Introduction
Malignant gliomas are the most common type of
primary brain tumors (1), of which
patients have a notably high likelihood of relapse after treatment
(2–4). Glioblastomas are understood to evolve
from brain glioma stem cells (BGSCs). BGSCs have the capabilities
of self-renewal, and may render themselves profitable to the
survival of tumors by conferring resistance to therapeutic
treatments (5,6). It was initially proposed that acute
myelogenous leukemia may evolve from a subset of precursor stem
cells, and considerable efforts have been devoted to studying this
sub-population of cells in numerous cancers since (7). Stem cells, for example, exhibit many
of the hallmarks inherent to malignant tumors, including the
abilities to proliferate, invade and metastasize (5).
Stem cells expressing the CD133 membrane protein are
markedly pronounced within cells of malignant gliomas (7,8).
Furthermore, the encephalic inoculation of CD133+ stem
cells into NOD-SCID mice results in tumor growth (9). Thus, CD133+ stem cells are
likely to be important for tumor recurrence. Paired box 3 protein
(Pax3) is a member of the paired box (Pax) family of transcription
factors that is normally expressed during embryonic development,
but has recently been implicated in tumorigenesis (8). The expression of Pax3, for example,
was found to be positively correlated with melanoma progression
(10–12). In addition, GFAP is a major
intermediate filament protein, where it likely plays a critical
role in mature astrocytes owing to its high abundance and strong
conservation among vertebrates. Pax3 has previously been reported
to negatively regulate the expression of GFAP during the
differentiation of astrocytes from neural stem cells (NSCs)
(13,14). In addition, Pax3 overexpression can
be detected during the process of the astrocyte precursor cell
proliferation and can maintain the high malignancy in gliomas by
negatively regulating GFAP expression in glioma cells (15). Thus, we speculated that GFAP is a
Pax3-responsive gene in BGSCs. In these studies, we attempted to
examine the expression of Pax3 and to determine whether Pax3 could
bind to the element of the GFAP promoter in BGSCs.
Materials and methods
Cell lines and reagents
Human BGSCs were kindly gifted by Soochow
University, China. Human malignant glioma cell line (U87MG) and
normal human astrocytes (1800) were obtained from the Cell Library
of the Chinese Academy of Sciences (Shanghai, China). BGSCs were
cultured at 37°C in 5% CO2 in a 1:1 mix of Dulbecco's
modified Eagle's medium and Ham's F-12 medium (DMEM/F-12) (Gibco,
Grand Island, NY, USA) containing 10 ml B27, 10 µg EGF, 10
µg FGF, 5 ml L-glutamine, 5 ml MEM-vitamin solution, 5 ml
sodium pyruvate, 5 ml MEM non-essential amino acids (Gibco), and 5
ml penicillin-streptomycin solution (HyClone, Logan City, UT, USA).
U87MG cells were cultured at 37°C in 5% CO2 in DMEM
supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine
and 100 U/ml penicillin-streptomycin (Gibco). The normal astroctyes
(1800) were cultured at 37°C and 5% CO2 in modified
RPMI-1640 medium (HyClone) supplemented with 10% FBS, 2 mM
L-glutamine and 100 U/ml penicillin-streptomycin (Gibco). Culture
medium was altered every 3–4 days. U87MG and 1800 cells were split
using 0.25% trypsin. BGSCs were split using StemPro Accutase. The
present study obtained ethical approval from the Affiliated
Hospital of Nantong University, China [approval ID: (2013)040].
RT-PCR
Total RNA was extracted from cells with TRIzol
(Invitrogen, Carlsbad, CA, USA), from which cDNA was reverse
transcribed using the Omniscript RT kit (Qiagen) aaccording to the
manufacturer's instructions. The sequences for primers used were as
follows: H-Pax3-F, 5′-AAGCCCA AGCAGGTGACAA-3′ and H-Pax3-R,
5′-ATGGAACTCACTGACGGCAC-3′; H-GFAP-F, 5′-AGATCCGCACGCAGTA TGA-3′
and H-GFAP-R, 5′-AGTCGTTGGCTTCGTGCTT-3′; H-β-actin-F,
5′-CATGAAGTGTGACGTGGACATC-3′ and H-β-actin-R,
5′-GGACTCGTCATACTCCTGATTG-3′. The thermal cycles were: 95°C for 30
sec, 68°C for 30 sec, and 72°C for 30 sec for 35 cycles for
β-actin, and 95°C for 30 sec, 58°C for 30 sec, and 72°C for 30 sec
for 35 cycles for Pax3.
Western blotting
Expression levels of β-actin, Pax3 and GFAP proteins
were determined via western blotting with specific antibodies as
previously described (16). The
following antibodies were used: primary antibodies against β-actin
(1:2,000 dilution), Pax3 (1:500) and GFAP (1:1,000); all of which
were obtained from Abcam (Cambridge, MA, USA).
Immunofluorescence microscopy
Dispersed BGSCs were smeared onto a Poly-L-lysine
glass slide. Cells were washed with phosphate-buffered saline
(PBS), fixed in 4% paraformaldehyde for 30 min, permeabilized in
0.1% Triton X-100 for 30 min, and blocked against non-specific
binding in 5% BSA (Amresco, Solon, OH, USA) for 45 min at room
temperature. Subsequently, cells were incubated with rabbit
anti-Pax3 polyclonal (1:100), GFAP monoclonal (1:100) (both from
Abcam), anti-human nestin (1:100; Alexa Fluor 488l eBioscience, San
Diego, CA, USA), and anti-human CD133-APC antibodies (1:100;
Miltenyi Biotec) overnight at 4°C. The slides were then washes 3
times with PBS, and incubated with Cy3 or FITC
fluorescently-labeled secondary antibodies for 2 h at room
temperature. DNA was stained by incubating the slides in
4′,6-diamidino-2-phenylindole (DAPI) (0.2 mg/ml) for 2 min
immediately following incubation with secondary antibodies. Slides
were stored at 4°C in the dark and visualized with a Leica
fluorescence microscope (Germany). All assays were performed 3
times in duplicate.
siRNA transfection
Pax3 was knocked down as previously described
(15), using siRNA sequences
purchased from Biomics (Jiangsu, China). The siRNA sequences were:
Pax3 sense, 5′-CGCAUCCUGAGAAGUAAAUdTdT-3′ and Pax3 antisense,
5′-AUUUACUUCUCAGGAUGCGdTdT-3′; negative control (NC) sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and NC antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. siRNAs were transfected into BGSCs in
6-well plates (1 mg/ml) using the MicroPoly-Transfecter Cell
Reagent (Invitrogen) completed according to the manufacturer's
instructions.
Transient transfections and
differentiation
Plasmid vectors for overexpressing Pax3 were
constructed by GeneChem (Shanghai, China). Transient transfections
were carried out using the MicroPoly-Transfecter Cell Reagent
according to the manufacturer's instructions. To induce
differentiation, the transfected BGSCs were seeded onto coverslips
coated with poly-L-lysine within 24-well plates, and cultured in
DMEM/F-12 supplemented with 1% FBS for 3 days. Next, the
neurospheres were fixed and processed for immunofluorescence
microscopy as previously described. A subset of cells were omitted
from immunofluorescence microscopy, and were incubated at 37°C for
24 h after transfection.
CCK-8 cell proliferation assay
Cells at 12 h post-transfection were seeded into
96-well plates at a density of 4,000 cells/well. Next, cell
viability was determined using the Cell Counting Kit-8 (CCK-8;
Dojindo, Shanghai, China). Briefly, 100 µl DMEM/F-12 and 10
µl CCK-8 reagent were added to each pre-cultured film,
whereupon the plates were incubated for 1.5 h at 37°C. The
absorbance was determined at a wavelength of 450 nm using a
Multiskan MK33 microplate reader (Thermo Electron Corporation,
Shanghai, China).
Cell invasion assay
A cell invasion assay was performed using 24-well
tissue culture plates (BD Biosciences, Bedford, MA, USA) consisting
of an 8-µm polyethylene terephthalate membrane coated with a
Matrigel basement membrane matrix (100 µg/cm2).
In brief, the Matrigel (R&D Systems, Minneapolis, MN, USA) was
rehydrated overnight at 4°C. The Transwell membranes were
pre-coated with 24 mg/ml Matrigel. Two days after transfection, the
cells (5×104) were seeded into the upper chamber with
DMEM/F-12, and the lower chamber was filled with DMEM containing 10
ml B27, 10 µg EGF, 10 µg FGF, 5 ml L-glutamine, 5 ml
MEM-vitamin-solution, 5 ml sodium pyruvate, 5 ml MEM non-essential
amino acids, and 5 ml penicillin-streptomycin solution as a
chemoattractant. The invasion assay was carried out in a 5%
CO2 humidified chamber at 37°C for 48 h, and cells on
the upper surface of the filters were removed by wiping the upper
surface of the membrane with a cotton swab. The filter membrane was
fixed in 4% paraformaldehyde, and stained with Coomassie blue. The
degree of invasion was quantified by counting the cells that
migrated to the lower side of the filter in at least six random
fields at a magnification of ×100 using a fluorescence microscope.
Experiments were repeated 3 times in triplicate.
Apoptosis detection
Cells were trypsinized, counted, washed twice with
ice-cold PBS, and resuspended in 1X binding buffer (pH 7.4)
containing 10 mM HEPES/NaOH, 140 mM NaCl, and 2.5 mM
CaCl2. Next, 100 µl cell suspension was stained
with 5 µl Annexin V-FITC fluorescent label (R&D Systems
Europe Ltd., Abingdon, UK) and 10 µl propidium iodide (PI)
(Sigma) for 15 min at room temperature in the dark. Finally, 400
µl of binding buffer was added to each sample, which was
immediately held on ice prior until analysis on a FACSCalibur flow
cytometer (BD Pharmingen, San Diego, CA, USA), for which
105 cells/sample were analyzed.
Monoclonal formation
The original neural cells were digested with StemPro
Accutase into single cell suspension. Next, the digests were
diluted to 40 cells/ml with serum-free medium containing B27, bFGF
and EGF, of which 50 µl of this cell suspension was
inoculated/microwell of a 96-well plate. Cells were centrifuged at
1,000 × g, to which 3 parts supernatant was combined with 1 part
fresh serum-free culture medium, and bFGF and EGF were added for a
final concentration of 20 ng/ml. Next, 100 µl serum-free
culture mixture was added to each microhole, whereupon cells were
cultivated at 37°C and 5% CO2. Finally, the division and
cloning of the single cells were observed with microscopy.
Chromatin immunoprecipitation
Chromatin was immuneprecipitated with a ChIP assay
kit (Upstate Biotechnology, Lake Placid, NY, USA) completed as per
the manufacturer's instructions. Briefly, BGSCs were lysed, where
upon the chromatin was immunoprecipitated with anti-Pax3 polyclonal
antibodies (sc-376215; Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA).
Luciferase reporter assay
The length promoter plasmid pGFAP1600 of the human
GFAP gene was cloned into a pGL3-Basic luciferase reporter vector
(Promega, Madison, WI, USA) and amplified via PCR. The
oligonucleotide PCR primer sequences used to detect the fragment of
the pGFAP were as follows: GFAP F (KpnI),
5′-CCCGGTACCAAGCAGAC CTGGCAGCATTG-3′ and GFAP R (HindIII),
5′-CGGAAGC TTGCCCGGGTGCCCCTGGCAAC-3′. Mutant-promoter constructs
were created by deleting the nucleotides corresponding to the P1
and P2 probes (P1 F5, 5′-CATGCCCATGA CTCACCTTGGCACAG-3′ and R5,
5′-GTGAGTCATGGGCATGAAGAGGAGGC; P2 F7,
5′-GGGATTACGCCACCCCACTCAGCCCT-3′ and R7,
5′-GGGGTGGCGTAATCCCAGCACTTTGG-3′) of pGFAP1600, either individually
or in combination, using a Pfu Turbo DNA polymerase from a
QuikChange kit (Stratagene, La Jolla, CA, USA). The mutant-promoter
constructs were termed pGFAP1600-DelP1 (deletion of 5 nucleotides
within P1) or pGFAP1600-DelP2x (deletion of 8 nucleotides within
P2). The human Pax3 was achieved by cloning the cDNA fragment into
a pCI vector in HEK 293T cells. All constructs were confirmed by
sequence validation. Normal or mutated GFAP promoter reporter
plasmids were transfected together with the PRL (Promega) vector
into HEK 293T cells using Lipofectamine 2000. HEK293T cells were
used for luciferase assay since these cells are more easily
transfected in comparison with BGSCs. Twenty-four hours after
transfection, the cells were lysed and assayed using the
Dual-Luciferase Assay kit (Promega). Promoter activity was measured
by comparing luciferase levels.
Electrophoretic mobility shift assay
Putative interactions between Pax3 and GFAP mRNA
transcripts were monitored via the electrophoretic mobility shift
assay. Nuclear components of HEK 293T cells transfected with
overexpression-Pax3 were extracted according to NE-PER Nuclear and
Cytoplasmic Extraction Reagents (Pierce, Rockford, IL, USA) and
electrophoretic mobility shift assays (EMSA) were exactly performed
as previously described (17).
First, we designed fluorescently-labeled normal and mutated probes.
The sequence of the normal and mutated oligo nucleotides used in
this experiment were: P1 probe Fw, 5′-ATGCCCAGTGAATGACTCAC-3′ and
Rw, 5′-GTGAGTCATTCACTGGGCAT-3′; PIM probe Fm,
5′-ATGCCCACTGTATGACTCAC-3′ and Rm, 5′-GTGAGTCATACAGTGGGCAT-3′. For
supershift experiments, nuclear extracts were pre-incubated with
anti-Pax3 polyclonal antibodies (Abcam) before adding the labeled
probes.
Statistical analysis
All statistical analyses, including the t-test was
carried out using GraphPad Prism software (version 6; GraphPad
Software, La Jolla, CA, USA).
Results
Identification of BGSCs and Pax3 and GFAP
mRNA expression
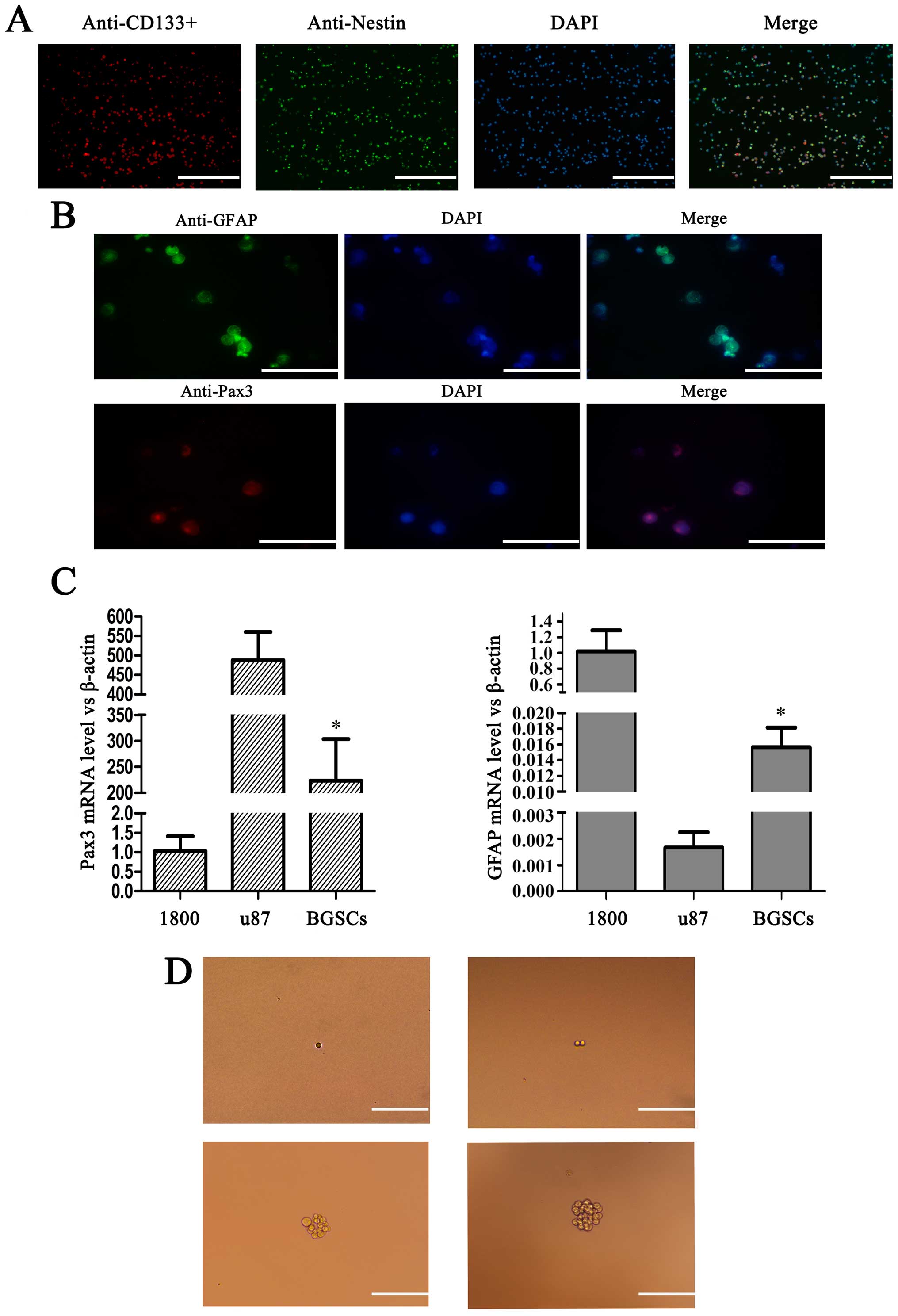
We performed immunofluorescence staining with
CD133+ and nestin antibodies to identify BGSCs. Through
immunofluorescence, the nuclei of the supposed BGSCs were stained
blue, the cytoplasm red, and the cell membrane stained green,
therefore confirming the expression of CD133+ and nestin
BGSC markers (Fig. 1A). We also
determined whether Pax3 enhanced GFAP expression in the BGSCs using
immunohistochemical staining (Fig.
1B), and observed that GFAP and Pax3 proteins co-localized in
the cytoplasm of the BGSCs. Furthermore, we measured the expression
of Pax3 and GFAP in the U87MG human glioma cell, BGSC and 1800
normal human astrocyte lines by RT-PCR. Pax3 was highly expressed
in the BGSCs compared with that noted in the normal human
astrocytes (Fig. 1C). In addition,
we found that a single BGSC could form a colony of 50 BGSCs by
proliferation, suggesting the observed colony formation resulted
from the ability of BGSCs to self-renew and proliferate, rather
than the recruitment of individual BGSCs (Fig. 1D).
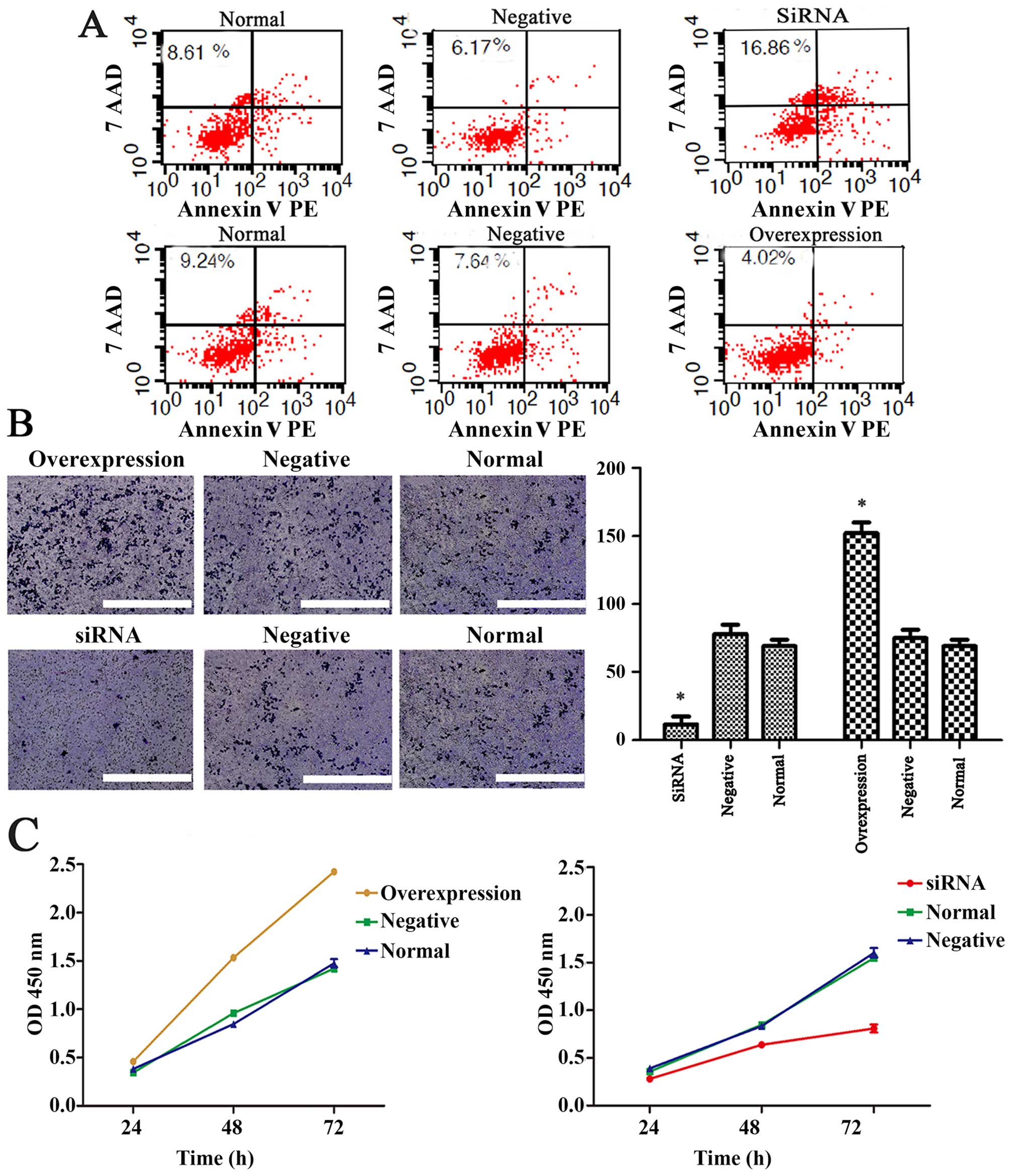
Pax3 decreases the apoptosis, enhances
the invasion and promotes the proliferation of BGSCs
The proportion of BGSCs undergoing apoptosis was
determined using an Annexin V/PI apoptosis detection kit coupled to
flow cytometry. We found that a significantly greater proportion of
siPax3-transfected BGSCs underwent apoptosis relative to the
control BGSCs (P<0.01), while the apoptotic index of BGSCs
transfected to overexpress Pax3 was significantly lower than that
of the untransfected cells (P<0.01; Fig. 2A). Subsequently, we performed a cell
invasion assay to determine the influence of Pax3 on the invasion
of BGSCs. The number of invading BGSCs transfected with a
Pax3-knockdown siRNA was significantly lower (14±5) than that of
the untransfected cells (78±12; P<0.01). Moreover, the number of
BGSCs transfected with a Pax3-overexpression vector (152±6) was
markedly greater than that of the normal control cells (P<0.01).
The above results suggest that Pax3 enhances BGSC invasion in vivo
(Fig. 2B).
We also determined the influence of Pax3 expression
status on the proliferation of BGSCs using the CCK-8 assay. We
found that the proliferation of siPax3-transfected BGSCs was
significantly inhibited (P<0.05), but there was a significant
increase in proliferation in the overexpression Pax3
plasmid-transfected BGSCs (P<0.05; Fig. 2C). These data suggest that Pax3
promotes the proliferation of BGSCs.
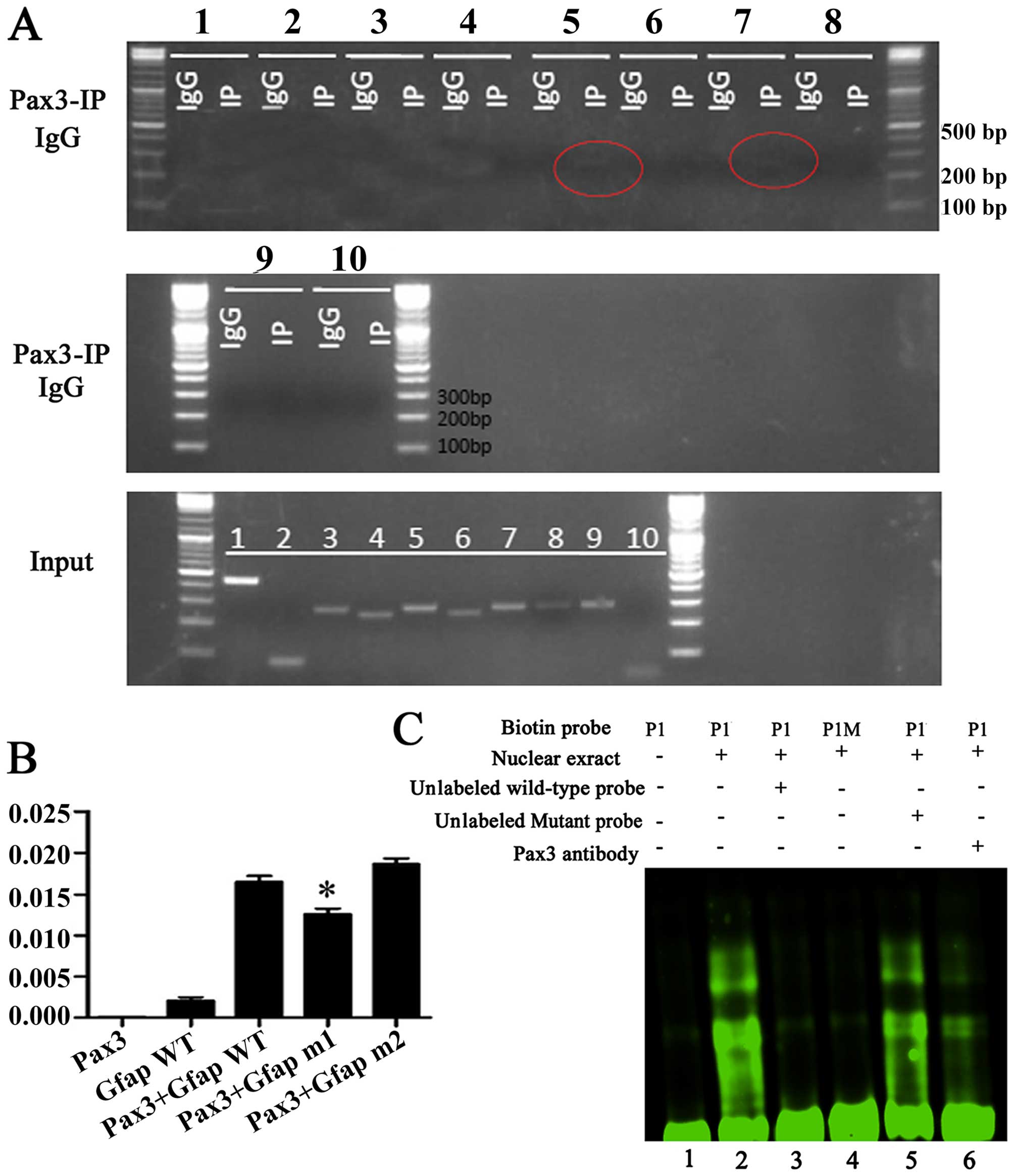
Pax3 binds the GFAP promoter
Pax3 is a transcription factor containing a paired
domain that recognizes consensus sequences harboring a GTTCC motif
and a paired-type homeodomain that binds with a consensus sequence
harboring an ATTA motif (13). We
used chromatin immunoprecipitation to determine whether Pax3 can
bind to the promoter region of GFAP in the context of native
chromatin. First, we analyzed the gene promoter sequences within
2.0 kb upstream of the transcriptional start site of genes using a
pair of primers of 250–300 bp (GFAP F1, 5′-CCAGGTCCCCAGTTCATAGCA-3′
and GFAP R1, 5′-TCCTTCCACATCAGCCTCCC-3′; GFAP F2,
5′-TGTCCAAATGCAGAGCATACCC-3′ and GFAP R2,
5′-GGCGCAACCACGACTCACTG-3′; GFAP F3, 5′-AGGCTGAGGTGAGGGGATCA-3′ and
GFAP R3, 5′-AATGCTGCCAGGTCTGCTTG-3′; GFAP F4,
5′-CAAGCAGACCTGGCAGCATT-3′ and GFAP R4,
5′-CTGAATAGAGCCTTGTTCTCCACC-3′; GFAP F5, 5′-CACCGGCGGTGGAGAACAAG-3′
and GFAP R5, 5′-TCTGGGGATGGATGGGTTTG-3′; GFAP F6,
5′-TCCCCAGAGGTTCTTCCCATC-3′ and GFAP R6,
5′-GTGGCAGTGGAGGTCCTGATAG-3′; GFAP F7,
5′-CAGGACCTCCACTGCCACATAGA-3′ and GFAP R7,
5′-TTTCATAACCCAGGCATTATCTCACT-3′; GFAP F8,
5′-AGTTGGAAAGCAGGTCAGAGGTCA-3′ and GFAP R8,
5′-GGAAGGTGGGTCAAGAAAGGGTT-3′; GFAP F9,
5′-ACCCTTTCTTGACCCACCTTCC-3′ and GFAP R9,
5′-TCTGGCTCTGCTCGCTCCTG-3′; GFAP F10, 5′-CCTCAGTGGGGTGAGGGGAGC-3′
and GFAP R10, 5′-GGGGCATTCGAGCCAGGGAG-3′). We also used an
anti-Pax3 antibody specific for targeting an intron sequence of
GFAP. The binding of Pax3 to the P1 and P2 regions was
subsequently demonstrated by PCR performed with primers specific to
BGSCs (Fig. 3A).
Subsequently, we referred to the study by Cao et
al that described two Pax3 protein binding sites, designated P1
(5′-ATGCCAGTGAATGACTCAC-3′, spanning from −949 to −979 bp on the
positive strand) and P2 (5′-GGGATTACAAGCATGAGCCACC-3′, spanning
from −2,172 to −2,183 bp on the positive strand) (18). According to these results, we
constructed two recombinant normal and mutated promoters of
GFAP, and performed luciferase reporter gene assays to
determine mRNA transcript expression. The normal pGFAP1600
luciferase promoter construct showed low transcription activity,
while inducing overexpression of Pax3 resulted in a notable
increase in transcriptional activity. Furthermore, the absence of
P1 in the promoter region resulted in a notable decrease in
transcription activity, whereas the absence of the P2 promoter
region had no obvious suppression on transcription (Fig. 3B). Together, these results indicated
that: i) Pax3 can regulate GFAP; and ii) that P1 is the
binding region for transcription factors. Next, the EMSA revealed
that Pax3 binds to the P5 promoter region of GFAP. The
double-stranded oligonucleotides corresponding to the predicted
cis-element of the GFAP promoter region formed
sequence-specific DNA/protein complexes with the nuclear extracts
of Pax3-expressing HEK293T cells, while the mutated P1 probes
failed to generate such a DNA/protein complex during EMSA (Fig. 3C). Finally, we performed
EMSA-supershift analysis using a fluorescently-labeled anti-Pax3
antibody to stain the nuclear extract of HEK 293T cells with
overexpression of Pax3. The presence of the anti-Pax3 antibody
generated the supershift band.
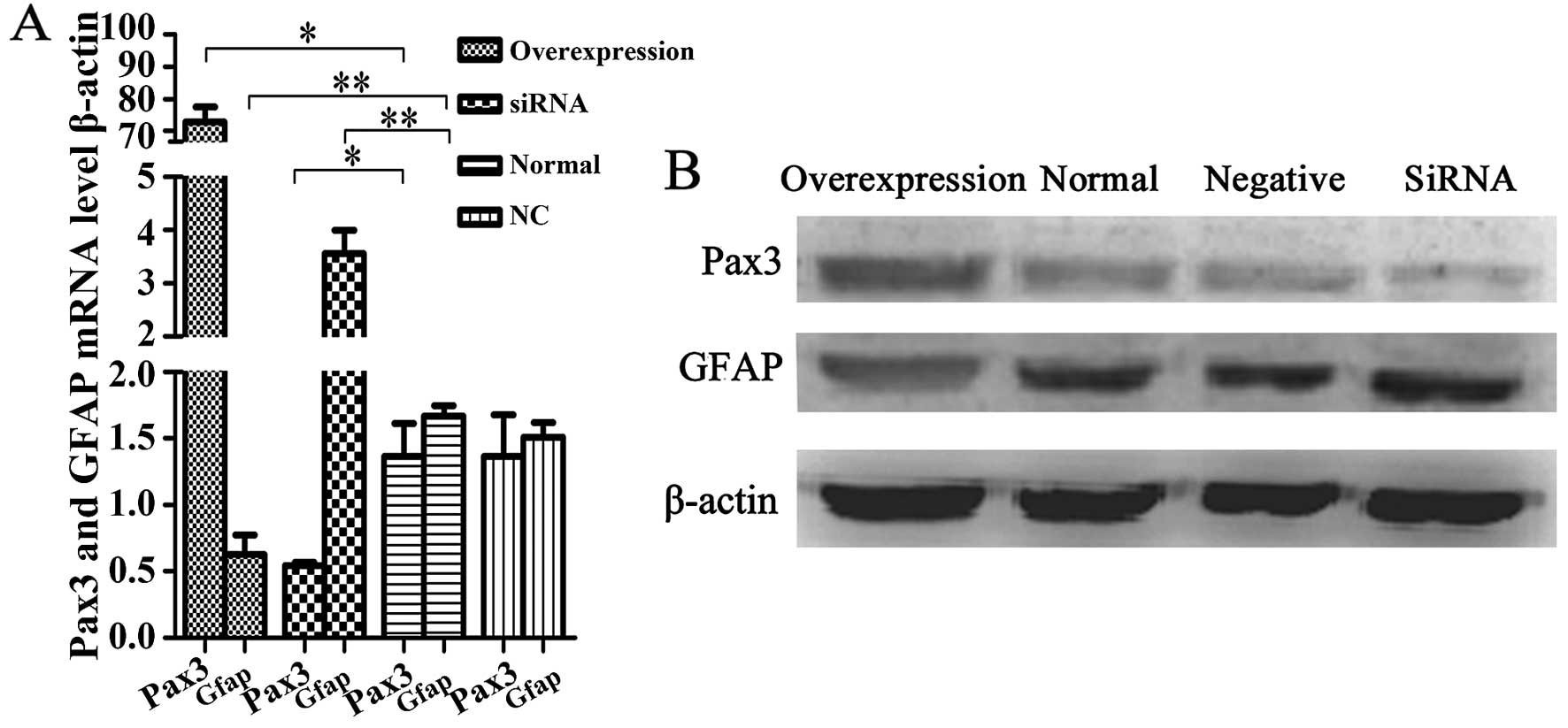
Pax3 inhibits GFAP expression in BGSCs
and has an effect on BGSC differentiation
To ascertain that Pax3 is a negative regulator of
GFAP transcription, we suppressed Pax3 expression with an
siRNA that specifically targets Pax3 mRNA, while creating a Pax3
enhanced-expression model via transfection with an
overexpression-Pax3 plasmid. Then, we performed RT-PCR and western
blotting to determine the expression of GFAP at the mRNA and
protein expression levels, respectively, with the Pax3 knockdown
and overexpression BGSC models. We found that the expression of
GFAP mRNA and protein was significantly greater after Pax3
knockdown compared to that noted in the untransfected cells
(P<0.05). In addition, GFAP mRNA and protein were both decreased
in the Pax3-overexpression model (P<0.05). The results implicate
that Pax3 negatively regulates the expression of GFAP at both the
mRNA and protein levels (Fig. 4).
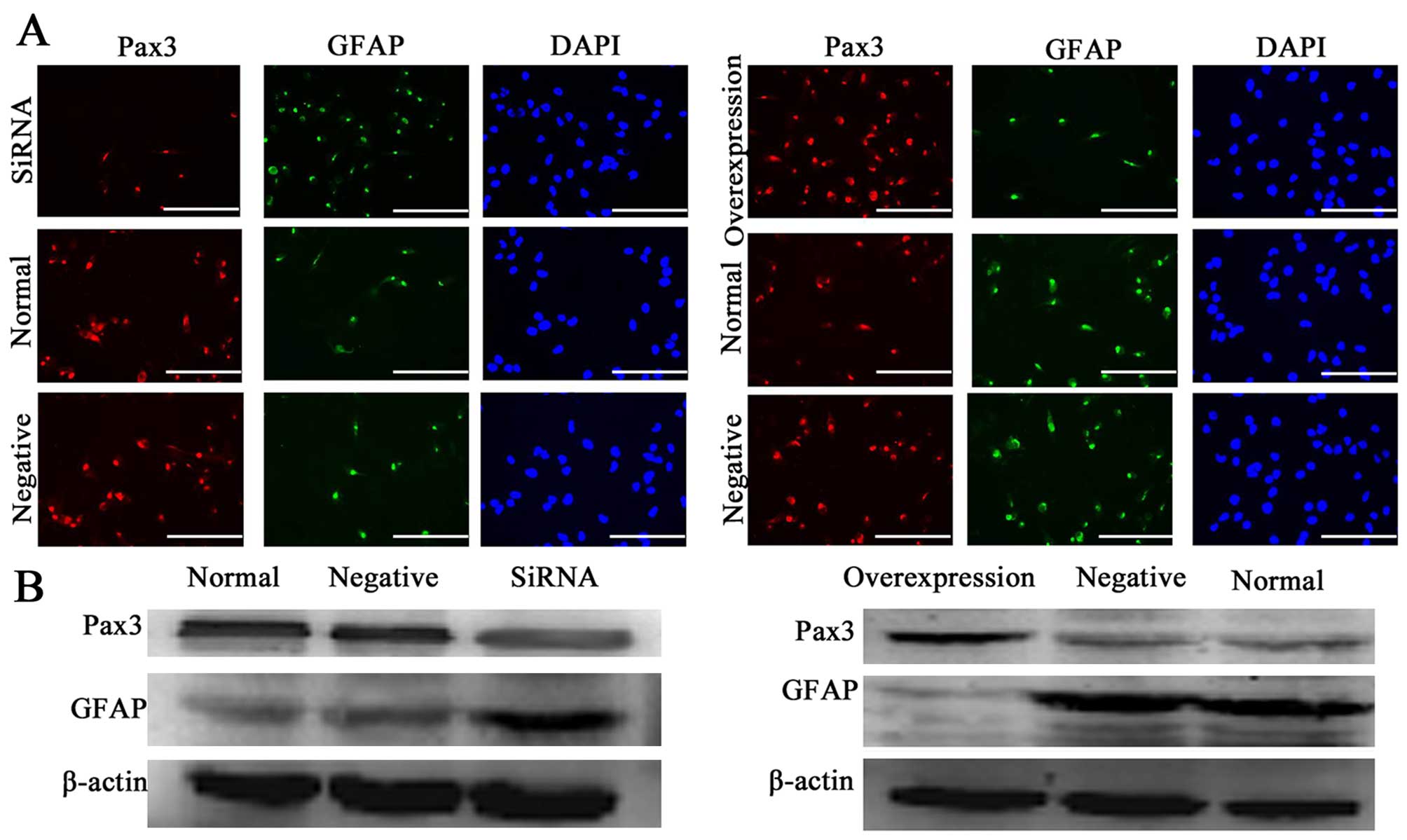
To determine whether overexpression and suppression of Pax3
modulated the differentiation of BGSCs, we transfected siRNA and an
overexpression-Pax3 plasmid into the BGSCs. We performed
immunofluorescence with anti-GFAP and showed that the proportion of
GFAP-positive cells was 20, 24 and 67% in the negative controls,
untransfected cells and BGSCs transfected with siPax3,
respectively. We also observed that, in association with Pax3
silencing via siRNA, the expression of GFAP significantly increased
compared to the untransfected cells (P<0.05; Fig. 5A). Furthermore, western blotting
also showed that the expression of GFAP protein in the BGSCs was
significantly increased after the knockdown of Pax3 compared to the
untransfected cells (P<0.05; Fig.
5B). These results demonstrated that Pax3 negatively regulates
the expression of GFAP and affects the differentiation in
BGSCs.
Discussion
Brain glioma stem cells (BGSCs) were initially
isolated from leukemic cells (19).
Following Reya et al recognition of the similarities in the
biological hallmarks of stem and tumor cells, it was proposed that
cancers may evolve from a subset of precursor stem cells (20). Since then, stem cells have been
isolated from the tumor microenvironment of numerous types of
cancers, including those of the nasopharynx, breast and prostate
cancer (21). Although Ignatova
et al previously reported on the isolation of BGSCs from
cortical glial tumors (22), it is
apparent that we currently know very little in regards to the
contribution of BGSCs to tumorigenesis. Accordingly, the present
study endeavored to investigate the putative roles of BGSCs, with
particular prominence on their contribution to the hallmarks of
cancers.
CD133+ cells exist in various tissues,
including umbilical cord blood, fetal brain, fetal liver and
placental trophoblasts. Moreover, immunohistochemical staining has
shown that CD133 is distributed throughout stem cells; on the
membrane and within the cytoplasm. Recent studies have shown that
CD133 is expressed in neural stem cells (NSCs) (23,24).
Furthermore, nestin, a protein marker for NSCs, is also expressed
in follicle stem cells. In the present study, by
immunohistochemical staining, we identified CD133 and nestin as
putative markers of BGSCs.
Prior research has implicated that Pax3 binds to the
promoter region of GFAP, thereby negatively regulating
GFAP expression and subsequently arresting the
differentiation of astrocytes from NSCs (13). Furthermore, the overexpression of
Pax3 has been observed within glioma tissues, in which it was
reported to regulate GFAP expression in glioma cells
(15). However, no studies have
focused on the transcriptional relationship between Pax3 and
GFAP in BGSCs. Herein, we found that Pax3 was expressed at a
higher level in BGSCs than in U87MG malignant glioma cell and 1800
normal astrocyte lines. Then, we identified a binding element
within the promoter region of GFAP to which Pax3 could
associate, and subsequently demonstrated that Pax3 inhibits the
transcription of GFAP in BGSCs.
To describe the influence of Pax3 on the behavior of
BGSCs, Pax3 was silenced via the transfection of small interfering
RNA (siRNA) into BGSCs. The subsequent cell proliferation, invasive
and apoptosis assays showed that this siRNA-induced downregulation
of Pax3 inhibited proliferation, induced apoptosis and decreased
the invasiveness of transfected BGSCs. In contrast, upregulation of
Pax3 via transfection of BGSCs with an overexpression vector
induced increased cell proliferation and invasiveness, while
suppressing apoptosis. Together, these results suggests that Pax3
plays a vital role in the growth and evolution of BGSCs.
To further determine the effects of Pax3 on BGSC
differentiation and GFAP expression, siPax3-transfected BGSCs were
cultivated in culture medium containing serum, to which the ability
of BGSCs to differentiate was decreased. To the contrary,
differentiation was increased in the overpression Pax3
plasmid-transfected BGSCs. Our results indicate that Pax3 acts as a
transcriptional repressor during serum-induced differentiation of
BGSCs. Thereby, it is reasonable to speculate that Pax3 suppresses
the expression of GFAP during oncogenesis.
In conclusion, we firstly demonstrated that Pax3
binds to the promoter region of GFAP, and consequently
negatively regulates GFAP expression in BGSCs. Pax3 can be
considered a regulator of BGSC differentiation and may determine
the degree of the malignancy of gliomas. Pax3 also plays a crucial
role in the regulation of the growth and invasion of BGSCs. The
results of the present study imply that Pax3 is a putative target
for novel therapies endeavoring to treat gliomas.
Acknowledgments
Brain glioma stem cells were kindly gifted by
Soochow University, China. The present study was supported by the
Natural Science Foundation of Jiangsu Province (BK20130386), the
National Natural Science Foundation of China (81402447), the
Chinese Projects for Postdoctoral Science Funds (no. 2015M571792),
the Jiangsu Planned Projects for Postdoctoral Research Funds (no.
1402200C), the Six Major Human Resources Project of Jiangsu
Province (2014-WSW-028), and the Technology Project of Nantong
(HS2014053).
References
|
1
|
Fang JS, Deng YW, Li MC, Chen FH, Wang YJ,
Lu M, Fang F, Wu J, Yang ZY, Zhou XY, et al: Isolation and
identification of brain tumor stem cells from human brain
neuroepithelial tumors. Zhonghua Yi Xue Za Zhi. 87:298–303. 2007.In
Chinese. PubMed/NCBI
|
|
2
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al European organisation for Research and Treatment of Cancer
Brain Tumor and Radiotherapy Groups; National Cancer Institute of
Canada Clinical Trials Group: Radiotherapy plus concomitant and
adjuvant temozolomide for glioblastoma. N Engl J Med. 352:987–996.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Komotar RJ, Otten ML, Moise G and Connolly
ES Jr: Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma-a critical review. Clin Med Oncol. 2:421–422.
2008.PubMed/NCBI
|
|
4
|
Enting RH, van der Graaf WT, Kros JM,
Heesters M, Metzemaekers J and den Dunnen W: Radiotherapy plus
concomitant and adjuvant temozolomide for leptomeningeal pilomyxoid
astrocytoma: A case study. J Neurooncol. 80:107–108. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Frosina G: Stem cell-mediated delivery of
therapies in the treatment of glioma. Mini Rev Med Chem.
11:591–598. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dietrich J, Diamond EL and Kesari S:
Glioma stem cell signaling: Therapeutic opportunities and
challenges. Expert Rev Anticancer Ther. 10:709–722. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galli R, Binda E, Orfanelli U, Cipelletti
B, Gritti A, De Vitis S, Fiocco R, Foroni C, Dimeco F and Vescovi
A: Isolation and characterization of tumorigenic, stem-like neural
precursors from human glioblastoma. Cancer Res. 64:7011–7021. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Plummer RS, Shea CR, Nelson M, Powell SK,
Freeman DM, Dan CP and Lang D: PAX3 expression in primary melanomas
and nevi. Mod Pathol. 21:525–530. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ryu B, Kim DS, Deluca AM and Alani RM:
Comprehensive expression profiling of tumor cell lines identifies
molecular signatures of melanoma progression. PLOS One. 2:e5942007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu F, Cao J, Lv J, Dong L, Pier E, Xu GX,
Wang RA, Xu Z, Goding C and Cui R: TBX2 expression is regulated by
PAX3 in the melanocyte lineage. Pigment Cell Melanoma Res.
26:67–77. 2013. View Article : Google Scholar
|
|
13
|
Liu Y, Zhu H, Liu M, Du J, Qian Y, Wang Y,
Ding F and Gu X: Downregulation of Pax3 expression correlates with
acquired GFAP expression during NSC differentiation towards
astrocytes. FEBS Lett. 585:1014–1020. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gomes FC, Paulin D and Moura Neto V: Glial
fibrillary acidic protein (GFAP): Modulation by growth factors and
its implication in astrocyte differentiation. Braz J Med Biol Res.
32:619–631. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen J, Xia L, Wu X, Xu L, Nie D, Shi J,
Xu X, Ni L, Ju S, Wu X, et al: Clinical significance and prognostic
value of PAX3 expression in human glioma. J Mol Neurosci. 47:52–58.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia L, Huang Q, Nie D, Shi J, Gong M, Wu
B, Gong P, Zhao L, Zuo H, Ju S, et al: PAX3 is overexpressed in
human glioblastomas and critically regulates the tumorigenicity of
glioma cells. Brain Res. 1521:68–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cebolla B and Vallejo M: Nuclear factor-I
regulates glial fibrillary acidic protein gene expression in
astrocytes differentiated from cortical precursor cells. J
Neurochem. 97:1057–1070. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cao L, Yu Y, Bilke S, Walker RL,
Mayeenuddin LH, Azorsa DO, Yang F, Pineda M, Helman LJ and Meltzer
PS: Genome-wide identification of PAX3-FKHR binding sites in
rhabdomyosarcoma reveals candidate target genes important for
development and cancer. Cancer Res. 70:6497–6508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Blair A, Hogge DE, Ailles LE, Lansdorp PM
and Sutherland HJ: Lack of expression of Thy-1 (CD90) on acute
myeloid leukemia cells with long-term proliferative ability in
vitro and in vivo. Blood. 89:3104–3112. 1997.PubMed/NCBI
|
|
20
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Guo LP, Chen LZ, Zeng YX and Lu
SH: Identification of cancer stem cell-like side population cells
in human nasopharyngeal carcinoma cell line. Cancer Res.
67:3716–3724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ignatova TN, Kukekov VG, Laywell ED,
Suslov ON, Vrionis FD and Steindler DA: Human cortical glial tumors
contain neural stem-like cells expressing astroglial and neuronal
markers in vitro. Glia. 39:193–206. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rafii S: Circulating endothelial
precursors: Mystery, reality, and promise. J Clin Invest.
105:17–19. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Uchida N, Buck DW, He D, Reitsma MJ, Masek
M, Phan TV, Tsukamoto AS, Gage FH and Weissman IL: Direct isolation
of human central nervous system stem cells. Proc Natl Acad Sci USA.
97:14720–14725. 2000. View Article : Google Scholar : PubMed/NCBI
|