Introduction
Cholangiocarcinoma (CCA) is a highly malignant
adenocarcinoma that originates in the extrahepatic and liver bile
ducts, which terminate at the ampulla of Vater (1). CCA is generally classified into 3
forms: intrahepatic cholangiocarcinoma (ICC), hilar
cholangiocarcinoma and extrahepatic cholangiocarcinoma (ECC)
(2). Although CCA is a relatively
rare neoplasm with an annual incidence rate of 1–2 cases/100,000 in
the Western world, the overall incidence and mortality rates of
this neoplasm appear to be rising worldwide over the past several
decades (3,4). Due to its late clinical presentation
and the lack of effective non-surgical therapeutic modalities, the
disease is difficult to diagnose and is frequently fatal (5). Over the past 30 years, the overall
5-year survival rate for patients with CCA, including those who
have undergone resection, is less than 10% (6). Moreover, high drug resistance usually
lowers the efficacy of chemotherapy drugs (7). Therefore, novel effective strategies
and drugs are urgently needed to improve outcomes for patients with
CCA.
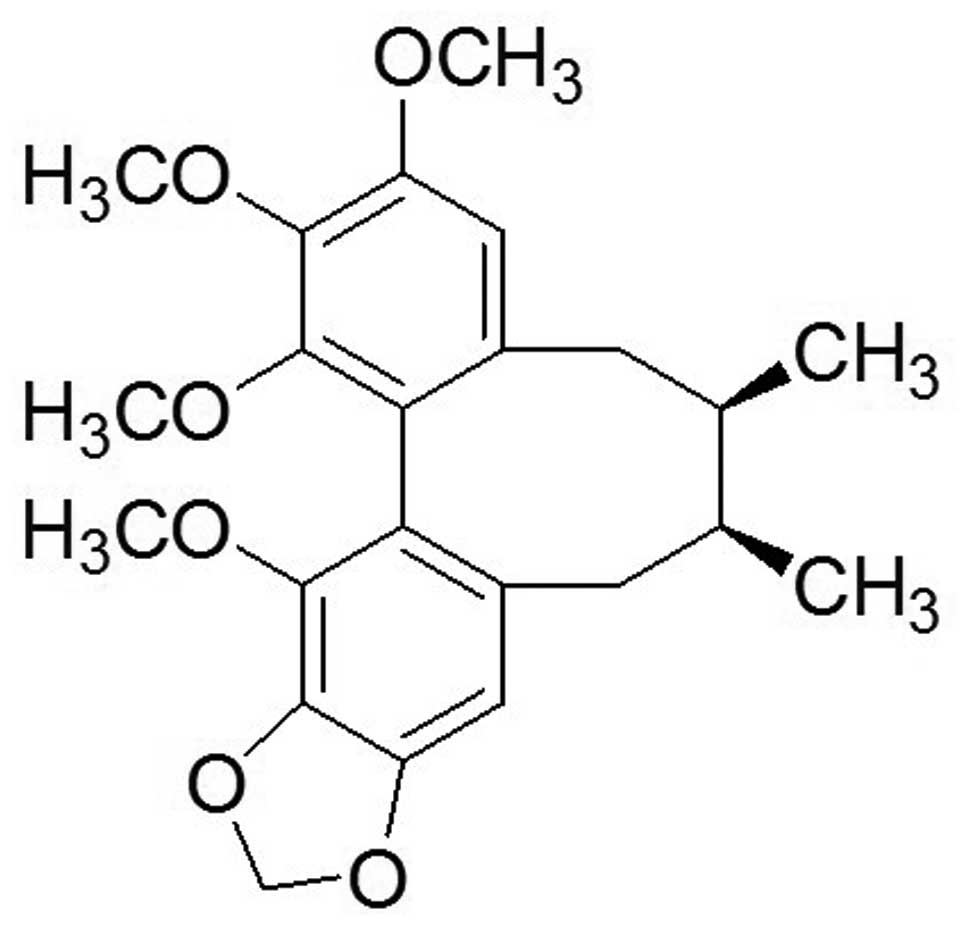
For centuries, schisandrin B (Sch B; Fig. 1) has been a major traditional
Chinese medicine. Sch B is extracted from the fruit of
Schisandra chinensis Baill., and has been used to treat
several human diseases, including hepatitis and myocardial
disorders (8). In addition, more
studies have increasingly shown that Sch B possesses antitumor
activity in various types of human cancers, including glioma,
gastric and breast cancer, and hepatoma (9–12).
Previous studies have shown that Sch B attenuates cancer invasion
and metastasis with very low toxicity (13), and it inactivates ATR when DNA
damage occurs (14). However, to
the best of our knowledge, the effects of Sch B on CCA cells and
the underlying mechanisms of these effects have not been previously
reported. In the present study, we investigated the anticancer
effects of Sch B on human CCA cell lines (HCCC-9810 and RBE) and
the possible molecular mechanisms underlying these actions, which
provided experimental evidence for the potential application of Sch
B as a new natural antitumor medicine for CCA.
Materials and methods
Cell lines and culture
The human CCA HCCC-9810 and RBE cell lines were
purchased from the Shanghai Institute of Cell Biology, Chinese
Academy of Sciences (CAS; Shanghai, China). All cells were grown in
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS)
(both from Gibco, Grand Island, NY, USA), 100 µg/ml
streptomycin and 100 U/ml penicillin (HyClone, Logan, UT, USA). The
cells were then stored in a 5% CO2 atmosphere at 37°C.
After reaching 80% confluency, the cells were digested with 0.25%
pancreatin-ethylene diamine tetraacetic acid diluted in a ratio of
1:3.
Drugs and antibodies
Sch B and Rhodamine 123 (Rho 123) were purchased
from Sigma-Aldrich (St. Louis, MO, USA). Sch B was dissolved in
dimethyl sulfoxide (DMSO; Sigma-Aldrich) to create a 100 mM stock
solution. The stock solution was further diluted with culture media
to acquire the desired concentrations. The control groups were
treated with equal volumes of DMSO. Cell Counting Kit-8 (CCK-8) was
purchased from Dojindo Laboratories (Tokyo, Japan). The Annexin
V/PI apoptosis kit was obtained from BD Biosciences (San Diego, CA,
USA). Primary antibodies against cleaved PARP, cleaved caspase-3,
CDK4, Bcl-2, Bax, cleaved caspase-9 and cyclin D1, and secondary
antibodies (goat anti-rabbit) were all purchased from Cell
Signaling Technology (Danvers, MA, USA). β-actin was purchased from
Abways Technology (Shanghai, China).
Colony formation assay
HCCC-9810 and RBE cells were suspended in RPMI-1640
medium at a final concentration of 200 cells/ml and seeded onto
6-well plates (Corning, Corning, NY, USA). Mediim containing 10%
FBS was replaced every 3 days. The cells were fixed and stained
when a signal colony contained >50 cells. Then, colony-forming
units were microscopically photographed. The results are expressed
as the average of 3 independent experiments.
Cell viability assay
Cells were diluted into single cell suspensions and
seeded in 96-well plates (1×103 cells/well) in 100
µl culture medium. Following overnight incubation, the cells
were treated with various concentrations (0, 10, 20, 40, 80 and 160
µmol/l) of Sch B. At each time point (24, 48 and 72 h), 10
µl CCK-8 solution was added to each well and incubated for 3
h in the incubator, and the absorbance was determined at 450 nm
using a microplate reader (Bio-Tek, Winooski, VT, USA). The cell
survival rate was calculated as the percentage of absorbance in
individual Sch B-treated wells vs. untreated control wells. The
results represent the average of 5 parallel samples. All
experiments were repeated at least 3 times.
Cell cycle analysis
After treatment with different concentrations of Sch
B for 48 h, HCCC-9810 and RBE cells were harvested, fixed in 70%
ethanol and stored at 4°C. On the day of analysis, the cells were
washed and incubated in staining buffer [10 mg/ml RNase and 1 mg/ml
propidium iodide (PI) (Sigma-Aldrich)] at 37°C in the dark for 30
min. Next, the samples were analyzed using flow cytometry (BD
Biosciences). Cell Quest acquisition software was used to calculate
the percentages of cells in the G0/G1, S and G2/M phases.
Mitochondrial membrane potential (ΔΨm)
assay
Rho 123 was used to measure the ΔΨm according to
previously reported methods with some modifications (15). Briefly, after treatment with Sch B
(20, 40 and 80 µmol/l) for 48 h, cells were harvested and
incubated with medium containing Rho 123 (10 µg/ml) for 30
min in a 5% CO2 atmosphere at 37°C in the dark. The
samples were then analyzed by flow cytometry. In all cases, the
samples were gated to exclude cellular debris. Each assay was
carried out in triplicate, and the results are expressed as the
mean ± SD.
Cell apoptosis assay
Apoptosis was detected using the FITC Annexin V
apoptosis detection kit (BD Biosciences) according to the
manufacturer's instructions. Briefly, cells were treated with Sch B
(0, 20, 40 and 80 µmol/l) for 48 h. The cells were then
washed and resuspended in 1X binding buffer at a density of
1×106 cells/ml. Then, the cells were incubated with 5
µl Annexin V-FITC and 5 µl PI (100 µg/ml) for
15 min in the dark at room temperature. The stained cells were
diluted with 400 µl binding buffer and immediately detected
by flow cytometry (BD Biosciences). Each sample was assayed in
duplicate, and the experiment was repeated 3 times.
Western blot analysis
HCCC-9810 and RBE cells were treated with Sch B (0,
20, 40 and 80 µmol/l) for 48 h. The cells were harvested and
lysed in RIPA buffer (Beyotime Institute of Biotechnology, Beijing,
China) supplemented with protease inhibitor (Roche Applied Science,
Indianapolis, IN, USA) at 4°C for 15 min. Then, cells were
centrifuged in a microcentrifuge at 12,000 × g for 15 min at 4°C to
collect the supernatant. The protein concentration was measured
using the bicinchoninic acid (BCA) assay kit (Beyotime). A total of
30 µg of protein/lane was subjected to SDS-polyacrylamide
gel electrophoresis (PAGE) after boiling for 5 min in 1X SDS sample
buffer, and then electrophoretically transferred to polyvinylidene
difluoride membranes (Millipore, Bedford, MA, USA). After blocking
with 5% skim milk for 1–2 h at room temperature, the membranes were
then incubated with the indicated primary antibodies against Bcl-2,
Bax, cleaved caspase-9, cleaved PARP, cyclin D1, cleaved caspase-3,
CDK4 and β-actin (1:1,000) at 4°C overnight. After washing with
Tris-buffered saline and Tween-20 (TBST) buffer for 3×5 min, the
blots were then re-probed with the secondary antibodies conjugated
to horseradish peroxidase for 1 h at room temperature. The protein
bands were detected using enhanced chemiluminescence and visualized
using a Gel Doc 2000 (Bio-Rad, Hercules, CA, USA).
In vivo tumor xenograft study
Male athymic nude mice (5–6 weeks old) were
purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai,
China). All animal experiments were performed in strict accordance
with the Institutional Animal Care and Use Committee-approved
protocol. The mice were housed in a specific pathogen-free
environment. HCCC-9810 cells in log-phase growth
(2.5×106) suspended in 100 µl phosphate-buffered
saline (PBS) were subcutaneously inoculated into the right flank of
the nude mice. Sch B treatment was started 7 days after inoculation
of the cells. The control group (n=8) was administered PBS
intraperitoneally, whereas the other two treatment groups (n=8)
were administered Sch B (20 and 80 mg/kg) intraperitoneally every 2
days for up to 30 days. On day 30, all mice were sacrificed, and
the tumors were removed and weighed.
Statistical analysis
All of the experiments were independently repeated 3
times. Numerical data are presented as the mean ± standard
deviation. Statistically significant differences between samples
were determined using the Student's t-test. P<0.05 was
considered to indicate a statistically significant result.
Results
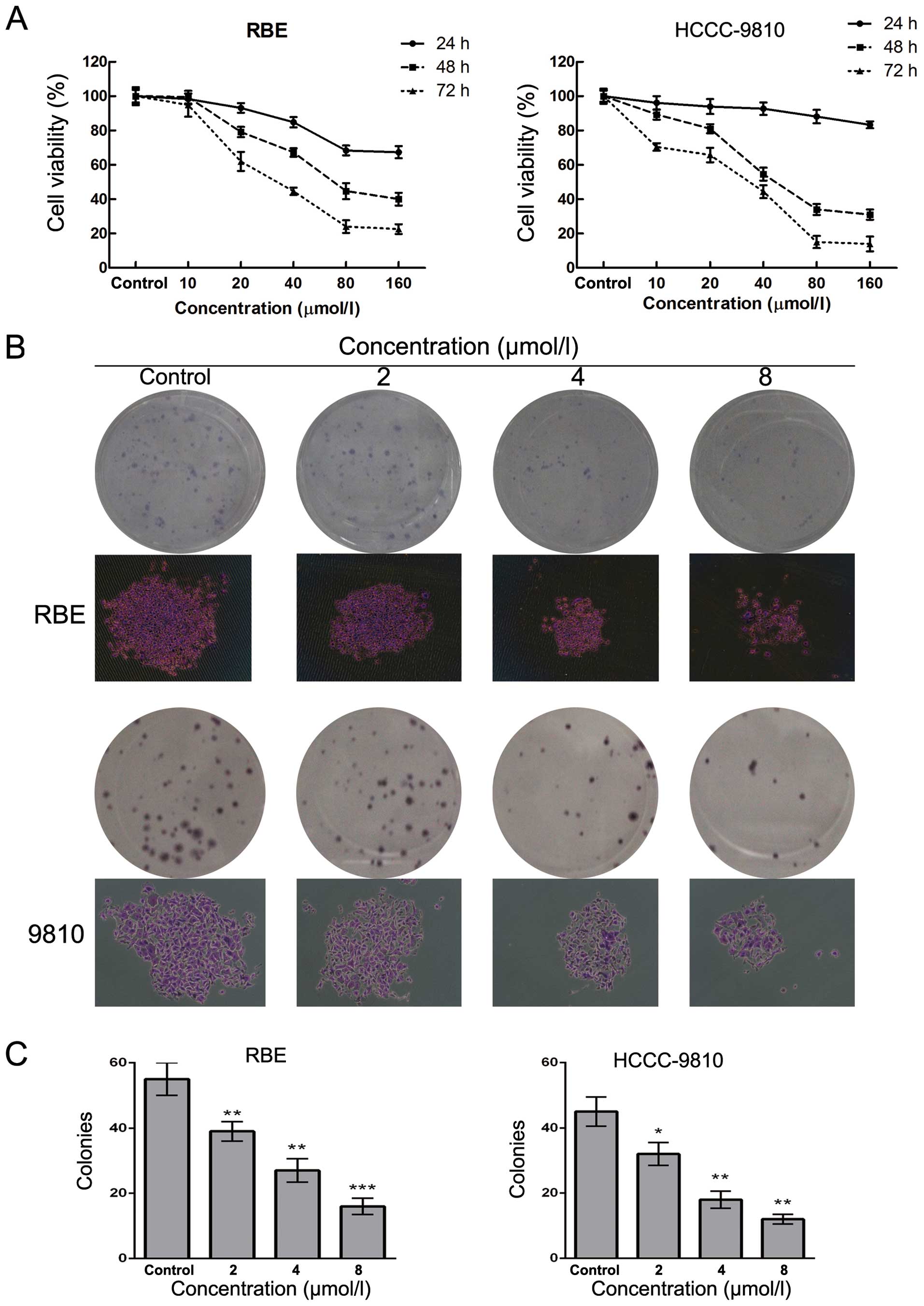
Sch B inhibits the proliferation and
decreases the cell viability of CCA cells in a dose-dependent
manner
To investigate the effect of Sch B on human CCA
cells, HCCC-9810 and RBE cells were treated with Sch B at
concentrations of 0–160 µM for 24, 48 and 72 h followed by
detection of cell viability. The results of the CCK-8 assay showed
that Sch B decreased the cell viability of the HCCC-9810 and RBE
cells in a dose- and time-dependent manner (Fig. 2A). The IC50 values (the
drug concentration that inhibited 50% of the cells) of Sch B in the
HCCC-9810 and RBE cells at 48 h were 40±1.6 and 70±2.6
µmol/l, respectively. As shown in Fig. 2B and C, the numbers of colonies of
RBE and HCC-9810 cells were significantly reduced in a
concentration-dependent manner after treatment with Sch B. The
individual colony size in Sch B-treated group was also smaller than
that of control group as shown in Fig.
2B. These results suggested that Sch B inhibited the
proliferation of CCA cells.
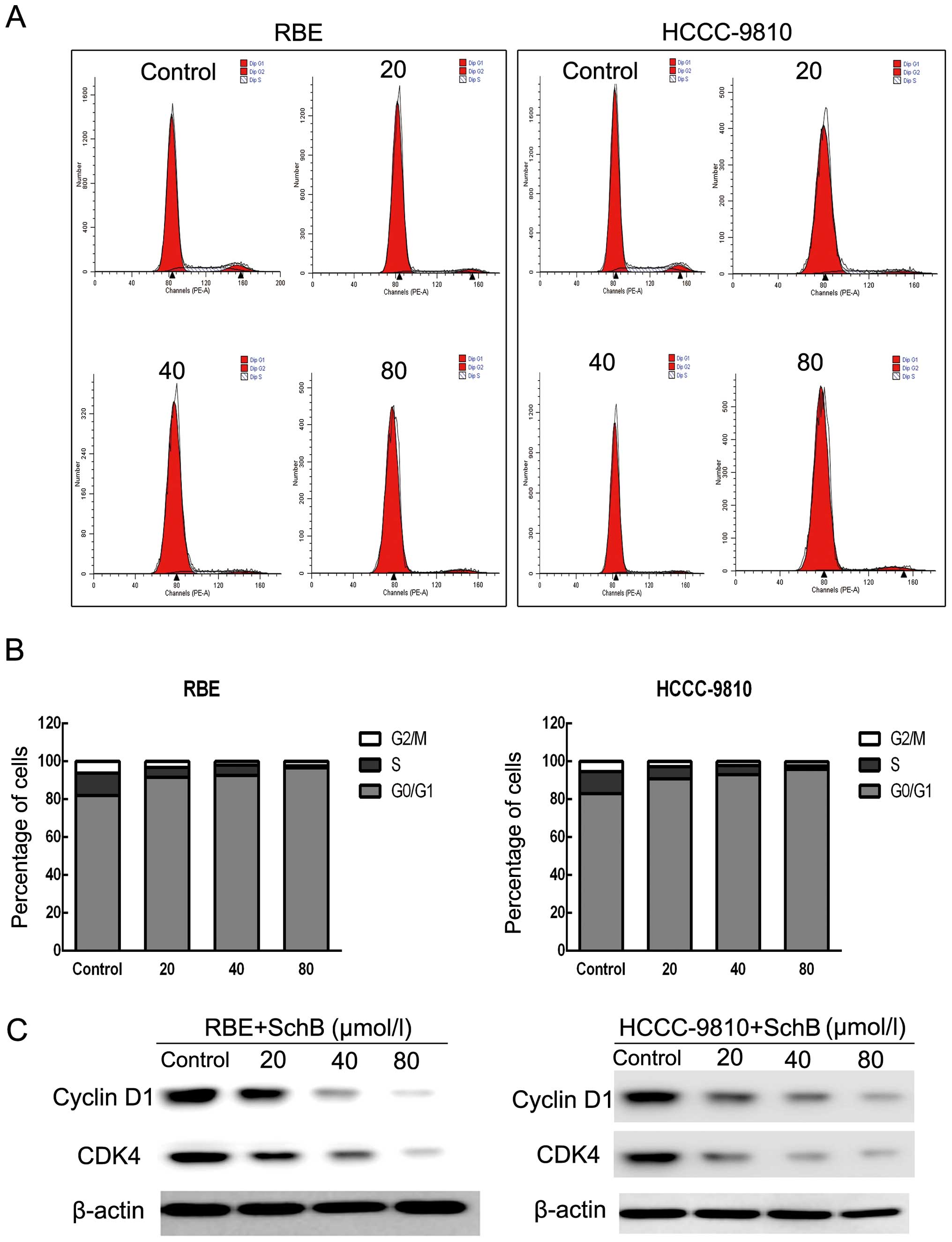
Sch B induces G0/G1 phase arrest in
HCCC-9810 and RBE cells
To determine whether Sch B-induced cytotoxicity was
caused by cell cycle arrest, we performed cell cycle distribution
analysis in the HCCC-9810 and RBE cells after exposure to Sch B. As
shown in Fig. 3A and B, Sch B
induced G0/G1 phase arrest (from 82.94 to 95.75% for HCCC-9810
cells and from 81.9 to 96.62% for RBE cells) with a slight S phase
decrease compared with that of the control cells.
Next, we assessed the protein expression of the
G0/G1 phase regulatory genes after Sch B treatment. As shown in
Fig. 3C, the expression levels of
cyclin D1 and CDK4 were reduced in a dose-dependent manner after
treatment with Sch B. These results illustrated that Sch B blocked
the cell cycle to inhibit CCA cell proliferation.
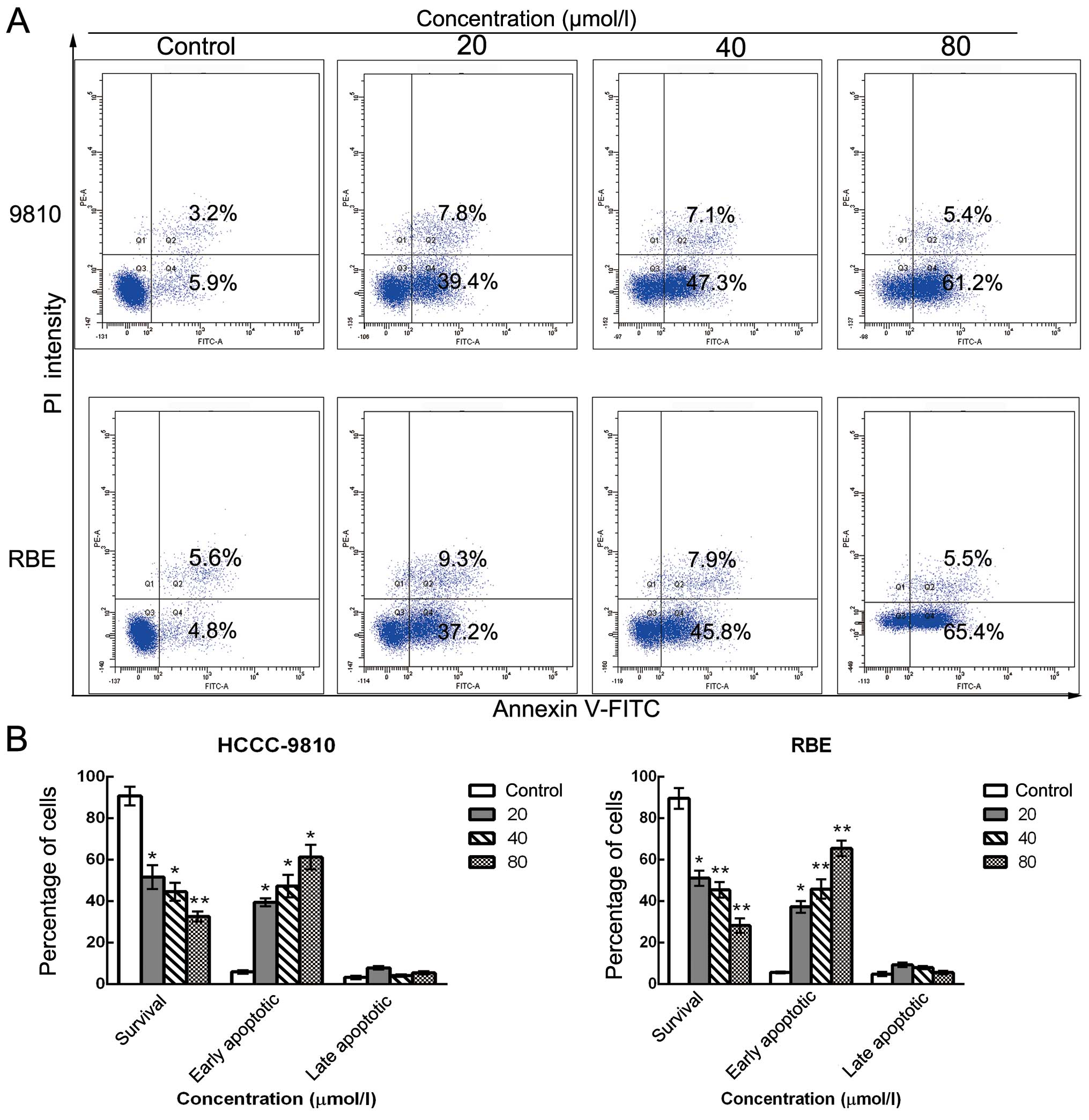
Sch B induces apoptosis in human CCA
cells
To further elucidate whether the inhibition of cell
growth was caused by apoptosis, we performed Annexin V/PI dual
staining followed by flow cytometric analysis after stimulation
with various doses of Sch B. The translocation of
phosphatidylserine (PS) from the inner leaflet to the outer leaflet
of the plasma membrane is a marker of apoptosis. Apoptotic and
necrotic cells can be identified via fluorophore-labeled Annexin V,
which has a high affinity for PS. In addition, PI nucleic acid dye
gains entry into late apoptotic and necrotic cells, but not into
early apoptotic and living cells. Therefore, various cell
populations can be easily distinguished using Annexin V/PI
staining. As assessed by flow cytometry and as shown in Fig. 4A and B, the number of surviving
cells was decreased (from 90.1 to 33.2% for HCCC-9810 cells and
from 89.4 to 28.9% for RBE cells) and the number of early apoptotic
cells was markedly increased (from 5.9 to 61.2% for HCCC-9810 cells
and from 4.8 to 65.4% for RBE cells) in a dose-dependent manner
after Sch B treatment compared with the control group. These
results are consistent with the cell viability results as
determined by the CCK-8 assay, suggesting that the apoptotic cell
death pathway may play a key role in the antiproliferative effect
of Sch B on HCCC-9810 and RBE cells.
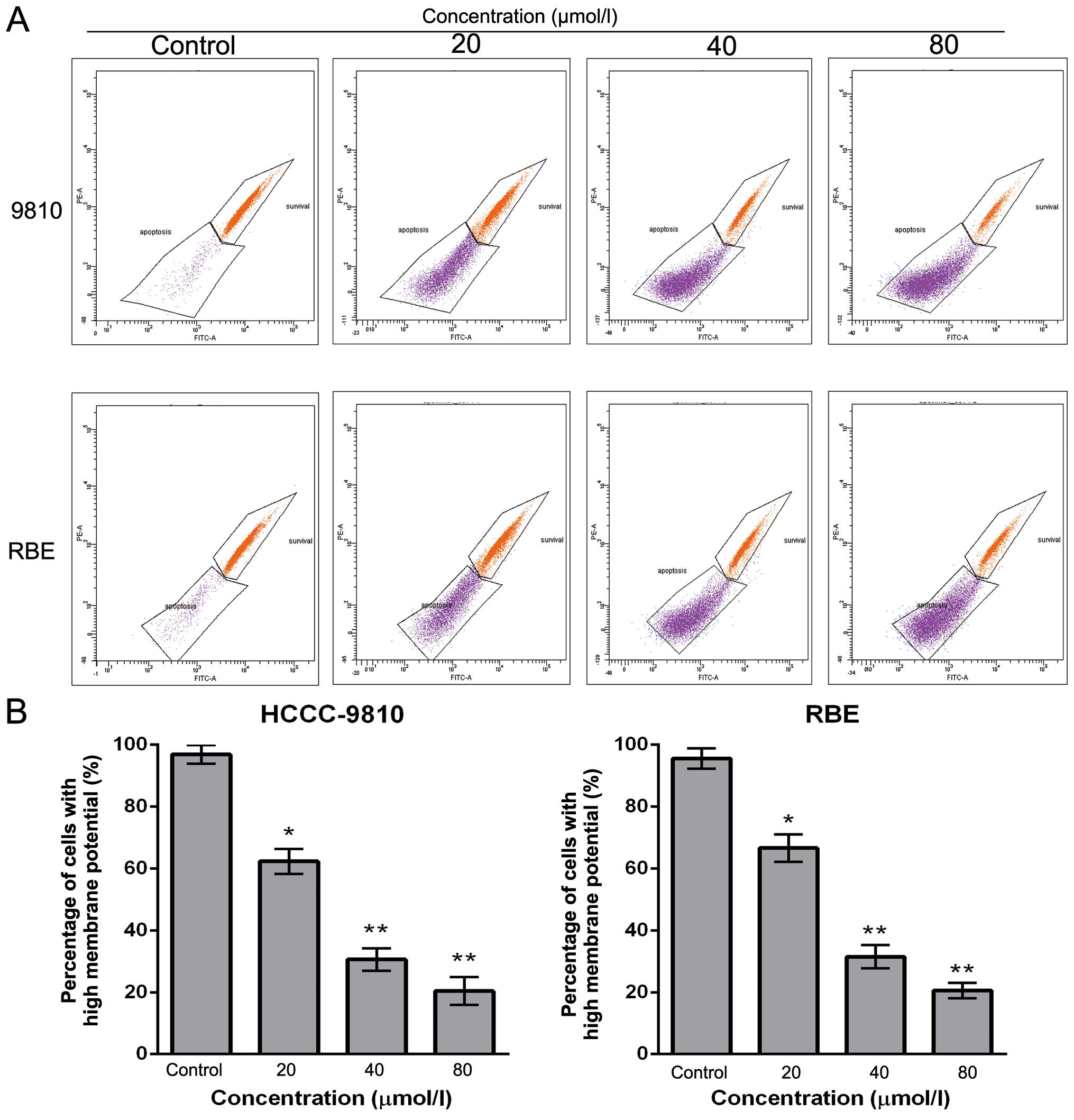
Sch B decreases ΔΨm in CCA cells
To examine whether mitochondrial membrane integrity
is disrupted by Sch B treatment, cells were stained with Rho 123,
and staining was detected by flow cytometry. The loss of the ΔΨm
was reflected by a decrease in the intensity of Rho 123 fluorescent
staining. Compared with the control cells, Sch B treatment markedly
decreased Rho 123-positive cells (from 96.9 to 18.2% for HCCC-9810
cells and from 95.6 to 20.5% for RBE cells) in a dose-dependent
manner (Fig. 5A and B). These
findings suggest that Sch B promotes the apoptosis of CCA cells
through a mitochondrial-dependent apoptotic pathway.
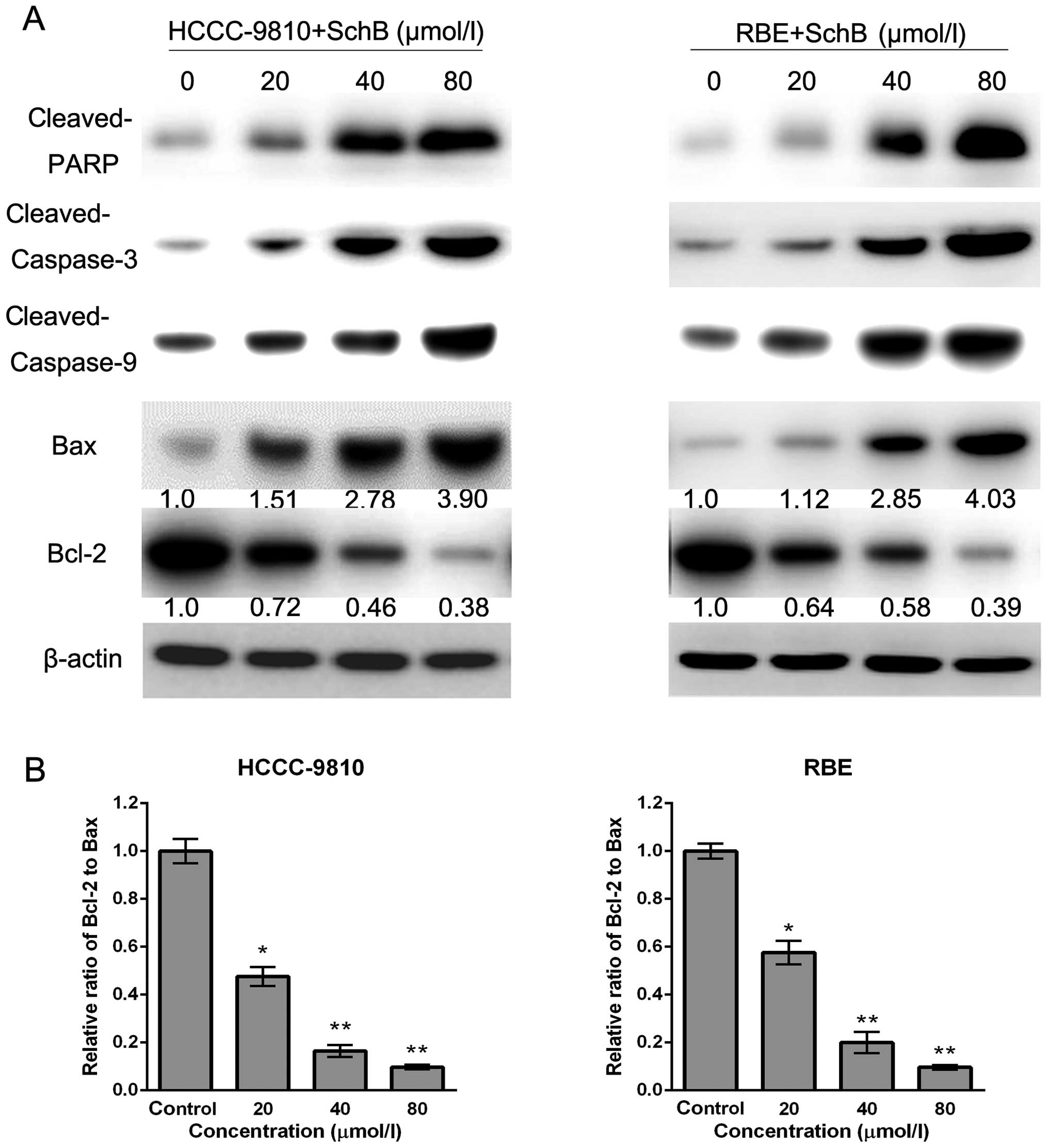
Sch B induces apoptosis via the
regulation of caspase-3 and Bcl-2 family members in CCA cells
To investigate the underlying molecular mechanisms
responsible for Sch B-induced apoptosis of HCCC-9810 and RBE cells,
we assessed the expression of apoptosis-related factors (cleaved
PARP, Bax, Bcl-2, cleaved caspase-3 and cleaved caspase-9) after
Sch B treatment by western blot analysis. As shown in Fig. 6A, increased expression of cleaved
PARP, Bax, cleaved caspase-9 and cleaved caspase-3, and decreased
expression of Bcl-2 was observed after Sch B treatment, which was
consistent with apoptosis induced by Sch B. Furthermore, compared
with the control group, the Bcl-2 (anti-apoptotic) to Bax
(pro-apoptotic) ratio was significantly decreased (Fig. 6B). Our results suggest that Sch B
promotes apoptosis through the regulation and activation of
apoptosis-related proteins in CCA cells.
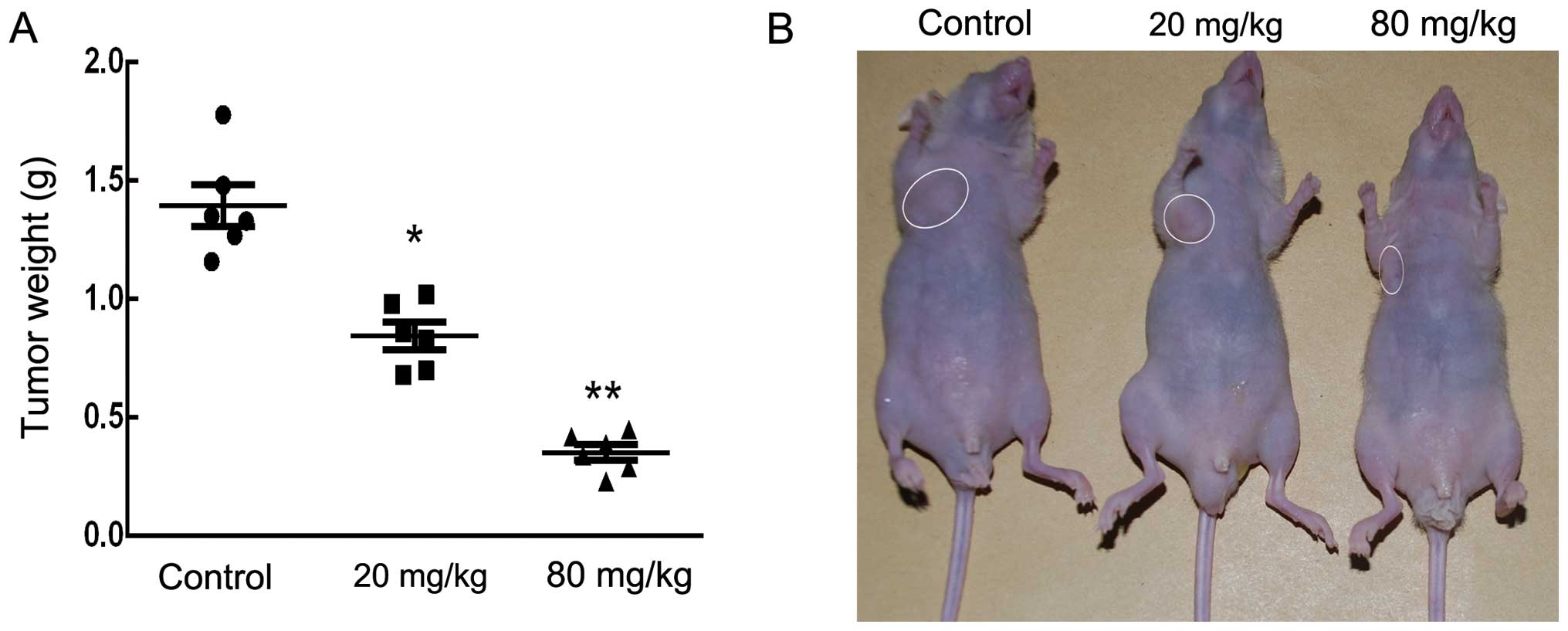
Sch B inhibits tumor growth in vivo
The anticancer effect of Sch B on tumor growth in
vivo was further analyzed by intraperitoneally injecting
vehicle (PBS) or Sch B (20 and 80 mg/kg) into nude mice bearing
subcutaneous HCCC-9810 tumor xenografts every 2 days for up to 30
days. As shown in Fig. 7A and B,
there was a marked dose-dependent reduction in tumor weight in mice
treated with oridonin compared with the control mice.
Discussion
Previous studies have demonstrated that Sch B
promoted apoptosis in a wide range of human cancers (9–12).
However, little is known concerning the antitumor activity of Sch B
on cholangiocarcinoma (CCA) cells and its underlying mechanisms. In
the present study, for the first time, we examined the inhibitory
effect and mechanism of Sch B on CCA cells. We showed that Sch B
dose-dependently decreased cell viability and induced cell cycle
arrest and apoptosis in CCA cells. Our in vivo study showed
that Sch B strongly suppressed tumor growth. Thus, Sch B may be a
promising drug for CCA prevention and treatment.
To examine the mechanism of Sch B-induced inhibition
of cell survival, we performed the cell cycle distribution
experiment in the presence of Sch B and found that it induced G0/G1
phase arrest. Further study showed that Sch B treatment resulted in
a downregulation of the expression of cyclin D1 and CDK4 in both
HCCC-9810 and RBE cells (Fig. 3C).
It is well known that the cell cycle is a tightly regulated process
consisting of 4 distinct phases: G0/G1, S, G2 and M. Activation of
each phase is dependent on the regulation of various cyclins and
cyclin-dependent kinases (Cdks) (16,17).
The complex of cyclin D1 and CDK4 is the main driver of the
transition of cells from G0/G1 to S phase by phosphorylating of
retinoblastoma (Rb) (18–20). Hyper-phosphorylated Rb activates E2F
after its dissociation from the E2F/DP1/Rb complex (21). As suggested by our cell cycle
analysis data, Sch B arrested HCCC-9810 and RBE cells at the G0/G1
phase (Fig. 3A), which may be due
to the downregulation of cyclin D1 and CDK4.
Apoptosis, genetically programmed cell death that
plays crucial roles in cell death and survival, is considered to be
one of the main contributors to cancer development. Chemical
compounds that induce cancer cell apoptosis are considered
promising anticancer drugs (22,23).
Caspases, a family of cysteine proteases, play essential roles in
apoptosis. Both the death-receptor-induced extrinsic pathway and
the mitochondria-apoptosome-mediated intrinsic pathway ultimately
activate caspases (24,25). Among them, caspase-3, which is
activated by caspase-8 or -9, is one of the most important
executioner caspases (26).
Caspase-3 cleaves several cellular proteins, including the PARP
protein, resulting in morphological changes and DNA fragmentation
that eventually lead to apoptosis (27). In the present study, we observed the
activation of caspase-9 and -3 and the cleavage of PARP following
Sch B treatment in HCCC-9810 and RBE cells, indicating that the
mitochondrial pathway was involved in Sch B-induced apoptosis.
Bcl-2 family proteins, including Bax, Bad, Bid,
Bcl-2 and Bcl-xL, play essential roles in the mitochondria-mediated
apoptosis pathway by regulating the release of cytochrome c
(28). The pro-apoptotic members
Bax and Bad induce the release of cytochrome c from the
mitochondria within cells, whereas anti-apoptotic Bcl-2 and Bcl-xL
promote cell survival by preventing cytochrome c release
from the mitochondria (29,30). A low Bcl-2 to Bax ratio can induce
ΔΨm collapse, the release of cytochrome c, and subsequent
apoptosis (31). The present study
showed that the apoptosis that occurs in response to Sch B
treatment is accompanied by decreased expression of anti-apoptotic
genes and increased expression of pro-apoptotic genes. Consistent
with this result, an obvious decrease of ΔΨm was observed in CCA
cells treated with Sch B. Taken together, these results indicate
that the mitochondrial pathway is involved in Sch B-induced
apoptosis. Sch B may be a promising drug for CCA treatment.
However, further studies are certainly needed to elucidate the
mechanism underlying Sch B-induced apoptosis.
Acknowledgments
The present study was supported by grants from the
Major Science and Technology Projects of Zhejiang Province
(2014C04008-1).
References
|
1
|
de Groen PC, Gores GJ, LaRusso NF,
Gunderson LL and Nagorney DM: Biliary tract cancers. N Engl J Med.
341:1368–1378. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tyson GL, Ilyas JA, Duan Z, Green LK,
Younes M, El-Serag HB and Davila JA: Secular trends in the
incidence of cholangiocarcinoma in the USA and the impact of
misclassification. Dig Dis Sci. 59:3103–3110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Landis SH, Murray T, Bolden S and Wingo
PA: Cancer statistics, 1998. CA Cancer J Clin. 48:6–29. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Patel T: Worldwide trends in mortality
from biliary tract malignancies. BMC Cancer. 2:102002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Khan SA, Davidson BR, Goldin R, Pereira
SP, Rosenberg WM, Taylor-Robinson SD, Thillainayagam AV, Thomas HC,
Thursz MR and Wasan H: British Society of Gastroenterology:
Guidelines for the diagnosis and treatment of cholangiocarcinoma:
Consensus document. Gut. 51(Suppl 6): VI1–VI9. 2002. View Article : Google Scholar
|
|
6
|
Zhang TC, Cao EH, Li JF, Ma W and Qin JF:
Induction of apoptosis and inhibition of human gastric cancer
MGC-803 cell growth by arsenic trioxide. Eur J Cancer.
35:1258–1263. 1999. View Article : Google Scholar
|
|
7
|
Shaib Y and El-Serag HB: The epidemiology
of cholangiocarcinoma. Semin Liver Dis. 24:115–125. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu GT: Pharmacological actions and
clinical use of fructus schizandrae. Chin Med J. 102:740–749.
1989.PubMed/NCBI
|
|
9
|
Li L, Wang T, Xu ZL, Yu Y, Chen W and Chen
F: Effects of schisandrin B on reversing multidrug resistance in
human breast cancer cells transfected with mdr1 gene. Zhonghua Yi
Xue Za Zhi. 85:1633–1637. 2005.In Chinese. PubMed/NCBI
|
|
10
|
Liu XN, Zhang CY, Jin XD, Li YZ, Zheng XZ
and Li L: Inhibitory effect of schisandrin B on gastric cancer
cells in vitro. World J Gastroenterol. 13:6506–6511. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu YF, Cao MF, Gao YP, Chen F, Wang T,
Zumbika EP and Qian KX: Down-modulation of heat shock protein 70
and up-modulation of caspase-3 during schisandrin B-induced
apoptosis in human hepatoma SMMC-7721 cells. World J Gastroenterol.
10:2944–2948. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Q, Lu XH, Wang CD, Cai L, Lu JL, Wu JS,
Zhuge QC, Zheng WM and Su ZP: Antiproliferative and
apoptosis-inducing activity of schisandrin B against human glioma
cells. Cancer Cell Int. 15:122015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Z, Zhang B, Liu K, Ding Z and Hu X:
Schisandrin B attenuates cancer invasion and metastasis via
inhibiting epithelial-mesenchymal transition. PLoS One.
7:e404802012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nishida H, Tatewaki N, Nakajima Y, Magara
T, Ko KM, Hamamori Y and Konishi T: Inhibition of ATR protein
kinase activity by schisandrin B in DNA damage response. Nucleic
Acids Res. 37:5678–5689. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu YP, Tan ZJ, Wu XS, Liu TY, Jiang L, Bao
RF, Shu YJ, Li ML, Weng H, Ding Q, et al: Triptolide induces s
phase arrest and apoptosis in gallbladder cancer cells. Molecules.
19:2612–2628. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bloom J and Cross FR: Multiple levels of
cyclin specificity in cell-cycle control. Nat Rev Mol Cell Biol.
8:149–160. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nigg EA: Cyclin-dependent protein kinases:
Key regulators of the eukaryotic cell cycle. BioEssays. 17:471–480.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Harbour JW, Luo RX, Dei Santi A, Postigo
AA and Dean DC: Cdk phosphorylation triggers sequential
intramolecular interactions that progressively block Rb functions
as cells move through G1. Cell. 98:859–869. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ezhevsky SA, Ho A, Becker-Hapak M, Davis
PK and Dowdy SF: Differential regulation of retinoblastoma tumor
suppressor protein by G1 cyclin-dependent kinase
complexes in vivo. Mol Cell Biol. 21:4773–4784. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hinds PW, Mittnacht S, Dulic V, Arnold A,
Reed SI and Weinberg RA: Regulation of retinoblastoma protein
functions by ectopic expression of human cyclins. Cell.
70:993–1006. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Massagué J: G1 cell-cycle control and
cancer. Nature. 432:298–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chinkwo KA: Sutherlandia frutescens
extracts can induce apoptosis in cultured carcinoma cells. J
Ethnopharmacol. 98:163–170. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Asgar MA, Senawong G, Sripa B and Senawong
T: Synergistic anticancer effects of cisplatin and histone
deacetylase inhibitors (SAHA and TSA) on cholangiocarcinoma cell
lines. Int J Oncol. 48:409–420. 2016.
|
|
24
|
Hu W and Kavanagh JJ: Anticancer therapy
targeting the apoptotic pathway. Lancet Oncol. 4:721–729. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Reed JC: Apoptosis-regulating proteins as
targets for drug discovery. Trends Mol Med. 7:314–319. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng L, Zheng J, Wu LJ and Zhao YY:
Julibroside J8-induced HeLa cell apoptosis through caspase pathway.
J Asian Nat Prod Res. 8:457–465. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Antonsson B: Mitochondria and the Bcl-2
family proteins in apoptosis signaling pathways. Mol Cell Biochem.
256–257:141–155. 2004. View Article : Google Scholar
|
|
28
|
Pal S, Pal PB, Das J and Sil PC:
Involvement of both intrinsic and extrinsic pathways in
hepatoprotection of arjunolic acid against cadmium induced acute
damage in vitro. Toxicology. 283:129–139. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsujimoto Y: Bcl-2 family of proteins:
Life-or-death switch in mitochondria. Biosci Rep. 22:47–58. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Das A, Banik NL, Patel SJ and Ray SK:
Dexamethasone protected human glioblastoma U87MG cells from
temozolomide induced apoptosis by maintaining Bax:Bcl-2 ratio and
preventing proteolytic activities. Mol Cancer. 3:362004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang YH, Wu YL, Tashiro S, Onodera S and
Ikejima T: Reactive oxygen species contribute to oridonin-induced
apoptosis and autophagy in human cervical carcinoma HeLa cells.
Acta Pharmacol Sin. 32:1266–1275. 2011. View Article : Google Scholar : PubMed/NCBI
|