Introduction
Breast cancer, a leading type of cancer occurring in
women, tends to invade into adjacent regions and to metastasize to
lymph nodes and adjacent organs (1). During the process of metastasis,
tumor-suppressor genes are inactivated, which may be responsible
for breast cancer metastasis (2).
Promoter hypermethylation is a type of epigenetic alteration
associated with gene silencing. Aberrant hypermethylation of tumor
suppressor genes is an important epigenetic event in the
development and progression of breast cancer (3).
PAQR3, also known as Raf kinase trapping to Golgi
(RKTG), belongs to the family of progestin and adipoQ receptor
(PAQR) and is a seven-transmembrane protein specifically localized
in the Golgi apparatus in mammalian cells (4,5).
Subsequent characterization of PAQR3 indicates that it might
negatively modulate Ras-mediated signaling by isolating Raf kinase
to the Golgi apparatus (4). PAQR3
acts as a tumor suppressor mainly via its inhibitory activity on
Raf/MEK/ERK signaling (6–8). For example, the PAQR3 expression level
was markedly decreased in colorectal cancer samples compared with
adjacent normal tissues and the expression level of PAQR3 was
inversely associated with tumor grade in colorectal cancer samples
(9). PAQR3 also inhibited cell
proliferation, migration, sprouting and angiogenesis of endothelial
cells, and the expression level of PAQR3 was found to be
significantly downregulated in clinical clear-cell renal cell
carcinoma samples, with an inverse correlation with VEGF expression
level (7). Furthermore, it was
reported that PAQR3 was downregulated in gastric cancer and was
closely associated with metastasis progression and survival in
patients with gastric cancer (10).
Moreover, some data strongly suggest that downregulation of PAQR3
promotes tumor metastasis and proliferation through induction of
ERK phosphorylation in osteosarcoma (11). Recently, it has been reported that
PAQR3 expression is downregulated in human breast cancers and plays
an important role in carcinogenesis. However, the molecular
mechanisms of expression regulation that lead to tumorigenesis and
progression of breast cancer are still not clearly understood.
In the present study, we reported that PAQR3 is
important for breast tumorigenesis. Decreased PAQR3 expression was
observed in breast cancer patient samples. PAQR3 overexpression in
breast cancer cells inhibited tumor growth and invasion in
vitro. We examined the methylation status of PAQR3 in breast
cancer and matched non-tumor samples and determined whether
promoter methylation was associated with decreased gene expression
in breast cancer cell lines. We also examined associations between
PAQR3 methylation and several clinicopathological parameters, and
concluded that PAQR3 reactivation is associated with demethylation
of the PAQR3 gene by 5-Aza-dC treatment in MCF-7 cells.
Materials and methods
Tissue samples and cell lines
A total of 46 breast cancer tissues and paired
adjacent non-tumor tissues obtained from surgically resected
specimens at the Affiliated Hospital of Nantong University during
the period from 2010 to 2011 were analyzed. All specimens were
immediately frozen in liquid nitrogen and kept at −80°C until RNA
and protein extractions were performed. None of the patients had
received any preoperative adjuvant therapy. The resected tissue
specimens from these patients were fixed in 10% formalin and
embedded in paraffin. Written informed consent was obtained from
all patients. The study was approved by the Ethics Committee of the
Affiliated Hospital of Nantong University.
Human breast cancer cell lines MCF-7, MDA-MB-231,
MDA-MB-453, MDA-MB-468 and T47D were cultured in medium
supplemented with 10% fetal bovine serum (FBS) (Gibco) at 37°C in a
5% CO2 incubator. For overexpression of endogenous
PAQR3, the coding sequence of PAQR3 was amplified and subcloned
into the pcDNA3.1(+) vector (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's instructions.
Real-time quantitative PCR
Total RNA was extracted using the TRIzol protocol
(Invitrogen). cDNA was subsequently synthesized from total RNA
using an Omniscript RT kit (Qiagen, Valencia, CA, USA). A
25-μl reaction mixture contained 1 μl of cDNA from
samples, 12.5 μl of 2X Fast EvaGreen™ qPCR Master Mix, 1
μl primers (10 mM), and 10.5 μl of RNase/DNase-free
water. The amplification conditions for 40 cycles consisted of
denaturation at 96°C for 2 min, annealing at 96°C for 15 sec, and
extension at 60°C for 1 min. Quantitative PCR analysis was then
performed for PAQR3 mRNA expression, and data were normalized to
GAPDH levels and determined by the 2−ΔΔCt method. All
analyses were performed using Eppendorf Mastercycler® ep
realplex (2S; Eppendorf, Hamburg, Germany). The sequences of the
primers for PAQR3 were as follows: PAQR3 forward,
5′-TTCAAGACCCACATCAACC-3′ and reverse,
5′-TTTCCCTTGTATTTCCATTC-3′.
Western blotting
Total protein was extracted by lysis buffer
containing protease inhibitors (Promega, Madison, WI, USA). Equal
amounts of protein were separated by 12% polyacrylamide gel
electrophoresis and then transferred to a polyvinylidene fluoride
(PVDF) membrane. After being blocked with 5% non-fat milk, the
membrane was incubated with the primary antibodies overnight at
4°C. A goat polyclonal antibody against PAQR3 (1:500; Santa Cruz
Biotechnology, USA) was used and membranes were washed three times
in TBST for 5 min and subsequently incubated with a secondary
antibody, anti-goat IgG-conjugated IRDye 800 (1:5,000; Rockland,
Gilbertsville, PA, USA) at room temperature for 2 h, followed by
scanning with an Odyssey Infrared Imaging system (LI-COR
Biosciences, Lincoln, NE, USA), and analyzed with PDQuest 7.2.0
software (Bio-Rad). β-actin was used as the loading control.
Methylation-specific PCR and bisulfite
sequencing PCR
The methylation status of the PAQR3 promoter was
determined by methylation-specific PCR (MSP) and bisulfite
sequencing PCR (BSP). Genomic DNA extracted from tissues and cells
was modified with bisulfite reagents following the manufacturer's
instructions. This modification converts unmethylated cytosine to
thymine, whereas methylated cytosine remained unchanged. PCR
amplification was performed using 2.0 μl of
bisulfite-modified DNA in a total volume of 50 μl reaction
containing 2 μl of each primer, 5 μl of 10X DreamTaq
Buffer, and 2.0 mM dNTP Mix and 1.25 U DreamTaq (Fermentas). The
MSP conditions were as follows: 94°C for 5 min, 40 cycles of 94°C
for 30 sec, 60°C for 30 sec, 72°C for 45 sec and 72°C for 10 min.
Methylation-specific PCR products were analyzed by a 2% agarose
gel. Forward and reverse primers for the methylated sequence (M)
were 5′-TTGTTGAAGAGCGCG TATTATATC-3′ and
5′-TAAAAAACCCGAAAATCTACTCGTA-3′, respectively, and for the
unmethylated sequence (U) 5′-TTGTTGAAGAGTGTGTATTATATTGA-3′ and
5′-TAAAAAACCCAAAAATCTACTCATA-3′, respectively. Moreover, amplified
MSP products were analyzed using BioEdit and ClustalW alignment
tools. The BSP conditions were as follows: 94°C for 5 min, 40
cycles of 94°C for 30 sec, 56°C for 30 sec, 72°C for 45 sec and
72°C for 10 min, and directly sequenced using the ABI 3700
automated sequencing system (Applied Biosystems, Foster City, CA,
USA). Forward and reverse primers for BSP were
5′-GATGTATTAGAAGTTGTTGAAGAG-3′ and
5′-AACAAAAAAAATATAAATAAAAAAAA-3′, respectively.
Treatment of cells with 5-Aza-dC
MCF-7 cells were seeded at a density of
5×105 cells/well in 6-well culture plates in Dulbecco's
modified Eagle's medium (DMEM) (Invitrogen) containing 10% FBS, and
incubated in a humidified atmosphere of 5% CO2 at 37°C.
After overnight culture, the cells were incubated with medium
containing 0, 5 and 10 μmol/l 5-aza-2′-deoxycytidine
(5-Aza-dC) (Sigma, St. Louis, MO, USA) for 3 days. Total RNA and
protein were isolated after treatment, and PAQR3 mRNA and protein
were analyzed by qPCR and western blotting as aforementioned.
Cell invasion assay
For the invasion assays, the cells were suspended in
serum-free medium and plated in duplicate in the top well of
Matrigel invasion chambers (8-mm pore size; Corning, Inc., Corning,
NY, USA). Complete medium was placed in the lower chamber and cells
were allowed to invade for 24 h at 37°C in 5% CO2.
Non-invading cells in the upper chamber were removed using a cotton
swab, and cells on the lower chamber were fixed in methanol and
stained with 0.1% crystal violet. The number of invasive cells was
counted in three random fields per experiment from three
independent experiments.
Colony formation assay
Cells were seeded to 1.0×103 cells/well
in 6-well plates and cultured in DMEM medium for 3 weeks at 37°C in
5% CO2. Surviving colonies were stained with 0.5%
crystal violet and visible colonies were counted. All the
experiments were performed in triplicate wells three times.
Statistical analysis
All quantified data represent an average of at least
triplicate samples. SPSS 17.0 (SPSS, Inc., Chicago, IL, USA) was
used for statistical analysis. Data are presented as means ± SD.
One-way ANOVA or two-tailed Student's t-test was used for
comparisons between groups. A chi-square test or Fischer's exact
test was used to identify differences between categorical
variables. Survival analysis was performed using the Kaplan-Meier
method. P<0.05 was considered to indicate statistically
significant results.
Results
Expression of PAQR3 is downregulated in
breast cancer tissues
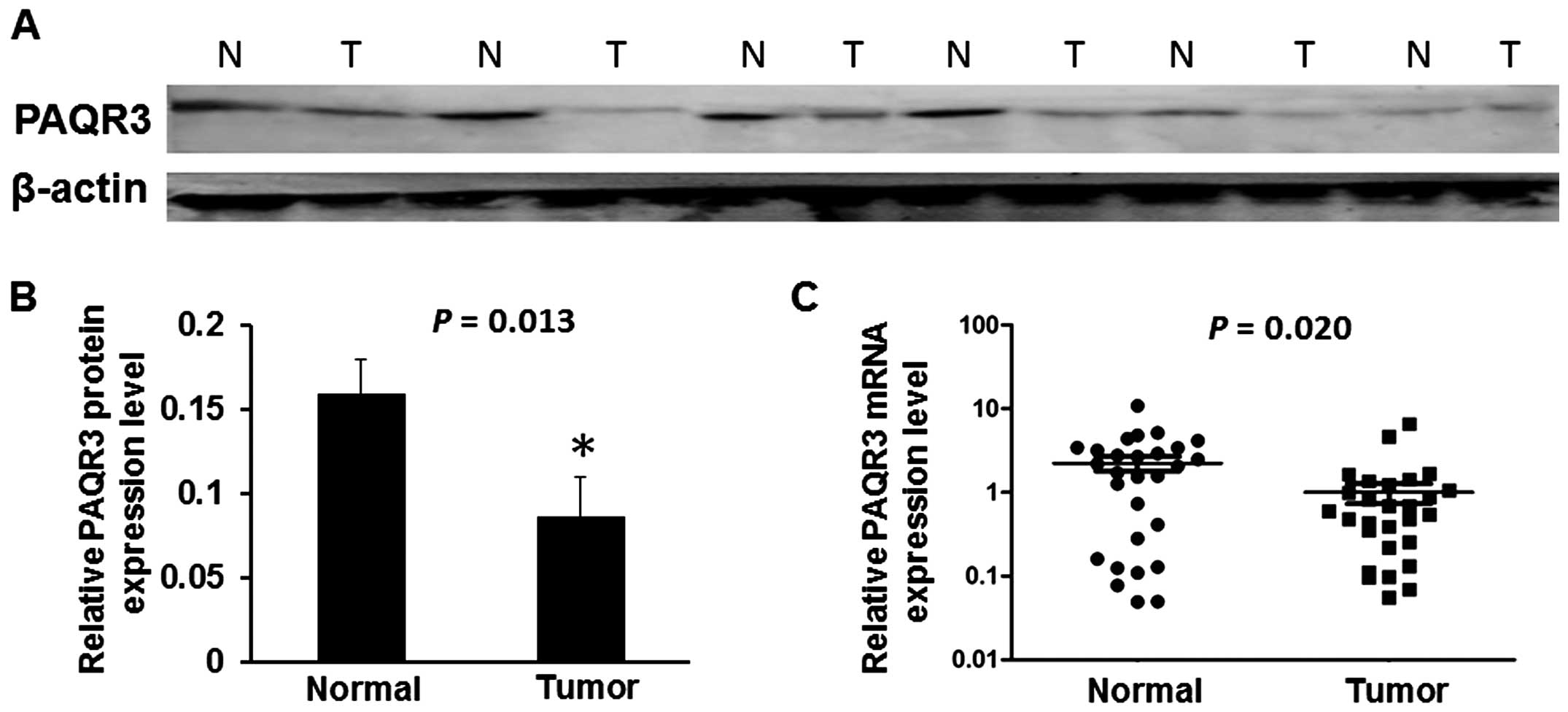
In order to assess the expression of PAQR3 in breast
cancer tissues, we performed real-time PCR and western blotting to
measure the expression of PAQR3 mRNA and protein in 28 freshly
collected breast cancer tissues and corresponding paracancerous
normal tissues. We found that the average relative expression of
PAQR3 mRNA was mar kedly downregulated compared to that in the
corresponding adjacent non-cancerous normal tissues (P=0.020,
Fig. 1C). We also found, compared
with adjacent non-tumor tissues, that the level of PAQR3 protein in
breast cancer tissues was significantly decreased in 22 of the 28
cases (P=0.013, Fig. 1B). Six cases
represented the control non-tumor tissues (Fig. 1A).
Overexpression of PAQR3 inhibits cell
invasion and cell colony formation in vitro
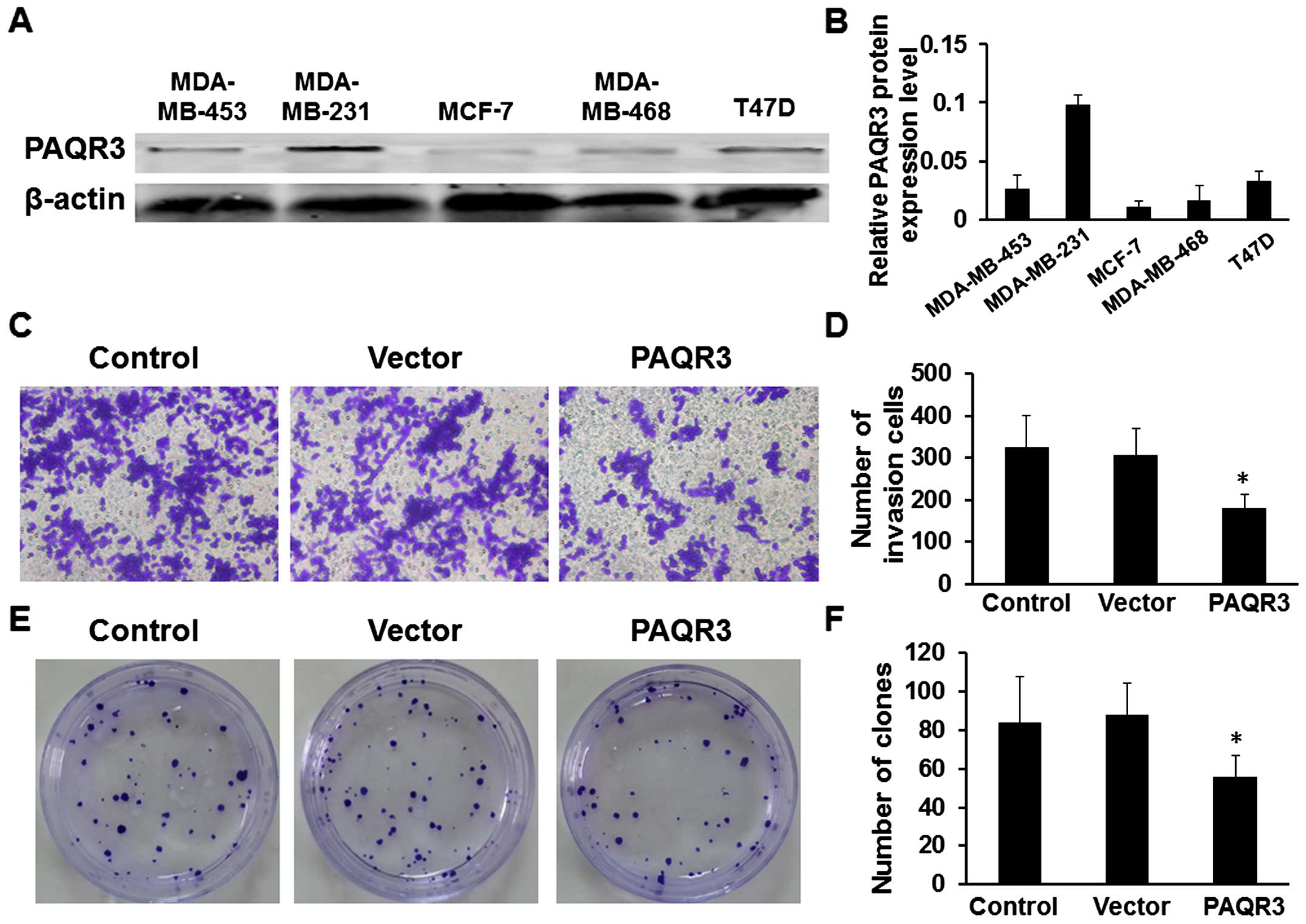
We first detected the expression level of the PAQR3
protein in cell lines, and five breast cancer cell lines were
analyzed using western blotting. The expression of PAQR3 was
detected in all cell lines and the lowest PAQR3 expression level
was detected in the MCF-7 cell line when compared with the other
breast cancer cell lines (Fig. 2A and
B). In the present study, we chose the MCF-7 cells for further
investigation, and then, we evaluated the potential role of PAQR3
on cellular invasion by Transwell assays in the MCF-7 cell line.
MCF-7 cells were transfected with the PAQR3 overexpressing or empty
vector plasmid and seeded in the chamber and their invasion
abilities were determined 48 h later. The results showed that
overexpression of PAQR3 was associated with a significant reduction
of invasion ability compared to the empty vector (Fig. 2C and D, P<0.05). We further
evaluated the potential effect of PAQR3 on cell colony formation
and found that the overexpres sion of PAQR3 significantly inhibited
the colony formation of the MCF-7 cells (Fig. 2E and F, P<0.05).
Promoter hypermethylation is involved in
decreased PAQR3 expression
It has been reported that hypermethylation is one of
the main causes of decreased gene expression in most types of
cancer. To identify whether reduced PAQR3 expression is due to
promoter hypermethylation, we studied the methylation status of the
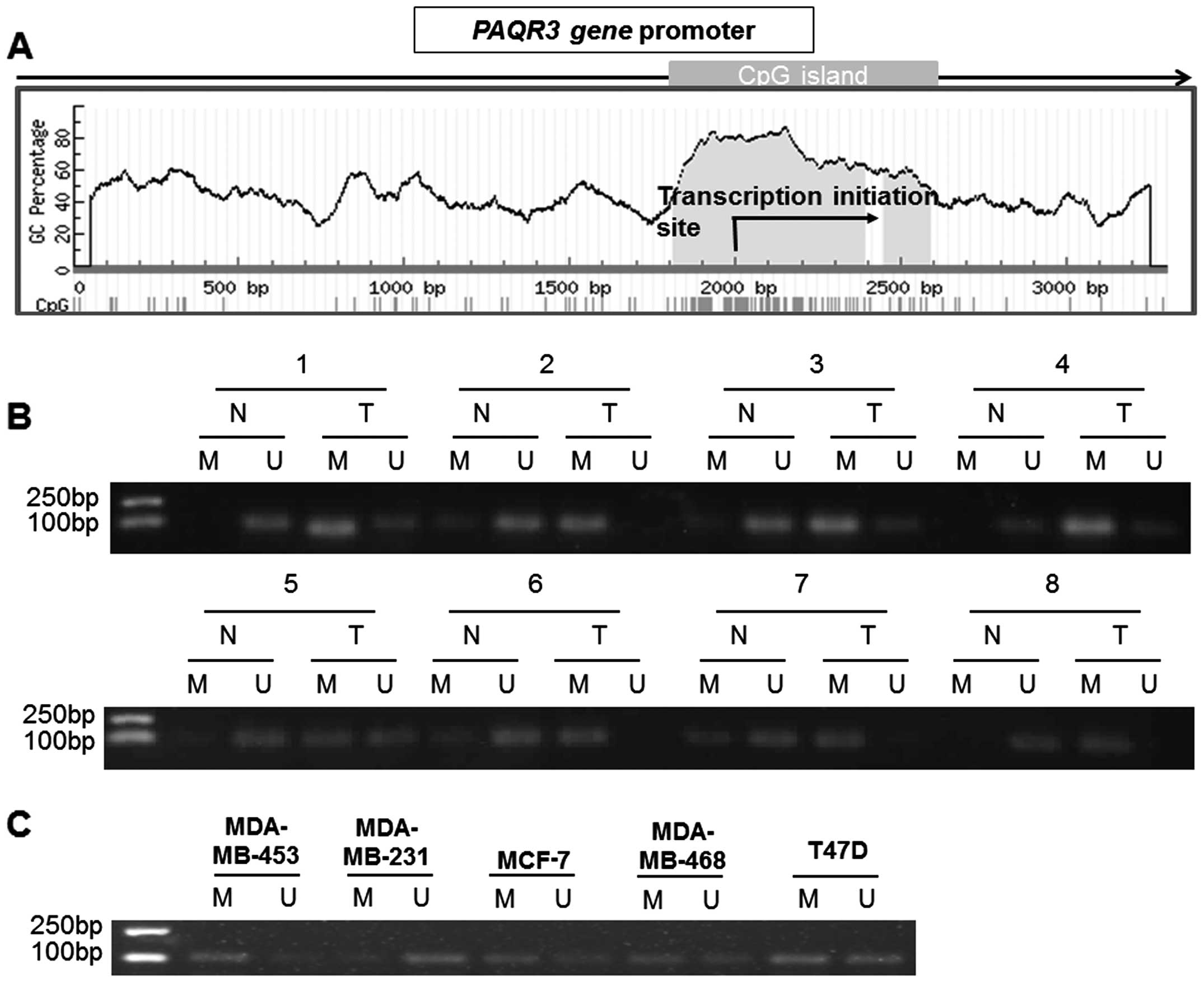
PAQR3 gene promoter in breast cancer. We searched for CpG islands
in the PAQR3 gene promoter by using the online accessible software
MethPrimer (Fig. 3A). Then we
analyzed the methylation status of the PAQR3 gene promoter in
cancerous tissue samples and their paired adjacent non-tumor
tissues from 46 breast cancer patients by using MSP analysis. As
expected, we observed that hypermethylation of PAQR3 was detected
in 71.8% (33/46) of the breast cancer tissues, and 28.2% (13/46) of
the adjacent non-tumor tissues. The difference in PAQR3 methylation
between breast cancer and adjacent non-tumor tissue specimens was
significant (P<0.001). Representative MSP results are shown in
Fig. 3B, and the first to the sixth
sample in Fig. 3B correspond to the
six cases in Fig. 1A. We also
examined the DNA methylation status of the PAQR3 gene promoter in
breast cancer cell lines. Data from MSP analysis showed
hypermethylation of the PAQR3 gene promoter in the MCF-7 and
MDA-MB-453 cells, and hypomethylation in the MDA-MB-231 cells
(Fig. 3C).
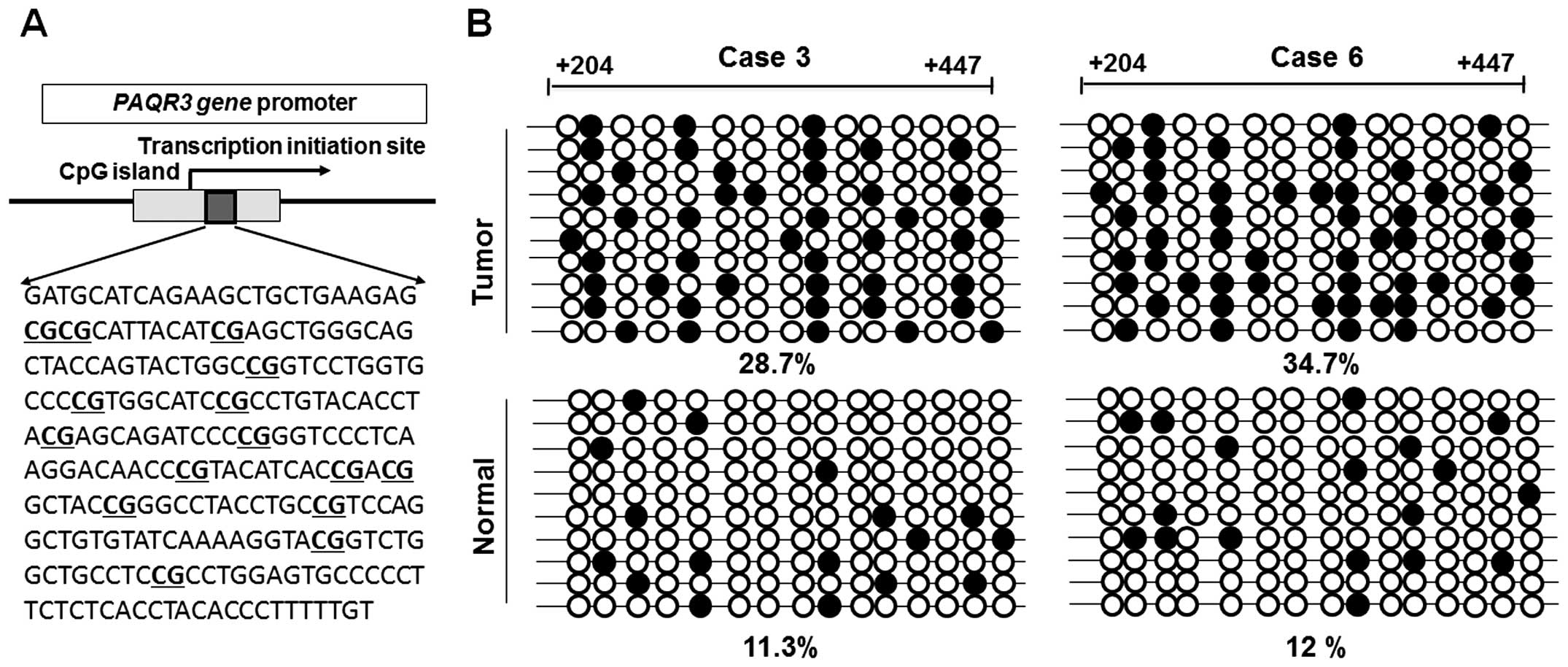
To further illustrate the methylation status of the
PAQR3 promoter, we performed bisulfite sequencing around the
promoter region of the PAQR3 gene in some of the breast cancer
tissues. Specific primers without CpG sites were used to amplify
the region spanning position from +204 to +447. The sequence
including the 15 CpG sites is shown in Fig. 4A. Bisulfite sequencing of 10
individual clones of PCR products from breast cancer tissues
revealed densely methylated CpGs within the promoter region
compared with the non-tumor tissues (P<0.05, Fig. 4B). These results indicated that
hyper-methylation may be involved in the transcriptional repression
of PAQR3.
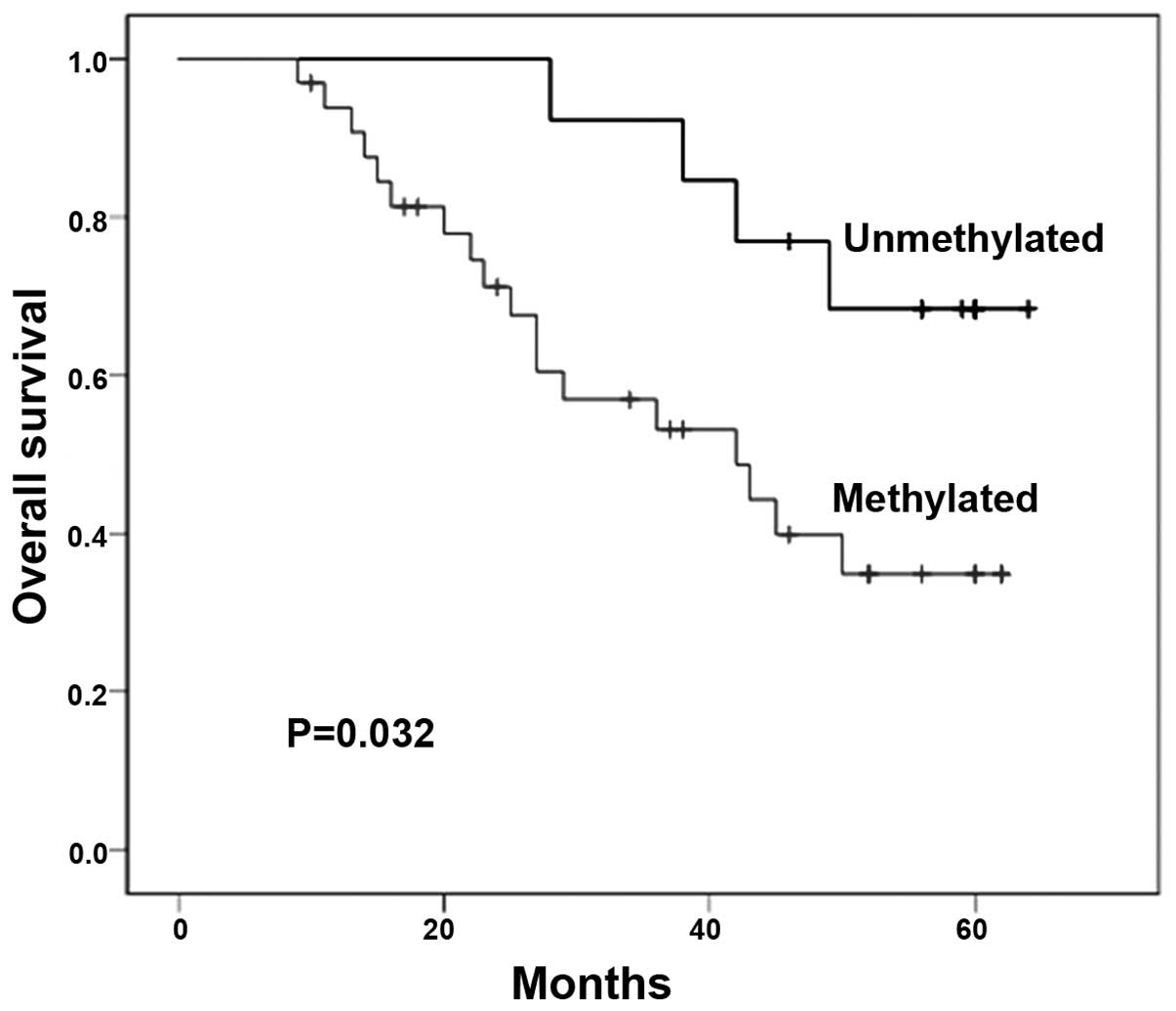
PAQR3 promoter methylation is associated
with clinicopathological features and poor prognosis
The correlations between methylation of PAQR3 and
the clinicopathologic features of these patients are summarized in
Table I. We found that the PAQR3
gene promoter methylation status was related to tumor lymph node
metastasis (P=0.010), but there was no significant difference in
clinicopathologic features, including age, tumor size, TNM stage,
ER, PR and HER2/neu status between methylated and unmethylated
tumors from these patients. Moreover, hypermethylation of the PAQR3
promoter in breast cancer was significantly correlated with poor
prognosis by Kaplan-Meier curve analysis with the log-rank test
(χ2=4.598, P=0.032, Fig.
6).
 | Table IClinicopathological parameters of the
breast cancer samples and PAQR3 methylation. |
Table I
Clinicopathological parameters of the
breast cancer samples and PAQR3 methylation.
| Clinical
parameters | Total (n=46) | PAQR3 expression
| P-valuea |
|---|
| Methylated
(n=33) | Unmethylated
(n=13) |
|---|
| Age (years) | | | | 0.068 |
| <50 | 22 | 13 | 9 | |
| ≥50 | 24 | 20 | 4 | |
| Tumor size (cm) | | | | 0.278 |
| <2 | 19 | 12 | 7 | |
| ≥2 | 27 | 21 | 6 | |
| Lymph node
metastasis | | | | 0.010b |
| Positive | 21 | 19 | 2 | |
| Negative | 25 | 14 | 11 | |
| TNM stage | | | | 0.057 |
| I + II | 29 | 18 | 11 | |
| III | 17 | 15 | 2 | |
| ER status | | | | 0.667 |
| Positive | 26 | 18 | 8 | |
| Negative | 20 | 15 | 5 | |
| PR status | | | | 0.887 |
| Positive | 24 | 17 | 7 | |
| Negative | 22 | 16 | 6 | |
| HER2/neu
status | | | | 0.161 |
| Positive | 18 | 15 | 3 | |
| Negative | 28 | 18 | 10 | |
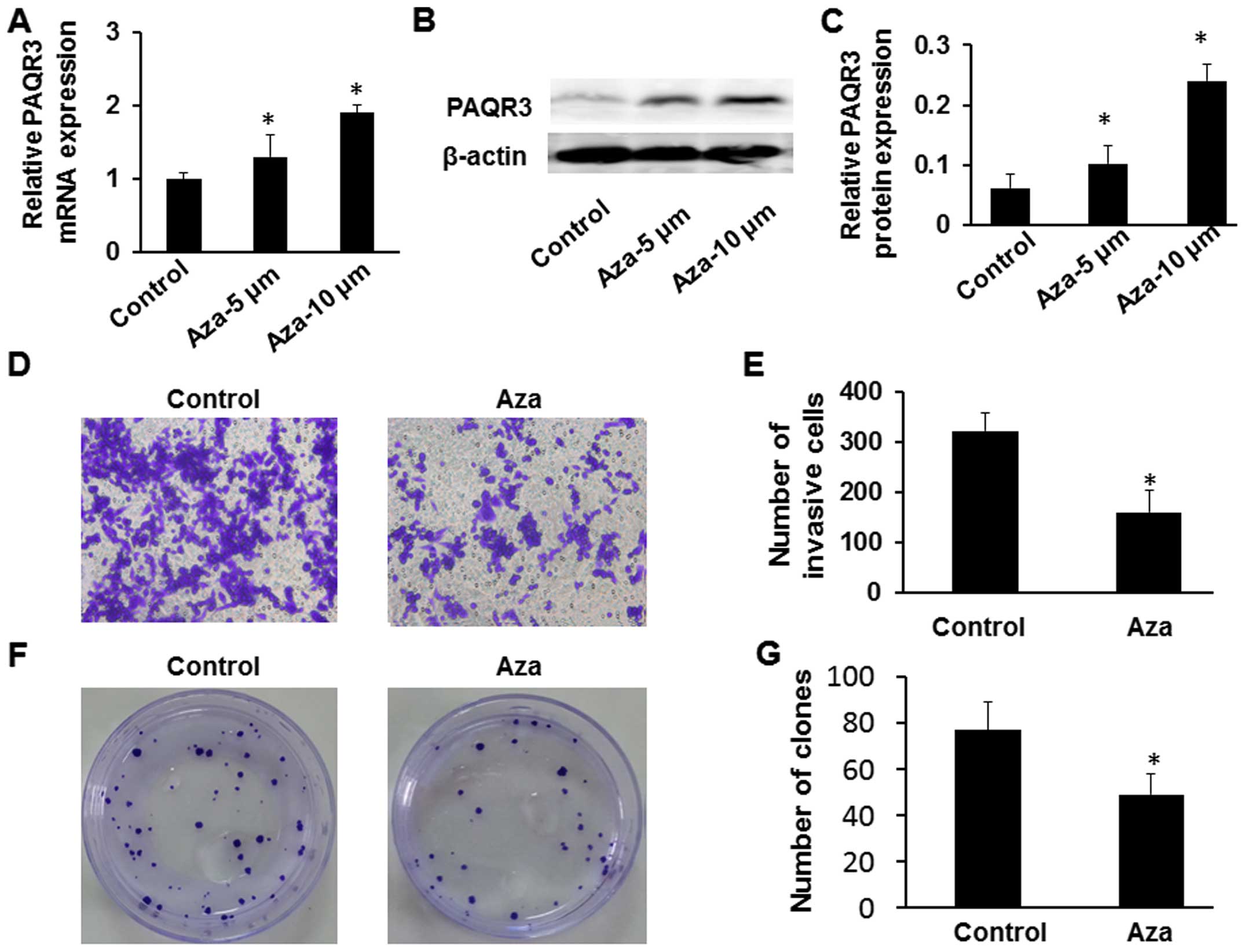
Reactivation of PAQR3 expression after
treatment with 5-Aza-dC
To further confirm that aberrant methylation was
responsible for suppressing PAQR3 expression, we treated the MCF-7
breast cancer cells with the demethylating agent 5-Aza-dC. We found
that the expression of PAQR3 mRNA and protein was significantly
elevated in the MCF-7 cells with the highest expression occurring
at a concentration of 10 μmol/l (Fig. 5A–C). Moreover, to further detect
whether the reactivation of PAQR3 expression can regulate breast
cancer proliferation and invasion, we analyzed the capability of
growth and invasion in the MCF-7 cells using colony formation and
Transwell assays. The Transwell assay showed that the number of
invading cells that were treated with 5-Aza-dC was markedly reduced
compared with the untreated group (Fig.
5D and E, P<0.05). A significant reduction in colony numbers
was observed in the MCF-7 cells treated with 5-Aza-dC, compared
with the control untreated group in the plate assays (Fig. 5F and G, P<0.05). Thus, it is
clear that the colo ny formation rate and invasive capacity of
MCF-7 cells treated with 5-Aza-dC was markedly decreased compared
with the untreated group as a result of the effect of
demethylation.
Discussion
The genomic map of the PAQR3 gene, which resides on
chromosome 4q21.21 and encodes one protein in most species, has
been described (4). In contrast to
the other PAQR family members, PAQR3 diverged quite early from the
family and exhibited relatively independent differentiation
(12). On the one hand, PAQR3 was
proven to regulate cell proliferation and invasion by inhibiting
ERK phosphorylation. On the other hand, PAQR3 can block cell
proliferation and survival due to the negative regulation of the
Ras/MAPK and the PI3K/Akt pathways, or promote tumor metastasis and
proliferation through induction of ERK phosphorylation (4,11,13).
Hence, we determined that the expression level of PAQR3 is closely
associated with the progression and metastasis of cancers (9). Collectively, these findings indicated
that PAQR3 is actively implicated in the regulation of cell
proliferation and migration and plays an important role in tumor
development (6,14). In the present study, the results
from qPCR and western blotting showed that PAQR3 mRNA and protein
expression was significantly downregulated in breast cancer tissues
demonstrating that PAQR3 acted as a tumor suppressor gene, as
previously reported in other studies as well (15). We also detected the expression level
of the PAQR3 protein in five breast cancer cell lines by western
blotting, and found that the lowest PAQR3 expression level was
detected in the MCF-7 cell line, and the highest expression level
in the MDA-MB-231 cells. However, the proliferation and migration
potential of the MCF7 cells were less than these parameters in the
MDA-MB-231 cells and it is unknown why PAQR3 expression and
aggressiveness do not correlate, but it would be significant to
investigate this in future studies. We also found that
overexpression of the PAQR3 gene inhibited the invasion and colony
formation of breast cancer cells in vitro.
Furthermore, some recent studies have indicated that
PAQR3 has a functional interaction with p53 in cancer formation and
epithelial-to-mesenchymal transition (14). In addition, some data have provided
convincing evidence that PAQR3 plays an important role in
regulating obesity and energy homeostasis accompanied by modulation
of leptin signaling (13). PAQR3
also modulates insulin sensitivity, energy metabolism, as well as
obesity in mice partly via negative regulation of PI3K (13,16).
Moreover, it is likely that the reduced expression of PAQR3 in
human tumors could relieve the inhibitory effect of PAQR3 on
histone H3 lysine 4 (H3K4) methylation, leading to facilitation of
hypoxia-induced H3K4 methylation and activation of mesenchymal gene
expression (17).
There is increasing evidence showing that gene
silencing due to aberrant DNA methylation is an early event in
carcinogenesis and may serve as a potential diagnostic and
prognostic biomarker in some cancers (18,19).
Recently, in humans, it was found that the expression level of
PAQR3 was downregulated in many types of cancers including
colorectal cancer, gastric cancer, osteosarcoma, laryngeal squamous
cell carcinoma, liver cancer and breast cancer (9,10,20–22).
In the present study, MSP analysis showed that PAQR3 promoter
methylation was significantly higher in breast cancer tissues than
that in adjacent non-tumor tissues. In addition, bisulfite
sequencing analysis of the CpG islands around the PAQR3 promoter
was used to detect the methylation status, indicating dense
methylation of CpG sites in breast cancer tissues when compared
with adjacent non-tumor tissues. Further analysis showed that
aberrant PAQR3 promoter methylation in breast cancer was associated
with poor overall survival, which can provide important prognostic
information for patients with breast cancer. These findings
suggested that epigenetic silencing of the PAQR3 promoter via
hypermethylation may be one of the major mechanisms for
inactivation of this gene in breast cancer.
The recognition that silencing of tumor suppressor
genes through promoter hypermethylation plays a significant role in
tumorigenesis has led to the clinical use of hypomethylating agents
including 5-Aza-dC (23), which has
been approved for the treatment of cancer. In the present study,
the results from qPCR and western blotting showed that 5-Aza-dC
treatment significantly promoted expression of the PAQR3 gene at
both the mRNA and protein levels in the MCF-7 cancer cell line in a
dose-dependent manner with the highest expression at 10
μmol/l, indicating that demethylation by 5-Aza-dC
contributed to the reactivation of PAQR3, thus resulting in a
decrease in colo ny formation rate and the invasive capacity of the
MCF-7 cells as detected using Transwell and colony formation
assays. However, 5-Aza-dC demethylation is not specific. It can
activate many other genes, thus 5-Aza-dC may directly promote PAQR3
gene promoter demethylation, or it may act through the regulation
of other genes that promote demethylation, indirectly mediating
PAQR3 gene expression.
In conclusion, PAQR3 promoter hypermethylation is
observed in breast cancer, and promoter hypermethylation of PAQR3
is significantly correlated with poorer survival in breast cancer
patients. Bearing these findings in mind, it will be important in
the future to elucidate the mechanism underlying PAQR3 regulation
on breast cancer cells. Further research is needed to determine the
mechanism of promoter methylation of PAQR3 in breast cancer
tumorigenesis.
Acknowledgments
This study was supported by the grants from the
Jiangsu Postdoctoral Science Foundation (grant no. 1402202C).
References
|
1
|
Hinestrosa MC, Dickersin K, Klein P, Mayer
M, Noss K, Slamon D, Sledge G and Visco FM: Shaping the future of
biomarker research in breast cancer to ensure clinical relevance.
Nat Rev Cancer. 7:309–315. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Li H, Feng J, Cui X, Huang W, Li Y,
Su F, Liu Q, Zhu J, Lv X, et al: Lin28 induces
epithelial-to-mesenchymal transition and stemness via
downregulation of let-7a in breast cancer cells. PLoS One.
8:e830832013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dworkin AM, Huang TH and Toland AE:
Epigenetic alterations in the breast: Implications for breast
cancer detection, prognosis and treatment. Semin Cancer Biol.
19:165–171. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Feng L, Xie X, Ding Q, Luo X, He J, Fan F,
Liu W, Wang Z and Chen Y: Spatial regulation of Raf kinase
signaling by RKTG. Proc Natl Acad Sci USA. 104:14348–14353. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo X, Feng L, Jiang X, Xiao F, Wang Z,
Feng GS and Chen Y: Characterization of the topology and functional
domains of RKTG. Biochem J. 414:399–406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xie X, Zhang Y, Jiang Y, Liu W, Ma H, Wang
Z and Chen Y: Suppressive function of RKTG on chemical
carcinogen-induced skin carcinogenesis in mouse. Carcinogenesis.
29:1632–1638. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Y, Jiang X, Qin X, Ye D, Yi Z, Liu
M, Bai OW, Xie X, Wang Z, et al: RKTG inhibits angiogenesis by
suppressing MAPK-mediated autocrine VEGF signaling and is
down-regulated in clear-cell renal cell carcinoma. Oncogene.
29:5404–5415. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan F, Feng L, He J, Wang X, Jiang X,
Zhang Y, Wang Z and Chen Y: RKTG sequesters B-Raf to the Golgi
apparatus and inhibits the proliferation and tumorigenicity of
human malignant melanoma cells. Carcinogenesis. 29:1157–1163. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang X, Li X, Fan F, Jiao S, Wang L, Zhu
L, Pan Y, Wu G, Ling ZQ, Fang J, et al: PAQR3 plays a suppressive
role in the tumorigenesis of colorectal cancers. Carcinogenesis.
33:2228–2235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ling ZQ, Guo W, Lu XX, Zhu X, Hong LL,
Wang Z, Wang Z and Chen Y: A Golgi-specific protein PAQR3 is
closely associated with the progression, metastasis and prognosis
of human gastric cancers. Ann Oncol. 25:1363–1372. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma Z, Wang Y, Piao T, Li Z, Zhang H, Liu Z
and Liu J: The tumor suppressor role of PAQR3 in osteosarcoma.
Tumour Biol. 36:3319–3324. 2015. View Article : Google Scholar
|
|
12
|
Tang YT, Hu T, Arterburn M, Boyle B,
Bright JM, Emtage PC and Funk WD: PAQR proteins: A novel membrane
receptor family defined by an ancient 7-transmembrane pass motif. J
Mol Evol. 61:372–380. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang L, Wang X, Li Z, Xia T, Zhu L, Liu B,
Zhang Y, Xiao F, Pan Y, Liu Y, et al: PAQR3 has modulatory roles in
obesity, energy metabolism, and leptin signaling. Endocrinology.
154:4525–4535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang Y, Xie X, Li Z, Wang Z, Zhang Y,
Ling ZQ, Pan Y, Wang Z and Chen Y: Functional cooperation of RKTG
with p53 in tumorigenesis and epithelial-mesenchymal transition.
Cancer Res. 71:2959–2968. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu X, Li Z, Chan MT and Wu WK: PAQR3: A
novel tumor suppressor gene. Am J Cancer Res. 5:2562–2568.
2015.PubMed/NCBI
|
|
16
|
Wang X, Wang L, Zhu L, Pan Y, Xiao F, Liu
W, Wang Z, Guo F, Liu Y, Thomas WG, et al: PAQR3 modulates insulin
signaling by shunting phosphoinositide 3-kinase p110α to the Golgi
apparatus. Diabetes. 62:444–456. 2013. View Article : Google Scholar :
|
|
17
|
Liu C, Zhang Y, Hou Y, Shen L, Li Y, Guo
W, Xu D, Liu G, Zhao Z, Man K, et al: PAQR3 modulates H3K4
trimethylation by spatial modulation of the regulatory subunits of
COMPASS-like complexes in mammalian cells. Biochem J. 467:415–424.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jones PA and Baylin SB: The epigenomics of
cancer. Cell. 128:683–692. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Esteller M: Cancer epigenomics: DNA
methylomes and histone-modification maps. Nat Rev Genet. 8:286–298.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu Q, Zhuang K and Li H: PAQR3 plays a
suppressive role in laryngeal squamous cell carcinoma. Tumour Biol.
37:561–565. 2016. View Article : Google Scholar
|
|
21
|
Wu HG, Zhang WJ, Ding Q, Peng G, Zou ZW,
Liu T, Cao RB, Fei SJ, Li PC, Yang KY, et al: Identification of
PAQR3 as a new candidate tumor suppressor in hepatocellular
carcinoma. Oncol Rep. 32:2687–2695. 2014.PubMed/NCBI
|
|
22
|
Li Z, Ling Z-Q, Guo W, Lu XX, Pan Y, Wang
Z and Chen Y: PAQR3 expression is downregulated in human breast
cancers and correlated with HER2 expression. Oncotarget.
6:12357–12368. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fenaux P: Inhibitors of DNA methylation:
Beyond myelodys-plastic syndromes. Nat Clin Pract Oncol. 2(Suppl
1): S36–S44. 2005. View Article : Google Scholar
|