeting CCNE1 in human hepatocellular
carcinoma cells. Biochem Biophys Res Commun. 443:1078–1084. 2014.
View Article : Google Scholar
|
|
12
|
Babae N, Bourajjaj M, Liu Y, Van Beijnum
JR, Cerisoli F, Scaria PV, Verheul M, Van Berkel MP, Pieters EH,
Van Haastert RJ, et al: Systemic miRNA-7 delivery inhibits tumor
angiogenesis and growth in murine xenograft glioblastoma.
Oncotarget. 5:6687–6700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Garrett-Sinha LA, Eberspaecher H, Seldin
MF and de Crombrugghe B: A gene for a novel zinc-finger protein
expressed in differentiated epithelial cells and transiently in
certain mesenchymal cells. J Biol Chem. 271:31384–31390. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takahashi K and Yamanaka S: Induction of
pluripotent stem cells from mouse embryonic and adult fibroblast
cultures by defined factors. Cell. 126:663–676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Atkins GB and Jain MK: Role of
Krüppel-like transcription factors in endothelial biology. Circ
Res. 100:1686–1695. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mun GI and Boo YC: A regulatory role of
Kruppel-like factor 4 in endothelial argininosuccinate synthetase 1
expression in response to laminar shear stress. Biochem Biophys Res
Commun. 420:450–455. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ohnesorge N, Viemann D, Schmidt N, Czymai
T, Spiering D, Schmolke M, Ludwig S, Roth J, Goebeler M and Schmidt
M: Erk5 activation elicits a vasoprotective endothelial phenotype
via induction of Kruppel-like factor 4 (KLF4). J Biol Chem.
285:26199–26210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Zhou M, Qin G, Weintraub NL and
Tang Y: MiR-92a regulates viability and angiogenesis of endothelial
cells under oxidative stress. Biochem Biophys Res Commun.
446:952–958. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cui Y, Xiao Z, Chen T, Wei J, Chen L, Liu
L, Chen B, Wang X, Li X and Dai J: The miR-7 identified from
collagen biomaterial-based three-dimensional cultured cells
regulates neural stem cell differentiation. Stem Cells Dev.
23:393–405. 2014. View Article : Google Scholar :
|
|
20
|
Hale AT, Tian H, Anih E, Recio FO III,
Shatat MA, Johnson T, Liao X, Ramirez-Bergeron DL, Proweller A,
Ishikawa M, et al: Endothelial Kruppel-like factor 4 regulates
angiogenesis and the Notch signaling pathway. J Biol Chem.
289:12016–12028. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pan C, Lu B, Chen H and Bishop CE:
Reprogramming human fibroblasts using HIV-1 TAT recombinant
proteins OCT4, SOX2, KLF4 and c-MYC. Mol Biol Rep. 37:2117–2124.
2010. View Article : Google Scholar
|
|
22
|
Thomas M, Lieberman J and Lal A and Lal A:
Desperately seeking microRNA targets. Nat Struct Mol Biol.
17:1169–1174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang GJ, Liu GH, Ye YW, Fu Y and Zhang XF:
The role of microRNA-1274a in the tumorigenesis of gastric cancer:
Accelerating cancer cell proliferation and migration via directly
targeting FOXO4. Biochem Biophys Res Commun. 459:629–635. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hao Z, Yang J, Wang C, Li Y, Zhang Y, Dong
X, Zhou L, Liu J, Zhang Y and Qian J: MicroRNA-7 inhibits
metastasis and invasion through targeting focal adhesion kinase in
cervical cancer. Int J Clin Exp Med. 8:480–487. 2015.PubMed/NCBI
|
|
25
|
Huynh FC and Jones FE: MicroRNA-7 inhibits
multiple oncogenic pathways to suppress HER2Δ16 mediated breast
tumorigenesis and reverse trastuzumab resistance. PLoS One.
9:e1144192014. View Article : Google Scholar
|
|
26
|
Shi Y, Luo X, Li P, Tan J, Wang X, Xiang T
and Ren G: miR-7-5p suppresses cell proliferation and induces
apoptosis of breast cancer cells mainly by targeting REGγ. Cancer
Lett. 358:27–36. 2015. View Article : Google Scholar
|
|
27
|
Suto T, Yokobori T, Yajima R, Morita H,
Fujii T, Yamaguchi S, Altan B, Tsutsumi S, Asao T and Kuwano H:
MicroRNA-7 expression in colorectal cancer is associated with poor
prognosis and regulates cetuximab sensitivity via EGFR regulation.
Carcinogenesis. 36:338–345. 2015. View Article : Google Scholar
|
|
28
|
Li J, Zheng Y, Sun G and Xiong S:
Restoration of miR-7 expression suppresses the growth of Lewis lung
cancer cells by modulating epidermal growth factor receptor
signaling. Oncol Rep. 32:2511–2516. 2014.PubMed/NCBI
|
|
29
|
D'Alessandris QG, Martini M, Cenci T, Capo
G, Ricci-Vitiani L, Larocca LM and Pallini R: VEGF isoforms as
outcome biomarker for anti-angiogenic therapy in recurrent
glioblastoma. Neurology. 84:1906–1908. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zheng X, Li A, Zhao L, Zhou T, Shen Q, Cui
Q and Qin X: Key role of microRNA-15a in the KLF4 suppressions of
proliferation and angiogenesis in endothelial and vascular smooth
muscle cells. Biochem Biophys Res Commun. 437:625–631. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ray A, Alalem M and Ray BK: Loss of
epigenetic Kruppel-like factor 4 histone deacetylase
(KLF-4-HDAC)-mediated transcriptional suppression is crucial in
increasing vascular endothelial growth factor (VEGF) expression in
breast cancer. J Biol Chem. 288:27232–27242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Z, Liu Y, Li L, Xu Z, Bi B, Wang Y and
Li JY: MiR-7-5p is frequently downregulated in glioblastoma
microvasculature and inhibits vascular endothelial cell
proliferation by targeting RAF1. Tumour Biol. 35:10177–10184. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koch AE, Harlow LA, Haines GK, Amento EP,
Unemori EN, Wong WL, Pope RM and Ferrara N: Vascular endothelial
growth factor. A cytokine modulating endothelial function in
rheumatoid arthritis. J Immunol. 152:4149–4156. 1994.PubMed/NCBI
|
|
34
|
Melter M, Reinders ME, Sho M, Pal S,
Geehan C, Denton MD, Mukhopadhyay D and Briscoe DM: Ligation of
CD40 induces the expression of vascular endothelial growth factor
by endothelial cells and monocytes and promotes angiogenesis in
vivo. Blood. 96:3801–3808. 2000.PubMed/NCBI
|