Introduction
Pancreatic cancer (PC) is a fatal disease. It
commonly presents at an advanced stage, with synchronous metastases
and poor prognosis. PC has a median survival of 6 months and an
overall 5-year survival rate that is less than 5% (1). PC is expected to become the second
leading cause of cancer-related deaths in the US within the next
decade (2). PC is also highly
chemoresistant (3). Currently, the
most effective chemotherapies for advanced PC are a combination of
5-fluorouracil (5-FU)/leucovorin with irinotecan and oxaliplatin
(FOLFIRINOX) (4), or nab-paclitaxel
with gemcitabine, which only provide modest response rates, ranging
from 23 to 31% (5) and have limited
tolerability. Thus, discovery of new drugs or drug combination
therapies for patients with PC is critically important.
Over the past three decades, a growing number of
traditional Chinese medicines have been widely used in China as
adjuvant treatments during chemotherapy and radiotherapy for cancer
(6). One of the well-known drugs
includes the seeds of Coix lacryma-jobi (family
Cramineae), which have been widely used as a herbal medicine
for the treatment of hypertension, arthritis, cancer and
immunological disorders in China (7,8). The
oily substance extracted from coix seeds has been prepared in an
injectable emulsion form, and has successfully been used in China
to treat more than 500,000 patients for various types of cancer,
including non-small cell lung (9)
and advanced gastric cancer (10),
and hepatoma (11). In recent
years, the use of coix seed emulsion was tested in the US in a
phase II clinical trial for patients with advanced PC (12,13).
The results revealed that coix seed emulsion, when combined with
gemcitabine, can overcome chemoresistance, increase the tolerance
of patients, and significantly improve the lifespan and quality of
life of patients. Preclinical studies also indicate that coix seed
emulsion blocks the tumor cell cycle at the G2/M phase (14), and induces tumor cell apoptosis by
upregulating the expression of caspase-8 and downregulating the
expression of Bcl-2 and Cox-2 (15,16).
However, the mechanisms that underlie the synergistic effect of
coix seed emulsion and gemcitabine have not yet been
elucidated.
Molecular therapy that is targeted against specific
components of defined signaling pathways has recently attracted
much attention. Nuclear factor-κB (NF-κB) plays a critical role in
the development and progression of various human cancers that
exhibit constitutive and continuous NF-κB activity (17,18).
However, the induction of this signaling pathway is also associated
with gemcitabine chemoresistance in PC cells (19–21).
We thus reasoned that agents that block NF-κB activation may reduce
chemoresistance to gemcitabine, and may be effective when used in
combination with gemcitabine as a novel therapeutic regimen for
treating PC. We demonstrated that coix seed emulsion markedly
decreased expression levels of NF-κB in the nucleus, and of IkBα,
IKK and EGFR in the cytoplasm of Lewis lung carcinoma in
vivo (22). The emulsion also
reduced NF-κB-dependent reporter activity in a dose-dependent
manner in MDA-MB-231 breast cancer cells (16).
In the present study, we report on the synergistic
action of coix seed emulsion and gemcitabine when both are used in
PC cell lines PANC-1, AsPC-1 and BxPC-3, and also in a xenograft
mouse model. We also assessed NF-κB activity and the expression
levels of apoptosis-related components that lie downstream of
NF-κB, to explore the mechanism of action of coix seed emulsion
plus gemcitabine in treating PC.
Materials and methods
Antibodies and reagents
Antibodies were purchased as follows: anti-rabbit
cyclooxygenase-2 (Cox-2), survivin, Bcl-2, Bax, β-actin (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), cleaved-polyADP ribose
polymerase (PARP), Ki-67 and p65 antibody (Cell Signaling
Technology, Danvers, MA, USA). Coix seed emulsion
(Kanglaite® injection) was obtained from Zhejiang
Kanglaite Pharmaceutical Co., Ltd. (Hangzhou, China).
Gemcitabine-HCl was purchased from Eli Lilly and Company
(Indianapolis, IN, USA). All other chemical reagents were purchased
from Sigma-Aldrich (St. Louis, MO, USA).
Cell culture
The human PC cell lines PANC-1, BxPC-3 and AspC-1,
and the human embryonic kidney cell line HEK-293T were obtained
from the Type Cell Collection of the Chinese Academy of Sciences
(Shanghai, China). PANC-1, BxPC-3 and AspC-1 cells were cultured in
RPMI-1640 medium, and HEK-293T in Dulbecco's modified Eagle's
medium (DMEM) (Gibco, Grand Island, NY, USA) in a 5% CO2
and 95% air humidified atmosphere at 37°C. All media were
supplemented with 10% fetal bovine serum (Gibco) and 100 U/ml of
penicillin-streptomycin.
Xenograft mouse model
Male nude BALB/c mice (6 week of age) were obtained
from the Zhejiang Chinese Medical University Experimental Animal
Center (Hangzhou, China), and housed under pathogen-free conditions
and were provided with standard laboratory food and water. All
animal-handling protocols were approved by the Experimental Animal
Use and Management Committee of Zhejiang Chinese Medical
University. To induce tumor growth, the right flank of each mouse
was subcutaneously injected with 5×106 PC BxPC-3 cells
suspended in 0.1 ml phosphate-buffered saline (PBS). When
subcutaneous tumors developed to ~30 mm3 in size (7 days
after inoculation), the mice (n=24) were randomly divided into four
groups: a control group (0.9% saline, i.p. daily), a coix seed
emulsion alone group (12.5 ml/kg, i.p. daily), a gemcitabine alone
group (50 mg/kg, i.p. 3/week) and a coix seed emulsion and
gemcitabine combination group (coix seed emulsion 12.5 ml/kg, i.p.
daily; gemcitabine 50 mg/kg, i.p. 3/week). Body weights and tumor
volumes were measured every three days. Tumor volumes were
calculated with the formula: Tumor volume (mm3) =
maximal length × maximal width2/2. After 24 days of
treatment, all mice were overdosed with anesthesia, and tumors were
harvested and weighed. The rate of inhibition of tumor growth was
calculated using the formula: [(Mean tumor weight of the control
group - mean tumor weight of the treatment group)/mean tumor weight
of the control group] × 100%. Tumors tissue were fixed in buffered
formalin for further analyses.
Cell viability as detected by MTS
assay
Cells were seeded in 96-well microplates at a
density of 4×104 cells/ml and cultured overnight; they
were exposed to various concentrations of coix seed emulsion and
gemcitabine, alone or in combination for the desired time. Control
cells received only dimethyl sulfoxide (DMSO). Cell viability was
assessed using the 3-(4,5-dimethylthiazol-2yl)-5-
(3-carboxymethoxypheny l)-(4-sulfophenyl)-2H-tetrazolium (MTS)
assay (Promega, Madison, WI, USA) according to the manufacturer's
instructions. Cell viability is directly proportional to the
absorbance at 490 nm of a formazan product that is bio-reduced from
MTS in living cells as previously described (23). To determine the synergetic effect
between coix seed emulsion and gemcitabine, the cells were exposed
to drugs in a fixed ratio and the combination index (CI) was
calculated by CalcuSyn software (Biosoft, Cambridge, UK). All
experiments were carried out in hexaplicate, and were independently
repeated at least twice.
Caspase-3 activity assay
Apoptosis of cells after the treatment of coix seed
emulsion and gemcitabine, alone or in combination, was evaluated
with the use of the Caspase-3/CPP32 Fluorometric Assay kit
(BioVision Research Products, Mountain View, CA, USA; according to
the manufacturer's instructions), as previously described (23). Cellular lysates (50 μl) were
extracted and protein concentration was measured using BCA protein
assay reagents (Pierce, Rockford, IL, USA). Cleavage of DEVD-AFC, a
substrate of caspase-3, was quantified using a fluorescence
microplate reader, with 400 nm excitation and 505 nm emission
filters.
Protein extraction and western blot
analysis
Cells were seeded in 100-mm tissue culture dishes
and grown to 80% confluency. Medium was then replaced with fresh
medium containing selected drugs at the various concentrations. The
control group was treated with the same volume of DMSO. After 48 h,
all cells were washed twice with ice-cold PBS, collected in RIPA
lysis buffer (20 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 5 mM EDTA, 1
mM Na3VO4, pH 7.5) supplemented with a
protease inhibitor mixture (Sigma-Aldrich, St. Louis, MO, USA), and
incubated on ice for 20 min. Afterwards, the cell lysate was
centrifuged at 14,000 rpm for 10 min, and the pellet was diluted in
SDS-sample buffer. Protein concentration was determined using BCA
protein reagents (Pierce). The nuclear and cytoplasmic proteins
were extracted with the Nuclear and Cytoplasmic Protein Extraction
kit (Pierce; according to the manufacturer's instructions). The
cell lysate (30 μg) was subjected to electrophoresis on a
4–12% sodium dodecyl sulfate (SDS)-polyacrylamide gel, and then
transferred onto a polyvinylidene difluoride (PVDF) membrane
(Bio-Rad, Hercules, CA, USA). Blots were probed with appropriate
primary antibodies and IRDye® 800CW secondary antibody
(LI-COR Biotechnology, Lincoln, NE, USA). Bands were visualized by
Odyssey infrared imaging system (LI-COR Biotechnology) and
quantified by densitometry analysis, using ImageJ software (NIH,
Bethesda, MA, USA).
Cell transfection and reporter assay
HEK-293T cells (at 70% confluency) were transfected
with the NF-κB-EGFP construct (GeneChem, Shanghai, China), which
links an enhanced GFP reporter to an NF-κB response element as
previously described (16). To
prepare DNA for transfection, 10 μl Lipofectamine 2000 and 2
μg DNA were diluted with 0.2 ml of Opti-MEM (both from
Invitrogen, Carlsbad, CA, USA), according to the manufacturer's
instructions. After incubation for 24 h, the transfection rate was
tested (data not shown), and HEK-293T cells for which the
transfection rate was >90% were treated with the desired
concentration of coix seed emulsion alone, with gemcitabine alone,
with the coix seed emulsion and gemcitabine combination or with
DMSO. After an additional 48 h, the expression of EGFP in each
group was captured on a fluorescence microscope system (Olympus,
Tokyo, Japan); the cells (1×106 cells/ml) were then
washed twice with cold PBS, half of them were re-suspended in
fixing buffer and analyzed by flow cytometry (Millipore, Billerica,
MA, USA) within 1 h. The other half of the cells were lysed and the
protein concentration was quantified. Data are reported as the
average of three independent experiments.
Electrophoretic mobility shift assay
The electrophoretic mobility shift assay (EMSA) was
used to assess NF-κB activation as previously described (24). Briefly, nuclear extracts prepared
from cells treated with nuclear and cytoplasmic extraction reagents
(Pierce) were incubated with biotin-labeled double-stranded NF-κB
oligonucleotide (Beyotime, Beijing, China) for 30 min at 37°C, and
EMSA was performed, following the instructions in the LightShift
Chemiluminescent EMSA kit (Pierce).
Immunohistochemical staining and
assessment
Formalin-fixed, paraffin-embedded tumor tissues were
sectioned (4-μm) and incubated with antibodies against Ki-67
and P65, at 1:100 dilutions. All stained slides were examined by an
independent observer; and the tissue was assessed for protein
expression in neoplastic areas at a magnification of ×40.
Percentage of Ki-67 and P65 expression was recorded for each area
(cytosolic or nuclear), and then averaged for each mouse. A total
of five fields were examined in six samples from each of the
treatment groups.
Statistical analysis
All data were represented as mean ± standard
deviation (SD) for at least three independent experiments;
representative examples are shown. Statistical analysis of multiple
group comparisons was performed by one-way analysis of variance
(ANOVA). Comparisons between two groups were analyzed using
Student's t-tests. A P-value of <0.05 was considered to indicate
a statistically significant result.
Results
Coix seed emulsion inhibits the viability
and induces the apoptosis of PC cells in a dose-dependent
manner
To investigate the cytotoxicity of coix seed
emulsion on PC cell lines in vitro, we exposed BxPC-3,
PANC-1 and AsPC-1 cells for 72 h to coix seed emulsion at
increasing concentrations (0–10.0 mg/ml), and examined the cell
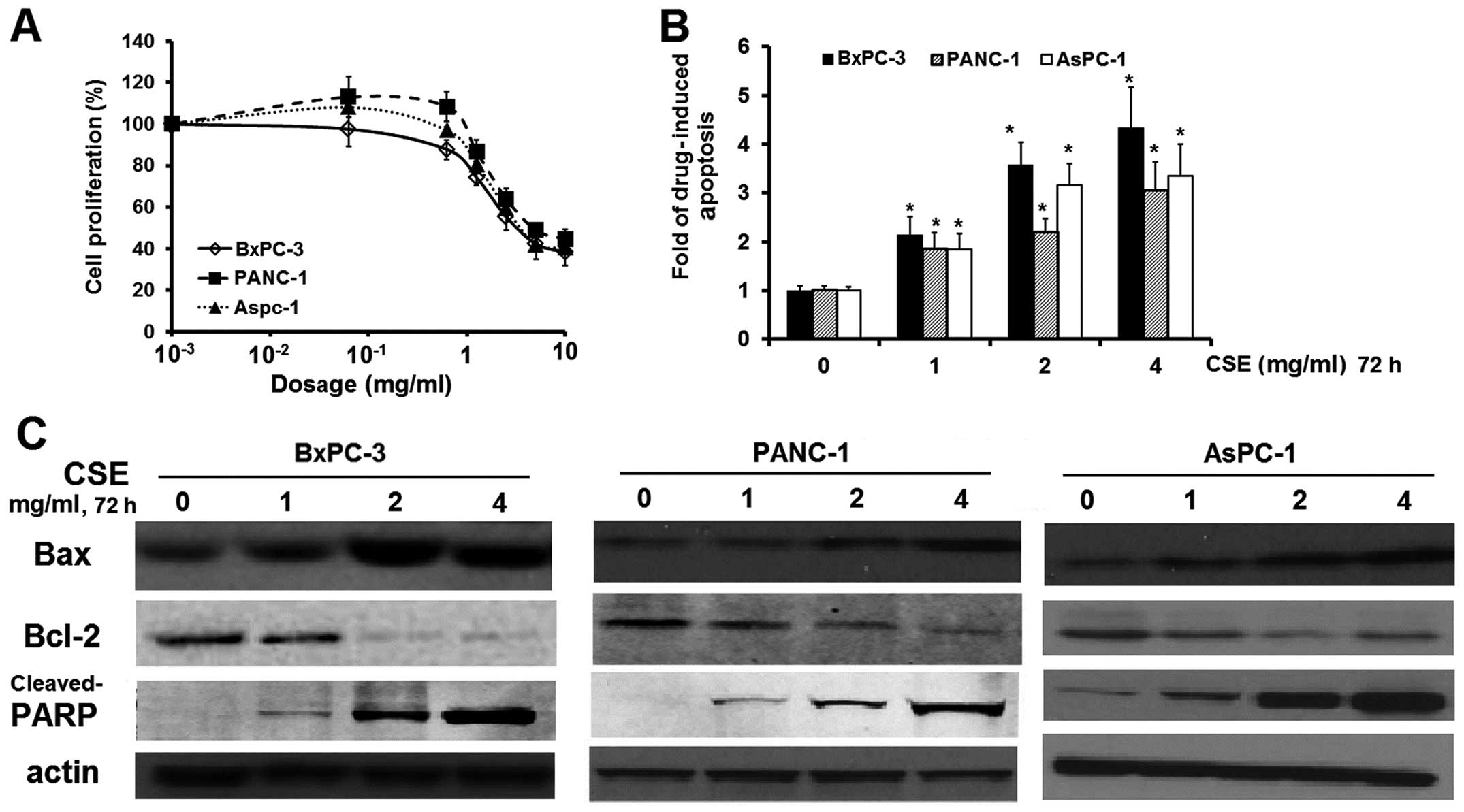
viability by the MTS assay. As shown in Fig. 1A, coix seed emulsion exhibited
dose-dependent cytotoxicity in the BxPC-3, PANC-1 and AsPC-1 cells
with an IC50 value of 1.50, 1.75 and 1.80 mg/ml,
respectively.
To further study whether emulsion treatment
decreases cell viability through apoptosis, we measured caspase-3
activity in all three cell lines and found that, compared to
untreated cells, caspase-3 activity decreased from 1.84 to
4.33-fold in the treated PC cells, in a dose-dependent manner
(Fig. 1B). To clarify the molecular
mechanism of coix seed emulsion-induced apoptosis, we determined
apoptosis-related protein expression in the BxPC-3, Panc-1 and
AsPC-1 cells, and reported that expression of the anti-apoptotic
protein Bcl-2 was downregulated, whereas that of the pro-apoptotic
protein Bax was upregulated, also in a dose-dependent manner; these
findings indicate that the apoptosis-inducing effect of coix seed
emulsion can be partly attributed to an altered Bax/Bcl-2 protein
ratio, which is a critical determinant of apoptosis (21). The caspase-3 downstream substrate
cleaved-PARP was also increased in the emulsion-treated PC cells.
Our results confirm previous studies on the use of coix seed
emulsion with breast (25) and
liver cancer cells (15). Even
though coix seed emulsion does not inhibit the viability of
cultured cancer cells as much as do other small molecular
inhibitors, it can alter apoptosis signaling in PC cells.
Pretreatment with coix seed emulsion
potentiates gemcitabine-induced cytotoxicity
Gemcitabine is the only first-line drug for patients
with advanced PC; however it only provides a modest improvement in
survival (26), in part due to the
development of drug resistance. NF-κB regulation is involved in the
formation of such resistance (27,28).
Since coix seed emulsion inhibits NF-κB activity in MDA-MB-231
breast cancer cells, we next assessed whether coix seed emulsion
enhances the cytotoxicity of gemcitabine in PC cells. We found that
pretreatment of coix seed emulsion was more efficacious in
sensitizing the cells to gemcitabine compared to co-treatment of
the two drugs or whether the emulsion was washed away before adding
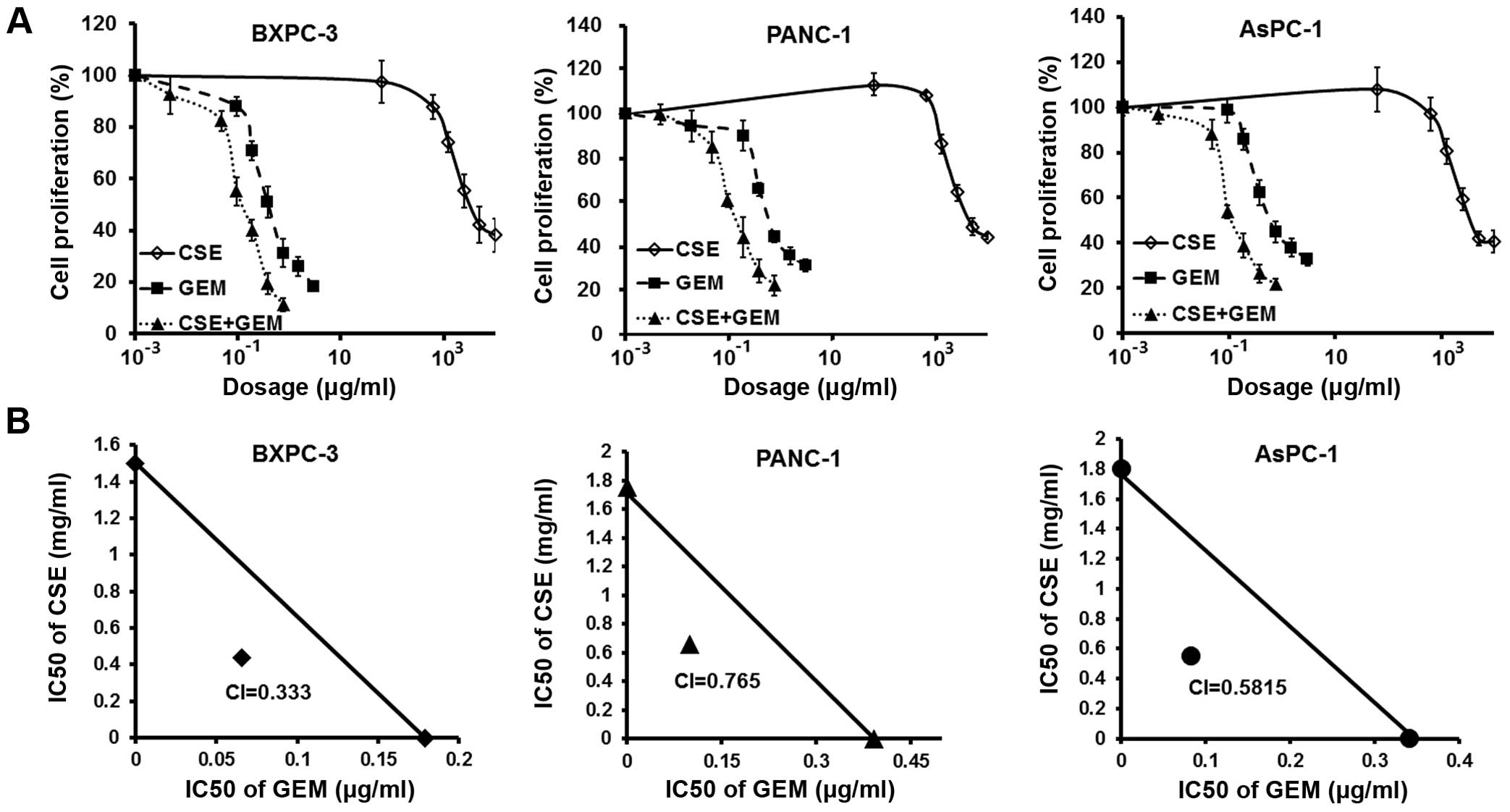
drugs (data not shown). To confirm synergism between the emulsion
and gemcitabine, we pretreated BxPC-3, PANC-1 and AsPC-1 cells with
coix seed emulsion alone (0–5.0 mg/ml) for 48 h, following by
co-treatment with varying concentrations of gemcitabine (0–0.75
μg/ml) for another 24 h. Our results showed that
pretreatment with coix seed emulsion effectively decreased the
IC50 value of gemcitabine from 0.18–0.39 to 0.066–0.099
μg/ml (Fig. 2A). We then
determined CI values for the combination treatment group. CI is a
quantitative measure of the degree of drug interaction; CI <1
indicates synergism, CI >1 indicates antagonism and CI=1
indicates an additive effect. As shown in Fig. 2B, BxPC-3, PANC-1 and AsPC-1 cells
pretreated with coix seed emulsion showed synergistic loss of cell
viability, when the pretreatment was combined with gemcitabine
(CI=0.333, 0.765 and 0.282, respectively).
Pretreatment with coix seed emulsion
augments apoptosis induced by gemcitabine in PC cells
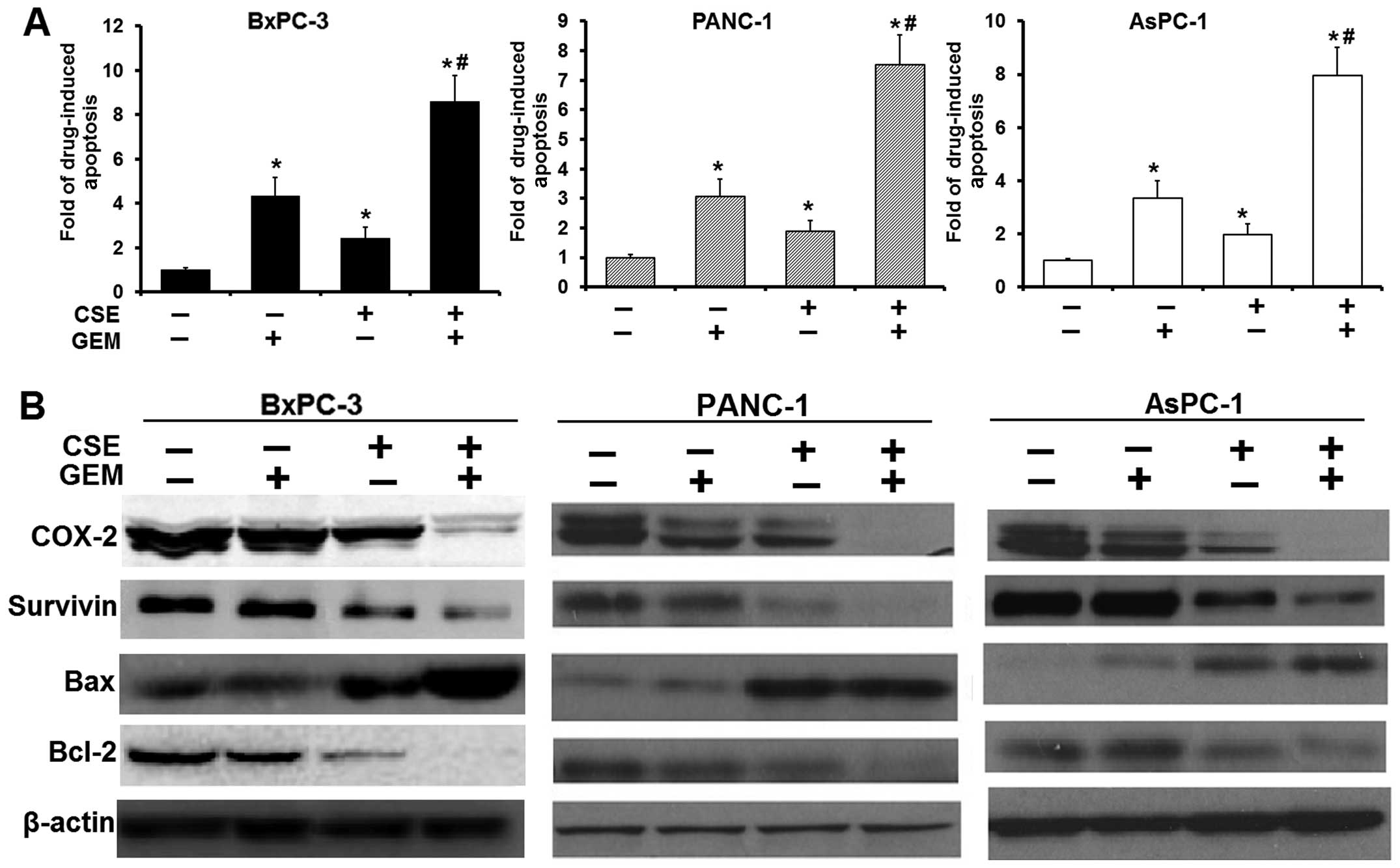
To confirm whether the enhanced loss of cell
viability resulting from pretreatment with coix seed emulsion is
mediated by apoptosis, the cells were pretreated (4.0 mg/ml) with
emulsion, followed by gemcitabine. This significantly elicited
gemcitabine-induced caspase-3 activity from 1.89- to 2.43-fold, to
7.52- to 8.61-fold (P<0.05) in all PC cell lines used (Fig. 3A), suggesting that the loss of
viable cells by coix seed emulsion and gemcitabine was due to the
induction of the cell death pathway. In agreement with these
results, we also found significant upregulation of Bax and
downregulation of Bcl-2 protein expression in the combination
treatment group (Fig. 3B), and that
the combination treatment significantly suppressed other
pro-survival molecules such as survivin and Cox-2. Our results
indicate that coix seed emulsion and gemcitabine indeed synergized
the cytotoxic effect of gemcitabine on PC cells.
Coix seed emulsion abolishes
gemcitabine-induced NF-κB activation and nucleus translocation
Since Bcl-2, survivin and Cox-2 proteins are
downstream effectors regulated by NF-κB (29,30),
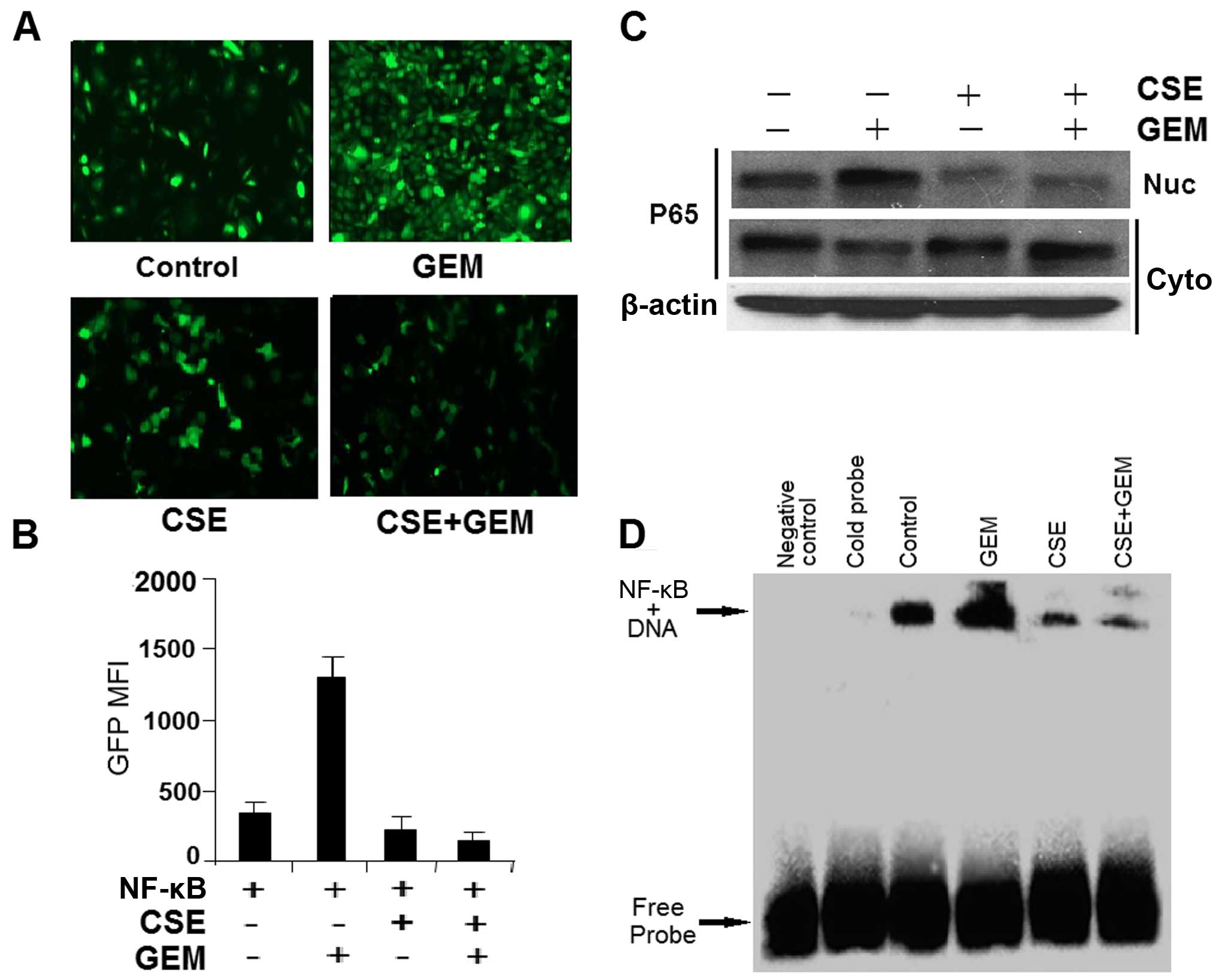
we next assessed the possible mechanistic role of NF-κB in our
experimental system. Employing an NF-κB-dependent enhanced green
fluorescent protein (EGFP) reporter assay, we directly measured the
effects of the drugs on a gene promoter element that has a
consensus NF-κB response element in HEK-293T cells. Gemcitabine
(0.18 μg/ml) treatment alone stimulated the reporter
activity; meanwhile coix seed emulsion (4.0 mg/ml) effectively
diminished gemcitabine-induced or non-induced reporter activity
(Fig. 4A and B). Activation of
NF-κB is associated with translocation to the nucleus of the
p65/Rel-A subunit of this transcription factor, as well as with the
NF-κB DNA-binding activity (31).
PANC-1 cells were exposed to coix seed emulsion (4.0 mg/ml) for 72
h, followed by 3 h of gemcitabine (0.50 μg/ml) treatment;
the nuclear and cytoplasmic fractions of the cell lysate were
subjected to electrophoresis and immunoblotting by an anti-p65
antibody. Meanwhile, an electrophoretic mobility shift assay was
used to assess the NF-κB DNA-binding activity in the nuclear
extracts. Fig. 4C shows that
gemcitabine stimulated the translocation of p65 from the cytoplasm
to the nucleus, consistent with activation of the NF-κB
transcription factor; this effect was markedly attenuated by
treating the cells with coix seed emulsion. These observations were
further supported by the finding that coix seed emulsion
sequestered the basal level of NF-κB DNA-binding activity in
unstimulated PC cells, and gemcitabine induced NF-κB DNA-binding
activity (Fig. 4D). Our results
indicate that the chemosensitizing effect of coix seed emulsion is,
in part, due to inactivation of NF-κB and its downstream genes.
Coix seed emulsion enhances the
therapeutic effects of gemcitabine in xenografts derived from
Panc-1 cells by inhibiting NF-κB activity
Based on the in vitro finding that
pretreatment of coix seed emulsion sensitizes gemcitabine-induced
cytotoxicity in PC cells, we evaluated the therapeutic advantage of
using coix seed emulsion and gemcitabine in a xenograft nude mouse
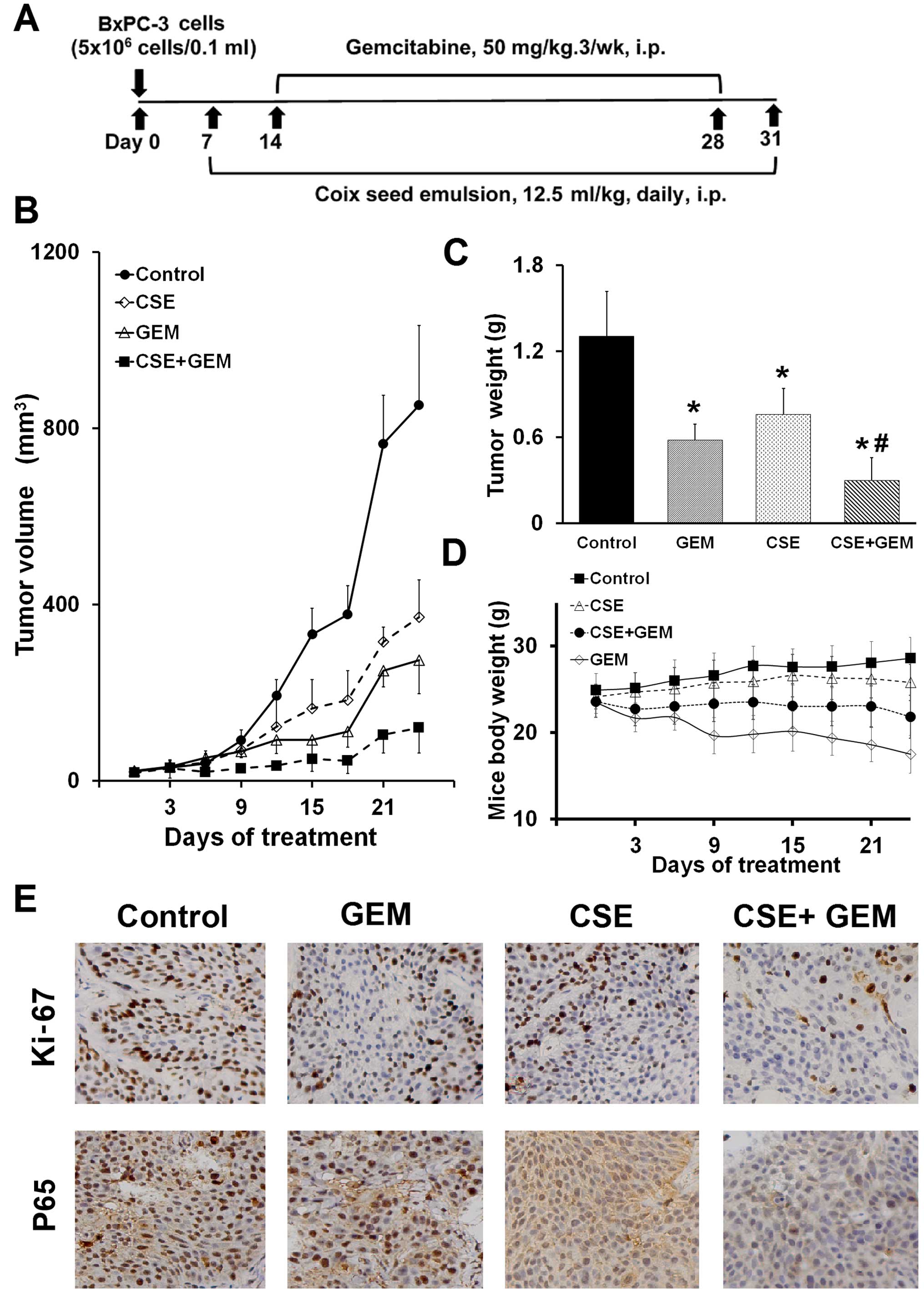
model implanted with PANC-1 cells (see experimental protocol in
Fig. 5A). Relative to the untreated
control group, administration of coix seed emulsion by
intraperitoneal injection caused a 30% reduction in tumor weight
(P<0.05), while gemcitabine treatment alone caused a 50%
reduction in tumor volume and weight (P<0.05) (Fig. 5B and C). However, the combination of
coix seed emulsion and gemcitabine demonstrated a 68% reduction in
tumor volume and weight, a result of which was superior to
treatment with coix seed emulsion alone (P<0.05) or gemcitabine
alone (P<0.05) (Fig. 5B and C).
Moreover, coix seed emulsion partly rescued the severe weight loss
induced in the mice by gemcitabine (Fig. 5D). We additionally investigated
whether the coix seed emulsion alone, or in combination with
gemcitabine, downregulated NF-κB in the tumor tissue.
Immunohistochemistry revealed significant downregulation of p65
expression in groups treated with coix seed emulsion alone
(P<0.05) and with the combination (P<0.05), compared with the
control; only modest alterations were noted in the gemcitabine
alone group (Fig. 5E). These
findings paralleled the study on a significant reduction in Ki-67
staining in tumors derived from the mice treated with the
combination of the two drugs (P<0.05), indicating diminished
cellular viability in the tumors.
Taken together, our in vivo results are
consistent with our in vitro findings and collectively
provide convincing evidence in support of the superior antitumor
efficacy of the combination of coix seed emulsion and
gemcitabine.
Discussion
We demonstrated that coix seed emulsion effectively
induces apoptosis by activating caspase-3 and increasing the
Bax/Bcl-2 ratio in pancreatic cancer (PC) cells; also, that
pretreatment of these cells with coix seed emulsion exerts
chemosensitization effects that are synergistic with those of
gemcitabine.
The major impediments to and challenges of using
available treatments for patients with advanced PC are the
dose-limiting toxicity to normal tissues (32,33)
and the increase in acquisition of drug resistance during
gemcitabine-based chemotherapies (34). Substantial evidence implicates the
constitutive activation of the transcription factor NF-κB in de
novo and acquired resistance to the therapeutic response in PC
progression (31,35). A possible strategy for overcoming
this problem is the use of combinatorial treatments (36); for example, the application of NF-κB
inhibitors (37) or natural
products (38) to inactive NF-κB
before beginning treatment with conventional therapeutics. Zhou
et al (29) recently
reported that 6-Shogaol (a phenolic alkanone derived from ginger)
potentiates PC to gemcitabine treatment in vitro and in
vivo, and that such sensitization is achieved by suppressing
the activation signaling via toll-like receptor 4 (TLR4)/NF-κB. We
therefore reasoned that coix seed emulsion, an anticancer agent
developed from natural products, may similarly serve as a novel
chemosensitizing drug, and that its mechanism of action may also be
the abrogation of the chemotherapeutic agent-induced activation of
NF-κB, which would then inactivate multiple downstream NF-κB-driven
survival factors.
Given the robust antitumor activity of the combined
use of coix seed emulsion and chemotherapeutic agents in the
treatment of different types of cancer, we were interested in
ascertaining whether coix seed emulsion is specifically effective
in synergistically sensitizing PC cells to chemotherapeutics.
Results from a phase II clinical trial (38 patients) in the US
showed that patients treated with a combination of coix seed
emulsion and gemcitabine had a median progression-free survival
(PFS) of 114 days, which was significantly longer than the median
PFS of 57.5 days in the gemcitabine only group (P=0.008) (12). In another phase II clinical trial
(58 patients) in China, addition of coix seed emulsion to
gemcitabine treatment led to a (non-significant) trend towards
improved survival of patients with advanced PCs (13). We examined this issue further in our
in vitro and in vivo studies. The in vitro
experiments showed that treatment with coix seed emulsion alone
caused a modest loss of viable PC cells; this effect varied with
the cell type used (IC50=1.50, 1.75 and 1.80 mg/ml in
BxPC-3, PANC-1 and AsPC-1 cells, respectively); however,
pretreatment with coix seed emulsion effectively decreases the
IC50 value of gemcitabine; we believe that the lower
dose may lead to less toxic side-effects on normal cells in
vivo. The value of the CI also confirmed the synergism of coix
seed emulsion and gemcitabine combination. Additionally, our
results revealed that coix seed emulsion alone is a general inducer
of apoptosis in PC, likely due to downregulation of anti-apoptotic
Bcl-2, upregulation pro-apoptotic Bax and cleavage of PARP. This is
consistent with previous studies that the drug inhibits and
activates anti- and pro-apoptotic genes, respectively, in various
solid tumors (39,40). Furthermore, we noted that
pretreatment with coix seed emulsion elicited significantly higher
apoptosis in the PC cell lines tested. We conclude that the
synergistic effect of coix seed emulsion and gemcitabine in PC
cells may be due to enhanced gemcitabine-induced apoptosis.
Next, we addressed the molecular mechanisms whereby
coix seed emulsion sensitizes PC cells to gemcitabine. Preclinical
studies found that coix seed emulsion markedly decreased the level
of NF-κB in the nucleus in Lewis lung carcinoma (22), and inhibited protein kinase C/NF-κB
signaling in a dose-dependent manner in MDA-MB-231 breast cancer
cells (16). We thus investigated
the expression of anti-apoptotic proteins survivin and Bcl-2, and
of the proliferation protein Cox-2, both of which are regulated by
NF-κB at the transcriptional level and can be inhibited by
suppressing NF-κB (41). We also
studied the expression of the pro-apoptosis protein Bax and
survivin, a member of the IAP protein family that is constitutively
activated in PCs, and contributes to the development of resistance
to the chemotherapeutic response (42); in fact, a direct knockdown of
survivin and p65 enhanced the chemosensitivity of PC cells to
gemcitabine (30). Our
immunoblotting results were consistent with these published
findings and document that the coix seed emulsion and gemcitabine
combination significantly suppressed the level of expression of
survivin in the PC cell lines. Additionally, overexpression of
Bcl-2 in PC cells has been reported to contribute to
chemoresistance (43). Our findings
also demonstrated that the combination of the two drugs effectively
downregulated Bcl-2 expression and elevated Bax expression, which
in turn enhanced the apoptosis of the cancer cells. Moreover,
constitutively overexpressed Cox-2 has been validated as a
promising therapeutic target for reversing chemoresistance in
cancer cells (44). Our
observations reiterate this potential therapeutic benefit in that
coix seed emulsion complements the downregulation of Cox-2 by
inhibiting NF-κB, and increases the chemosensitivity of PC cells.
The most common form of NF-κB is a dimer of RelA (p65) and NF-κB 1
(p50); in an inactive state, this form is sequestered in the
cytoplasm; however, following cellular stimulation, active NF-κB
translocates into the nucleus, where it binds with specific
response elements in DNA sequences to switch on gene transcription
(45). Collectively, our
observations thus provide multiple lines of evidence that coix seed
emulsion attenuates gemcitabine-induced NF-κB activation by
abrogating gemcitabine-induced activation of NF-κB response
element, inhibiting gemcitabine associated translocation of the
Rel-A/p65 subunit to the nucleus, and reducing the DNA binding
activity of NF-κB.
Our in vivo study was performed in nude mice
with subcutaneous BxPC-3 human PC xenografts. When 12.5 ml/kg coix
seed emulsion was administered for 24 days, tumor volumes in the
treated group were considerably lower than those in the control
group (P<0.05), confirming the findings that coix seed emulsion
contributes to the inhibition of tumor growth in xenograft mouse
models that bear various types of solid tumors. Notably, coix seed
emulsion also markedly augmented the anticancer efficacy of
gemcitabine, when compared to mice treated with gemcitabine alone.
Additionally, tumor samples significantly exhibited decreased Ki-67
immunoreactivity (indicating reduced viability of cells in tumors
treated with the two drugs), as well as diminished p65
immunostaining in the combination group, supporting the finding of
decreased NF-κB transcriptional activity. The in vivo
results are thus consistent with the in vitro molecular
investigations, which clearly support the finding that coix seed
emulsion sensitizes PC cancer cells to gemcitabine treatment by
inhibiting the constitutive and gemcitabine-induced activation of
NF-κB.
In conclusion, our evidence shows that coix seed
emulsion pretreatment synergistically abrogates de novo and
acquired resistance to gemcitabine in PC cells. Studies on the
molecular mechanism underlying this finding point to suppression of
NF-κB activity and of its downstream target genes as contributing
to the synergistic effect. We recommend that coix seed emulsion be
developed as a non-toxic adjuvant to conventional chemotherapeutics
for the treatment of patients with advanced PC.
Acknowledgments
The present study was financially supported by
grants from the Natural Science Foundation of China (nos. 81202926,
81303235, 81473434 and 81373982), the Zhejiang Provincial Natural
Science Foundation of China (LQ12H29002), the Program for Zhejiang
Leading Team of S&T Innovation (2012R10044_05), the Zhejiang
Science and Technology Project (2009C33162), the Research Fund for
the Doctoral Program of Higher Education (20113322120001), the
Zhejiang Health Science and Technology Project (2011KYA119) and the
Zhejiang TCM Science and Technology Project (2008GA009). We
sincerely thank Dr Sonal Jhaveri Schneider (Dana-Farber Cancer
Institute, Boston, MA, USA) for her advice and assistance with the
writing of the manuscript.
References
|
1
|
Wolfgang CL, Herman JM, Laheru DA, Klein
AP, Erdek MA, Fishman EK and Hruban RH: Recent progress in
pancreatic cancer. CA Cancer J Clin. 63:318–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paulson AS, Tran Cao HS, Tempero MA and
Lowy AM: Therapeutic advances in pancreatic cancer.
Gastroenterology. 144:1316–1326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Conroy T, Desseigne F, Ychou M, Bouché O,
Guimbaud R, Bécouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de
la Fouchardière C, et al Groupe Tumeurs Digestives of Unicancer;
PRODIGE Intergroup: FOLFIRINOX versus gemcitabine for metastatic
pancreatic cancer. N Engl J Med. 364:1817–1825. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Von Hoff DD, Ervin T, Arena FP, Chiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Qi F, Li A, Inagaki Y, Gao J, Li J, Kokudo
N, Li XK and Tang W: Chinese herbal medicines as adjuvant treatment
during chemo- or radio-therapy for cancer. Biosci Trends.
4:297–307. 2010.
|
|
7
|
Normile D: Asian medicine. The new face of
traditional Chinese medicine. Science. 299:188–190. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li D: The anticancer drug Kang-Lai-Te
emulsion for infusion. Vestn Ross Akad Med Nauk. 9:32–37. 2005.In
Russian.
|
|
9
|
Liu X, Yang Q, Xi Y, Yu K, Wang W, Zhao X
and Kou X: Kanglaite injection combined with chemotherapy versus
chemotherapy alone in the treatment of advanced non-small cell lung
carcinoma. J Cancer Res Ther. 10(Suppl 1): S46–S51. 2014.
View Article : Google Scholar
|
|
10
|
Zhan YP, Huang XE, Cao J, Lu YY, Wu XY,
Liu J, Xu X, Xiang J and Ye LH: Clinical safety and efficacy of
Kanglaite® (Coix Seed Oil) injection combined with
chemotherapy in treating patients with gastric cancer. Asian Pac J
Cancer Prev. 13:5319–5321. 2012. View Article : Google Scholar
|
|
11
|
Fu F, Wan Y and Wu T: Mulati: Kanglaite
injection combined with hepatic arterial intervention for
unresectable hepatocellular carcinoma: A meta-analysis. J Cancer
Res Ther. 10(Suppl 1): S38–S41. 2014. View Article : Google Scholar
|
|
12
|
Tagliaferri MA, Schwartzberg LS, Chen MM,
Camacho LH, Kaplan EH, Arena FP, Bienvenu BJ, North SE, Patel H and
Li D: A phase IIb trial of coix seed injection for advanced
pancreatic cancer. J Clin Oncol. 31(Suppl): e150232013.
|
|
13
|
Sun Y, Li Y, Qin S, Ma D, Jiao SC, Yu SY,
Li J, Liu D, Song D and Li D: A multicenter randomized phase II
trial on Kanglaite Injection (KLT) plus gemcitabine hydrochloride
(GEM) versus GEM in patients with local advanced and metastatic
pancreatic cancer. J Clin Oncol. 29(Suppl): e145102011.
|
|
14
|
Jiang Y, Yuan Q, Huang A and Fang Q: The
synergistic mechanism of pemetrexed followed by Kanglaite was due
to KLT subsequently inhibiting the pemetrexed-activated MAPK
signaling pathway. Clin Lab. 61:1353–1363. 2015.PubMed/NCBI
|
|
15
|
Lu Y, Zhang BY, Jia ZX, Wu WJ and Lu ZQ:
Hepatocellular carcinoma HepG2 cell apoptosis and caspase-8 and
Bcl-2 expression induced by injectable seed extract of Coix
lacryma-jobi. Hepatobiliary Pancreat Dis Int. 10:303–307. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Woo JH, Li D, Wilsbach K, Orita H, Coulter
J, Tully E, Kwon TK, Xu S and Gabrielson E: Coix seed extract, a
commonly used treatment for cancer in China, inhibits NFkappaB and
protein kinase C signaling. Cancer Biol Ther. 6:2005–2011. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Perkins ND: The diverse and complex roles
of NF-κB subunits in cancer. Nat Rev Cancer. 12:121–132.
2012.PubMed/NCBI
|
|
18
|
Li F and Sethi G: Targeting transcription
factor NF-kappaB to overcome chemoresistance and radioresistance in
cancer therapy. Biochim Biophys Acta. 1805:167–180. 2010.PubMed/NCBI
|
|
19
|
Arlt A, Gehrz A, Müerköster S, Vorndamm J,
Kruse ML, Fölsch UR and Schäfer H: Role of NF-kappaB and Akt/PI3K
in the resistance of pancreatic carcinoma cell lines against
gemcitabine-induced cell death. Oncogene. 22:3243–3251. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qanungo S, Uys JD, Manevich Y, Distler AM,
Shaner B, Hill EG, Mieyal JJ, Lemasters JJ, Townsend DM and
Nieminen AL: N-acetyl-L-cysteine sensitizes pancreatic cancers to
gemcitabine by targeting the NFκB pathway. Biomed Pharmacother.
68:855–864. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Banerjee S, Kaseb AO, Wang Z, Kong D,
Mohammad M, Padhye S, Sarkar FH and Mohammad RM: Antitumor activity
of gemcitabine and oxaliplatin is augmented by thymoquinone in
pancreatic cancer. Cancer Res. 69:5575–5583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pan P, Wu Y, Guo ZY, Wang R, Wang YJ and
Yuan YF: Antitumor activity and immunomodulatory effects of the
intraperitoneal administration of Kanglaite in vivo in Lewis lung
carcinoma. J Ethnopharmacol. 143:680–685. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gu M, Yu Y, Gunaherath GM, Gunatilaka AA,
Li D and Sun D: Structure-activity relationship (SAR) of
withanolides to inhibit Hsp90 for its activity in pancreatic cancer
cells. Invest New Drugs. 32:68–74. 2014. View Article : Google Scholar
|
|
24
|
Banerjee S, Wang Z, Kong D and Sarkar FH:
3,3′-Diindolylmethane enhances chemosensitivity of multiple
chemotherapeutic agents in pancreatic cancer. Cancer Res.
69:5592–5600. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chung CP, Hsu CY, Lin JH, Kuo YH, Chiang W
and Lin YL: Antiproliferative lactams and spiroenone from adlay
bran in human breast cancer cell lines. J Agric Food Chem.
59:1185–1194. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oettle H, Post S, Neuhaus P, Gellert K,
Langrehr J, Ridwelski K, Schramm H, Fahlke J, Zuelke C, Burkart C,
et al: Adjuvant chemotherapy with gemcitabine vs observation in
patients undergoing curative-intent resection of pancreatic cancer:
A randomized controlled trial. JAMA. 297:267–277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Arlt A, Vorndamm J, Müerköster S, Yu H,
Schmidt WE, Fölsch UR and Schäfer H: Autocrine production of
interleukin 1beta confers constitutive nuclear factor kappaB
activity and chemoresistance in pancreatic carcinoma cell lines.
Cancer Res. 62:910–916. 2002.PubMed/NCBI
|
|
28
|
Cao LP, Song JL, Yi XP and Li YX: Double
inhibition of NF-κB and XIAP via RNAi enhances the sensitivity of
pancreatic cancer cells to gemcitabine. Oncol Rep. 29:1659–1665.
2013.PubMed/NCBI
|
|
29
|
Zhou L, Qi L, Jiang L, Zhou P, Ma J, Xu X
and Li P: Antitumor activity of gemcitabine can be potentiated in
pancreatic cancer through modulation of TLR4/NF-κB signaling by
6-shogaol. AAPS J. 16:246–257. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu J, Ma J, Wu Z, Li W, Zhang D, Han L,
Wang F, Reindl KM, Wu E and Ma Q: Arginine deiminase augments the
chemosensitivity of argininosuccinate synthetase-deficient
pancreatic cancer cells to gemcitabine via inhibition of NF-κB
signaling. BMC Cancer. 14:6862014. View Article : Google Scholar
|
|
31
|
Maier HJ, Schmidt-Strassburger U, Huber
MA, Wiedemann EM, Beug H and Wirth T: NF-kappaB promotes
epithelial-mesenchymal transition, migration and invasion of
pancreatic carcinoma cells. Cancer Lett. 295:214–228. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Neoptolemos JP, Stocken DD, Friess H,
Bassi C, Dunn JA, Hickey H, Beger H, Fernandez-Cruz L, Dervenis C,
Lacaine F, et al European Study Group for Pancreatic Cancer: A
randomized trial of chemoradiotherapy and chemotherapy after
resection of pancreatic cancer. N Engl J Med. 350:1200–1210. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stathis A and Moore MJ: Advanced
pancreatic carcinoma: Current treatment and future challenges. Nat
Rev Clin Oncol. 7:163–172. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chiorean EG and Coveler AL: Pancreatic
cancer: Optimizing treatment options, new, and emerging targeted
therapies. Drug Des Devel Ther. 9:3529–3545. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Carbone C and Melisi D: NF-κB as a target
for pancreatic cancer therapy. Expert Opin Ther Targets. 16(Suppl
2): S1–S10. 2012. View Article : Google Scholar
|
|
36
|
Wang Z, Li Y, Ahmad A, Banerjee S, Azmi
AS, Kong D and Sarkar FH: Pancreatic cancer: Understanding and
overcoming chemoresistance. Nat Rev Gastroenterol Hepatol. 8:27–33.
2011. View Article : Google Scholar
|
|
37
|
Nakanishi C and Toi M: Nuclear
factor-kappaB inhibitors as sensitizers to anticancer drugs. Nat
Rev Cancer. 5:297–309. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luqman S and Pezzuto JM: NFkappaB: A
promising target for natural products in cancer chemoprevention.
Phytother Res. 24:949–963. 2010.PubMed/NCBI
|
|
39
|
Lu Y, Li CS and Dong Q: Chinese herb
related molecules of cancer-cell-apoptosis: A minireview of
progress between Kanglaite injection and related genes. J Exp Clin
Cancer Res. 27:312008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Y, Zhang W, Wang XJ and Liu S:
Antitumor effect of Kanglaite® injection in human
pancreatic cancer xenografts. BMC Complement Altern Med.
14:2282014. View Article : Google Scholar
|
|
41
|
Hill R, Li Y, Tran LM, Dry S, Calvopina
JH, Garcia A, Kim C, Wang Y, Donahue TR, Herschman HR, et al: Cell
intrinsic role of COX-2 in pancreatic cancer development. Mol
Cancer Ther. 11:2127–2137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jiang C, Tan T, Yi XP, Shen H and Li YX:
Lentivirus-mediated shRNA targeting XIAP and survivin inhibit
SW1990 pancreatic cancer cell proliferation in vitro and in vivo.
Mol Med Rep. 4:667–674. 2011.PubMed/NCBI
|
|
43
|
Dong J, Zhao YP, Zhou L, Zhang TP and Chen
G: Bcl-2 upregulation induced by miR-21 via a direct interaction is
associated with apoptosis and chemoresistance in MIA PaCa-2
pancreatic cancer cells. Arch Med Res. 42:8–14. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ali S, Banerjee S, Schaffert JM, El-Rayes
BF, Philip PA and Sarkar FH: Concurrent inhibition of NF-kappaB,
cyclooxygenase-2, and epidermal growth factor receptor leads to
greater anti-tumor activity in pancreatic cancer. J Cell Biochem.
110:171–181. 2010.PubMed/NCBI
|
|
45
|
Li L, Aggarwal BB, Shishodia S, Abbruzzese
J and Kurzrock R: Nuclear factor-kappaB and IkappaB kinase are
constitutively active in human pancreatic cells, and their
down-regulation by curcumin (diferuloylmethane) is associated with
the suppression of proliferation and the induction of apoptosis.
Cancer. 101:2351–2362. 2004. View Article : Google Scholar : PubMed/NCBI
|