Introduction
Colon cancer, a highly malignant tumor, is the 3rd
most common cause of cancer-related death according to global
epidemiological data and the prevalence is increasing in the face
of better nutrition and improved living conditions (1). Traditional therapeutic methods for
treating colon cancer include surgical tumor removal, chemotherapy
and radiotherapy, but these are limited by tumor recurrence and
significant side-effects. Specifically, 5-year survival of hepatic
metastases from large bowel carcinomas with surgical treatment is
less than 30% (2), thus, better
therapies or approaches are needed urgently. Investigations of
signaling molecules that are expressed in tumors may be a promising
avenue.
Gracilaria lemaneiformis (kelp referred to as
Nostoc), is a Gracilaria species (gigartinales rhodophyta) and a
rich source of polysaccharides, protein, vitamins and multiple
minerals such as phosphorus, calcium, iodine, iron, zinc and
magnesium (3). Phycoerythrin (PE)
from G. lemaneiformis is an important light-harvesting
protein with potential use in the food and drug industries with
potential antioxidant, immunomodulatory, antitumor,
radiation-resistant, anti-anemic and anti-aging properties as well
as few side-effects (4). Chen et
al (5) demonstrated that PE
facilitated macrophage phagocytic activity in tumor-bearing mice
and enhanced non-specific immunity. Gao et al (6) revealed that PE inhibited human breast
cancer MCF-7 cells and induced apoptosis related to cell cycle
arrest but the antitumor effects were complex and the molecular
mechanism for this outcome remains unclear.
Genomics permits the study of tumorigenesis at the
molecular level and proteomics allows a focus on expression and
function of tumor proteins (7) for
screening and identifying tumor biomarkers (8,9),
classifying tumors (10), and for
developing drugs as well as assessing mechanisms of oncogenesis
(11). Thus, with proteomic
technology and bioinformatic analysis we investigated molecular
mechanisms of PE inhibition of SW480 cells and identified the
proteins associated with proliferation and inhibition. These data
will offer a theoretical foundation for future development of
cancer therapeutics.
Materials and methods
Cell strain
The human colon cancer SW480 cell line was purchased
from the Cell Bank at the Chinese Academy of Sciences (Shanghai,
China). Cells were cultured in RPMI-1640 containing 10% fetal
bovine serum (FBS) in an incubator with 5.0% CO2 at
37°C.
Reagents
PE was extracted from fresh Gracilaria
lemaneiformis (12) and the
purity of PE was 5.37 (A565/280 nm). RPMI-1640 medium
and trypsin were products of Gibco (Waltham, MA, USA). FBS was
purchased from Hangzhou Chinese Holly Biotechnology Ltd. (Hangzhou,
China). CCK-8 was a product of Sigma-Aldrich (St. Louis, MO, USA).
An Annexin V/PI staining kit was purchased from Lianke Biotechno
Co., Ltd., (Hangzhou, China). Cell culture plates were obtained
from Corning Costar Corp. (Cambridge, MA, USA). Reverse
transcription and PCR kits were products of Takara Biotechnology
Ltd. (Dalian, China). Primary and secondary antibodies were from
Beyotime Institute of Biotechnology (Shanghai, China).
CCK-8 testing inhibition of PE on
SW480 growth
Cells (5×104/ml) were inoculated into
96-well culture plates (100 µl each well) and 24 h later,
supernatant was removed and PE medium was added at 6 different
concentrations (0, 5, 10, 20, 40 and 80 µg/ml) in sextuplicate.
Cisplatin was a positive control. After incubation CCK-8 was added
(20 µl/well) for another 4 h. Absorbance was read at 490 nm and
IC50 values for PE were calculated (R =
(Acontrol - Apositive)/Acontrol ×
100%) using the Logit method.
Microscopic observation
Polylysine-preprocessed coverslips were seeded in
6-well plates and 1.5 ml cell suspension (1.5×104/ml)
was added to each well. Next, 24 h later, cells were treated with
PE (0, 20, 40 and 80 µg/ml) for 48 h. Some samples were examined
under an inverted microscope, and others were collected, rinsed
three times with 0.01 mol/l PBS (pH 7.0), followed by fixation with
2.5% glutaraldehyde and 1% osmic acid. Ethanol was used for
dehydration. After drying under CO2 and ion sputtering
and metal spraying, samples were visualized under a scanning
electron microscope (Hitachi S-3000N; Hitachi, Ltd., Tokyo, Japan)
(13).
FCM measurement of apoptosis
Cells were seeded (5×104/ml) into 25-ml
culture flasks for 24 h and were treated with PE at different
concentrations (0, 10, 20, 40 and 80 µg/ml, respectively) for 48 h.
According to the Annexin V/PI kit instructions, primary medium was
removed and pancreatin solution without EDTA was added for
digestion which was terminated by adding medium. After
centrifugation, 0.5 ml PBS (10 mmol/l, pH 7.0), 10 ml media with
binding reagent, 1.25 µl FITC (200 µg/ml) and 10 µl PI (50 µg/ml)
were added into each tube successively. The reaction was at room
temperature and in the dark for 10 min and flow cytometry (BD
Biosciences, San Jose, CA USA) was used to measure cells and data
were analyzed using WinMDI 3.0 (14). All experiments were performed in
triplicate.
FCM cell cycle determination
Cell samples were prepared and analyzed as described
above. Cells treated with PE for 48 h were collected and fixed with
pre-cooled 70% ethanol overnight at 4°C and washed with 0.01 mol/l
PBS three times. After permeabilization, cells were treated with
RNAse (final concentration, 10 µg/ml) for 1 h at 37°C and then
stained with PI (final concentration, 50 µg/ml). The reaction was
incubated at 37°C in the dark for 30 min and flow cytometry was
used to measure cell cycle arrest. Data were analyzed using BD
FACSDiva software (14). All
experiments were repeated three times.
Proteomics for analyzing protein
Cell lysis buffer (200 µl, 7 mol/l Urea, 2 mol/l
thiourea, l4% CHAPS, 40 mmol/l Tris, 65 mmol/l DTT, 5 mmol/l PMSF,
1 mg/ml DNaseI and 0.25 mg/ml RNase A) was added to
1.5×106 cells and incubated for 30 min with intermittent
swirling. The solution was centrifuged at 15,000 rpm for 30 min at
4°C and supernatant was collected. Total protein was quantified
with a BCA protein assay kit (15)
and stored at −80°C.
First-dimension of analysis with isoelectric
focusing electrophoresis was used on 200 µg samples mixed with
loading buffer (7 mol/l urea, 2 mol/l thiourea, 4% CHAPS, 65 mmol/l
DTT, 0.2% BioLyte and 0.001% bromophenol blue). IPG glue solid
state strips were used as carriers. The maximum current was 60
µA/IPG strip, which were hydrated and focused at 20°C. Isoelectric
focusing was performed under 8,000 V (total voltage-time of 30,000
Vh). Focused tapes were put into equilibration buffer (6 mol/l
urea, 2% SDS, 50 mmol/l Tris-HCl, pH 8.8 and 20% glycerol)
containing 1% DTT and 4% iodoacetamide for 15 min. Then, tapes were
transferred to 12% PAGE covered with agarose. 2D analysis SDS-PAGE
was electrophoresed. The current (10 mA/gel) was maintained for 15
min and increased to 30 mA/gel until bromophenol blue reached the
bottom. After silver staining (16), GS-800 transmission and scans were
used to collect and analyze gel images (resolution 63.5×63.5). For
protein identification by MS, protein dot matching was performed
between analytical silver stained gels and preparative gels to
correlate the precise spot position to be excised. Protein spots
were compared by image-matching analysis using PDQuest 7.4.0.
Mass spectrum identification of
differential protein spots
Differential spots were excised from gels and placed
into 1.5-ml tubes, washed with Milli-Q water three times followed
by addition of destaining solution (15 mmol/l potassium
ferricyanide and 50 mmol/l sodium thiosulfate were premixed at a
volume ratio of 1:1) and samples were incubated for 20 min. After
two additional washes with Milli-Q water, pellets were immersed in
acetonitrile and dehydrated. Acetonitrile was discarded and trypsin
digestion (12.5 mg/l, performed with 20 mmol/l ammonium
bicarbonate) was added and incubated at 4°C. Then the gel was kept
at 37°C for 18 h for enzymatic hydrolysis. Enzymatic reaction
liquid was collected and the gel was washed with 0.1% TFA/50%
acetonitrile mixture twice with slight agitation to extract peptide
fragments. Extraction liquid was dried under nitrogen and
redissolved in 0.7 µl 5.0 g/l CHCA matrix (dissolved in 50%
acetonitrile and 0.1% TFA). Samples were loaded on a stainless
steel 192-well target plate and air-dried. Peptide mass
fingerprints were obtained with MALDI-TOF-M (Bruker Daltonics,
Inc., Billerica, MA, USA) and precision peaks quality was corrected
according to automatic switching peaks of trypsin as an interior
reference. Mass spectrometric data were retrieved using Mascot
software (www.matrixscience.co.uk) in Swiss-Prot and NCBI nr
database (retrieval parameters: trypsin digestion, permissible
error ± 0.2 Da, oxidative modification of methionine, iodoacetamide
modification of cysteine) (17).
qPCR technique identifying mRNA
expression of differential proteins
The primer sets (Table
I) were designed for amplification of β-actin and the expressed
genes using the Premier 5.0 software and synthesized by Shanghai
Sunny Biological Technology Co, Ltd. (Shanghai, China). Total RNA
was extracted from PE-treated and untreated SW480 cells with Dnase
I by RNase-Free Dnase Set (Qiagen, Hilden, Germany). cDNA was
obtained by reverse transcription using PrimeScript RT-PCR kit
(Takara). SYBR-Green I-based qPCR was performed according to
routine protocols (18) using the
ABI StepOnePlus Real-Time PCR system (Applied Biosystems, Carlsbad,
CA, USA) and three replicates were performed for all genes in each
sample. Relative changes of gene expression were measured using the
2−ΔΔCt method. Relative quantification of targets in
each sample was carried out using β-actin as a control.
 | Table I.Primer sequences and product
sizes. |
Table I.
Primer sequences and product
sizes.
| Gene symbol | Nucleotide
sequence | Product size
(bp) | Temperature
(°C) |
|---|
| GRP78 | Forward:
GGGCCCTGTCTTCTCAACAT | 210 | 59 |
|
| Reverse:
GAGTCGAGCCACCAACAAGA |
|
|
| NPM1 | Forward:
GGAGGTGGTAGCAAGGTTCC | 149 | 60 |
|
| Reverse:
GATTTCTTCACTGGCGCTTTT |
|
|
| Annexin
A2 | Forward:
GGACGCGAGATAAGGTCCTG | 145 | 59 |
|
| Reverse:
GCTTTCTGGTAGTCGCCCTT |
|
|
| Ezrin | Forward:
CGCTCTAAGGGTTCTGCTCTG | 114 | 60 |
|
| Reverse:
TTGGTTTCGGCATTTTCGGT |
|
|
| HSP60 | Forward:
ACAAGAACATTGGAGCTAAACTTGT | 104 | 59 |
|
| Reverse:
TTGGCTATAGAGCGTGCCAG |
|
|
| MTHSP75 | Forward:
AGGGAGCTCCTGGCTAGAAA | 144 | 59 |
|
| Reverse:
GCCAGAACTTCCAGAGCCTT |
|
|
| p53 | Forward:
GAGCACTGCCCAACAACAC | 224 | 58 |
|
| Reverse:
ATGGCGGGAGGTAGACTGA |
|
|
|
Caspase-3 | Forward:
ATGGAAGCGAATCAATGGAC | 242 | 56 |
|
| Reverse:
ATCACGCATCAATTCCACAA |
|
|
|
Caspase-9 | Forward:
GCGAACTAACAGGCAAGCA | 144 | 58 |
|
| Reverse:
CCAAATCCTCCAGAACCAAT |
|
|
| Bcl-2 | Forward:
GTGGAGGAGCTCTTCAGGGA | 304 | 56 |
|
| Reverse:
AGGCACCCAGGGTGATGCAA |
|
|
| β-actin | Forward:
CTTCCAGCCTTCCTTCCTGG | 110 | 60 |
|
| Reverse:
CTGTGTTGGCGTACAGGTCT |
|
|
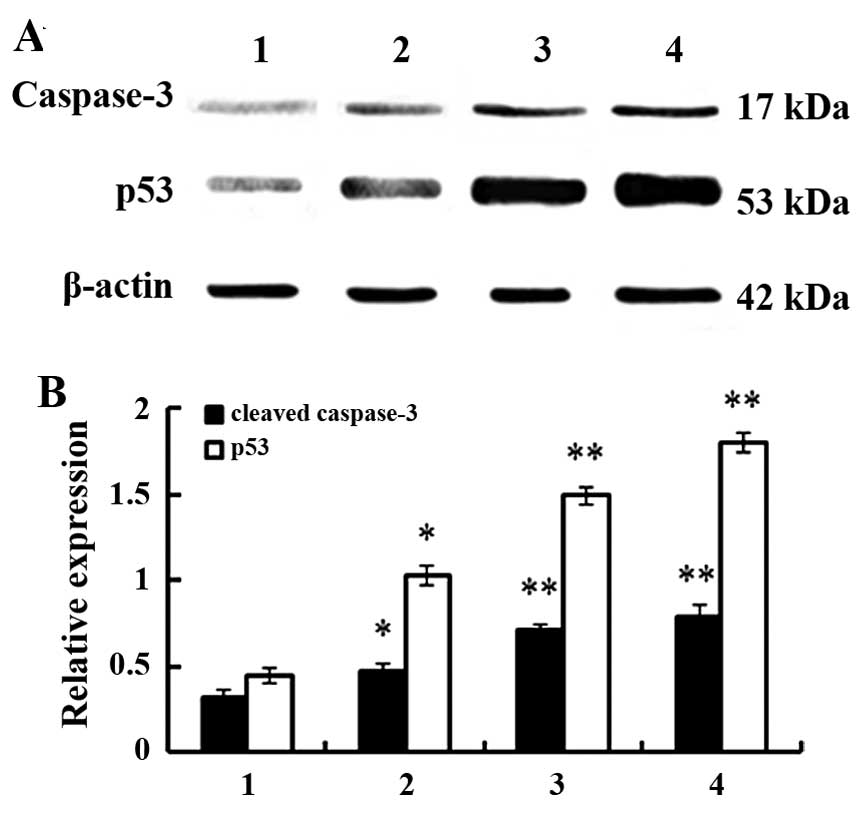
Western blotting testing
Samples were mixed with Laemmli's loading buffer,
boiled for 5 min, and subjected to 12% SDS-PAGE at 120 V followed
by electroblotting to nitrocellulose membrane for 2 h at 80 V.
Membranes were blocked for 1 h with 5% skim milk in TBS at room
temperature and subsequently probed overnight with anti-p53,
anti-caspase-3 and anti-β-actin. The membranes were rinsed and
incubated with an HRP-conjugated secondary antibody. Following the
secondary antibody incubation, the membranes were rinsed and bound
antibodies were detected using enhanced chemiluminescence according
to the manufacturer's instructions. The protein bands were
quantified by densitometry analysis.
Statistical methods
The SPSS 16.0 software was used to carry out
statistical analysis and data are means ± SD. A Students t-test was
used to assess differences which were considered statistically
significant at P<0.05.
Results
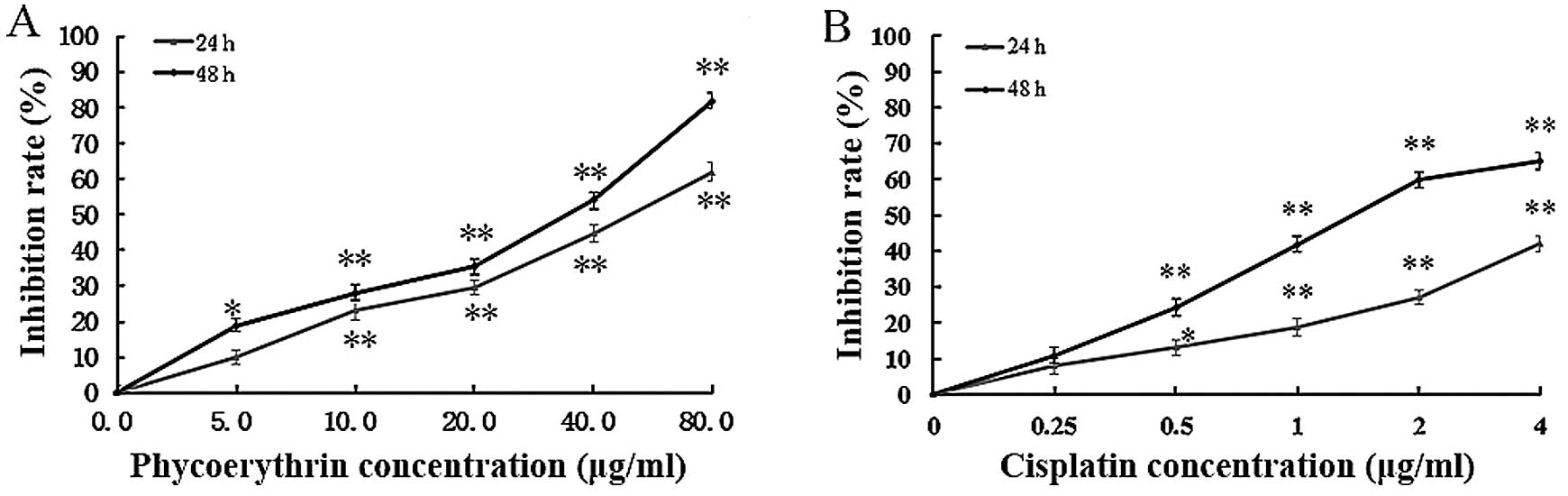
PE inhibits SW480 cell growth
PE inhibited SW480 cell growth in a dose- and
time-dependent manner (Fig. 1A). In
addition, IC50 values of PE for 24 and 48 h of exposures
were 48.2 and 27.4 µg/ml, respectively. PE inhibited cell growth at
80 µg/ml for 24 h and data for beyond 48 h with 40 µg/ml were
similar to those of positive controls (Fig. 1B).
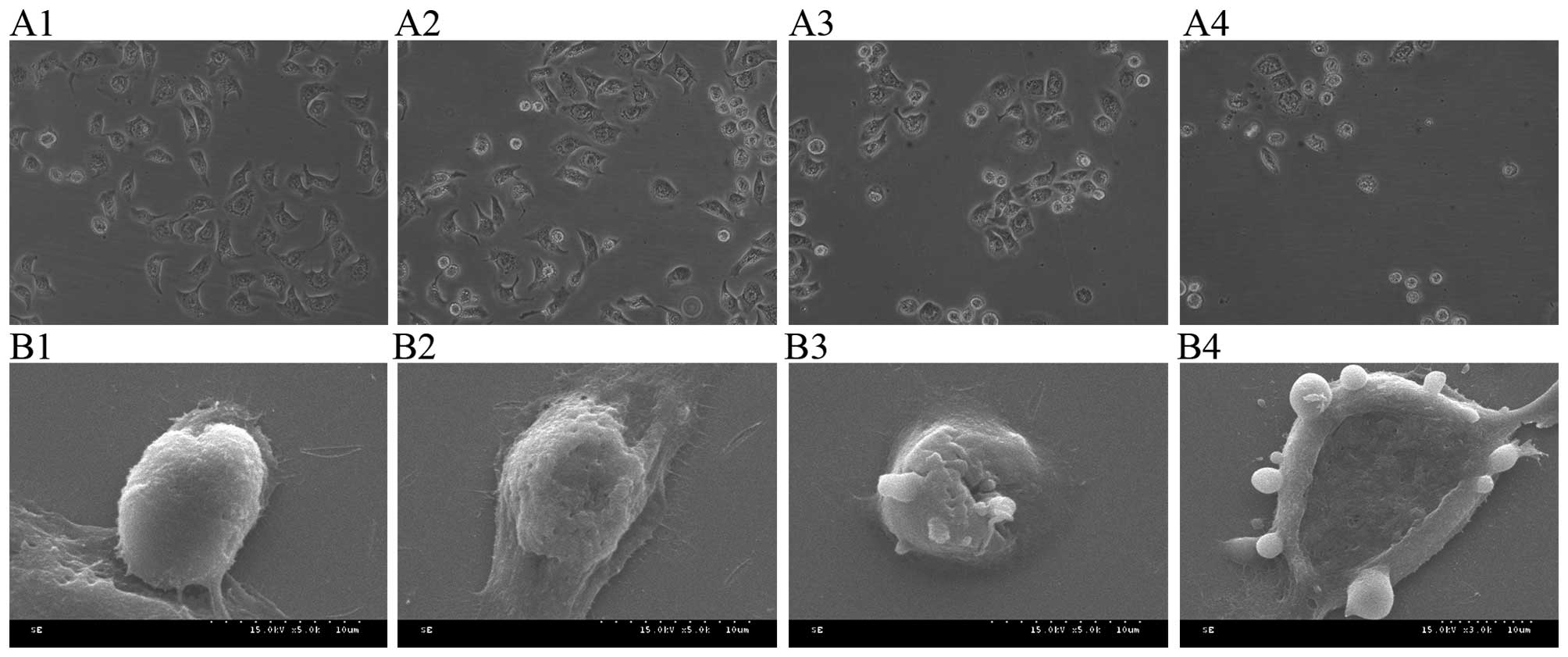
Morphological effect of PE on SW480
cells
Microscopic data indicate that control SW480 cells
had spindle shapes, adhered to the cell wall and tight junctions
and were partially overlapped. Nuclei were large and the nucleolus
was clear and dark. Increasing PE concentration caused cell
shrinkage, loosening of cell junctions and some cell detachment and
floating. In addition, nuclei were shrunk and some were broken
(Fig. 2A). Control SW480 cells had
denser and bigger microvilli on the surface, radially stretching
outward and regularly arranged as observed under a scanning
electron microscope. Increasing PE concentrations caused shortening
of microvilli and cell morphology was abnormal and damaged. After
treatment with 40 µg/ml PE cell shrinkage was obvious, and many
apoptotic bodies budded from the cell membrane (Fig. 2B).
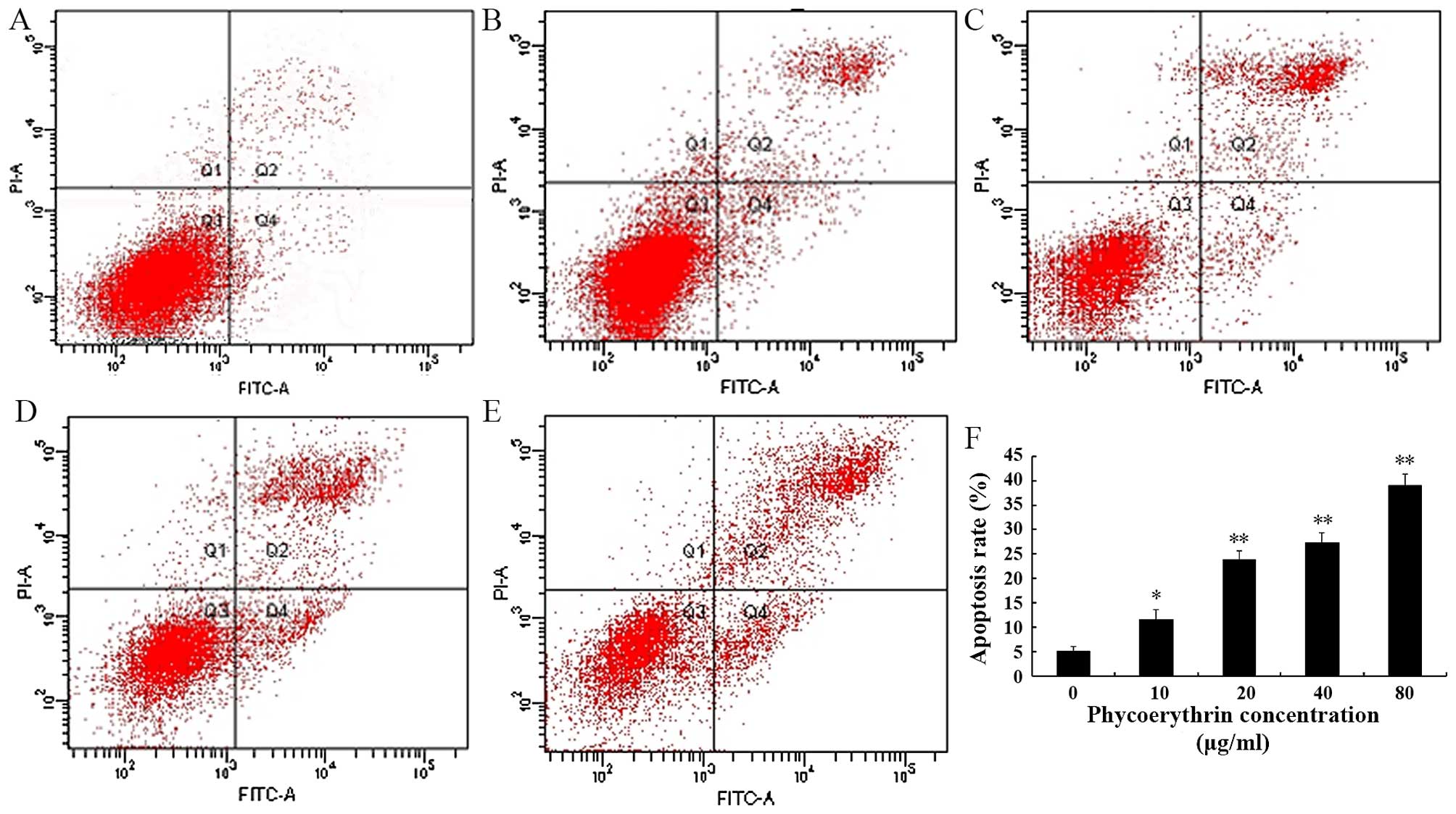
PE-induced apoptosis in SW480
cells
The results of the Annexin V-FITC/PI staining showed
that, with the increase of the PE concentration, the apoptosis rate
correspondingly increased (Fig. 3).
The early stage apoptosis rate increased from 3.8 to 29.0% while in
the later stage it increased from 3.3 to 10.0%.
PE affects cell cycle distribution in SW480 cells.
After PE treatment for 48 h, FCM was used to analyze the cell
cycle. Table II shows that most
cells were arrested in the G0/G1 phase and
cells were arrested in the S stage in controls. PE treatment
increased cells arrested in G2/M phases. Flow cytometry
confirmed that PE induced G2/M cell cycle arrest in
SW480 cells and this was concentration-dependent. PE-induced
apoptosis may be related to cell cycle inhibition at
G2/M stages.
| Table II.Effect of PE on SW480 cell cycle. |
Table II.
Effect of PE on SW480 cell cycle.
| Group |
G0/G1 (%) | S (%) | G2/M
(%) |
|---|
| Control | 91.04±0.03 | 8.83±0.05 | 0.13±0.03 |
| 10 µg/ml PE | 69.45±0.04 | 22.92±0.04 |
7.63±0.06a |
| 20 µg/ml PE | 69.52±0.05 | 20.06±0.09 |
10.41±0.12a |
| 40 µg/ml PE | 66.34±0.04 | 20.20±0.08 |
13.46±0.12a |
| 80 µg/ml PE | 47.66±0.07 | 23.39±0.06 |
28.95±0.09b |
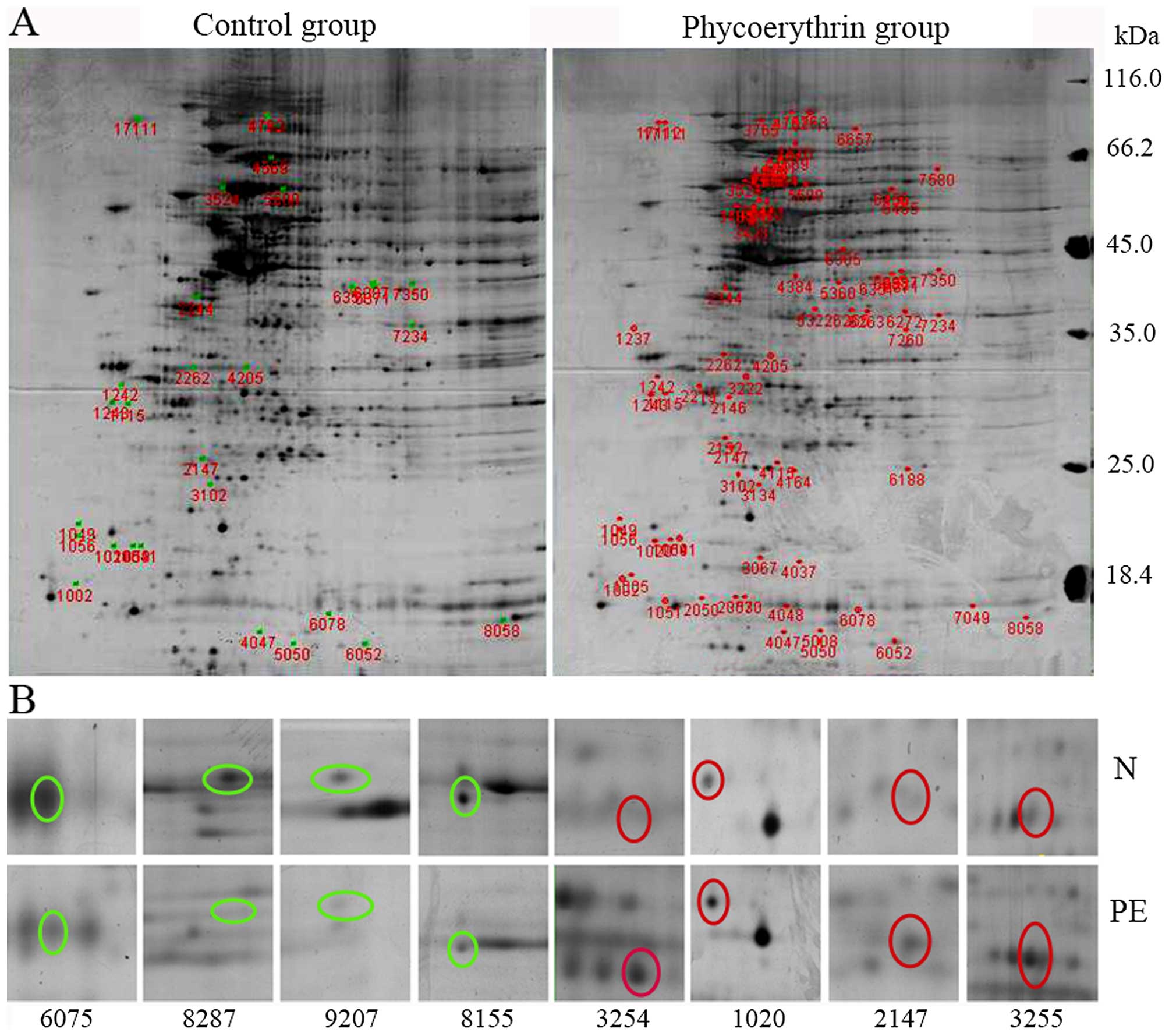
DE atlas analysis
Total proteins of control and PE-treated group were
subjected to 2DE analysis and data show that 3 gels from controls
generated an average of 1010±60 protein dots whereas 3 gels from
the treatment group had 1018±60 dots and matching score was 99%.
Comparing two groups of proteins with expression that was 2-fold
more or less were consistent with the data from the gels and these
proteins were selected for additional analysis. A total of 379
proteins were identified (Fig. 4A),
including 188 upregulated and 191 downregulated proteins. Of these
40 protein dots were chosen for MS analysis (Fig. 4B). Fifteen of their positive protein
spots (Table III) are implicated
in apoptosis, oxidative stress, lipid and ion transport and tumor
cell growth. To validate the MS data, qPCR and western blotting
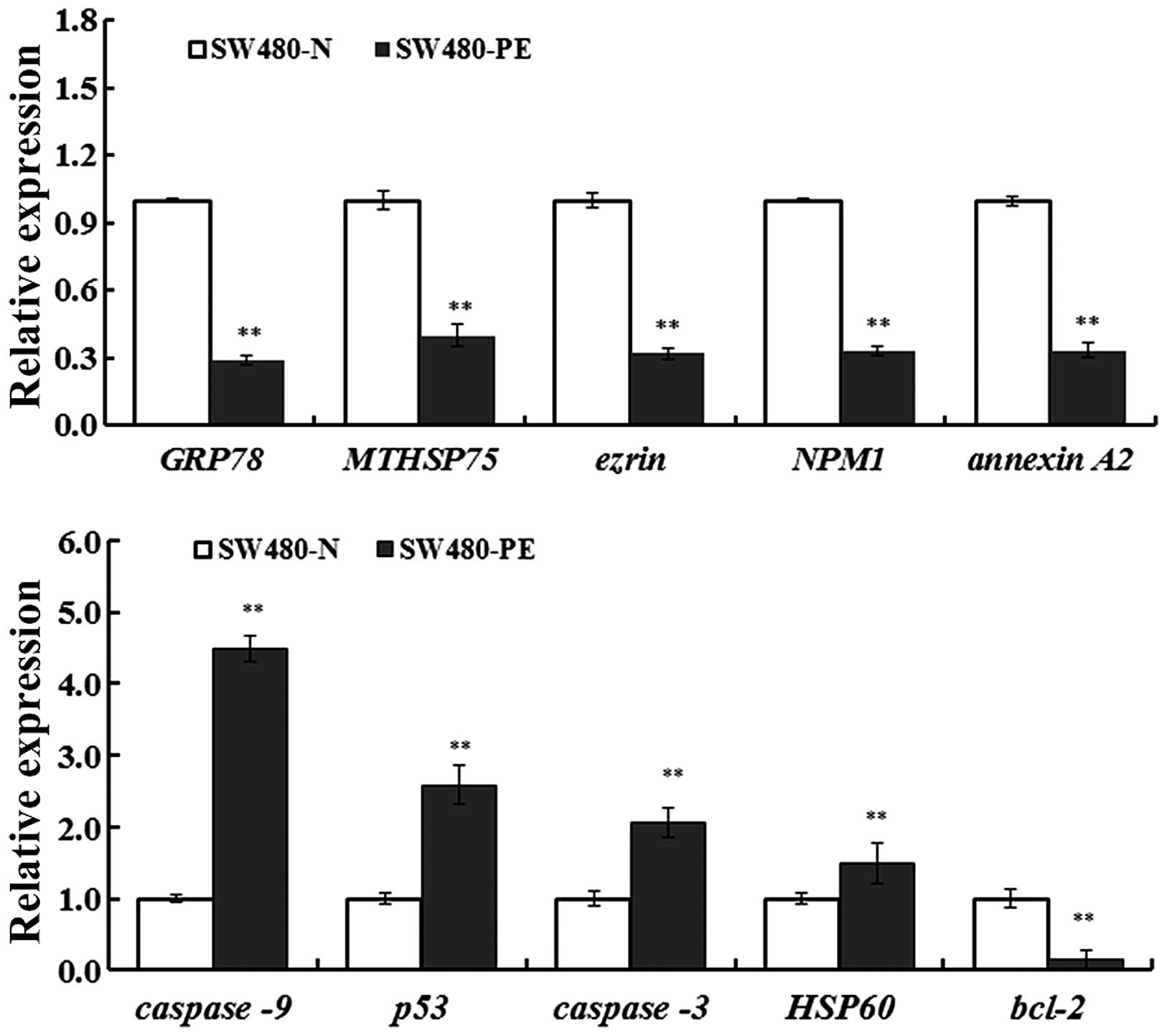
were used to analyze effects of PE on the expression of GRP78,
MTHSP75, HSP60, NPM1, Ezrin, Annexin A2, p53, caspase-3 and Bcl-2
(Figs. 5 and 6). Compared with controls, differences
were statistically significant (P<0.05 or P<0.01) and these
data agreed with protein analyses.
| Table III.Protein spot data identified by
MS. |
Table III.
Protein spot data identified by
MS.
| No. | Spot no. | Match to | Protein | Relative
expression | Match | Sequence
coverage | Score | PI | Mr |
|---|
| 1 | 2655 | gi|16507237 | 78 kDa
glucose-regulated protein precursor (GRP78) | $ | 396 (328) | 53% | 8355 | 5.07 | 72402 |
| 2 | 2531 | gi|21961605 | Keratin 10 | $ | 27 (16) | 36% | 536 | 5.09 | 59020 |
| 3 | 3330 | gi|10835063 | Nucleophosmin
isoform 1 (NPM1) | $ | 68 (49) | 37% | 1645 | 4.64 | 32726 |
| 4 | 4373 | gi|119607256 | hCG1988300, isoform
CRA_a | $ | 98 (89) | 41% | 2895 | 5.09 | 47241 |
| 5 | 4485 | gi|181573 | Cytokeratin 8
(CK8) | $ | 266 (226) | 59% | 5414 | 5.52 | 53529 |
| 6 | 5164 | gi|1568551 | Histone (H2B) | $ | 27 (53) | 26% | 1555 | 10.31 | 13928 |
| 7 | 6082 | gi|435476 | Cytokeratin 9 | $ | 32 (28) | 17% | 926 | 5.19 | 62320 |
| 8 | 6302 | gi|4757756 | Annexin A2 isoform
2 | $ | 104 (87) | 61% | 2523 | 7.57 | 38808 |
| 9 | 6379 | gi|28317 | Unnamed protein
product | $ | 19 (18) | 4% | 648 | 5.17 | 59720 |
| 10 | 6658 | gi|21614499 | Ezrin | $ | 152 (98) | 41% | 1920 | 5.94 | 69484 |
| 11 | 17111 | gi|182855 | 80K-H protein | # | 21 (21) | 8% | 320 | 4.34 | 60228 |
| 12 | 3524 | gi|31542947 | 60 kDa heat shock
protein, mitochondrial HSP60 | # | 123 (107) | 46% | 3440 | 5.7 | 61187 |
| 13 | 4762 | gi|292059 | MTHSP75 | $ | 167 (138) | 58% | 3974 | 5.97 | 74019 |
| 14 | 2344 | gi|306875 | C protein | # | 87 (70) | 28% | 2315 | 5.1 | 32004 |
| 15 | 1041 | gi|7331218 | Keratin 1 | # | 16 (16) | 0.05 | 453 | 8.16 | 66149 |
Discussion
PE, a phycobiliprotein found in plants such as red
alga and G. lemaneiformis, is the product of covalent
binding of apoprotein and an open-chain tetrapyrrole chromophore
via the formation of thioether and it fluoresces orange (19). Its antitumor effects are of interest
for treating tumors as a photosensitizer or a fluorescent probe but
few studies suggest any direct cancer inhibition (20). Recent reports of antitumor effects
of PE indicate that PE significantly suppresses growth of human
cervical cancer HeLa cells and this is dose-dependent. With PE
treatment, HeLa cells were arrested at the G2/M phase
and cell proliferation was inhibited and apoptosis occurred
(21) but how this happened was
unclear. Previous studies suggest that PE induced cell apoptosis
perhaps by reducing mitochondrial membrane potentials and damaging
their integrity (22), which was
essential for live cancer cells. Also, PE caused cell cycle arrest
and apoptosis by upregulating expression of cyclin dependent kinase
CDC25A which increased cyclin at G1 and S stages and
indirectly downregulated its binding protein, CDK2 (23). With proteomics, we screened and
identified 15 proteins related to PE inhibition of colon cancer,
including metabolism- and apoptosis-related proteins, a
cytoskeletal and a carrier protein and another protein of unknown
function. Thus, 6 proliferation-associated protein proteins had
modified expression and most were related to apoptosis, or signal
pathway interference.
Heat shock proteins (HSPs) have highly conserved
structures and are common to all biological cells as they
participate in cell growth and metabolism. HSPs exist as HSP70, 90,
60 and micromolecular HSP (24).
Among the Hsp70 family are cytoplasmic HSP70, mitochondria HSP75
and endoplasmic reticulum (ER) glucose regulatory protein GRP78 or
immune globulin heavy chain binding protein, which can promote
correct protein folding to maintain normal ER function of and
interaction with enzyme 1 (IRE1), PKR-like ER kinase (PERK) and
activating transcription factor 6 (ATF6) to turn on/ER stress for
tumor cells to escape stress-induced apoptosis. Also, GRP78 may
upregulate expression of anti-apoptotic molecule Bc1-2 within the
nucleus to bind with caspase-7 and caspase-12 in the cytoplasm to
prevent ER stress and subsequent cell cascades (25). GRP78 was reported to be critical to
tumor cell growth by inhibiting expression of fibrosarcoma cells
and constraining tumor formation in animals. Neuroglioma cells
proliferate well when GRP78 is highly expressed. When the GRP78
gene was knocked out (26), cancer
cell apoptosis was accelerated via induction of ER stress. Dong
et al (27) reported that
GRP78 facilitated tumor growth by improving proliferation and the
anti-apoptotic ability of cancer cells. MTHSP75 is a chaperone
located in mitochondria that helps regulate biological functions
including cell survival, proliferation, chondriosome synthesis and
transfer of intracellular protein (28). MTHSP75 decreases cell damage by
regulating opening of the mitochondrial permeability transition
pore (29) and high expression of
HSP75 may enhance anti-apoptotic capacities and motility of cancer
cells and cause cell proliferation and transfer. Chen and Hang
(30) observed changes in tumor
growth and transfer ability using siRNA to silence HSP75 in a human
osteosarcoma cell line and reported that this decreased cancer cell
growth and transference. In most apoptotic systems, Hsp60 can be
released from the mitochondria and aggregate cytoplasmically and
enhance apoptosis. Cell death was also induced by active substance
BMD188. Hsp60 accelerated caspase maturity and activation of
caspase-3 (31). In the marrow
stromal HS-5 cell line, HSP60 could activate caspase-3 and
caspase-9 but not caspase-8, prompting release of mitochondrial
cytochrome c to the cytoplasm, causing apoptosis (32). In the present study, we found that
SW480 cells underwent cell cycle arrest and increased apoptosis
after PE treatment and that GRP78, mtHSP75 and HSP60 were
downregulated, indicating similar functions.
NPM (or B23, NO38 and nurnatri), is a
multifunctional phosphorylated shuttle protein (33) important to various physiological and
pathological processes including centrosome duplication, ribosome
synthesis, DNA repair, molecular chaperone and stress response
caused by stimulants in vitro and in vivo. Previous
studies confirmed that downregulation of mRNA of NPM slowed cell
cycle progression and mitosis, and induced apoptosis and this may
occur via p53 inhibition (34).
After UV radiation cells were damaged and NPM reset the nucleoplasm
and regulated p53 and HDM2. By binding to HDM2, NPM negatively
regulated HDM2-p53 and stabilized p53 (35). Colombo et al (36) demonstrated that NPM interacted with
p53 after stress and the p53 enhanced function may be attributable
to transcriptional activity of downstream factor p21 to inhibit
tumorigenesis. When NPM siRNA transfected A431 cells were tested,
cell proliferation decreased and stopped at the S stage, and fewer
cells were in the mitotic phase, inhibiting cell growth (37). NPM1 in SW480 cells was
down-regulated significantly after PE treatment and was similar to
NPM1 silencing: SW480 cell growth was reduced indicating that NPM1
may be a target of PE to inhibit tumorigenesis.
Ezrin, the coding gene of vli2, as a member of the
ezrin-radixin-moesin (ERM) family is a membrane cytoskeletal
cross-linker protein involved in cell interaction and cell matrices
via regulation of actin, controlling cellular morphology, adjusting
cell adhesion molecules, signal transduction and engulfing tumor
cells (38). Ezrin has a
significant role in the development, infiltration and transfer of
tumor cells. Localized to a specific membrane area, ezrin combines
with membrane proteins (such as CD44, CD95 and ICAM-2) and
scaffolding proteins (such as E3KARR and EBP50) to activate the
membrane cytoskeletal chain via signal transduction. Also, tyrosine
phosphorylation sites on ezrin, when linked to PIP2, are activated
and polar-oriented (39). As a
receptor of tyrosine kinase, ezrin can intracellularly signal to
control cell proliferation and differentiation after a particular
tyrosine kinase phosphorylation. Decreasing expression of ezrin
protein inhibits cell proliferation and cells in metaphase cells
decrease (40). Also, a significant
reduction of cellular pseudopod formation and their migration and
invasiveness was reduced. Ezrin may do this through many pathways,
but specifics are not known. Similarly, in SW480 cells, PE can
reduce the expression of ezrin which could be the target spot of
inhibiting proliferation of colon cancer cells.
Annexin A2 (ANXA2), a calcium-dependent phospholipid
binding protein, is widely distributed in eukaryotic cell
membranes, the cytoplasm and extracellular media, accounting for up
to 0.5–2.0% of total cell protein (41). It mainly participates in membrane
transportation and in activities dependent on membrane calmodulin
such as membrane fusion of exocytosis, vesicle trafficking, cell
adhesion, proliferation, apoptosis, DNA replication, signal
transduction and ion channel formation (42,43).
Reports suggest that highly expressed ANXA2 in human colorectal
cancer tissues/cells may be a diagnostic marker as well as a target
for treatment and tumor prognosis (44). Here, ANXA2 in SW480 control cells
were more highly expressed than cells treated with PE which
inhibited proliferation and induced apoptosis and this likely
occurred via regulation of the epidermal growth factor receptor
signal transduction pathway with phosphorylation of tyrosin
residues that signal through the Ras and PI3K pathways to control
nuclear target gene expression (45). In addition, ANXA2 may regulate
expression of several apoptosis-related proteins (such as
caspase-3, p53, or Bc1-2; Figs. 5
and 6) to block apoptosis-related
pathways (46). Given the essential
role in intracellular information conduction, ANXA2 may be an
essential therapeutic target for tumor therapy by inhibiting its
expression or eliminating its influence on signaling pathway.
In conclusion, SW480 treated with PE underwent
significant changes in expression of various proteins that may
serve as related targets for PE-inhibition of tumors in
vitro. HSP, NPM1, Ezrin and Annexin A2 expression were
upregulated or downregulated by PE and had antitumor effects by
influencing cell apoptosis, proliferation and energy metabolism.
This is a novel theory for further study the mechanism of action at
a protein level and verification and functional analysis of these
proteins requires further study.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (nos. 30571009 and 81501808)
the Natural Science Foundation of Zhejiang Province, China (Z307471
and LQ17H160015) and the Science and Technology Foundation of
Zhejiang Province, China (2009C33040).
References
|
1
|
Dunn K Bullard: Retrorectal tumors. Surg
Clin North Am. 90:163–171. 2010.Table of Contents. doi:
10.1016/j.suc.2009.09.009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Seitz U, Bohnacker S, Seewald S, Thonke F,
Brand B, Bräiutigam T and Soehendra N: Is endoscopic polypectomy an
adequate therapy for malignant colorectal adenomas? Presentation of
114 patients and review of the literature. Dis Colon Rectum.
47:1789–1796; discussion 1796–1787. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang GR: Determination of seven trace
elements in Gracilaria Lemaneiformis by atomic absorption
spectrometry. Chin J Spectrosc Lab. 24:1005–1008. 2007.
|
|
4
|
Li BH, Xie SS and Lu ZK: Spectral
properties of new photosensitizers for photodynamic diagnosis and
therapy. Guang Pu Xue Yu Guang Pu Fen Xi. 22:902–904. 2002.(In
Chinese). PubMed/NCBI
|
|
5
|
Chen MZ, Du H and Xu BJ: Gracilaria
phycoerythrin influence on H-22 tumor immune function in mice. Chin
Tradit Herbal Drugs. 42:1329–1332. 2010.
|
|
6
|
Gao SY, Wang J and Ji YB: Study on
phycoerythrin induces MCF-7 apoptosis by controlling variation of
ΔΨ_m. J Harbin University of Commerce. 28:8–13. 2012.
|
|
7
|
He QY and Chiu JF: Proteomics in biomarker
discovery and drug development. J Cell Biochem. 89:868–886. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seliger B and Kellner R: Design of
proteome-based studies in combination with serology for the
identification of biomarkers and novel targets. Proteomics.
2:1641–1651. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Srinivas PR, Verma M, Zhao Y and
Srivastava S: Proteomics for cancer biomarker discovery. Clin Chem.
48:1160–1169. 2002.PubMed/NCBI
|
|
10
|
Alaiya AA, Franzén B, Hagman A,
Silfverswärd C, Moberger B, Linder S and Auer G: Classification of
human ovarian tumors using multivariate data analysis of
polypeptide expression patterns. Int J Cancer. 86:731–736. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Utleg AG, Yi EC, Xie T, Shannon P, White
JT, Goodlett DR, Hood L and Lin B: Proteomic analysis of human
prostasomes. Prostate. 56:150–161. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu LN, Chen XL, Zhang XY, Zhang YZ and
Zhou BC: One-step chromatography method for efficient separation
and purification of R-phycoerythrin from Polysiphonia urceolata. J
Biotechnol. 116:91–100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jaganathan SK, Supriyanto E and Mandal M:
Events associated with apoptotic effect of p-Coumaric acid in
HCT-15 colon cancer cells. World J Gastroenterol. 19:7726–7734.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu W, Yu P and Li J: Induction of
apoptosis in human colon carcinoma COLO 205 cells by the
recombinant α subunit of C-phycocyanin. Biotechnol Lett.
33:637–644. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saini MK, Sanyal SN and Vaiphei K:
Piroxicam and C-phycocyanin mediated apoptosis in
1,2-dimethylhydrazine dihydrochloride induced colon carcinogenesis:
Exploring the mitochondrial pathway. Nutr Cancer. 64:409–418. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Görg A, Obermaier C, Boguth G, Harder A,
Scheibe B, Wildgruber R and Weiss W: The current state of
two-dimensional electrophoresis with immobilized pH gradients.
Electrophoresis. 21:1037–1053. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li C, Tan YX, Zhou H, Ding SJ, Li SJ, Ma
DJ, Man XB, Hong Y, Zhang L, Li L, et al: Proteomic analysis of
hepatitis B virus-associated hepatocellular carcinoma:
Identification of potential tumor markers. Proteomics. 5:1125–1139.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang H, Yang Y, Chen W, Ding L, Li P, Zhao
X, Wang X, Li A and Bao Q: Identification of differentially
expressed proteins of Arthrospira (Spirulina) plantensis-YZ under
salt-stress conditions by proteomics and qRT-PCR analysis. Proteome
Sci. 11:62013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bermejo R, Acién FG, Ibáñez MJ, Fernández
JM, Molina E and Alvarez-Pez JM: Preparative purification of
B-phycoerythrin from the microalga Porphyridium cruentum by
expanded-bed adsorption chromatography. J Chromatogr B Analyt
Technol Biomed Life Sci. 790:317–325. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang B, Wang GC and Li ZG: Antitumor
studies of C-phycocyanin chromophore peptides mediated photodynamic
therapy. Acta Laser Biology Sinica. 11:194–198. 2002.
|
|
21
|
Morcos NC, Berns M and Henry WL:
Phycocyanin: Laser activation, cytotoxic effects, and uptake in
human atherosclerotic plaque. Lasers Surg Med. 8:10–17. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Phillips D: Chemical mechanisms in
phothodynamic therapywith Phthal ocyanines. Prog Reaction Kinet.
22:175–300. 1997.
|
|
23
|
Xu BH: Study on the apoptosis induced by
phycoerythrin through cell cycle in human breast cancer MCF-7. D
Harbin University of Commerce. 33–34. 2010.
|
|
24
|
Joo M, Chi JG and Lee H: Expressions of
HSP70 and HSP27 in hepatocellular carcinoma. J Korean Med Sci.
20:829–834. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suzuki T, Lu J, Zahed M, Kita K and Suzuki
N: Reduction of GRP78 expression with siRNA activates unfolded
protein response leading to apoptosis in HeLa cells. Arch Biochem
Biophys. 468:1–14. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rauschert N, Brändlein S, Holzinger E,
Hensel F, Müller-Hermelink HK and Vollmers HP: A new tumor-specific
variant of GRP78 as target for antibody-based therapy. Lab Invest.
88:375–386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dong D, Stapleton C, Luo B, Xiong S, Ye W,
Zhang Y, Jhaveri N, Zhu G, Ye R, Liu Z, et al: A critical role for
GRP78/BiP in the tumor microenvironment for neovascularization
during tumor growth and metastasis. Cancer Res. 71:2848–2857. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hua G, Zhang Q and Fan Z: Heat shock
protein 75 (TRAP1) antagonizes reactive oxygen species generation
and protects cells from granzyme M-mediated apoptosis. J Biol Chem.
282:20553–20560. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiang F, Huang YS, Shi XH and Zhang Q:
Mitochondrial chaperone tumour necrosis factor receptor-associated
protein 1 protects cardiomyocytes from hypoxic injury by regulating
mitochondrial permeability transition pore opening. FEBS J.
277:1929–1938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen JH and Hang W: Effects of HSP75 small
interfering RNA on cell proliferation and migration in human
osteosarcoma cell line U2-OS. J China Med Univ. 42:1145–1147.
2013.
|
|
31
|
Chandra D, Choy G and Tang DG: Cytosolic
accumulation of HSP60 during apoptosis with or without apparent
mitochondrial release: Evidence that its pro-apoptotic or
pro-survival functions involve differential interactions with
caspase-3. J Biol Chem. 282:31289–31301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim YS, Koh JM, Lee YS, Kim BJ, Lee SH,
Lee KU and Kim GS: Increased circulating heat shock protein 60
induced by menopause, stimulates apoptosis of osteoblast-lineage
cells via up-regulation of toll-like receptors. Bone. 45:68–76.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grisendi S, Mecucci C, Falini B and
Pandolfi PP: Nucleophosmin and cancer. Nat Rev Cancer. 6:493–505.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen D, Yoon JB and Gu W: Reactivating the
ARF-p53 axis in AML cells by targeting ULF. Cell Cycle.
9:2946–2951. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kurki S, Peltonen K, Latonen L, Kiviharju
TM, Ojala PM, Meek D and Laiho M: Nucleolar protein NPM interacts
with HDM2 and protects tumor suppressor protein p53 from
HDM2-mediated degradation. Cancer Cell. 5:465–475. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Colombo E, Marine JC, Danovi D, Falini B
and Pelicci PG: Nucleophosmin regulates the stability and
transcriptional activity of p53. Nat Cell Biol. 4:529–533. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen JH and Hang W: Effects of regulating
NPM on the proliferative activity of human SCC A431 cells. Soochow
University. 18–22. 2013.
|
|
38
|
Jonstrup SP, Koch J and Kjems J: A
microRNA detection system based on padlock probes and rolling
circle amplification. RNA. 12:1747–1752. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fais S, De Milito A and Lozupone F: The
role of FAS to ezrin association in FAS-mediated apoptosis.
Apoptosis. 10:941–947. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lozupone F, Lugini L, Matarrese P, Luciani
F, Federici C, Iessi E, Margutti P, Stassi G, Malorni W and Fais S:
Identification and relevance of the CD95-binding domain in the
N-terminal region of ezrin. J Biol Chem. 279:9199–9207. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lokman NA, Ween MP, Oehler MK and
Ricciardelli C: The role of annexin A2 in tumorigenesis and cancer
progression. Cancer Microenviron. 4:199–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fatimathas L and Moss SE: Annexins as
disease modifiers. Histol Histopathol. 25:527–532. 2010.PubMed/NCBI
|
|
43
|
Zhang X, Liu S, Guo C, Zong J and Sun MZ:
The association of annexin A2 and cancers. Clin Transl Oncol.
14:634–640. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Duncan R, Carpenter B, Main LC, Telfer C
and Murray GI: Characterisation and protein expression profiling of
annexins in colorectal cancer. Br J Cancer. 98:426–433. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Grewal T and Enrich C: Annexins -
modulators of EGF receptor signalling and trafficking. Cell Signal.
21:847–858. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Huang Y, Jin Y, Yan CH, Yu Y, Bai J, Chen
F, Zhao YZ and Fu SB: Involvement of Annexin A2 in p53 induced
apoptosis in lung cancer. Mol Cell Biochem. 309:117–123. 2008.
View Article : Google Scholar : PubMed/NCBI
|