Introduction
Gastric cancer (GC) is one of the most common
malignant tumors, and is responsible for many cancer-related deaths
worldwide (1,2). Although several treatments for GC
including excision surgery, radiotherapy and chemotherapy have been
developed recently, the clinical outcome continues to be poor in
patients with advanced GC (1). The
mortality of GC is intrinsically related to metastasis, in which
angiogenesis plays a crucial role (3). Angiogenesis is also indispensable to
the continuous growth of the tumor. Therefore, one of the most
promising yet challenging therapeutic approaches to cure GC is to
develop safe, effective, and affordable anti-angiogenic
therapies.
Tribbles homolog 3 (TRIB3, also named TRB3, NIPK,
and SKIP3) belongs to the tribbles family of pseudokinases that
were first identified in Drosophila to regulate cell
division and migration (4–6). TRIB3 also participates in the
activation of multiple signaling pathways, such as
mitogen-activated protein kinase (MAPK) pathways (7–9). TRIB3
expression is upregulated by endoplasmic reticulum stress, hypoxia,
and nutrient starvation (6,10,11).
Recent studies suggest that TRIB3 is a potential oncogene, as
evidenced by its elevated expression in colorectal cancer (12), breast cancer (13), liver cancer and other cancer tissues
(14). Furthermore, TRIB3 is
associated with an adverse prognosis in these cancers. However,
whether TRIB3 is involved in the development and progression of GC
has not been reported.
Previous studies have shown that TRIB3 expression is
closely related to the progression of type 2 diabetes mellitus, and
that TRIB3 mediated apoptosis in islet β cells, as well as insulin
resistance. Together, these effects impair insulin-stimulated
glucose uptake and maintain hyperglycemia in diabetes (15–17).
Chronic hyperglycemia is a major initiator of diabetic angiopathy
(18). Because impaired
angiogenesis is a key pathological characteristic of diabetic
microangiopathy (19), and it is
well established that some inducers of TRIB3, such as endoplasmic
reticulum stress, hypoxia, and glucose deprivation can also induce
angiogenesis, we propose that the expression of TRIB3 is related to
angiogenesis.
The present study analyzed TRIB3 expression in GC
tissues from 191 GC patients categorized from stage I to IV, to
examine the role of TRIB3 in GC. The study also examined the
relationship between TRIB3 and tumor angiogenesis. We found that
TRIB3 expression was elevated in GC tissues, and that TRIB3
overexpression is correlated to the severity and poor prognosis in
GC. We also showed that TRIB3 suppression downregulated the
expression of VEGF-A in GC cells, which subsequently suppressed the
recruitment of endothelial cells and formation of vessels. Thus,
TRIB3 may be a promising target for anti-angiogenic therapy in
GC.
Materials and methods
Patients and tumor tissue samples
Our study was approved by the Ethics Committee of
Nanfang Hospital, the Southern Medical University. All tissues for
diagnostic purposes were obtained with the consent of each patient
diagnosed with primary GC and receiving resection surgery between
2004 and 2008. The clinical stages of the tumors were defined
according to the National Comprehensive Cancer Network Guidelines
(http://www.nccn.org/; version 1.2014, Gastric
Cancer). No patient received any pre-operative chemotherapy or
radiotherapy. A total of 191 tissue samples were used in this
study.
Cells and cell culture
The human gastric epithelial cell line GES-1, and GC
cell lines BGC803, BGC823, MGC803, MKN28, MKN45, and SGC7901 were
obtained from Foleibao Biotechnology Development Company (Shanghai,
China). Human umbilical vein endothelial cells (HUVECs) were
purchased from Sciencell Research Laboratories (Carlsbad, CA, USA).
All cells were cultured in RPMI-1640 medium (Hyclone, Logan, UT,
USA) with 10% fetal bovine serum at 37°C and with 5%
CO2.
Immunohistochemistry
Immunohistochemical staining was used to evaluate
the expression of TRIB3, VEGF-A, and CD31, as described previously
(20). The primary antibodies used
in this study were polyclonal rabbit antibodies for TRIB3 (Abcam,
San Francisco, CA, USA), VEGF-A, and CD31 (ABclonal, Boston, MA,
USA). The HRP-conjugated anti-rabbit secondary antibody was from
CWBIO (Beijing, China). Specifically bound antibodies were detected
with a 3,3′-diaminobenzidine staining kit (CWBIO). The percentage
of positive cells was calculated and categorized as follows: 0 (0%
of cells stained), 1 (1–25%), 2 (26–50%), 3 (51–75%), or 4
(76–100%). Staining intensity was visually scored as 0 (negative),
1 (weak), 2 (moderate), or 3 (strong). The final score (0–12) for
the expression of TRIB3 and VEGF-A was calculated as the product of
the percentage and the intensity scores for each case.
Tumor microvessel density (MVD) was determined by
counting the number of endothelial cells positively stained for
CD31, following the method described by Weidner et al
(21). Slides were scanned
initially at a low power (×100 magnification) in order to identify
areas with higher vascular density (hot spots). Subsequently,
counting of the stained microvessels was performed on four
consecutive high power (×400 magnification) fields within the
selected high density fields. Yellow-brown immunostained
endothelial cells or an endothelial cell cluster that was clearly
separated from adjacent microvessels were counted as vascular
structures. The average number of microvessels counted in four 400×
fields provided the MVD value for each case.
Gene silencing with siRNAs
Three siRNA sequences against TRIB3 were purchased
from RiboBio (Guangzhou, China). Transfection was carried out using
Lipofectamine® 2000 transfection reagent from Invitrogen
(Carlsbad, CA, USA) following the manufacturer's recommended
protocol. A negative control siRNA was used to examine the effect
of transfection alone.
Western blotting
Proteins extracted from the tissues and cells were
subjected to western blotting as described previously (20). Primary antibodies were polyclonal
rabbit antibodies for TRIB3 (Abcam), VEGF-A (ABclonal), and α-actin
(Proteintech, Wuhan, China). A secondary fluorescent goat
anti-rabbit antibody (LI-COR, Lincoln, NE, USA) was used in this
study. The Odyssey imaging system (LI-COR) was used to scan the
blots.
Quantitative real-time polymerase chain
reaction (qRT-PCR)
Total RNA of the cultured cells was extracted using
a TRIzol kit (Takara Bio, Inc., Shiga, Japan) according to the
manufacturer's protocols. The First Strand cDNA synthesis kit
(Takara Bio, Inc.) was used to synthesize cDNAs. qRT-PCR was
performed using the SYBR-Green dye (Roche, Mannheim, Germany). The
PCR primers used in this study were: 5′-ATTAGGCAGGGTCTGTCCTGTG-3′
(TRIB3, sense), 5′-AGTATGGACCTGGGATTGTGGA-3′ (antisense);
5′-CTTGCCTTGCTGCTCTACC-3′ (VEGF-A, sense),
5′-CACACAGGATGGCTTGAAG-3′ (antisense); 5′-TTCATTGACCTCAACTACATG-3′
(GAPDH, sense), 5′-GTGGCAGTGATGGCATGGAC-3′ (antisense).
Enzyme-linked immunosorbent assay
(ELISA)
GC cells (3×104/well) were seeded in
24-well plates and TRIB3 siRNA transfection was performed. The
culture medium was collected 48 h after transfection and secretion
of VEGF-A was determined using human VEGF-A ELISA kits (Baomanbio,
Shanghai, China) according to the manufacturer's instructions. The
results are presented as mean ± standard deviation from three
independent experiments.
HUVEC migration assay
GC cells (3×104/well) were seeded in
24-well plates and TRIB3 siRNA transfection was performed. After
culturing at 37°C for 48 h, a chamber with a porous (8.0 mm pore
size) polycarbonate membrane filter (Millipore Corp., Bedford, MA,
USA) containing 4×104 HUVECs in 0.2 ml RPMI-1640 medium
without fetal bovine serum, was inserted into each well. HUVECs
were fixed with 4% paraformaldehyde after co-culture for 12 h at
37°C, and subsequently stained with 0.1% crystal violet for 30 min.
The results were observed under an inverted microscope.
Scratch wound healing assay
GC cells (3×104/well) were seeded in
6-well plates and TRIB3 siRNA transfection was performed. The
culture medium was collected 48 h after transfection. Trypsinized
HUVECs were seeded in 6-well plates at a density of
3×105 cells per well. After reaching confluency,
cultured cells were scratched with a sterile 200-µl pipette
tip. The cells were then cultured in the GC cell-conditioned
medium. Wound closure was observed at 0, 24, and 48 h under an
inverted microscope.
Tubule formation by HUVECs
HUVECs mixed with 100 µl GC cell-conditioned
medium were seeded at a density of 2×104 cells per well
in 96-well plates containing 60 µl solidified Matrigel (BD
Biosciences, New York, NY, USA) and cultured for 6–8 h at 37°C.
Cultures were photographed under a microscope. The number of
tubules was counted in three individual wells and presented as the
mean ± standard deviation.
Statistical analyses
Statistical calculations were performed using SPSS
13.0 software (IBM, Chicago, IL, USA). Survival analysis was
performed according to the Kaplan-Meier method. Differences of
survival between groups were assessed with the log-rank test. The
relationship between TRIB3 and MVD was evaluated by linear
regression analysis. Student's t-test was performed to determine
significant differences between two experimental groups. One-way
ANOVA analysis was used to evaluate the statistical significance
among multiple groups. P<0.05 was considered to indicate a
statistically significant result.
Results
TRIB3 expression is elevated in advanced
GC and associated with a poor prognosis
In order to detect the expression of TRIB3,
immunohistochemical staining was performed in 27 normal gastric
tissues and 191 GC tissues. Patients were divided into two groups,
low (score 0–5) and high (score >5) expression levels, according
to the TRIB3 staining score. TRIB3 was markedly upregulated in GC
compared with normal gastric tissues. High expression levels of
TRIB3 were observed in 61.8% (118/191) of the GC tissues. This was
significantly >7.4% (2/27) seen in normal gastric tissues.
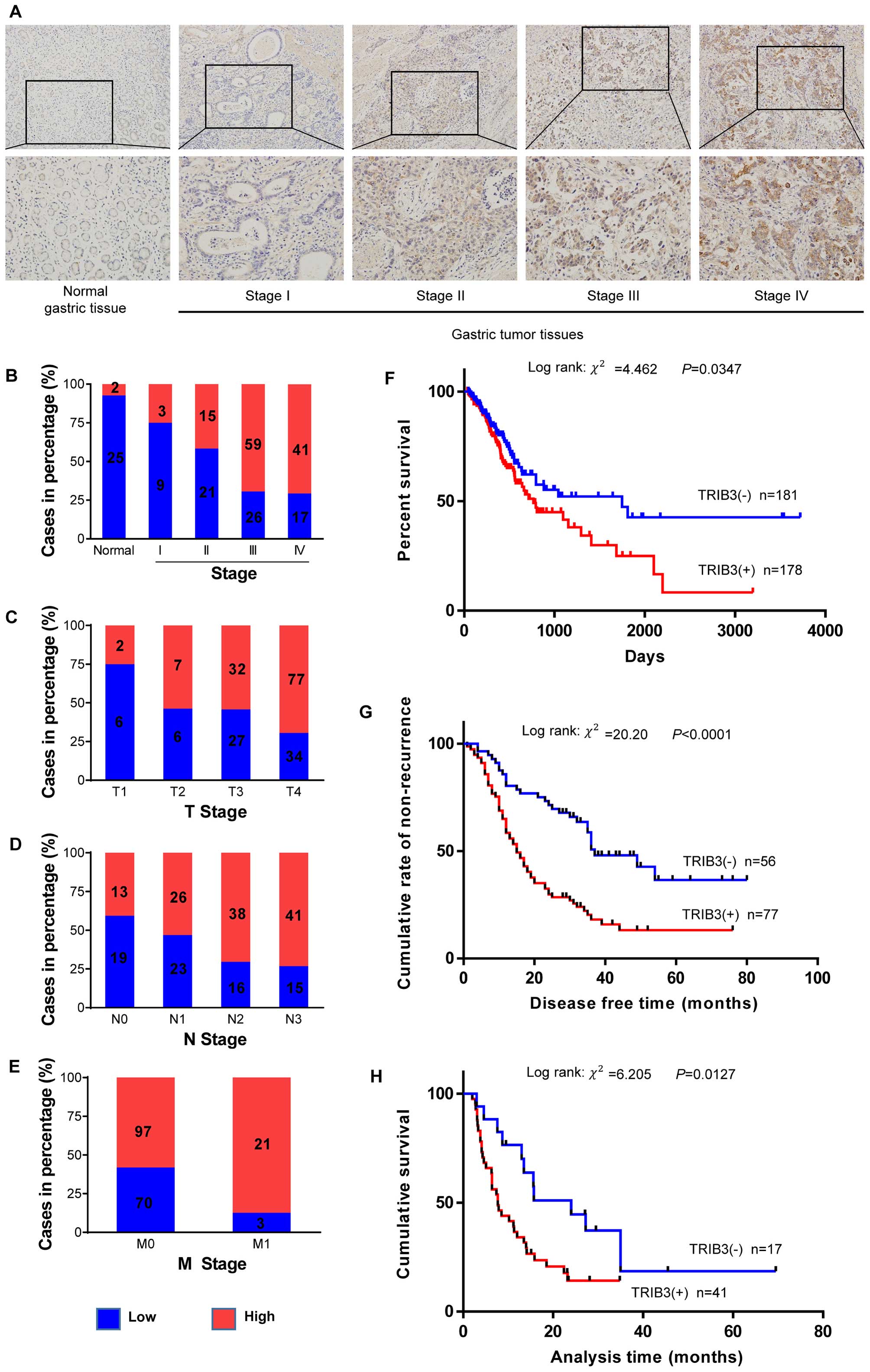
Representative staining examples of normal gastric and GC tissues
are shown in Fig. 1A.
The correlation analysis between pathological
characteristics and expression levels of TRIB3 showed that high
expression of TRIB3 was more frequent in patients with more
advanced overall TNM stage (P=0.001, Fig. 1B), T stage (P=0.029, Fig. 1C), N stage (P=0.006, Fig. 1D), and distant metastasis (P=0.006,
Fig. 1E). However, no statistical
significance was found with age and gender (P=0.339 and 0.578,
respectively, Table I).
 | Table ICorrelations between the
clinicopathological parameters and expression of TRIB3. |
Table I
Correlations between the
clinicopathological parameters and expression of TRIB3.
| Parameter | n | Expression of TRIB3
(n)
| P-value | χ2 |
|---|
| High | Low |
|---|
| Gender | | | | 0.578 | 0.309 |
| Male | 125 | 79 | 46 | | |
| Female | 66 | 39 | 27 | | |
| Age (years) | | | | 0.339 | 0.913 |
| ≤55 | 72 | 45 | 27 | | |
| >55 | 119 | 66 | 53 | | |
| TNM stage | | | | 0.001 | 17.089 |
| I | 12 | 3 | 9 | | |
| II | 36 | 15 | 21 | | |
| III | 85 | 59 | 26 | | |
| IV | 58 | 41 | 17 | | |
| Tumor invasion | | | | 0.029 | 9.059 |
| T1 | 8 | 2 | 6 | | |
| T2 | 13 | 7 | 6 | | |
| T3 | 59 | 32 | 27 | | |
| T4 | 111 | 77 | 34 | | |
| Lymph node
metastasis | | | | 0.006 | 12.431 |
| N0 | 32 | 13 | 19 | | |
| N1 | 49 | 26 | 23 | | |
| N2 | 54 | 38 | 16 | | |
| N3 | 56 | 41 | 15 | | |
| Distant
metastasis | | | | 0.006 | 7.69 |
| M0 | 167 | 97 | 70 | | |
| M1 | 24 | 21 | 3 | | |
Kaplan-Meier curves for overall survival duration of
GC patients, according to TRIB3 expression levels using data from
the TCGA database, showed that GC patients who had higher levels of
TRIB3 expression exhibited a worse prognosis (log-rank =4.462,
P=0.0347, Fig. 1F). Similar results
were observed in the present study where high expression levels of
TRIB3 were significantly correlated with short time to recurrence
among patients in stages I–III(log-rank =20.20, P<0.0001,
Fig. 1G), and short survival time
among patients in stage IV (log-rank =6.205, P=0.0127, Fig. 1H).
Overexpression of TRIB3 is associated
with tumor angiogenesis in GC
To clarify the potential relationship between TRIB3
and angiogenesis, a total of 91 GC tissues were immunostained for
TRIB3 and angiogenesis-related markers. VEGF-A is an important
hallmark of angiogenesis, while microvessels were identified by
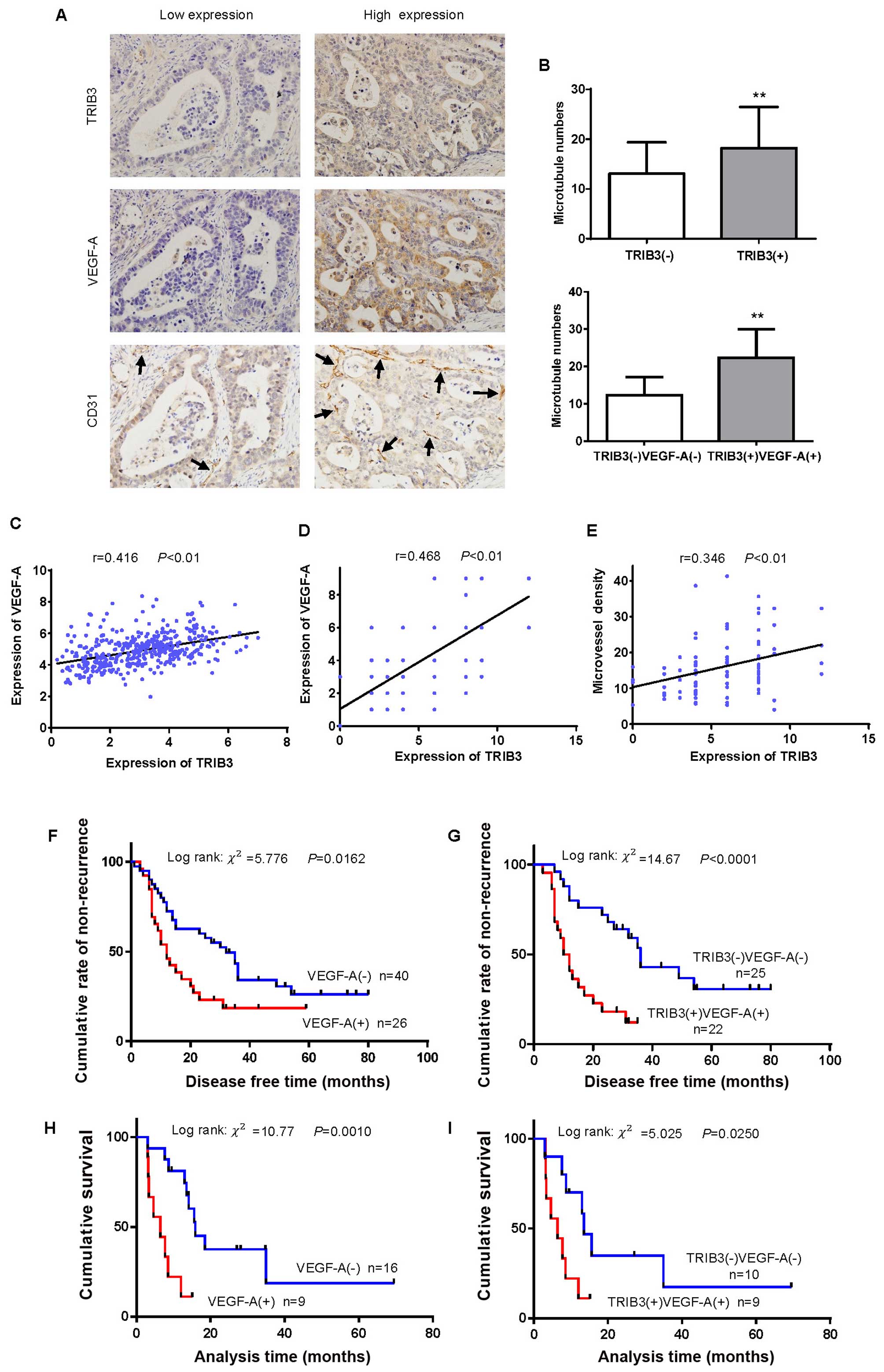
CD31 staining. Representative staining examples of low and high
expression of TRIB3, VEGF-A, and CD31 in GC are shown in Fig. 2A. Tumors with high expression levels
of TRIB3 had a significantly higher MVD than those with low
expression levels (18.2±1.1 vs. 13.1±1.0, t=3.261, P=0.0016,
Fig. 2B). Similar results were
observed in the groups where both TRIB3 and VEGF-A expression were
elevated (22.4±1.4 vs. 12.3±0.82, t=6.460, P<0.0001, Fig. 2B). Using data from the TCGA
database, linear regression analysis revealed that TRIB3 and VEGF-A
expressions were significantly correlated (r=0.416, P<0.01,
Fig. 2C). Similarly, a significant
positive correlation between the expression of TRIB3 and VEGF-A was
found in the present study (r=0.468, P<0.01, Fig. 2D). TRIB3 positivity was also
significantly correlated with MVD by linear regression analysis
(r=0.346, P<0.01, Fig. 2E).
Using Kaplan-Meier analysis with the log-rank test,
we found that high expression of VEGF-A was significantly
correlated with short time to recurrence among GC patients in
stages I–III (log-rank =5.776, P=0.0162, Fig. 2F). Similar results were observed in
patients expressing high levels of VEGF-A combined with high levels
of TRIB3 (log-rank =14.61, P<0.0001, Fig. 2G). In addition, high expression of
VEGF-A was associated with short survival time among stage IV GC
patients (log-rank =10.77, P=0.001, Fig. 2H). Similar results were observed in
patients with high levels of both VEGF-A and TRIB3 (log-rank
=5.025, P=0.025, Fig. 2I).
Expression of TRIB3 and VEGF-A in GC
surgical samples and GC cell lines
To confirm the elevated expression of TRIB3 and
VEGF-A in GC tissues, fresh surgically resected specimens were
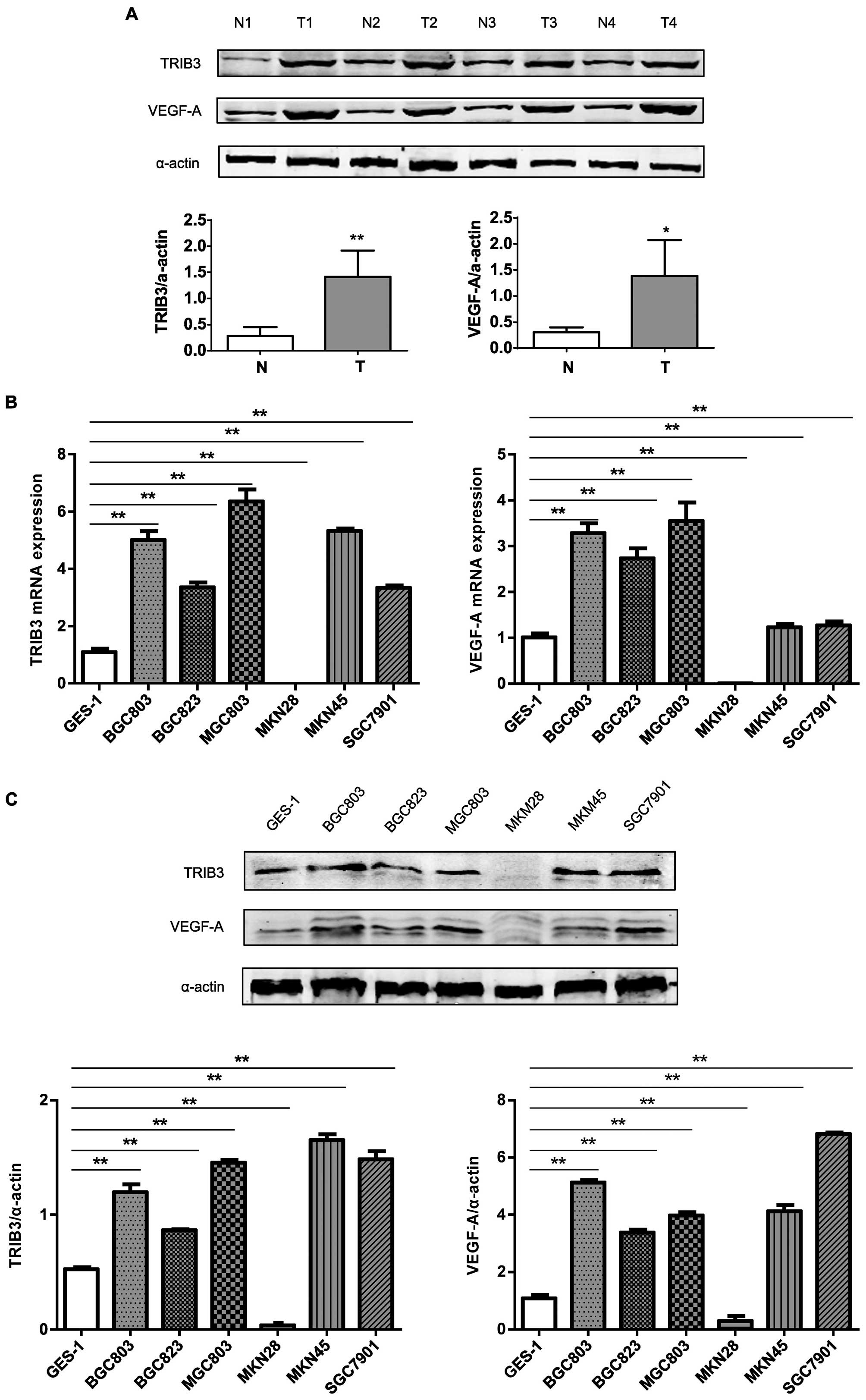
analyzed by western blotting. Both TRIB3 and VEGF-A were expressed
at higher levels in tumor tissues than in the adjacent non-tumorous
gastric tissues (Fig. 3A). To
better elucidate the expression profiles of TRIB3 and VEGF-A, we
examined their mRNA and protein expression levels in a human
gastric epithelial cell line (GES-1) and six GC cell lines (BGC803,
BGC823, MGC803, MKN28, MKN45, and SGC7901). Except for MKN28 cells,
the expression of TRIB3 mRNA was higher in GC (3- to 7-fold) than
in GES-1 cells (Fig. 3B).
Interestingly, a similar trend was observed in the expression of
VEGF-A mRNA (Fig. 3B). The
expression of TRIB3 and VEGF-A proteins were consistent with the
mRNA results (Fig. 3C).
TRIB3 silencing downregulates the
expression of VEGF-A in GC cells
The ability of three different TRIB3-siRNA sequences
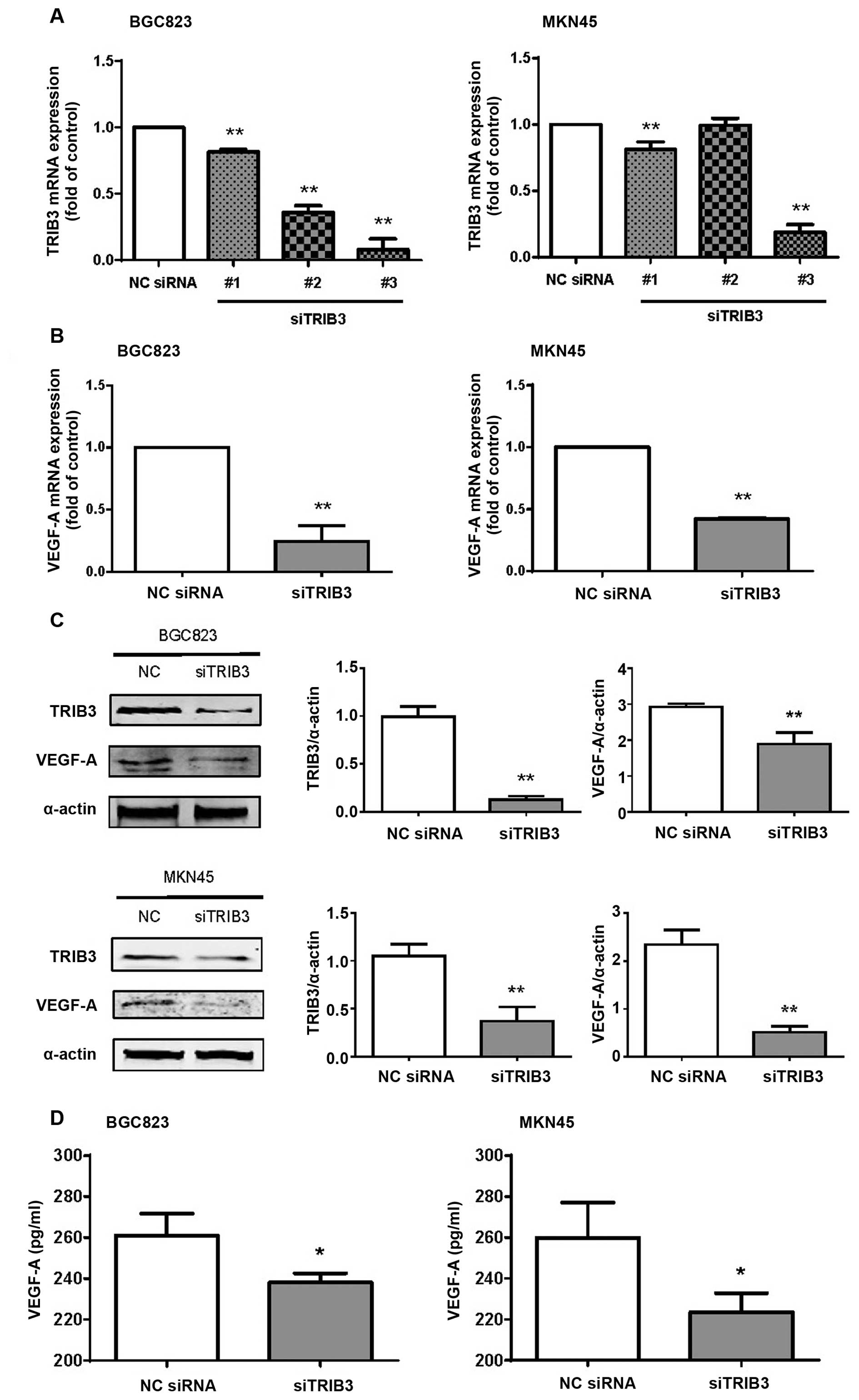
to downregulate TRIB3 was determined by qRT-PCR. Only siTRIB3-#3
consistently suppressed the expression of TRIB3 mRNA in the BGC823
and MKN45 cells (Fig. 4A).
Interestingly, the expression of VEGF-A mRNA was also downregulated
by TRIB3-#3-siRNA in the BGC823 and MKN45 cells (P=0.0005 and
P<0.0001, respectively, Fig.
4B). Both TRIB3 and VEGF-A protein levels were significantly
reduced by TRIB3-#3-siRNA in the BGC823 (P=0.0002 and 0.0062,
respectively, Fig. 4C) and MKN45
(P=0.0036 and 0.0006, respectively, Fig. 4C) cells. ELISA analyses showed that
secreted VEGF-A protein was also decreased in the TRIB3-knockdown
GC cells, when compared with control cells (P=0.0279 and 0.0329 in
BGC823 and MKN45 cells, respectively, Fig. 4D). These data showed that TRIB3
silencing contributed to the downregulation of VEGF-A mRNA and
protein expression in the GC cells.
TRIB3 silencing in GC cells compromises
their ability to recruit endothelial cells via soluble factors
Angiogenesis is dependent on the proliferation and
migration of vascular endothelial cells. To examine whether TRIB3
silencing influenced angiogenesis, we evaluated the responses of
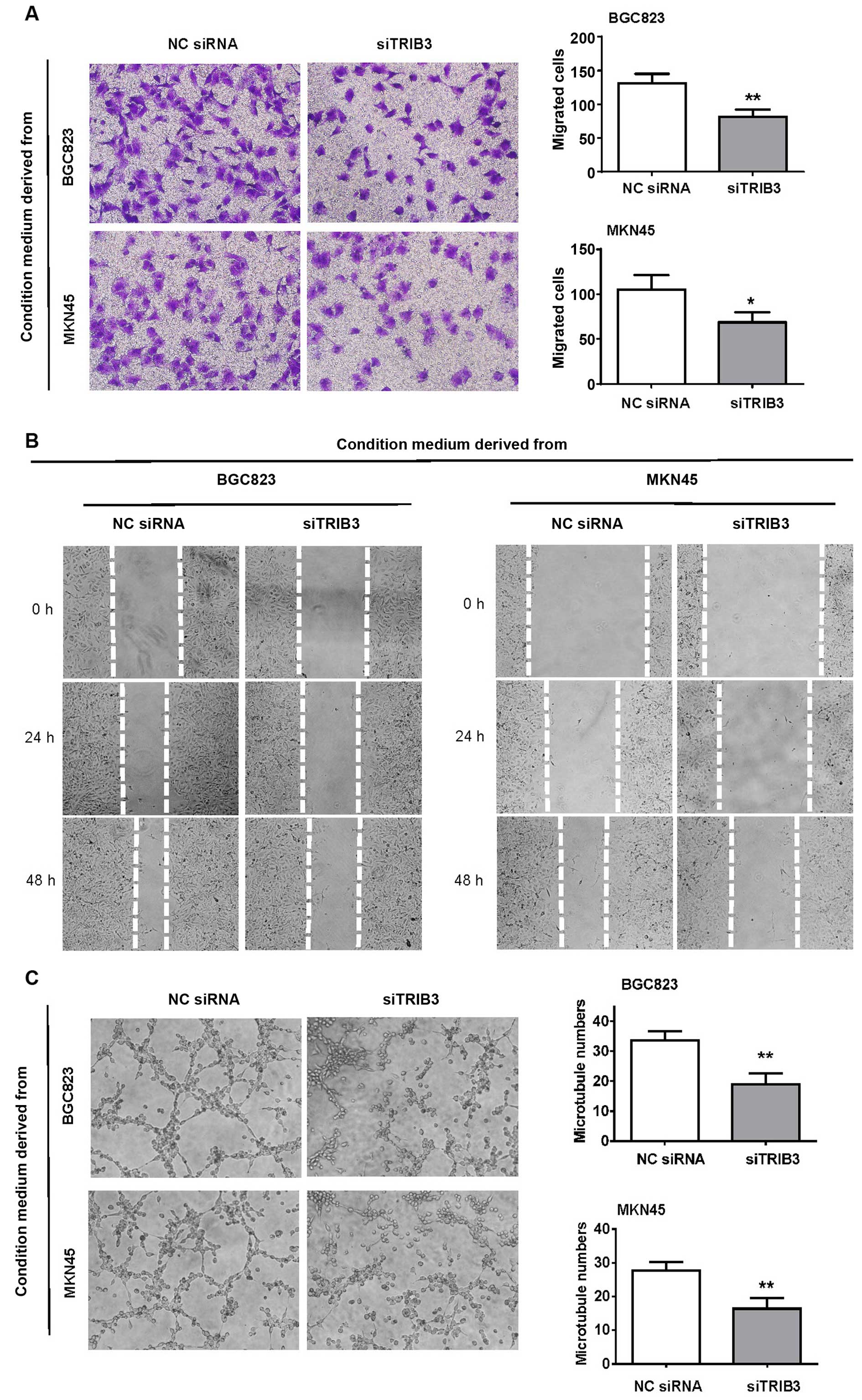
HUVECs in terms of migration and tubule formation. The migration
capacity of HUVECs was altered after co-culturing with GC
cell-conditioned media as shown by both the migration and scratch
wound healing assays (Fig. 5A and
B). Silencing of TRIB3 in the GC cells significantly impaired
their ability to recruit HUVECs. Significant differences in the
tubule-like structure formation capacity of HUVECs were also
observed between the control siRNA and TRIB3-#3-siRNA groups
(Fig. 5C).
Discussion
The purpose of the present study was to investigate
the oncological significance of TRIB3 in GC. We evaluated the
expression of TRIB3 in patients with GC in order to determine
correlations with pathological variables, including TNM stage,
survival time of patients, and incidence of cancer recurrence, and
to examine the relationship with angiogenesis.
Tribbles, first identified in Drosophila, are
members of the pseudokinase family of proteins with no associated
kinase activity. Instead of direct phosphorylation of target
proteins, tribbles act as adaptor or scaffold proteins, as well as
decoy kinases, that impede the function of other kinases through
obstructive binding (22,23). Currently, three mammalian homologs
of the tribbles gene are known. One of these, TRIB3, plays an
important role in multiple signaling pathways, coordinating crucial
cellular processes such as apoptosis, glucose and lipid metabolism,
adipocyte differentiation, and cell stress (15,24–27).
TRIB3 is overexpressed in many cancers, but whether TRIB3 is
upregulated in GC has not been reported previously.
To the best of our knowledge, this is the first
study to evaluate the expression of TRIB3 in GC patients and GC
cell lines, and examine the role of TRIB3 in tumor angiogenesis.
Our results showed that TRIB3 was significantly upregulated in GC
tissues compared with its level in adjacent non-tumor tissues, and
that high expression levels of TRIB3 were more frequent in patients
at a more advanced overall TNM stage, T stage, N stage, and with
distant metastasis. High expression levels of TRIB3 predicted high
cancer recurrence and mortality for GC patients. At the cellular
level, TRIB3 was constitutively expressed at higher levels in all
GC cell lines tested, except MKN28, when compared to a gastric
epithelial cell line. These data show the potential utility of
TRIB3 as a prognostic marker for GC.
In colorectal cancer, TRIB3 is a prognostic marker
and has functional relevance to cell growth (12). In breast cancer, TRIB3 is involved
in the ability of cancer cells to survive in hypoxic conditions and
is associated with a poor prognosis (13). Similar relationships were noted in
other types of cancer (14,28,29).
Knockdown of TRIB3 in tumor cells significantly inhibited the
invasive and metastatic ability of the cells by promoting
mesenchymal-epithelial transition (30). The finding of elevated TRIB3
expression in different malignant tumors supports the clinical
results of our study, implying that TRIB3 may be overexpressed
ubiquitously in tumors and play an important role in promoting
tumor progression.
The role of TRIB3 in multiple signaling pathways,
such as the MAPK pathway, combined with its role in hypoxia,
metabolism, and cell stress processes, originally spurred us to
investigate its role in angiogenesis. Because angiogenesis is
dependent on the proliferation and migration of vascular
endothelial cells, where VEGF-A plays an extremely important role,
specific studies were conducted to identify the role of TRIB3 in
angiogenesis. Firstly, immunohistochemical staining of 91 GC
tissues showed a significant positive correlation between the
expression of TRIB3 and VEGF-A. In addition, tumors with high
expression levels of TRIB3 had a higher MVD than those with low
expression levels of TRIB3. At the cellular level, the expression
of TRIB3 and VEGF-A were, with only one exception, consistent in a
human gastric epithelial cell line and GC cell lines, suggesting
that there is a correlation between the expression levels of the
two genes. Both mRNA and protein expression levels of VEGF-A were
significantly reduced by TRIB3-siRNA in GC cells. These data
strongly suggested that TRIB3 silencing downregulated the
expression of VEGF-A in GC cells. Further studies showed that TRIB3
silencing in GC cells significantly impaired their ability to
recruit endothelial cells via soluble factors, implying that TRIB3
silencing may downregulate angiogenesis in vitro.
Overall, tumor angiogenesis is a complex process.
Whether TRIB3 directly or indirectly modulates VEGF-A is still
unclear and requires further investigation.
In summary, our study shows that: i) the expression
of TRIB3 is significantly upregulated in GC; ii) high expression
levels of TRIB3 are associated with a shorter survival time and a
higher incidence of cancer recurrence and metastasis; and iii)
TRIB3 silencing contributes to the downregulation of VEGF-A
expression and angiogenesis in GC. Taken together, our data suggest
that suppression of TRIB3 may be a novel anti-angiogenic approach
for GC treatment.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 31271564 and 81472314),
the Special Foundation for National Clinical Specialties of China
(to The Department of Oncology, Nanfang Hospital), and the Team
Program of Natural Science Foundation of Guangdong Province, China
(no. 2011030003134).
References
|
1
|
Bang YJ, Van Cutsem E, Feyereislova A,
Chung HC, Shen L, Sawaki A, Lordick F, Ohtsu A, Omuro Y, Satoh T,
et al: ToGA Trial Investigators: Trastuzumab in combination with
chemotherapy versus chemotherapy alone for treatment of
HER2-positive advanced gastric or gastro-oesophageal junction
cancer (ToGA): A phase 3, open-label, randomised controlled trial.
Lancet. 376:687–697. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shen L, Shan YS, Hu HM, Price TJ, Sirohi
B, Yeh KH, Yang YH, Sano T, Yang HK, Zhang X, et al: Management of
gastric cancer in Asia: Resource-stratified guidelines. Lancet
Oncol. 14:e535–e547. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gacche RN: Compensatory angiogenesis and
tumor refractoriness. Oncogenesis. 4:e1532015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grosshans J and Wieschaus E: A genetic
link between morphogenesis and cell division during formation of
the ventral furrow in Drosophila. Cell. 101:523–531. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mata J, Curado S, Ephrussi A and Rørth P:
Tribbles coordinates mitosis and morphogenesis in Drosophila by
regulating string/CDC25 proteolysis. Cell. 101:511–522. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bowers AJ, Scully S and Boylan JF: SKIP3,
a novel Drosophila tribbles ortholog, is overexpressed in human
tumors and is regulated by hypoxia. Oncogene. 22:2823–2835. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kiss-Toth E, Bagstaff SM, Sung HY, Jozsa
V, Dempsey C, Caunt JC, Oxley KM, Wyllie DH, Polgar T, Harte M, et
al: Human tribbles, a protein family controlling mitogen-activated
protein kinase cascades. J Biol Chem. 279:42703–42708. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rzymski T, Paantjens A, Bod J and Harris
AL: Multiple pathways are involved in the anoxia HuR-regulated RNA
stability, NF-kappaB and ATF4. Oncogene. 27:4532–4543. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Du K, Herzig S, Kulkarni RN and Montminy
M: TRB3: A tribbles homolog that inhibits Akt/PKB activation by
insulin in liver. Science. 300:1574–1577. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ord D and Ord T: Characterization of human
NIPK (TRB3, SKIP3) gene activation in stressful conditions. Biochem
Biophys Res Commun. 330:210–218. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schwarzer R, Dames S, Tondera D, Klippel A
and Kaufmann J: TRB3 is a PI 3-kinase dependent indicator for
nutrient starvation. Cell Signal. 18:899–909. 2006. View Article : Google Scholar
|
|
12
|
Miyoshi N, Ishii H, Mimori K, Takatsuno Y,
Kim H, Hirose H, Sekimoto M, Doki Y and Mori M: Abnormal expression
of TRIB3 in colorectal cancer: A novel marker for prognosis. Br J
Cancer. 101:1664–1670. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wennemers M, Bussink J, Scheijen B,
Nagtegaal ID, van Laarhoven HW, Raleigh JA, Varia MA, Heuvel JJ,
Rouschop KM, Sweep FC, et al: Tribbles homolog 3 denotes a poor
prognosis in breast cancer and is involved in hypoxia response.
Breast Cancer Res. 13:R822011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hua F, Li K, Yu JJ, Lv XX, Yan J, Zhang
XW, Sun W, Lin H, Shang S, Wang F, et al: TRB3 links insulin/IGF to
tumour promotion by interacting with p62 and impeding
autophagic/proteasomal degradations. Nat Commun. 6:79512015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fang N, Zhang W, Xu S, Lin H, Wang Z, Liu
H, Fang Q, Li C, Peng L and Lou J: TRIB3 alters endoplasmic
reticulum stress-induced β-cell apoptosis via the NF-κB pathway.
Metabolism. 63:822–830. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang W, Liu J, Tian L, Liu Q, Fu Y and
Garvey WT: TRIB3 mediates glucose-induced insulin resistance via a
mechanism that requires the hexosamine biosynthetic pathway.
Diabetes. 62:4192–4200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu J, Zhang W, Chuang GC, Hill HS, Tian
L, Fu Y, Moellering DR and Garvey WT: Role of TRIB3 in regulation
of insulin sensitivity and nutrient metabolism during short-term
fasting and nutrient excess. Am J Physiol Endocrinol Metab.
303:E908–E916. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamagishi S and Imaizumi T: Diabetic
vascular complications: Pathophysiology, biochemical basis and
potential therapeutic strategy. Curr Pharm Des. 11:2279–2299. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng R and Ma JX: Angiogenesis in
diabetes and obesity. Rev Endocr Metab Disord. 16:67–75. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang L, Wu Y, Lin L, Liu P, Huang H, Liao
W, Zheng D, Zuo Q, Sun L, Huang N, et al: Metastasis-associated in
colon cancer-1 upregulation predicts a poor prognosis of gastric
cancer, and promotes tumor cell proliferation and invasion. Int J
Cancer. 133:1419–1430. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis-correlation in invasive breast
carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yokoyama T and Nakamura T: Tribbles in
disease: Signaling pathways important for cellular function and
neoplastic transformation. Cancer Sci. 102:1115–1122. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lohan F and Keeshan K: The functionally
diverse roles of tribbles. Biochem Soc Trans. 41:1096–1100. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Örd T, Örd D, Adler P, Vilo J and Örd T:
TRIB3 enhances cell viability during glucose deprivation in
HEK293-derived cells by upregulating IGFBP2, a novel nutrient
deficiency survival factor. Biochim Biophys Acta. 1853:2492–2505.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bezy O, Vernochet C, Gesta S, Farmer SR
and Kahn CR: TRB3 blocks adipocyte differentiation through the
inhibition of C/EBPbeta transcriptional activity. Mol Cell Biol.
27:6818–6831. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo L, Guo ZX, Gong HP, Shang YY, Zhong M,
Zhang Y and Zhang W: Tribbles homolog 3 is induced by high glucose
and associated with apoptosis in human endothelial cells. Mol Med
Rep. 12:1963–1970. 2015.PubMed/NCBI
|
|
27
|
Brisard D, Chesnel F, Elis S, Desmarchais
A, Sánchez-Lazo L, Chasles M, Maillard V and Uzbekova S: Tribbles
expression in cumulus cells is related to oocyte maturation and
fatty acid metabolism. J Ovarian Res. 7:442014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang J, Wen HJ, Guo ZM, Zeng MS, Li MZ,
Jiang YE, He XG and Sun CZ: TRB3 overexpression due to endoplasmic
reticulum stress inhibits AKT kinase activation of tongue squamous
cell carcinoma. Oral Oncol. 47:934–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou H, Luo Y, Chen JH, Hu J, Luo YZ, Wang
W, Zeng Y and Xiao L: Knockdown of TRB3 induces apoptosis in human
lung adenocarcinoma cells through regulation of Notch 1 expression.
Mol Med Rep. 8:47–52. 2013.PubMed/NCBI
|
|
30
|
Hua F, Mu R, Liu J, Xue J, Wang Z, Lin H,
Yang H, Chen X and Hu Z: TRB3 interacts with SMAD3 promoting tumor
cell migration and invasion. J Cell Sci. 124:3235–3246. 2011.
View Article : Google Scholar : PubMed/NCBI
|