Introduction
As the theory of tumor angiogenesis proposed by
Folkman in 1971 gained a firm foothold, studies concerning the
inhibition of tumor angiogenesis mushroomed worldwide. Angiogenesis
is the process by which new blood vessels are formed from
preexisting vasculature and is an essential step in normal tissue
growth and regeneration, immunity and nutrition (1). Based on the treatment strategy for
this theory, tumor growth and metastasis are suppressed due to the
lack of nutrients and oxygen following treatment with inhibitors of
tumor angiogenesis (2). In recent
years, several angiogenesis inhibitor agents such as bevacizumab
(monoclonal anti-VEGF antibody drug) and sunitinib
(second-generation multivariate receptor tyrosine kinase inhibitor
drug) have already 'hit' the market of antitumor drugs (3,4). As a
branch of the angiogenesis theory, integrin receptors are
attracting increased attention in the field of cancer research.
According to the theory, the expression levels of integrin
receptors in tumor tissues are relatively higher than levels in
normal tissues, which stimulate the overexpression of
angiogenesis-related proteins in downstream pathways leading to
tumor metastasis and angiogenesis.
HM-3, designed and synthesized with an (Arg-Gly-Asp)
RGD integrin ligand sequence, is a type of integrin blocker. A
preliminary study demonstrated that HM-3 significantly inhibited
the migration of endothelial cells in vitro, as well as
achieved a high inhibitory rate in SMMC-7721 (human hepatoma),
MGC-803 (human gastric) and H460 (human non-small cell lung cancer)
xenograft models (5). However,
similar to most polypeptide compounds, susceptibility to the
degradation of proteolytic enzymes and the lower stability in
vivo are two main defects associated with HM-3. Particularly in
pharmacokinetic studies in vivo, HM-3 has a relatively short
half-life (27.66±7.37 min in SD rats) (6), which bring inconvenience to its
clinical application. Therefore, prolonging the half-life of HM-3
in vivo by modification has been a significant topic in our
laboratory.
In the past several years, numerous covalent
modifications with polyethylene glycol (PEG) to HM-3 have been
carried out in our research. Four different types of PEG
[mPEG-ALD5k, mPEG-ALD10k (7), mPEG-SC10k and
mPEG-SC20k] were obtained to modify HM-3, and
mPEG-SC20k was determined to be the optimal molecule
after taking the reaction conditions (8), the antitumor activity of the modified
products and half-life in vivo into consideration. The
preliminary antitumor activity in vivo of
mPEG-SC20k-HM-3 in a human liver cancer SMMC-7721
xenograft model showed that mPEG-SC20k-HM-3 inhibited
tumor (SMMC-7721 xenograft) growth better than HM-3.
Pharmacokinetic assays in vivo indicated that the
elimination half-life in vivo of mPEG-SC20k-HM-3
subcutaneously administered was 20.13±0.64 h, which was 43.76 times
compared to HM-3. The primary target research confirmed that
integrin αvβ3 was the main target of
mPEG-SC20k-HM-3 in tumor cells (9).
To better evaluate the antitumor property of
mPEG-SC20k-HM-3, the present study aimed to
systematically investigate the antiangiogenesis and antitumor
metastasis activity in vitro, the antitumor activity in
vivo and explore the antitumor mechanism. The research provides
a good foundation for the medicinal development and application of
mPEG-SC20k-HM-3.
Materials and methods
Reagents
HM-3 [sequence: IVRRADRAAVPGGGGRGD; synthesized by
GL Biochem Ltd. (Shanghai, China)] and mPEG-SC20k-HM-3
were prepared and purified (with a purity of >98.5% as analyzed
by analytical high-performance liquid chromatography) as previously
described (8). Oxaliplatin (Lot no.
10702428) and docetaxel (Lot no. 12072811) were purchased from
Jiangsu Hengrui Medicine Co., Ltd. (Jiangsu, China) and Avastin
(Lot no. B3451) was purchased from Genentech Inc. (South San
Francisco, CA, USA), and were stored at 4°C.
Cell culture
The following human cell lines were used in the
present study: human umbilical vein endothelial cells (HUVECs)
obtained from American ScienCell Research Laboratory (Carlsbad, CA,
USA), human hepatoma cells (SMMC-7721), human gastric cancer cells
(MGC-803), and human non-small cell lung cancer cells (H460) all
purchased from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). HUVECs were maintained in endothelial cell
medium (ECM) supplemented according to the instructions of the
manufacturer. SMMC-7721, MGC-803 and H460 cells were incubated in
Dulbecco's modified Eagle's medium (DMEM) purchased from Gibco
(Grand Island, NY, USA).
HUVEC migration assay
The HUVEC migration assay was performed to
investigate the effect of mPEG-SC20k-HM-3 on antitumor
metastasis in vitro assays. Matrigel (purchased from BD
Biosciences) was coated uniformly on the surface of the Transwell
bottom after being diluted 1:2 with serum-free medium and cells
were seeded into 24-well culture plates and grown to 90%
confluency. Subsequently, the Transwell chamber and medium were
discarded after 24 h of incubation and migrated cells were stained
with 0.1% crystal violet for 10 min after being fixed with ethanol
for 30 min. Images of the cells were captured using a Zeiss Axio
Vert.A1 inverted microscope (Zeiss, Oberkochen, Germany) to
calculate the migration inhibitory rate.
Anti-angiogenesis assays in vitro
Tube formation, chick embryo chorioallantoic
membrane and the rat aortic ring assays were performed to
investigate the effect of mPEG-SC20k-HM-3 on
anti-angiogenesis in vitro. In the tube formation assay,
HUVECs were seeded on a 96-well plate coated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). Concerning the chick embryo
chorioallantoic membrane assay, 21 SPF chick embryos were purchased
and stored in an aseptic incubator at 37°C with 70% humidity after
disinfection with iodine tincture and 75% ethanol. A 1×1 cm window
was opened on the top of the embryo on the following day and sealed
with tapes after the upper air chamber membrane was turned on, with
subsequent incubation for 24 h. A piece of neutral filter paper of
1 cm diameter was settled in the embryo as a sample carrier on the
third day. Stationary liquid (identical volume of formaldehyde and
acetone mixed) was added into the chick chamber after
administration. With regard to the rat aortic ring assay,
extravascular fibrous and fatty tissues of the artery obtained from
rats were removed after being cleaned with phosphate-buffered
saline (PBS) containing high concentrations of antibiotics.
Condensing Matrigel was dropped into 96-well culture plates to
immerse the aortic rings. Samples, dissolved with DMEM, were dosed
into the ring when the Matrigel was curdled. Images were captured
by a Zeiss Axio Vert.A1 inverted microscope.
Xenograft experiments
Animal experiments were authorized by the Ethics
Committee of China Pharmaceutical University and carried out
according to the Guidelines for the Care and Use of Laboratory
Animals of China Pharmaceutical University and according to
principles of laboratory animal care and protection. Seventy-two
five-week-old SPF female BALB/c nude mice were purchased from SLRC
Animals Laboratory Co., Ltd. (Shanghai, China). Before the
experiment, eight animals/cage were kept in a pathogen-free
environment and every procedure was carried out in a laminar
airflow cabinet. Human non-small cell lung cancer H460 cells were
selected to construct the xenograft model. Cells (1×106)
were subcutaneously injected into the right flank of each mouse.
When tumor nodules reached a mean size of 80 mm3, the
animals were treated with 10 mg sample/kg body weight using
different modes of administration (dosing regimen is shown in
Table I). Animals in the negative
control group subcutaneously received the NS solvent. The
experiment was terminated after 23 days from transplantation by
cervical dislocation. The following parameters were evaluated:
daily body weight, tumor growth (assessed daily by caliper
measurement), and final tumor weight. Tumor volume was calculated
using the formula: length × width2/2.
 | Table IGroups and administration in the H460
xenograft models. |
Table I
Groups and administration in the H460
xenograft models.
| Groups | Drugs | Days, dosage and
administration |
|---|
| 1 | Control (NS) | 0–23 days, 0.2 ml,
subcutaneous |
| 2 | PEG | 0–23 days, 67.4
mg/kg/2 days, subcutaneous, 0.2 ml |
| 3 | Avastin | 0–23 days, 5 mg/kg,
tail vein injection, 0.2 ml |
| 4 |
mPEG-SC20k-HM-3 | 0–23 days, 73.4
mg/kg/2 days, subcutaneous, 0.2 ml |
| 5 |
mPEG-SC20k-HM-3 | 0–23 days, 36.7
mg/kg/2 days, subcutaneous, 0.2 ml |
| 6 |
mPEG-SC20k-HM-3 | 0–23 days, 18.35
mg/kg/2 days, subcutaneous, 0.2 ml |
| 7 |
mPEG-SC20k-HM-3 | 0–23 days, 36.7
mg/kg/2 days, tail vein injection, 0.2 ml |
| 8 | HM-3 | 0–23 days, 3
mg/kg/days, tail vein injection, 0.2 ml |
Mechanistic study
With regard to the mechanism of
mPEG-SC20k-HM-3 involved in its antitumor activity, we
performed two assays at different levels, including the genetic and
the protein levels. Originally, Tumor Metastasis PCR array,
Angiogenesis PCR array and Human Cancer PathwayFinder PCR array
(SABiosciences: http://www.sabiosciences.com) were performed to
identify key genes that were upregulated or downregulated after
incubation with effective dose of mPEG-SC20k-HM-3, by
which a possible mechanistic pathway could be predicted. All PCR
array assays were operated by Kangchen Biological Engineering Co.,
Ltd. (Shanghai, China) and negative HUVECs and testing HUVECs were
provided by our laboratory. Based on these findings, western blot
assay was performed to verify the expression variation of key
proteins in the pathways previously predicted. HUVECs were seeded
in 6-well culture plates and treated with different doses of
mPEG-SC20k-HM-3 for 24 h, and resuspended in 100
μl of mammalian protein extraction reagent (Shanghai Generay
Biotech Co. Ltd., Shanghai, China). Concentrated protein was
separated on 12% SDS-polyacrylamide gel (SDS-PAGE) for each
protein, and then the gel was transferred to a polyvinylidene
fluoride (PVDF) membrane (Millipore, Bedford, MA, USA). Briefly,
the PVDF membrane was incubated with a 1:1,000 dilution of VEGF,
Akt1, p-Akt1, ERK1/2, p-ERK1/2, MEK1, p-MEK1, integrin
αv, β3, α5, β1 and
β-actin antibodies (Cell Signaling Technology, Danvers, MA, USA).
Following incubation with a 1:2,000 dilution of secondary
antibodies, the blots were incubated with ECL reagents (Beyotime,
Jiangsu, China) and exposed to Tanon 5200-Multi to detect protein
expression. However, the HUVECs were first treated with
αvβ3 antibodies and then incubated with
mPEG-SC20k-HM-3 to detect the expression variation of
VEGF.
Statistical analysis
Statistical analysis was carried out using the
Student's t-test to evaluate the significance of differences
between different groups. P<0.05 was considered significant for
all tests.
Results
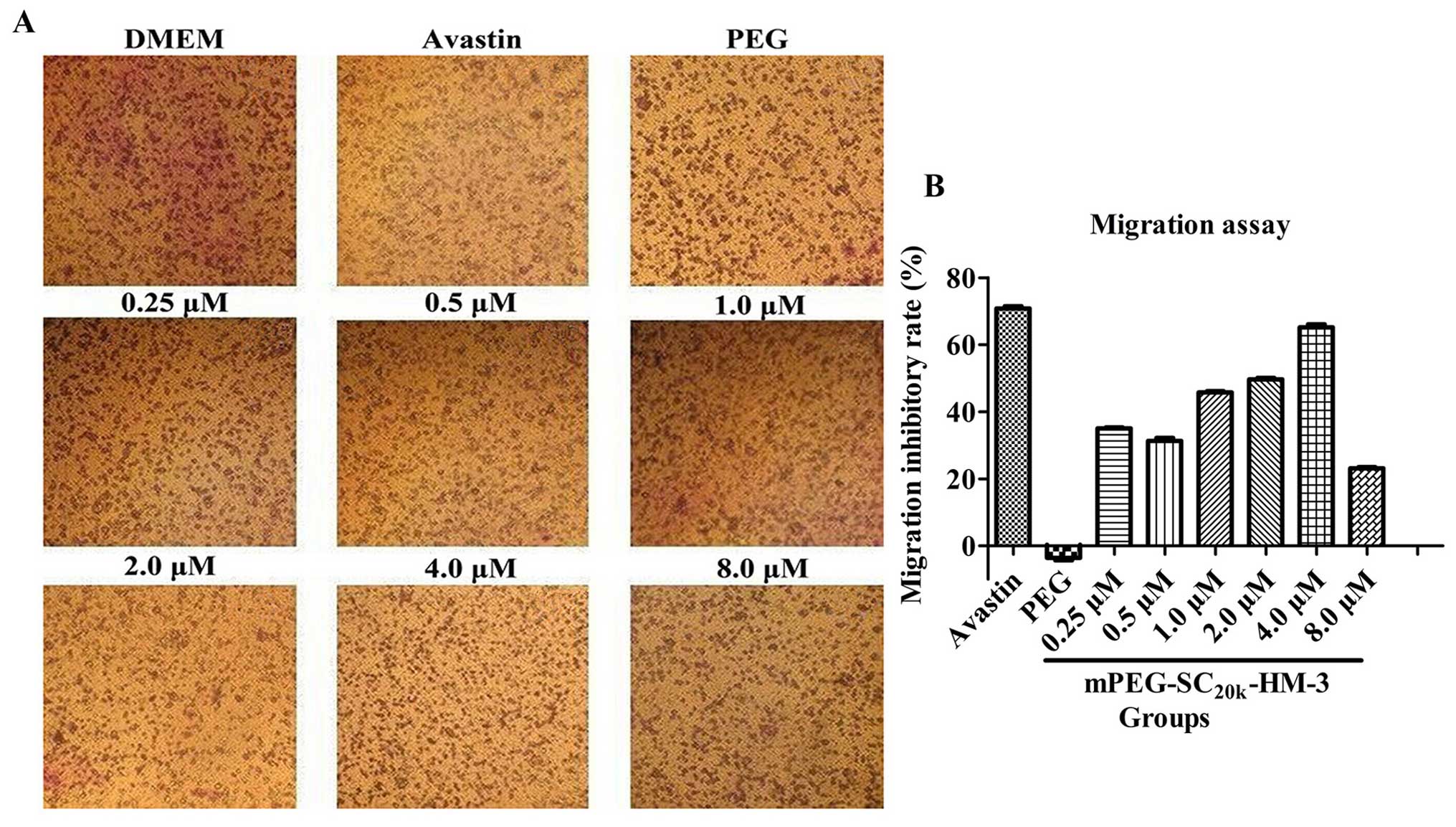
HUVEC migration assay
As for the antitumor metastasis activity in
vitro of mPEG-SC20k-HM-3, HUVEC migration assay was
performed. Compared with the negative control,
mPEG-SC20k-HM-3 exhibited an obvious inhibitory effect
on HUVEC migration in a dose-dependent manner at concentrations
ranging from 2.0 to 4.0 μM and from 4.0 to 10.0 μM,
as shown in Fig. 1, in which 4.0
μM (65.80%) was the optimal concentration for
mPEG-SC20k-HM-3 to inhibit the migration of HUVECs
compared with 10.0 mg/kg Avastin (70.85%). The above results
indicated that effective doses of mPEG-SC20k-HM-3
significantly inhibited HUVEC migration in vitro.
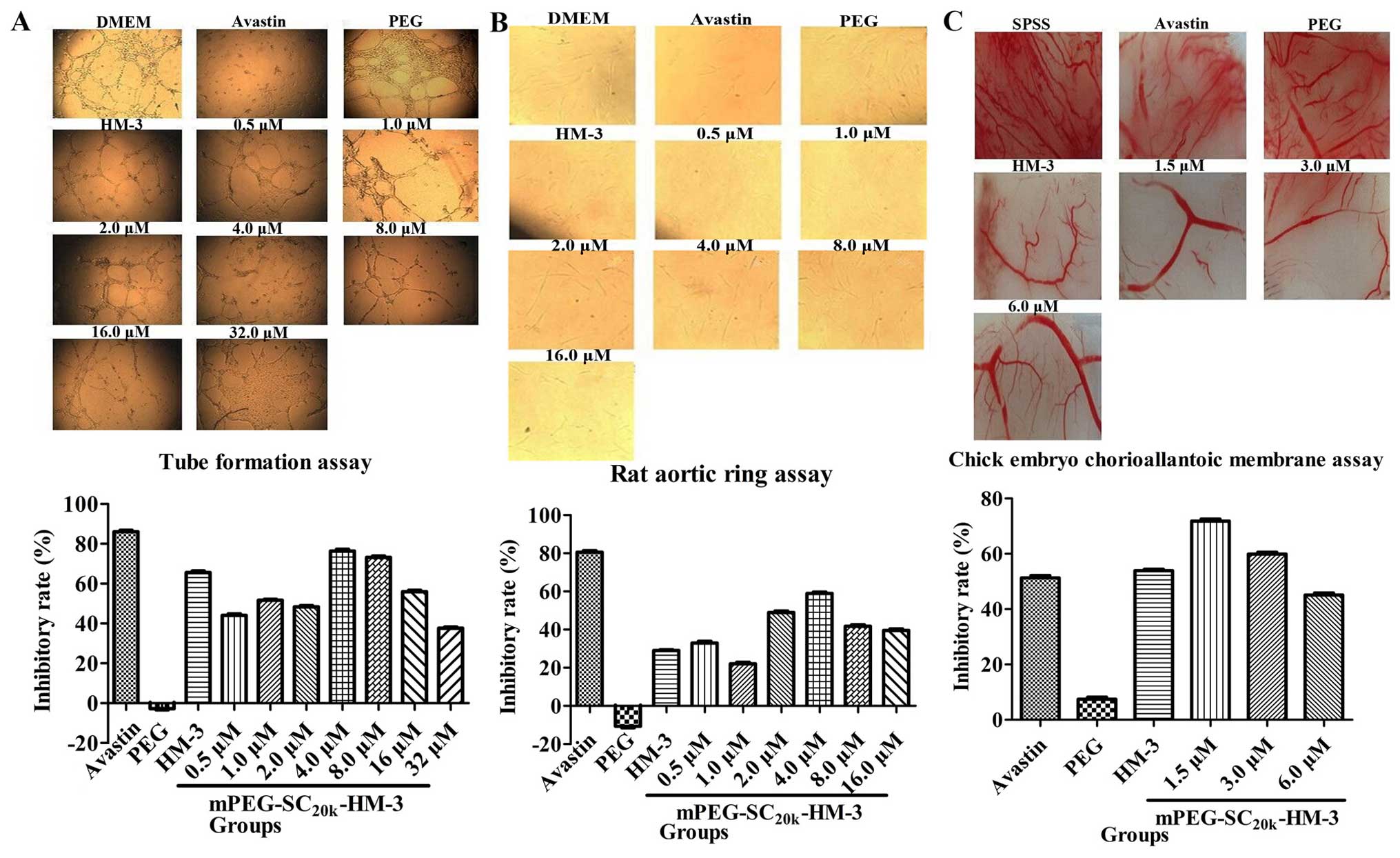
Anti-angiogenesis assays in vitro
Three independent assays were carried out to study
the anti-angiogenesis activity of mPEGSC20k-HM-3: tube
formation, rat aortic ring and chick embryo chorioallantoic
membrane assays. In the tube formation and rat aortic ring assays,
the optimum inhibitory concentration was 4.0 μM and the
inhibitory rates were 76.34% (Fig.
2A) and 58.9% (Fig. 2B),
compared with 86.02 and 80.6% for 10 mg/kg Avastin, respectively.
As for the chick embryo chorioallantoic membrane assay, the
angiogenesis inhibitory rate reached the highest level (71.8%) when
the concentration of mPEG-SC20k-HM-3 was 1.5 μM
(Fig. 2C), compared with 51.3% for
10 mg/kg Avastin. The assays in vitro above indicated that
effective doses of mPEG-SC20k-HM-3 significantly
inhibited angiogenesis.
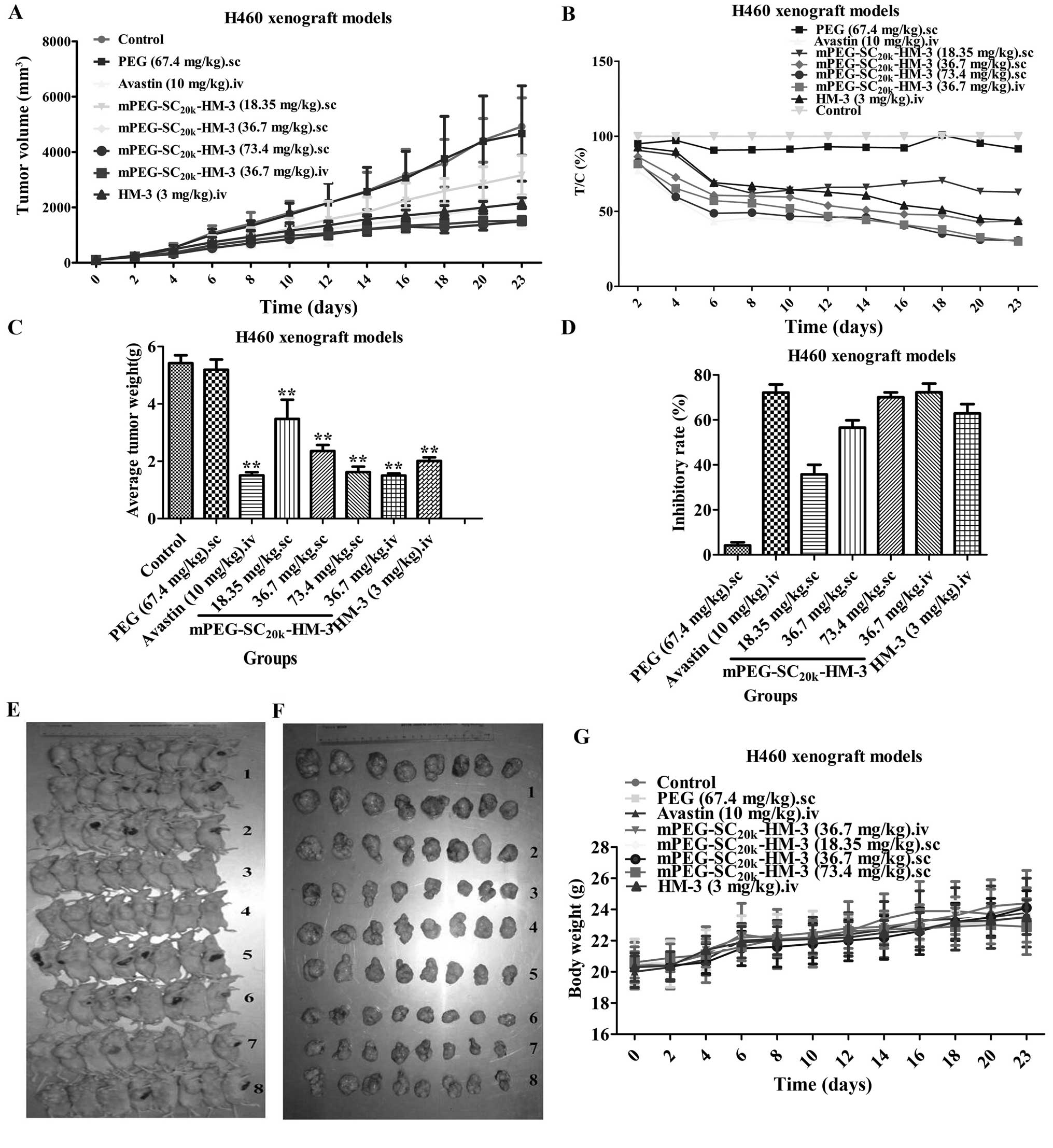
Xenograft experiments
The antitumor activities in vivo of
mPEG-SC20k-HM-3 were validated in a human non-small cell
lung cancer (H460) xenograft model. The results (Fig. 3) indicated that the T/Cs of each
group of mPEG-SC20k-HM-3 against NCI-H460 nude
xenografts were 62.7% (18.35 mg/kg, sc), 43.8% (36.7 mg/kg, sc),
30.6% (73.4 mg/kg, sc) and 30.0% (36.7 mg/kg, iv). The inhibitory
rates were 35.8, 56.6, 70.1 and 72.3%, respectively. The T/Cs of
HM-3 (3 mg/kg, iv), Avastin (10 mg/kg, iv) and PEG (67.4 mg/kg, sc)
were 43.7, 29.2 and 91.6%, while the inhibitory rates were 62.9,
72.2 and 4.2%, respectively. To assess the effects of the
treatments on toxicity, body weight and animal behavior were
monitored throughout the study. None of the
mPEG-SC20k-HM-3 groups was associated with any apparent
signs of toxicity such as reduced food and fluid consumption,
fatigue (data not shown) or body weight alterations (Fig. 3). In contrast, the weight of the
mice increased evidently after injection of different dose of
mPEG-SC20k-HM-3 during the 23 days of administration.
Taken together, the data obtained from the in vivo
experiments further confirmed the potential of
mPEG-SC20k-HM-3 administered subcutaneously against
human non-small cell lung cancer with a high inhibitory rate.
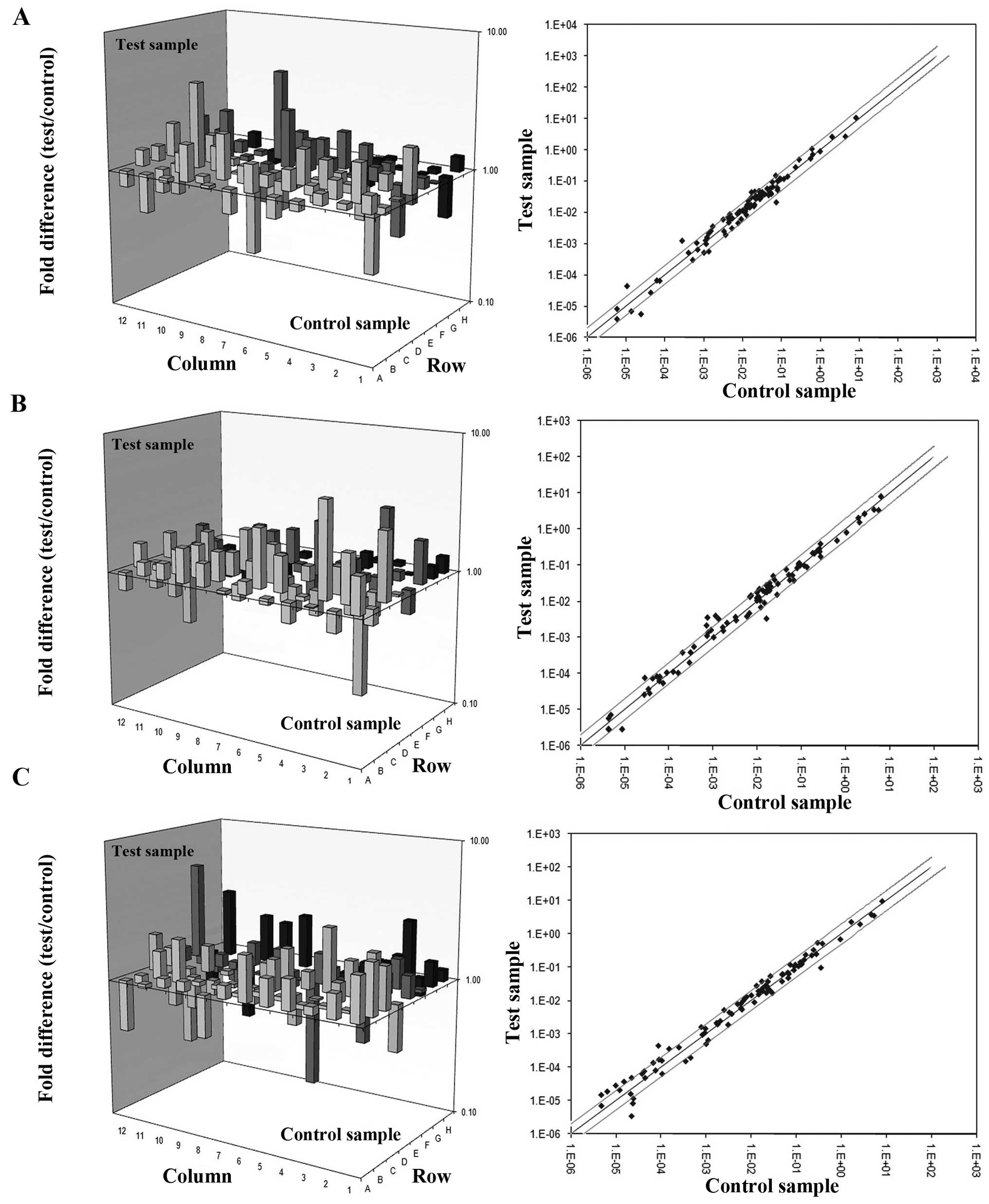
Mechanistic study
To better understand the antitumor property of
mPEG-SC20k-HM-3, the antitumor mechanism of
mPEG-SC20k-HM-3 was determined at the gene level
(RT2 Profiler PCR assays) and protein level (western
blot assays). According to the results of three PCR arrays
(Fig. 4A–C), 12 genes involved in
the three PCR arrays were downregulated in the test group when
compared with the negative group, and 27 genes were upregulated. We
classified these upregulated or downregulated genes into groups
with Gene ontology (GO) analysis system (Table II), and found that pathways of
angiogenesis and tumor metastasis were involved. The previous study
indicated that the main target of mPEG-SC20k-HM-3 in
cells was integrin αvβ3. Based on the results
of RT2 Profiler PCR assays and references on the
integrin-related pathways together, expression divergences of
various key proteins of the ERK and Akt signaling pathways were
detected by western blot assay before and after cells were
incubated with mPEG-SC20k-HM-3. The results shown in
Fig. 5 revealed that the expression
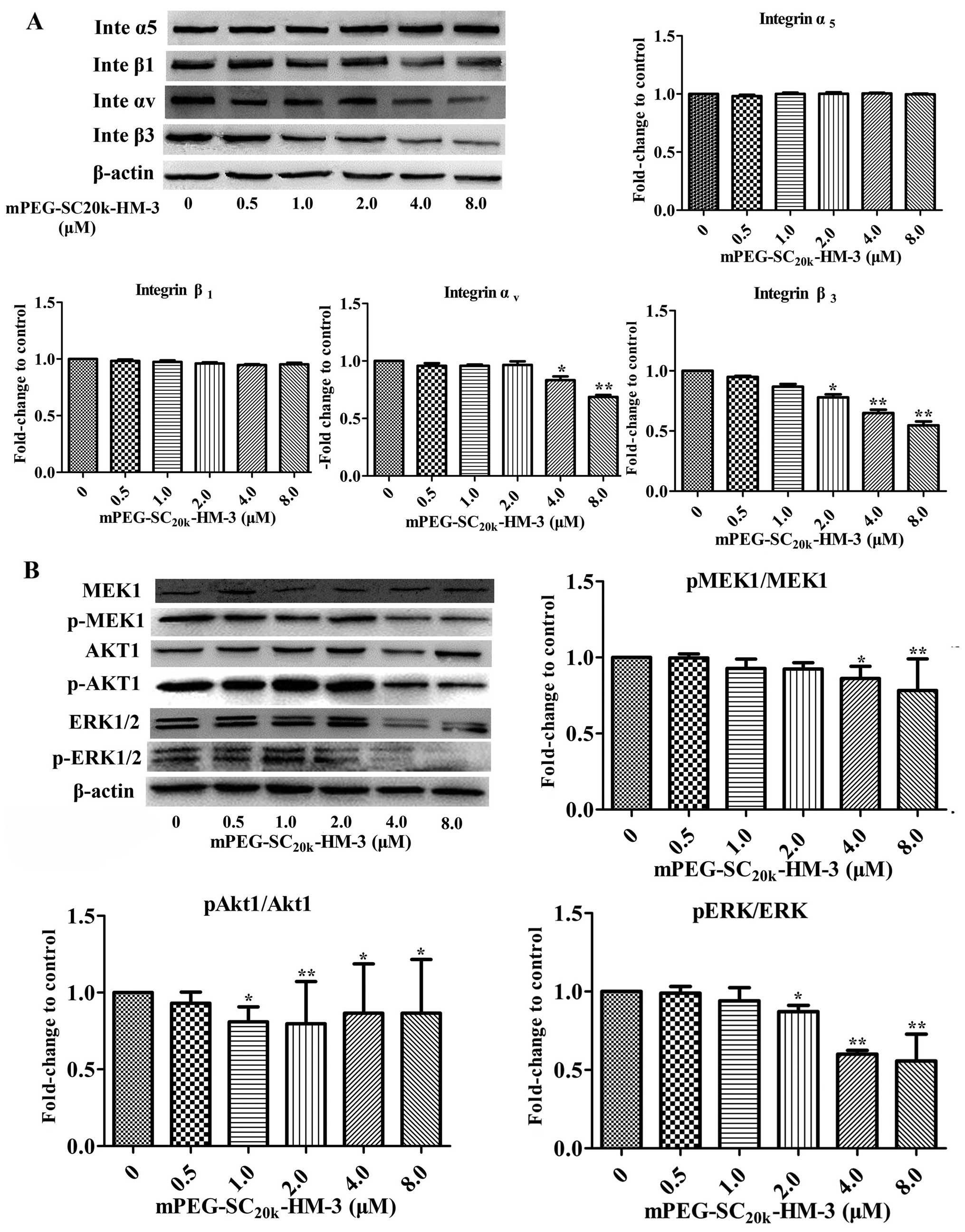
levels of Akt1, p-Akt1, ERK1/2, p-ERK1/2, MEK1, p-MEK1, integrin
αv and integrin β3 were reduced when the
concentration of mPEG-SC20k-HM-3 was 4.0 μM. For
integrin α5 and integrin β1, the expression
levels did not change as the concentration of
mPEG-SC20k-HM-3 changed. With regard to the impact of
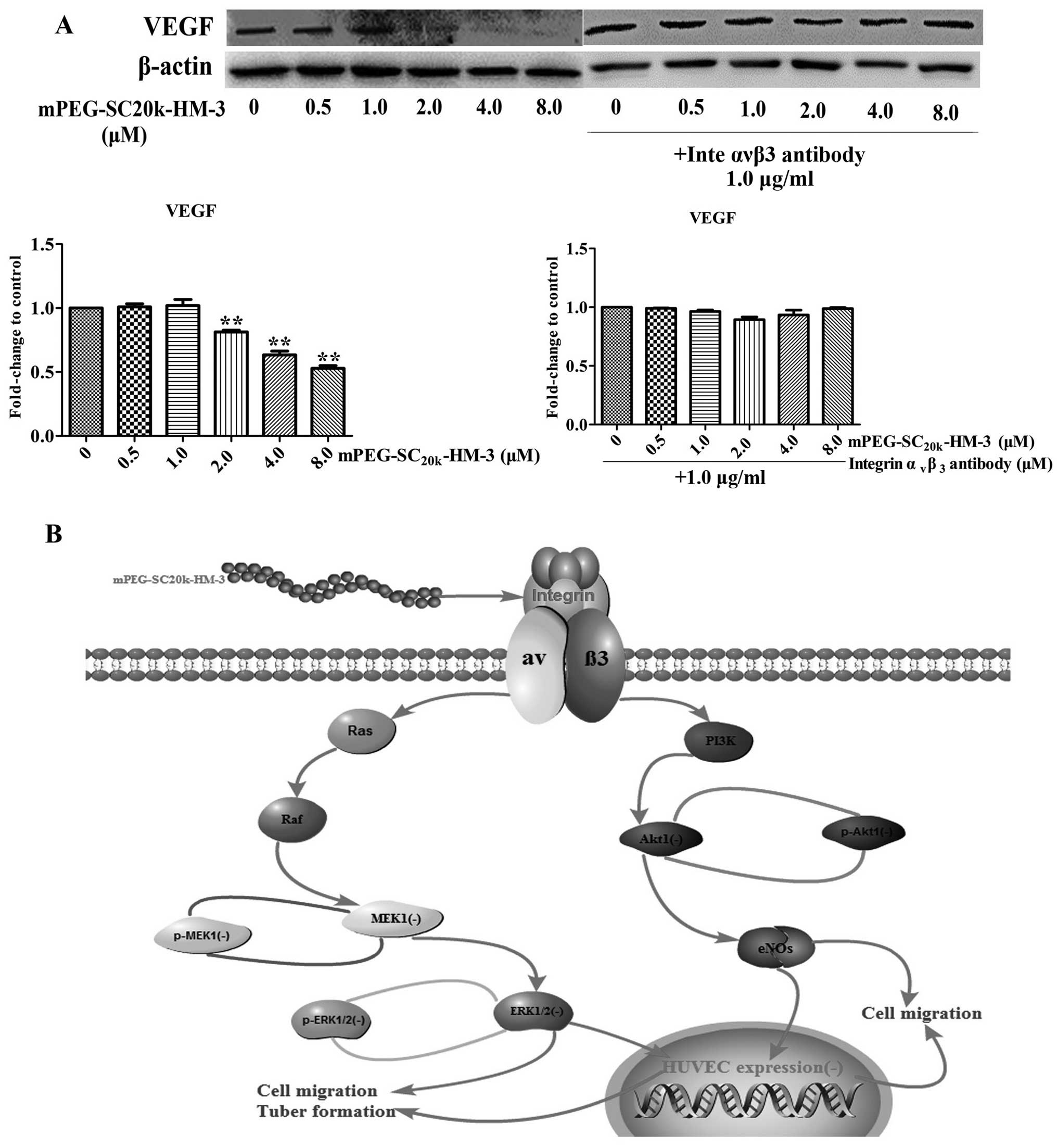
mPEG-SC20k-HM-3 on the expression of VEGF, we carried
out two independent assays. The expression of VEGF was reduced when
the concentration of mPEG-SC20k-HM-3 reached 2.0
μM; while the expression of VEGF did not change as the
concentration of mPEG-SC20k-HM-3 changed when the
antibody of integrin αvβ3 was administered
before mPEG-SC20k-HM-3 (Fig.
6A). In summary, mPEG-SC20k-HM-3 can target integrin
αvβ3 to reduce the expression of key proteins
in the ERK and Akt signaling pathways, by which tumor metastasis
and angiogenesis are inhibited and the expression of VEGF is
downregulated.
| Table IIGene ontology analysis of the
downregulated and upregulated genes identified from the PCR
arrays. |
Table II
Gene ontology analysis of the
downregulated and upregulated genes identified from the PCR
arrays.
| Categories | Downregulated | Upregulated |
|---|
Angiogenesis
Apoptosis | LECT1 | CXCL1, CXCL10,
CXCL9, ERBB2, ITGB3, MMP14, PLAU, FASLG, TIMP4, TNFSF10 |
| Cell cycle | | CDC20, MKI67,
TP53 |
| Cellular
senescence | IGFBP5 | |
| DNA damage and
repair | DDIT3 | |
| Metabolism | LPL | PFKL, SNAI2 |
| Angiogenic
factors | F3 | NOS3 |
| Cell adhesion | CDH6, CST7I,
TGA7 | CDH1, APC,
ITGB8 |
| ECM | MMP10, MMP3 | |
| Cell
proliferation | RORB, TRPM1 | CTBP1, FGFR4,
KISS1R, SRC, SYK, TSHR |
| Transcription
factors and regulators | | ETV4 |
Discussion
The aim of the present study was to investigate the
effects of mPEG-SC20k-HM-3, a modified product of the
antitumor peptide HM-3, which was found to overcome the short
half-life and high immunogenicity of HM-3.
mPEG-SC20k-HM-3 can be developed into antitumor drugs
with promising prospects. Preliminary research was performed in
regards to the reaction conditions of the modification, the
pharmacokinetics and pharmacodynamics (SMMC-7721 xenograft model)
of mPEG-SC20k-HM-3. However, systematic research on the
antitumor metastasis and anti-angiogenesis activities in
vitro, antitumor activity in vivo and a thorough
exploration of the antitumor mechanism of
mPEG-SC20k-HM-3 were not yet carried out, which were the
main goals of the present study.
In the pharmacodynamic trials in vitro of
mPEG-SC20k-HM-3, apart from tumor metastasis and
angiogenesis-related assays, MTT cytotoxicity, cell cycle and
apoptosis assays (data not shown) were also performed. According to
the results, mPEG-SC20k-HM-3 did not show any
cytotoxicity, cell cycle arrest or cell apoptosis inhibition. In
the xenograft pharmacodynamical experiments, however in the H460
xenograft model in the present study, mPEG-SC20k-HM-3
also showed a more marked inhibitory effect than HM-3 in hepatic
carcinoma (SMMC-7721), gastric (MGC-803) and colon cancer (HCT-116)
nude xenograft models (data not shown). A previous study indicated
that the mouse survival rate of the mPEG-SC20k-HM-3
(90.0 mg/kg) group was 90% after 15 days of therapy in a DBA/2
mouse lymphatic metastasis model (P388D1), compared with a 50%
survival rate of 3.0 mg/kg HM-3 (data not shown). In addition, the
number of surviving mice and body weight data after therapy suggest
that the modified peptide did not have any cytotoxicity compared
with conventional chemical drugs, which are consistent with the
results of the pharmacodynamic trials in vitro. For
combination therapy consideration, the pharmacodynamic trails in
vivo (data not shown) of the modified peptide combined with
other antitumor chemical drugs (docetaxel, oxaliplatin and
capecitabine) have already been launched and we obtained
considerable inhibitory rate data with these combination
therapies.
All the results above indicated that the
mPEG-SC20k-HM-3 has enhanced antitumor activities and
further study should be carried out to elucidate the mechanism of
the pharmacodynamical effects.
Since Judah Folkman proposed the theory of tumor
angiogenesis in 1971, tumor angiogenesis has been investigated
during the past 30 years and many achievements and new ideas based
on this theory have emerged. Integrins and their receptors are
derivatives of this theory and they have already been studied for
many years. Among all the receptors of the integrin family,
αvβ3 is considered as one of the most
significant factors with regard to its significance to tumor
angiogenesis (10,11) and it is the main target focused on
by current antitumor drug designers. Its expression in endothelial
cells in dormancy and other normal tissues is relatively low
compared with its expression in many types of cancer and
endothelial cells in tumor angiogenesis, which makes integrin
αvβ3 special and a hotspot in terms of tumor
angiogenesis and cancer treatment (12). In our preliminary study, flow
cytometric assay was performed to demonstrate that integrin
αvβ3 is the main target of
mPEG-SC20k-HM-3 (data not shown).
Current research indicates that the antitumor
property of mPEG-SC20k-HM-3 is closely related to the
target and its downstream pathways. Several signaling pathways are
switched on when the ligands bind with the integrin receptors,
including MAPK and Akt signaling pathways. The MAPK signaling
pathway is an important pipeline that transports signals from the
surface of cells to the nucleus. A variety of physiological
processes in cells, such as growth, development, differentiation,
division and apoptosis, are related to the regulation of the MAPK
signaling pathway. Sufficient evidence indicates that the MAPK
family participates in the process of cell adhesion, proteolysis of
extracellular matrix and cell migration. Members of the MAPK family
in mammals include extracellular signal regulated protein kinase
(ERK1/2), C-Jun N-terminal kinase (JNK) and p38 signaling pathways
(13). Among these members, the ERK
signaling pathway has been thoroughly studied, and was found to be
activated by Ras-Raf-MEK-ERK1/2 and participates in the migration
of different types of cells (14).
Abnormal PKB/Akt signaling is usually detected in human tumor
tissues and can be closely related to the process of tumor
development. The PKB/Akt signaling pathway participates in the
regulation of apoptosis, proliferation, differentiation and
metabolism of cells and plays an important role in angiogenesis.
PKB/Akt can be phosphorylated and activates endothelial nitric
oxide synthase (eNOS), which leads to angiogenesis and cell
migration (15). Another factor
connected with angiogenesis and cell migration is VEGF, with a
molecular weight of between 34 and 45 kb (16) and its sequences are highly
conserved. VEGF is a member of the platelet-derived growth factor
(PDGF) family isolated from the bovine pituitary follicle stellate
cell culture medium and can promote cell division, proliferation,
invasion and metastasis via binding to a specific receptor
(17). It is reported that the
specific receptor of VEGF is highly expressed in vascular
endothelial cells of tumor tissues compared with normal ones.
Based on the results of RT2 Profiler PCR
assays performed in the present study, mPEG-SC20k-HM-3
significantly down-regulated the expression of LECT1, F3, CDH6,
CST7I and TGA7, which are factors related to angiogenesis and tumor
metastasis. Concurrently, the results of the western blot assay
indicated that mPEG-SC20k-HM-3 inhibited the expression
of MEK1, p-MEK1, AKT1, p-AKT1, ERK1/2 and p-ERK1/2 in the ERK and
Akt signaling pathways. Therefore, the scheme of the mechanism of
action (Fig. 6B) based on the data
above can clearly explain the antitumor properties of
mPEG-SC20k-HM-3. Our previous research conducted on the
mechanism of HM-3 also involved the two signaling pathways
mentioned in the present study. However, in addition to the
expression of non-phosphorylated forms of the key proteins, the
expression divergences of phosphorylated forms of these proteins
were not taken into consideration, which was not rigorous (9). In addition, the explanation of the
relationship between HM-3 and VEGF is unreasonable as no trails
have demonstrated that HM-3 targets VEGF directly. Therefore, the
study on the mechanism of mPEG-SC20k-HM-3 is more
systematic and thorough. According to the results of RT2
Profiler PCR assays, the expression levels of MMP3 and MMP10 which
belong to the matrix metalloproteinase family were also
downregulated by mPEG-SC20k-HM-3. The MMP family are
zinc and calcium-dependent peptide chain endonucleases, which can
degradate various protein components in ECM to damage the
histological barrier of tumor cell invasion and as a result, tumor
metastasis can be inhibited (18).
The results that mPEG-SC20k-HM-3 downregulated MMP3 and
MMP10 expression indicated that mPEG-SC20k-HM-3 may also
target certain members of the MMP family. Certainly, the hypothesis
can be further demonstrated by a series of pharmacodynamic in
vitro studies and research regarding the related
mechanisms.
Acknowledgments
The present study was mainly supported by the 863
High-Technology Development Planning (no. SQ2011SF11B02030), the
Project Program of State Key Laboratory of national Medicines (no.
SKLNMBZ201403), the National Science and Techonology Major Projects
of New Drugs (nos. 2012ZX09103301-004 and 2014ZX09508007) in China
and a Project Funded by the Priority Academic Program Development
of Jangsu Higher Education Institutions.
References
|
1
|
Ryeom S and Folkman J: Role of endogenous
angiogenesis inhibitors in Down syndrome. J Craniofac Surg.
20(Suppl 1): S595–S596. 2009. View Article : Google Scholar
|
|
2
|
Ebos JM, Lee CR, Cruz-Munoz W, Bjarnason
GA, Christensen JG and Kerbel RS: Accelerated metastasis after
short-term treatment with a potent inhibitor of tumor angiogenesis.
Cancer Cell. 15:232–239. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Demetri GD, van Oosterom AT, Garrett CR,
Blackstein ME, Shah MH, Verweij J, McArthur G, Judson IR, Heinrich
MC, Morgan JA, et al: Efficacy and safety of sunitinib in patients
with advanced gastrointestinal stromal tumour after failure of
imatinib: A randomised controlled trial. Lancet. 368:1329–1338.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Motzer RJ, Michaelson MD, Redman BG, Hudes
GR, Wilding G, Figlin RA, Ginsberg MS, Kim ST, Baum CM, DePrimo SE,
et al: Activity of SU11248, a multitargeted inhibitor of vascular
endothelial growth factor receptor and platelet-derived growth
factor receptor, in patients with metastatic renal cell carcinoma.
J Clin oncol. 24:16–24. 2006. View Article : Google Scholar
|
|
5
|
Xu H, Pan L, Ren Y, Yang Y, Huang X and
Liu Z: RGD-modified angiogenesis inhibitor HM-3 dose: Dual function
during cancer treatment. Bioconjug Chem. 22:1386–1393. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yuan D, Shen H, Yuan S, Liu X, Xia X, Xie
P, Li W, Hu J, Liu Q and Xu H: Pharmacokinetics of HM-3 after
intravitreal administration in mice. Curr Eye Res. 39:837–844.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou K, Zheng X, Xu HM, Zhang J, Chen Y,
Xi T and Feng T: Studies of poly(ethylene glycol) modification of
HM-3 polypeptides. Bioconjug Chem. 20:932–936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu B, Xu HM, Zhao L, Huang X and Zhang F:
Site-specific modification of anti-angiogenesis peptide HM-3 by
polyethylene glycol molecular weight of 20 kDa. J Biochem.
148:341–347. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Z, Ren Y, Pan L and Xu HM: In vivo
anti-tumor activity of polypeptide HM-3 modified by different
polyethylene glycols (PEG). Int J Mol Sci. 12:2650–2663. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar
|
|
11
|
Silginer M, Burghardt I, Gramatzki D,
Bunse L, Leske H, Rushing EJ, Hao N, Platten M, Weller M and Roth
P: The aryl hydrocarbon receptor links integrin signaling to the
TGF-β pathway. Oncogene. 10:3872015.
|
|
12
|
Yan F, Xu X, Chen Y, Deng Z, Liu H, Xu J,
Zhou J, Tan G, Wu J and Zheng H: A lipopeptide-based
αvβ3 integrin-targeted ultrasound contrast agent for
molecular imaging of tumor angiogenesis. Ultrasound Med Biol.
41:2765–2773. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang C, Jacobson K and Schaller MD: MAP
kinases and cell migration. J Cell Sci. 117:4619–4628. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong F, Tian H, Yan S, Li L, Dong X, Wang
F, Li J, Li C, Cao Z, Liu X, et al: Dihydroartemisinin inhibits
endothelial cell proliferation through the suppression of the ERK
signaling pathway. Int J Mol Med. 35:1381–1387. 2015.PubMed/NCBI
|
|
15
|
Huang Y, Yu J, Wan F, Zhang W, Yang H,
Wang L, Qi H and Wu C: Panaxatriol saponins attenuated
oxygen-glucose deprivation injury in PC12 cells via activation of
PI3K/Akt and Nrf2 signaling pathway. Oxid Med Cell Longev.
2014:9780342014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Matsumoto K and Ema M: Roles of VEGF-A
signalling in development, regeneration, and tumours. J Biochem.
156:1–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ferrara N: Pathways mediating
VEGF-independent tumor angiogenesis. Cytokine Growth Factor Rev.
21:21–26. 2010. View Article : Google Scholar
|
|
18
|
Cathcart J, Pulkoski-Gross A and Cao J:
Targeting matrix metalloproteinases in cancer: Bringing new life to
old ideas. Genes Dis. 2:26–34. 2015. View Article : Google Scholar : PubMed/NCBI
|