Introduction
Epithelial ovarian cancer (EOC) is the most lethal
gynecological malignancy (1). Over
70% of EOC patients are diagnosed with distant metastasis and their
5-year survival rate is less than 30% (2). Surgery and platinum-based chemotherapy
are the main clinical treatments for most advanced EOC patients.
However, most patients eventually experience cancer recurrence.
Thus, an immunosuppressive EOC microenvironment should be overcome
and new therapeutic approaches, such as immunotherapy, are
needed.
Recently, many studies have shown that chronic
inflammation plays a significant role in tumor progression
(3). Many macrophages are found to
infiltrate the tumor tissue in many solid malignancies. These
tumor-associated macrophages (TAMs) are known to be recruited from
peripheral blood monocytes by chemokines (4) and are involved in tumor progression
(5). Many studies indicate that
TAMs are associated with poor prognosis in various types of cancers
(6–8). TAMs predominantly exhibit an M2
phenotype, which plays an important role in tumor angiogenesis,
aggressiveness and modifies the tumor microenvironment. TAMs have
the ability to produce high amounts of cytokines, which may
contribute to tumor formation and invasion (9).
The interleukin (IL) family is a group of
inflammatory cytokines. The major secretory characteristic of TAMs
includes the production of interleukin-10 (IL-10), which is an
immunosuppressive cytokine which participates in the growth and
metastasis of different human malignant tumors. Sources of IL-10
include monocytes, a subtype of dendritic cells which activates
macrophages and TAMs (10). IL-10
plays an important role in preventing inflammation in the negative
feedback loop (11). The expression
of IL-10 has been evaluated in many solid tumors, such as
esophageal squamous cell, non-small cell lung and hepatocellular
carcinoma (12–14).
Regulatory T cells (Tregs), a subpopulation of
CD4+ T lymphocytes, may suppress antitumor functions and
regulate immune tolerance (15,16).
Some studies have demonstrated that Treg cells can differentiate
from naïve CD4+ T cells in the presence of soluble
factors such as TGF-β and IL-10 (17). However, the significance of IL-10
secreted by TAMs and its mechanism in regulating T cells in EOC
remains unknown. In the present study, we aimed to explore the
relationship between IL-10 and the accumulation of Treg cells in
EOC and investigated the mechanism by which TAMs induce Treg cell
expression through IL-10.
Materials and methods
Patients
A total of 40 EOC samples and 20 benign ovarian
tumor samples, obtained from the Shanghai First Maternity and
Infant Hospital, Tongji University School of Medicine (Shanghai,
China) between January 2013 and 2015, were included in the present
study (Table I). Peripheral blood
mononuclear cells (PBMCs) were provided by the Shanghai Blood
Center (Shanghai, China). Ethical approval for the present study
was obtained from the Institutional Review Board of the Shanghai
First Maternity and Infant Hospital, Tongji University School of
Medicine.
 | Table I.Clinical characteristics of the 60
studied patients. |
Table I.
Clinical characteristics of the 60
studied patients.
| Variables | n=60 | Percent (%) |
|---|
| Diagnostic
categories |
|
|
| Benign
ovarian tumors | 20 | 33.33 |
|
Epithelial ovarian cancer | 40 | 66.67 |
| Histologic type |
|
|
|
Serous | 24 | 60.00 |
|
Non-serous | 16 | 40.00 |
| Tumor grade |
|
|
| Grade
I | 1 | 2.50 |
| Grade
II | 18 | 45.00 |
| Grade
III | 21 | 52.50 |
| Clinical stage |
|
|
| Stage
I | 11 | 27.50 |
| Stage
II | 9 | 22.50 |
| Stage
III | 19 | 47.50 |
| Stage
IV | 1 | 2.50 |
Immunofluorescence
Specimens were fixed in 4% neutral buffered
formalin, embedded in OCT and cut into 8-µm sections. Treg cells
were analyzed by immunofluorescence using double staining of CD4
and Foxp3. TAMs were analyzed using CD68. BSA (5%) (Sigma-Aldrich,
St. Louis, MO, USA) was used to block non-specific binding. Then,
sections were incubated with primary antibodies (anti-CD4,
anti-Foxp3 and anti-CD68; Abcam, Cambridge, MA, USA) in a
humidified chamber overnight at 4°C. After being washed, the
secondary anti-mouse IgG (for CD4; Invitrogen Life Technologies,
Carlsbad, CA, USA) and anti-rabbit IgG (for Foxp3 and CD68;
Invitrogen Life Technologies) were added for 45 min at room
temperature. All reagents were used except the primary antibodies
as negative controls. Sections were analyzed with a fluorescence
microscope (Leica, Germany). Treg cells and TAMs were
microscopically quantified by counting five fields of view at a
magnification of ×400.
ELISA assays for the detection of
inflammatory factors in ascites and blood serum
Ascites samples of 16 EOC patients and blood serum
samples of 7 healthy subjects were collected and stored at −80°C.
Subsequently, they were used for quantification of IL-10, IL-2 and
IL-12 expression levels by ELISA assays according to the
manufacturers instructions (RayBiotech, Inc., Atlanta, GA,
USA).
Immunohistochemistry
IL-10 was analyzed by immunohistochemistry. Sections
were incubated in 3% hydrogen peroxide to block endogenous peroxide
activity. BSA (5%) was used to block non-specific binding. After
being washed, the sections were incubated with primary antibodies
(mouse anti-human IL-10; Abcam) overnight at 4°C in a humidified
chamber. A secondary antibody (goat anti-mouse IgG) was added to
the sections and DAB kits (both from Vector Laboratories, Inc.,
Burlingame, CA, USA) were used. All reagents were used except for
the IL-10 as negative controls. The IL-10 content was
microscopically observed at a magnification of ×200 and ×400.
Detection of IL-10 in TAMs and T-cell
co-cultured supernatants
CD4+ T cells and fresh peripheral blood
CD14+ monocytes were purified with immunomagnetic beads
(Miltenyi Biotec Inc., Cambridge, MA, USA) from PBMCs provided by
the Shanghai Blood Center. Cells (107) were cultured
with 20 µl anti-CD4 beads (anti-CD14 beads) for 20 min at 4̊C.
Then, the cells were resuspended in 800 µl buffer and the
suspension was applied onto the column. The M1 macrophages were
induced by M-CSF (10 ng/ml; R&D Systems, Inc., Minneapolis, MN,
USA), LPS (500 ng/ml; Sigma-Aldrich) and IFN-r (20 ng/ml; R&D
Systems). The M2 macrophages were induced by M-CSF (10 ng/ml) and
IL-4 (20 ng/ml; both from R&D Systems). CD4+ T cells
were co-cultured with M1/M2 macrophages for 3 days and then the
IL-10 concentration in the supernatant was detected using an IL-10
ELISA kit (RayBiotech). After 30 min, the optical density was
detected at 450 nm using a microplate reader.
Quantitative real-time PCR
The RNA was extracted from T cells co-cultured with
the M1 group and M2 group using TRIzol. Takara Retroviral Reverse
Transcriptase kit (Takara Bio, Inc., Otsu, Japan) was used to
synthesize cDNA following the manufacturers instructions. Primers
for the Foxp3 gene were designed as follows: forward,
5-CCCTCAACTGAGAA CTCAAGTC-3 and reverse, 5-AGGTTGGCACCATAGTCT
CCA-3.
Western blotting
After 3 days of co-culture, CD4+ T cells
from the control, M1 and M2 group were treated with total protein
lysis buffer (DingGuo Biotechnology Co., Ltd., Shanghai, China).
Detection of Foxp3 was performed with a primary antibody against
Foxp3 (1:200; Abcam) by western blotting.
The co-cultured system building in
flow cytometric analysis of Treg cells
The macrophages were induced only by M-CSF (10
ng/ml; R&D Systems). Moreover, CD4+ T cells were
incubated with Mφ macrophages/Mφ macrophages + IL-10 (200 pg/ml) in
a 6-well plate (106 cells/well) for 3 days. T cells were
pre-stimulated with anti-CD28 (5 µg/ml) and anti-CD3 (10 µg/ml;
both from BD Biosciences, USA) for 3 days. After being co-cultured
with macrophages and IL-10, the proportion of
CD4+Foxp3+ Treg cells was detected by flow
cytometric analysis. Firstly, the cells were stimulated with a
T-cell stimulation cocktail and stained extracellularly with CD4
Pe-Cy5.5 (eBioscience, Inc., San Diego, CA, USA) for 45 min at 4̊C.
Then, the cells were fixed and infiltrated with Perm/Fix solution
(eBioscience) for 30 min. Subsequently, the cells were
intracellularly stained with anti-Foxp3-allophycocyanin (APC)
(eBioscience) for 45 min at 4̊C. All aforementioned steps were
protected from light. Data were analyzed by a
fluorescence-activated cell sorter (FACS).
The SKOV3 mouse subcutaneous
model
The human EOC cells SKOV3-luciferase (SKOV3-Luc)
were grown in Dulbeccos modified Eagles medium (DMEM) with 10%
fetal bovine serum (FBS) and 1% antibiotics. Peripheral blood
CD4+ T cells were purified from PBMCs with
immunomagnetic beads. Female nude mice (5 weeks of age, 15 g) were
purchased from Tongji University School of Medicine, Shanghai,
China and bred under SPF conditions; three groups were mixed with
the SKOV3-Luc cells for the establishment of the mouse model (four
mice per group). Nude mice from each group were injected with 0.1
ml of RPMI-1640 medium containing 1×106 SKOV3-Luc cells
and 1×105 CD4+ T cells with or without IL-10.
Tumor weight, tumor volume and total tumor flux were recorded. The
SKOV3-Luc tumors in vivo were tested for Luc expression by
D-luciferin (100 mg/kg; Life Technologies, Grand Island, NY, USA)
and captured with the IVIS Spectrum (PerkinElmer, Inc., Waltham,
MA, USA) for observation. The animal experiments were approved by
the Medical Animal Care Committee and all the animals were cared
for and only used in the laboratory.
Data and statistical analysis
Continuous data were evaluated using the LSD test,
Levenes test and the Mann-Whitney U test. Overall survival was
evaluated using Kaplan-Meier curves (log-rank test). Data are
presented as the mean ± standard deviation. Figures are presented
as the mean ± standard error of the mean. The statistical analysis
was carried out using SPSS 22.0 and GraphPad Prism 6 software.
P<0.05 was considered statistically significant.
Results
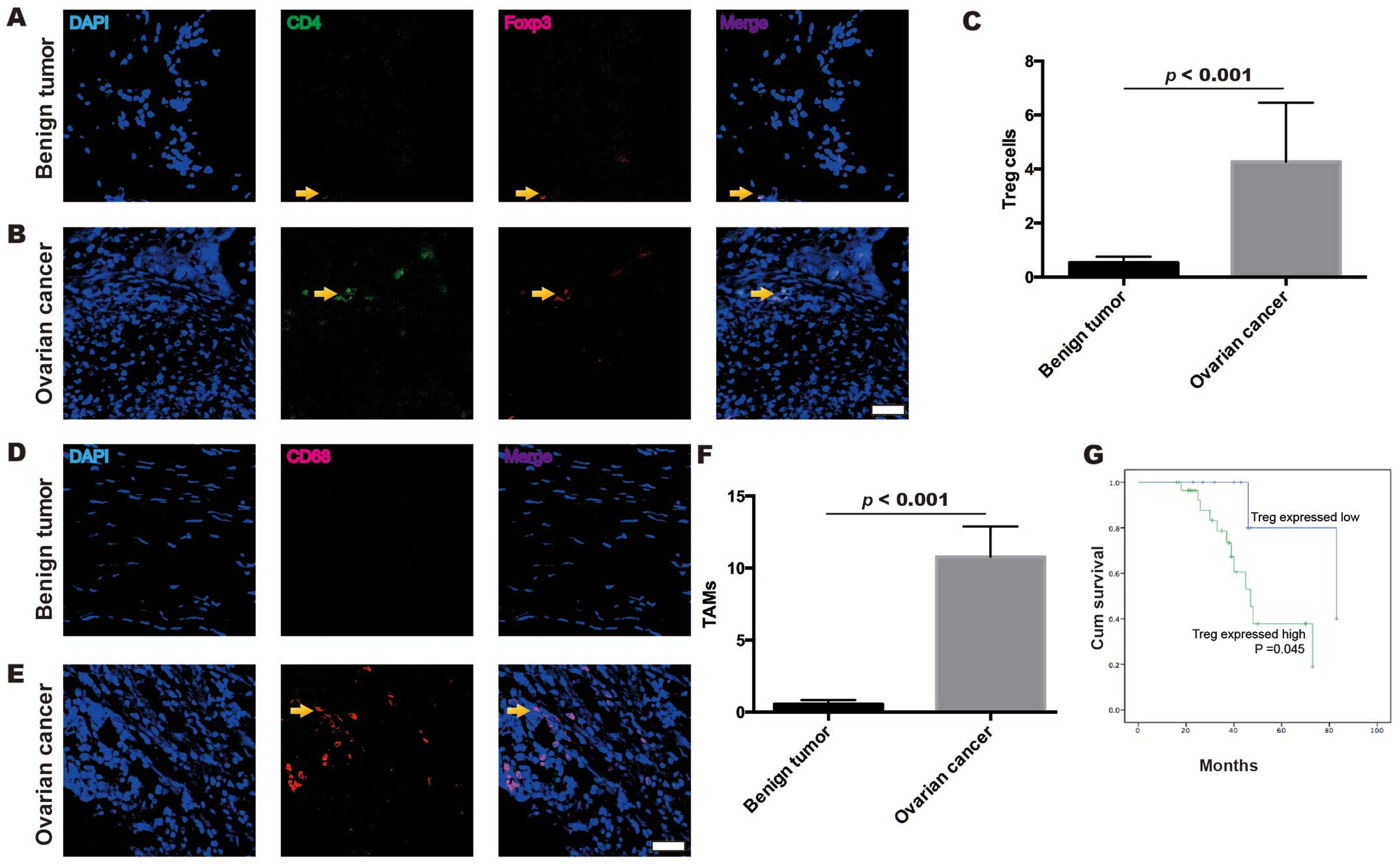
The frequencies of Treg cells and TAMs
in the EOC samples are higher than those in the benign ovarian
tumor samples
A total of 20 tissue samples from benign ovarian
tumors and 40 tissue samples from EOC were analyzed with
immunofluorescence confocal microscopy to evaluate the frequency of
Treg cells (Fig. 1A-C) and TAMs
(Fig. 1D-F). Significant
differences were identified between the frequency of Treg cells and
TAMs in the EOC and benign ovarian tumors. Treg cells were
identified as CD4+ and Foxp3+. The frequency
of Treg cells was confirmed to be higher in the EOC (4.26±2.19)
than that in the benign ovarian tumors (0.53±0.98, p<0.001,
Mann-Whitney U test) (Fig. 1C).
TAMs were identified as CD68+. The frequency of TAMs was
also confirmed to be higher in the EOC (10.79±13.26) than that in
the benign ovarian tumors (0.56±1.25, p<0.001, Mann-Whitney U
test) (Fig. 1F). Moreover, EOC
patients with high frequency of Treg cells exhibited a
significantly shorter overall survival time compared to those with
low frequency of Treg cells (p=0.045, Kaplan-Meier curves)
(Fig. 1G). Thus, these results
demonstrated that the distribution of Treg cells and TAMs was
different in the EOC and benign ovarian tumors.
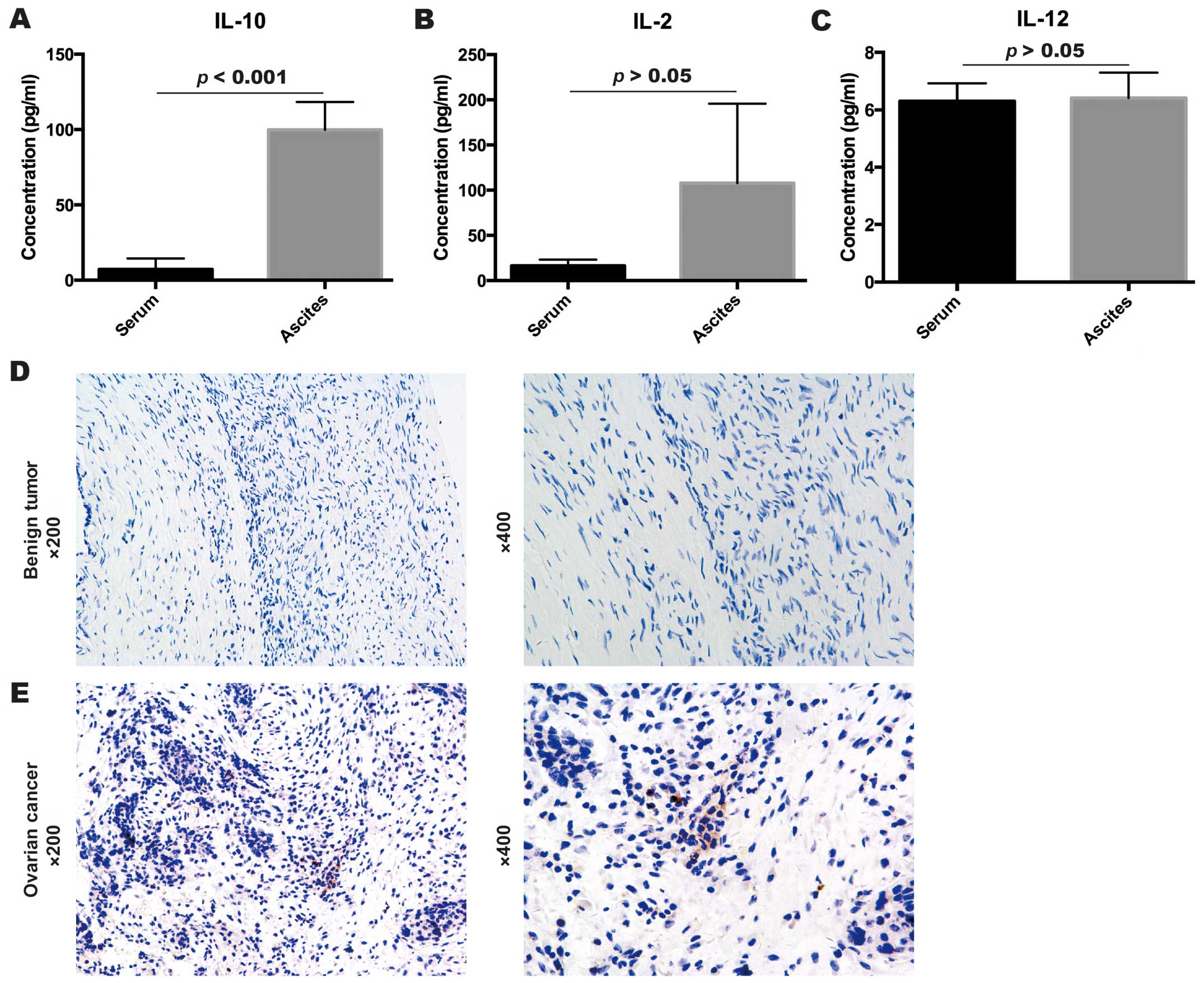
Inflammatory cytokines in ascites and
serum samples
Expression levels of inflammatory cytokines IL-2,
IL-10 and IL-12 in ascites samples of 16 EOC patients and blood
serum samples of 7 healthy subjects were detected by ELISA assays.
As shown in Fig. 2, the expression
level of IL-10 was significantly higher in the ascites of EOC
patients (99.81±71.67 pg/ml) than that in the blood serum of the
healthy subjects (p<0.001, 7.23±7.26 pg/ml, Levenes test)
(Fig. 2A). However, IL-2 and IL-12
expression levels had no significant difference between the ascites
of EOC patients and the serum of the healthy subjects (p>0.05,
IL-2, 107.76±351.55 pg/ml for the ascites vs. 16.38±6.98 pg/ml for
the serum, Mann-Whitney U test; p=0.35, IL-12, 6.41±3.55 pg/ml for
the ascites vs. 6.30±1.68 pg/ml for the serum, Mann-Whitney U test)
(Fig. 2B and C).
Expression of IL-10 in EOC and benign
ovarian tumor samples
To investigate the IL-10 expression in benign
ovarian tumor samples as well as EOC samples, the expression of
IL-10 was evaluated by immunohistochemistry. As shown in Fig. 2, compared to the negative expression
of IL-10 in the benign ovarian tumor samples (Fig. 2D), the EOC samples had strong
staining for IL-10 expression (Fig.
2E). Thus, we supposed that IL-10 may play an important role in
EOC progression.
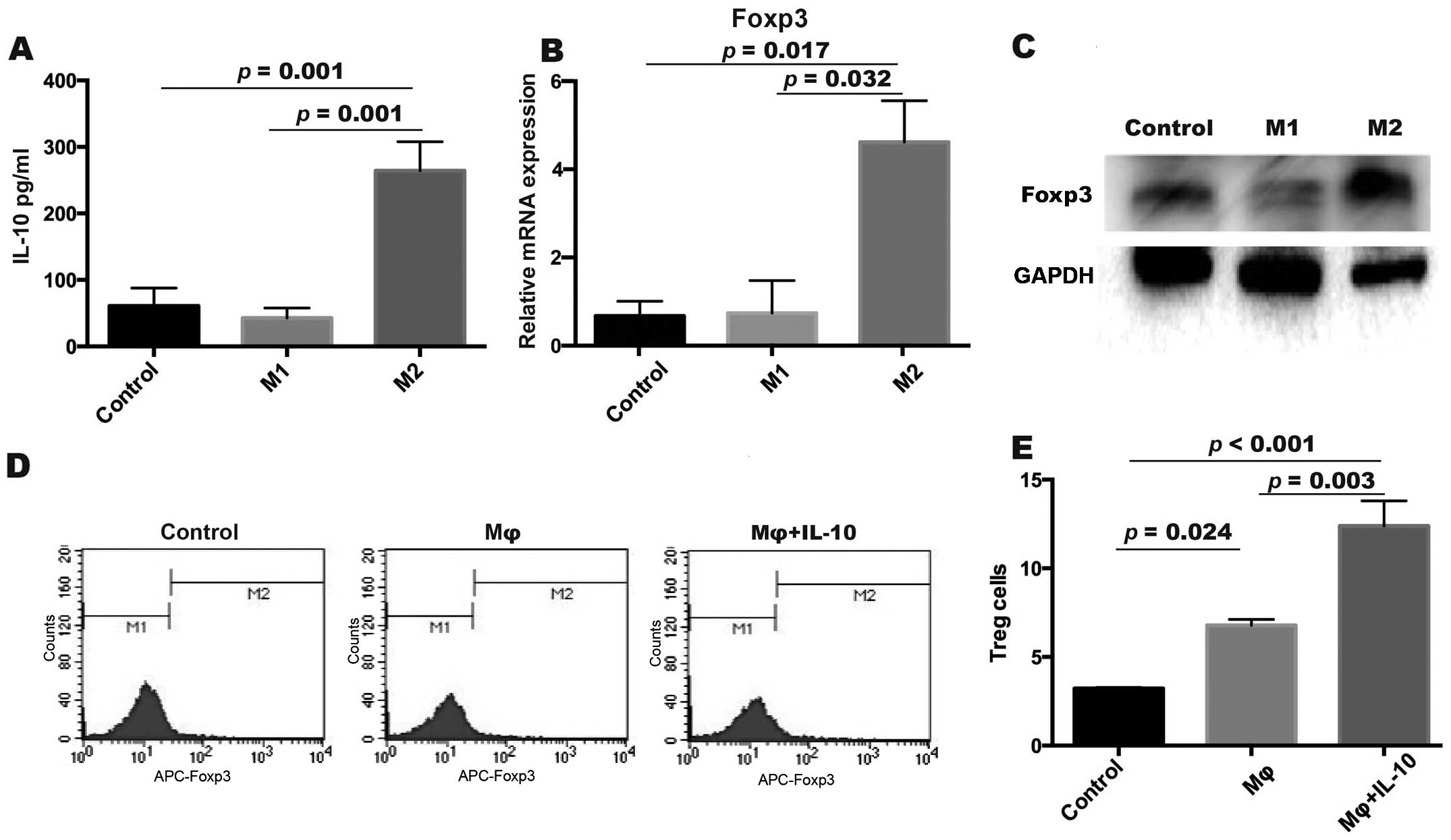
TAMs increase the frequency of Treg
cells by IL-10
TAMs reportedly make up to 70% of immune cells in
the EOC microenvironment (18) and
one feature of TAMs is to secrete IL-10 (19). We induced monocytes into macrophages
(M2, like TAMs) by M-CSF and IL-4 (Fig.
3). The M1 macrophage (M-CSF + LPS + IFN-r) was used as the M1
group. CD4+ T cells were co-cultured with the M1/M2
group. We then detected the IL-10 released in the co-cultured
supernatants, and the results indicated that IL-10 in the M2 group
(264.05±75.91 pg/ml) was significantly higher than that in the
other groups (p=0.001 both for the M1 and control groups, control:
60.89±46.55 pg/ml; M1: 42.71±26.09 pg/ml; LSD test) (Fig. 3A). Next, we detected the expression
of Foxp3 mRNA in CD4+ T cells after being cultured with
M1/M2 macrophages using real-time PCR. We found that the expression
of Foxp3 was higher in the M2 group (4.62±1.63) compared to the M1
group (0.74±1.28, p=0.032, Levenes test) and the control group
(0.67±0.58, p=0.017, Levenes test) (Fig. 3B). The same result was also
demonstrated by western blotting (Fig.
3C). These results suggested that IL-10 could upregulate Foxp3
expression to induce Treg cells during T differentiation.
Interleukin-10 induces the
upregulation of Treg cells
To investigate whether IL-10 plays a significant
role in the differentiation of Treg cells we next cultured
CD4+ T cells with macrophages (Mφ group) and macrophages
with IL-10 (Mφ + IL-10 group) for 3 days. After being co-cultured
with macrophages and IL-10, the proportion of Foxp3+
Treg cells was detected by flow cytometric analysis (Fig. 3D). The frequency of Treg cells was
higher in the Mφ + IL-10 group (12.39±2.45) when compared to the Mφ
group (6.79±0.58, p=0.003, LSD test) and the control group
(3.22±0.12, p<0.001, LSD test) (Fig.
3E). The results showed that IL-10 could increase the frequency
of Treg cells.
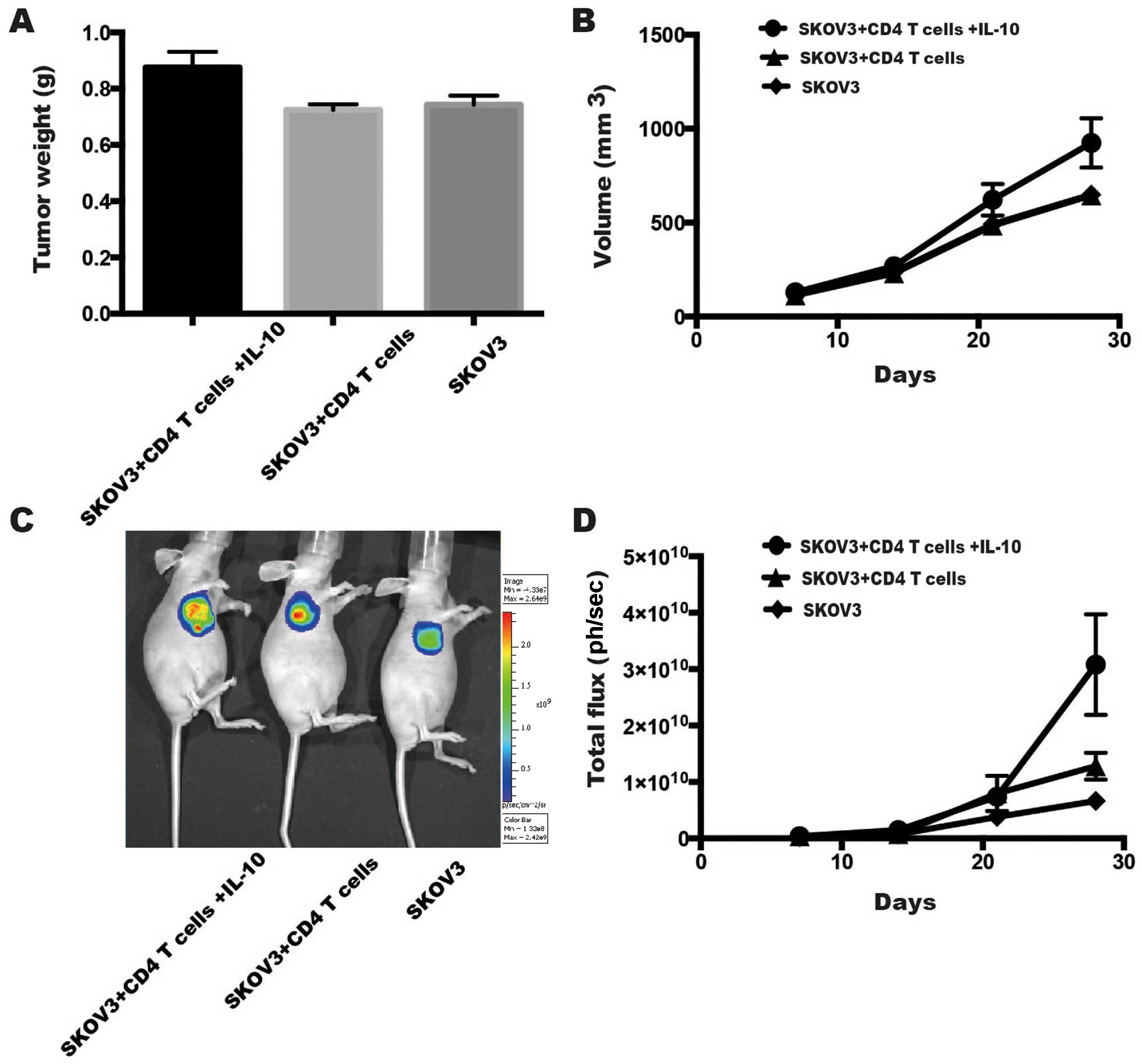
Effect of IL-10 and CD4+ T
cells on the growth of EOC tumors in vivo
To investigate the influence of IL-10 and
CD4+ T cells on EOC development, we established a
subcutaneous nude mouse model to observe EOC tumor growth (Fig. 4). The tumor weight (g) of the
SKOV3-Luc + CD4+ T cells + IL-10 group (0.88±0.06 g) was
much higher than that of the SKOV3-Luc + CD4+ T cell
group (p=0.024, LSD test, 0.72±0.02 g) and the SKOV3 group
(p=0.041, LSD test, 0.74±0.03 g) (Fig.
4A). Moreover, the tumor volume (mm3) of the
SKOV3-Luc + CD4+ T + IL-10 group (924.26±263.80
mm3) was also higher than that of the SKOV3-Luc +
CD4+ T cell group (p=0.032, LSD test, 645.96±41.17
mm3) and the SKOV3 group (p=0.033, LSD test,
649.40±14.02 mm3) on the 4th week (Fig. 4B). Furthermore, after injecting
SKOV3-Luc cells and CD4+ T cells with or without IL-10,
we used a luminescence imaging system to observe tumor growth. The
SKOV3-Luc + CD4+ T cells + IL-10 group
[3.08×1010±8.94×109 photons (ph)/sec] showed
an enhancement of tumor total flux (ph/sec) on the 4th week,
compared to the SKOV3-Luc + CD4+ T cell group (p=0.043,
LSD test, 1.28×1010±2.38×109 ph/sec) and the
SKOV3 group (p=0.012, LSD test,
6.65×109±1.51×109 ph/sec) (Fig. 4C and D).
Discussion
EOC contains malignant cells as well as a number of
TAMs. Different macrophages perform different functions. Therefore,
it is known that monocytes may be recruited by the tumor
microenvironment and acquire an M2 macrophage phenotype, which is
different from an M1 macrophage phenotype. M2 macrophages play a
key role in EOC growth, progression and peritoneal metastasis
through the secretion of cytokines and growth factors, thereby
contributing to tumor angiogenesis and immunosuppression (20). The poor prognosis of carcinomas,
which includes bladder, cervical and breast cancer, may be due to
the macrophage infiltration in tumors (21–23).
Macrophages are a key source of critical cytokine IL-10 (24). We therefore hypothesized that IL-10
secreted by TAMs may increase the frequency of Treg cells. In this
study, we investigated the frequency of TAMs and Treg cells in
human EOC and benign ovarian tumors. TAMs and Treg cells were
observed in patients with EOC more than in subjects with benign
ovarian tumors. In addition, the expression of IL-10 was also
observed to be higher in EOC patients. IL-10 is an
immunosuppressive cytokine, which is secreted by TAMs. It is known
that IL-10 contributes to the suppression of antitumor activity in
the tumor microenvironment. Our results also identified that the
expression level of IL-10 was significantly higher in the ascites
of EOC patients than that in the blood serum of the healthy
subjects. Thus, these data demonstrated that the soluble factor
IL-10 derived from TAMs was responsible for the EOC activity in the
tumor microenvironment and may promote EOC progression.
Treg cells are known to play a vital role in
suppressing antitumor functions. Some studies have shown that
during the early stage of differentiation, Treg cells may require
TGF-β, which is secreted by TAMs, to hamper immunoregulatory
responses (15). Other studies have
demonstrated that Treg cells may mediate immunosuppression by IL-10
in the tumor microenvironment (25–27).
With respect to Treg cells, Islas-Vazquez et al proposed
that IL-10 may participate in the differentiation process of Treg
cells in lung adenocarcinoma (28).
Our results indicated that IL-10 secreted by TAMs may participate
in the differentiation of T cells during development. Thus, IL-10
cytokine that is increased in the tumor microenvironment in EOC
patients promotes the presence of Treg cells and increases in Treg
cells is associated with EOC progression, peritoneal metastasis,
which eventually results in poor prognosis.
Treg cells are a subpopulation of T cells with
immunosuppressive response and express transcription factor Foxp3.
It is reported that Foxp3 plays an important role in regulatory
T-cell function (17).
Nevertheless, Foxp3 has its limited value in the isolation of Treg
cells in function assays, because it is a nuclear protein. Our data
demonstrated that the Foxp3 expression of Treg cells was higher in
the M2 group. Thus, IL-10 detected in this study may correlate with
Foxp3 expression in Treg cells and be involved in immunosuppression
in EOC. This knowledge may lead to the development of immunotherapy
that inhibits the suppressive activity mediated by IL-10 from TAMs.
Further research is required to explore this possibility.
In conclusion, our results helped clarify that IL-10
mediated interaction between TAMs and Treg cells, suggesting that
the targeting of TAMs and their cytokines may be potential targets
for novel immunotherapy in the treatment of ovarian cancer. This
conclusion may have important clinical value for the diagnosis,
therapy and prognosis of EOC patients.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81372787, 81072136), the
Shanghai Municipal Bureau of Health (nos. 20134033 and 20134225),
the Shanghai Pudong Research Project (no. PW2010D-5), the Shanghai
Health System Joint Research Project (no. 2013ZYJB0201) and the Top
100 Medical Elite in Shanghai (no. XBR 2011065).
References
|
1
|
Prat J: Ovarian carcinomas: Five distinct
diseases with different origins, genetic alterations, and
clinicopathological features. Virchows Archiv. 460:237–249. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Salani R and Bristow RE: Surgical
management of epithelial ovarian cancer. Clin Obstet Gynecol.
55:75–95. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Balkwill F, Charles KA and Mantovani A:
Smoldering and polarized inflammation in the initiation and
promotion of malignant disease. Cancer Cell. 7:211–217. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Loberg RD, Ying C, Craig M, Yan L, Snyder
LA and Pienta KJ: CCL2 as an important mediator of prostate cancer
growth in vivo through the regulation of macrophage infiltration.
Neoplasia. 9:556–562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qian BZ and Pollard JW: Macrophage
diversity enhances tumor progression and metastasis. Cell.
141:39–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kurahara H, Shinchi H, Mataki Y, Maemura
K, Noma H, Kubo F, Sakoda M, Ueno S, Natsugoe S and Takao S:
Significance of M2-polarized tumor-associated macrophage in
pancreatic cancer. J Surg Res. 167:e211–e219. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee CH, Espinosa I, Vrijaldenhoven S,
Subramanian S, Montgomery KD, Zhu S, Marinelli RJ, Peterse JL,
Poulin N, Nielsen TO, et al: Prognostic significance of macrophage
infiltration in leiomyosarcomas. Clin Cancer Res. 14:1423–1430.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shabo I, Stal O, Olsson H, Dore S and
Svanvik J: Breast cancer expression of CD163, a macrophage
scavenger receptor, is related to early distant recurrence and
reduced patient survival. Int J Cancer. 123:780–786. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sica A, Saccani A, Bottazzi B,
Polentarutti N, Vecchi A, van Damme J and Mantovani A: Autocrine
production of IL-10 mediates defective IL-12 production and NF-κB
activation in tumor-associated macrophages. J Immunol. 164:762–767.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lv M, Xiaoping X, Cai H, Li D, Wang J, Fu
X, Yu F, Sun M and Lv Z: Cytokines as prognοstic tool in breast
carcinoma. Front Biosci (Landmark Ed). 16:2515–2526. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sato T, Terai M, Tamura Y, Alexeev V,
Mastrangelo MJ and Selvan SR: Interleukin 10 in the tumor
microenvironment: A target for anticancer immunotherapy. Immunol
Res. 51:170–182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu JY, Li F, Wang LP, Chen XF, Wang D,
Cao L, Ping Y, Zhao S, Li B, Thorne SH, et al: CTL- vs
Treg lymphocyte-attracting chemokines, CCL4 and CCL20,
are strong reciprocal predictive markers for survival of patients
with oesophageal squamous cell carcinoma. Br J Cancer. 113:747–755.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang CY, Qi Y, Li XN, Yang Y, Liu DL,
Zhao J, Zhu DY, Wu K, Zhou XD and Zhao S: The role of CCL20/CCR6
axis in recruiting Treg cells to tumor sites of NSCLC patients.
Biomed Pharmacother. 69:242–248. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shen Y, Wei Y, Wang Z, Jing Y, He H, Yuan
J, Li R, Zhao Q, Wei L, Yang T, et al: TGF-β regulates
hepatocellular carcinoma progression by inducing Treg cell
polarization. Cell Physiol Biochem. 35:1623–1632. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma L, Zhang H, Hu K, Lv G, Fu Y, Ayana DA,
Zhao P and Jiang Y: The imbalance between Tregs, Th17 cells and
inflammatory cytokines among renal transplant recipients. BMC
Immunol. 16:562015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Peterson RA: Regulatory T-cells: Diverse
phenotypes integral to immune homeostasis and suppression. Toxicol
Pathol. 40:186–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shevach EM and Thornton AM: tTregs,
pTregs, and iTregs: Similarities and differences. Immunol Rev.
259:88–102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Deavers M, Patenia R, Bassett RL
Jr, Mueller P, Ma Q, Wang E and Freedman RS: Monocyte/macrophage
and T-cell infiltrates in peritoneum of patients with ovarian
cancer or benign pelvic disease. J Transl Med. 4:302006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang C, He L, He P, Liu Y, Wang W, He Y,
Du Y and Gao F: Increased drug resistance in breast cancer by
tumor-associated macrophages through IL-10/STAT3/bcl-2 signaling
pathway. Med Oncol. 32:3522015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mantovani A and Sica A: Macrophages,
innate immunity and cancer: Balance, tolerance, and diversity. Curr
Opin Immunol. 22:231–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hanada T, Nakagawa M, Emoto A, Nomura T,
Nasu N and Nomura Y: Prognostic value of tumor-associated
macrophage count in human bladder cancer. Int J Urol. 7:263–269.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fujimoto J, Sakaguchi H, Aoki I and Tamaya
T: Clinical implications of expression of interleukin 8 related to
angiogenesis in uterine cervical cancers. Cancer Res. 60:2632–2635.
2000.PubMed/NCBI
|
|
23
|
Mukhtar RA, Nseyo O, Campbell MJ and
Esserman LJ: Tumor-associated macrophages in breast cancer as
potential biomarkers for new treatments and diagnostics. Expert Rev
Mol Diagn. 11:91–100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang R, Lu M, Zhang J, Chen S, Luo X, Qin
Y and Chen H: Increased IL-10 mRNA expression in tumor-associated
macrophage correlated with late stage of lung cancer. J Exp Clin
Cancer Res. 30:622011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schmidt A, Oberle N and Krammer PH:
Molecular mechanisms of Treg-mediated T cell suppression. Front
Immunol. 3:512012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao L, Yang J, Wang HP and Liu RY:
Imbalance in the Th17/Treg and cytokine environment in peripheral
blood of patients with adenocarcinoma and squamous cell carcinoma.
Med Oncol. 30:4612013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shigematsu Y, Hanagiri T, Shiota H, Kuroda
K, Baba T, Ichiki Y, Yasuda M, Uramoto H, Takenoyama M, Yasumoto K,
et al: Immunosuppressive effect of regulatory T lymphocytes in lung
cancer, with special reference to their effects on the induction of
autologous tumor-specific cytotoxic T lymphocytes. Oncol Lett.
4:625–630. 2012.PubMed/NCBI
|
|
28
|
Islas-Vazquez L, Prado-Garcia H,
Aguilar-Cazares D, Meneses-Flores M, Galicia-Velasco M,
Romero-Garcia S, Camacho-Mendoza C and Lopez-Gonzalez JS: LAP
TGF-beta subset of CD4+CD25+CD127 Treg cells
is increased and overexpresses LAP TGF-beta in lung adenocarcinoma
patients. BioMed Res Int. 2015:4309432015. View Article : Google Scholar : PubMed/NCBI
|