Introduction
Ovarian cancer is one of the most lethal gynecologic
cancers with high morbidity in women worldwide (1). Surgical resection followed by
chemotherapy has been considered the most effective treatment for
ovarian cancer; however, most patients are diagnosed at advanced
stages and undergo recurrence after treatment, which leads to a
poor survival rate (2,3). To date, there is still lack of an
effective therapeutic method for ovarian cancer. It is of great
importance to develop promising strategies to improve the treatment
of ovarian cancer.
Cancer stem cells, a small subset of cancer cells,
are capable of self-renewal and multi-lineage differentiation into
cells that can form tumors (4).
These cells have been identified in various types of cancer and
contribute to cancer progression, metastasis, chemoresistance and
recurrence (5–8). Therefore, the elimination of cancer
stem cells may be a potential and promising therapy for the
treatment of cancer. Cancer stem cells express distinct cell
surface markers such as CD24, CD44, CD119 and CD133 (6,9,10).
Among these, CD133+ cancer stem cells have been widely
studied in ovarian cancers as CD133+ cells isolated from
ovarian cancer cells or primary ovarian tumor tissues show
self-renewal, multi-lineage differentiation and tumor initiation
capabilities (10–12). Given these findings, the elimination
of CD133+ ovarian cancer stem cells could be a novel and
effective therapeutic strategy for preventing ovarian cancer.
Truncated Bid (tBid), the C-terminal of full-length
Bid (a BH3-only subgroup of the Bcl-2 family member) cleaved by
caspase-8, with a molecular weight of 15 kDa, is a potent inducer
of cell apoptosis (13,14). tBid translocates to mitochondria and
activates the release of cytochrome c from mitochondria to
the cytosol wherein cytochrome c activates caspase-9/−3/−7
to trigger cell apoptosis (14–16).
Thus, tBid has been proposed as a promising molecular target for
killing cancer cells (17,18). Recombinant adenoviruses have been
suggested as an effective gene delivery tool for cancer treatment.
However, adenovirus-mediated tBid expression initiated by the CMV
promoter may kill packaging cells during the production of the
recombinant adenovirus, making it difficult to obtain high titers
of recombinant adenovirus. Therefore, a conditional expression
system is needed.
Cre (‘Causes recombination’) is a site-specific
recombinase that can mediate the specific recombinant process
between two LoxP (locus of crossover P1) sequences (19,20).
In the present study, we designed a recombinant adenovirus carrying
tBid (with CMV promoter) and a null gene expression cassette (Neo)
flanked by LoxP which was inserted upstream of tBid to block tBid
expression (Ad-CMV-LoxP-Neo-LoxP-tBid). To conditionally initiate
tBid expression, we introduced another recombinant adenovirus
carrying Cre (without CMV promoter), with a CD133 promoter inserted
upstream of Cre. Thus, Cre may only be expressed when the CD133
promoter was activated (Ad-CD133-Cre). We hypothesized that
infection of Ad-CD133-Cre into CD133+ ovarian cancer
stem cells may trigger the expression of Cre which may then mediate
the LoxP recombinant and cut-off the null gene expression cassette
between the two LoxP, ultimately leading to tBid overexpression.
The activity of the CD133 promoter is weaker than that of the CMV
promoter, but it has been reported that a very small amount of Cre
is able to cut-off LoxP sequences effectively (19,21).
Therefore, CD133 promoter-mediated Cre expression in
CD133+ ovarian cancer stem cells is sufficient to
trigger tBid overexpression. Together, this Cre/LoxP system
mediated-tBid overexpression could specifically function in and
kill CD133+ ovarian cancer stem cells, representing a
potential and promising adjuvant therapy for the prevention of
ovarian cancer metastasis and recurrence.
Materials and methods
Animals
Six-week-old female severe combined immune deficient
(SCID) mice (weighing 18–25 g) were purchased from the Guangdong
Medical Laboratory Animal Center (Guangzhou, China) and housed
under specific pathogen-free conditions (21±2°C and 12/12 h
light/dark) with free access to food and water. The animal
experimental procedures were reviewed and approved by the
Institutional Animal Care and Use Committee of Nanjing Medical
University Affiliated Suzhou Hospital.
Ovarian cancer cell culture
The ovarian cancer cell line A2780 purchased from
the American Type Culture Collection (ATCC; Manassas, VA, USA) was
cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% fetal bovine serum (FBS) (both from Gibco, Rockville, MD,
USA), 100 U/ml penicillin and 0.1 mg/ml streptomycin (Sigma, St.
Louis, MO, USA). Cells were routinely cultured in a humidified
incubator containing 5% CO2 at 37°C.
Generation of CD133+
ovarian cancer stem cells
The CD133+ ovarian cancer stem cells were
isolated by magnetic bead sorting as previously described (22). Briefly, cells were dissociated with
0.25% trypsin (Sigma) and suspended in ice-cold phosphate-buffered
saline (PBS). Then, single cells were incubated with microbeads
labeled with anti-CD133/1 monoclonal antibodies (Miltenyi Biotech,
Auburn, CA, USA) for 20 min at 4°C in the dark. The samples were
then washed with ice-cold PBS and sorted on a BD
fluorescence-activated cell sorting (FACS)Aria system (BD
Biosciences, San Jose, CA, USA). The CD133+ cells were
cultured in serum-free medium containing 10 ng/ml human recombinant
basic fibroblast growth factor and 20 ng/ml human recombinant
epidermal growth factor (Invitrogen, Carlsbad, CA, USA).
Construction, production and
purification of recombinant adenovirus
The cDNA fragments of the CD133 promoter and Cre
were amplified and inserted into adenovirus pShuttle plasmid
(Stratagene, Santa Clara, CA, USA) to construct the Ad-CD133-Cre
recombinant adenovirus. The cDNA fragments of LoxP-Neo-LoxP and
tBid were inserted into pShuttle-CMV plasmid (Stratagene) to
construct the Ad-CMV-LoxP-Neo-LoxP-tBid recombinant adenovirus. The
recombinant pShuttle and pAdEasy-1 plasmids (Stratagene) were
homologously recombined in BJ5183 bacteria. The recombined plasmids
were linearized and transfected into 293T cells (ATCC) to produce
the recombinant virus. After 14 days, the cells were harvested and
lyzed by freeze-thawing. The supernatants were collected and
concentrated by CsCl gradient centrifugation. The titers were
determined using the 50% tissue culture infectious dose method
(23). Cells were infected with
Ad-CD133-Cre at a multiplicity of infection (MOI) of 20 and
Ad-CMV-LoxP-Neo-LoxP-tBid at MOI 30.
Western blot analysis
Mitochondrial and cytosolic proteins were extracted
using a mitochondria isolation kit (Pierce, Rockford, IL, USA). The
proteins were separated by 12.5% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and then transferred
onto a nitrocellulose membrane (Bio-Rad, Hercules, CA, USA). The
membrane was then blocked in 2.5% non-fat milk at 37°C for 1 h. The
membrane was blotted with primary antibodies at 4°C overnight.
After washing three times with Tris-buffered saline (TBS)-Tween
(TBST) and once with TBS, the membrane was incubated with
horseradish peroxidase (HRP)-conjugated secondary antibody
(1:2,000; Bioss Antibodies, Beijing, China) for 1 h at 37°C. After
washing with TBST, the protein bands were visualized using a
chemiluminescence detection procedure (Amersham Biosciences,
Beijing, China). The band intensity was quantified using Image Pro
Plus 6.0 software (Media Cybernetics, Inc., Rockville, MD, USA).
The primary antibodies used in this experiment were as follows:
anti-Bid, anti-Cre, anti-cytochrome c, anti-cleaved
caspase-9 and anti-β-actin (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) and anti-cleaved caspase-3 and anti-COX IV (Abcam,
Cambridge, MA, USA).
Cell viability assay
Cell growth and viability were detected by MTT
assay. Briefly, cells were plated onto a 96-well plate at a density
of 1×104 cells/well and cultured overnight. Thereafter,
the cells were infected with Ad-CD133-Cre or/and
Ad-CMV-LoxP-Neo-LoxP-tBid and incubated for 48 h. The medium was
replaced with fresh medium and 20 µl of MTT stock solution (5
mg/ml; Sigma) was added to each well. After culturing for 4 h, 200
µl of dimethyl sulfoxide was added to each well to dissolve the
formazan crystals. Then, absorbance at a wavelength of 490 nm was
detected using a microplate reader (BioTek Instruments, Winooski,
VT, USA).
TUNEL assay
Cell apoptosis was assessed using the terminal
deoxynucleotidyl transferase dUTP nick end-labeling (TUNEL) kit
(Roche, Indianapolis, IN, USA). Briefly, the infected cells were
fixed with 4% paraformaldehyde and permeabilized with 0.2% Triton
X-100. Then, the cells were incubated with TUNEL reaction mixtures
for 1 h at 37°C. Then, the cells were observed using a fluorescence
microscope (Olympus, Tokyo, Japan), and apoptotic cells were
counted in five random fields/slide.
Flow cytometric analysis
Annexin V-fluorescein isothiocyanate (FITC) and
propidium iodide (PI) stained cells were detected by flow
cytometric analysis according to the manufacturer's instructions
(Jiancheng Bioengineering Institute, Nanjing, China). Briefly,
cells were dissociated with trypsin, washed with ice-cold PBS and
resuspended in binding buffer at 1×106 cells/ml. Then,
Annexin V and PI stock solution were added and incubated at 4°C in
the dark. Thereafter, the samples were detected by FACS and the
data were analyzed by FlowJo software (BD Biosciences).
In vivo xenograft experiments
CD133+ ovarian cancer stem cells
(1×105 cells) were suspended in 200 µl of PBS and
subcutaneously injected into the right flank of SCID mice (n=6
mice/group). Approximately 1×1010 plaque-forming units
of recombinant adenovirus diluted in 50 µl of PBS were
intratumorally injected every 3 days. Tumor volume (V) was measured
using an external caliper and calculated according to the formula:
V = length × width2 × π/6. The mice were sacrificed by
subcutaneous injection with sodium pentobarbital (100 mg/kg) 21
days after inoculation and the tumors were excised for western blot
analysis and TUNEL assay.
Data analysis
Data are presented as mean ± standard deviation
(SD). Statistical analyses were performed by one-way analysis of
variance followed by Bonferroni post hoc test using SPSS version
11.5 (SPSS, Inc., Chicago, IL, USA). Differences were considered
statistically significant at p<0.05.
Results
Cre/LoxP system-mediated tBid
overexpression in CD133+ ovarian cancer stem cells
To achieve conditional tBid overexpression, we
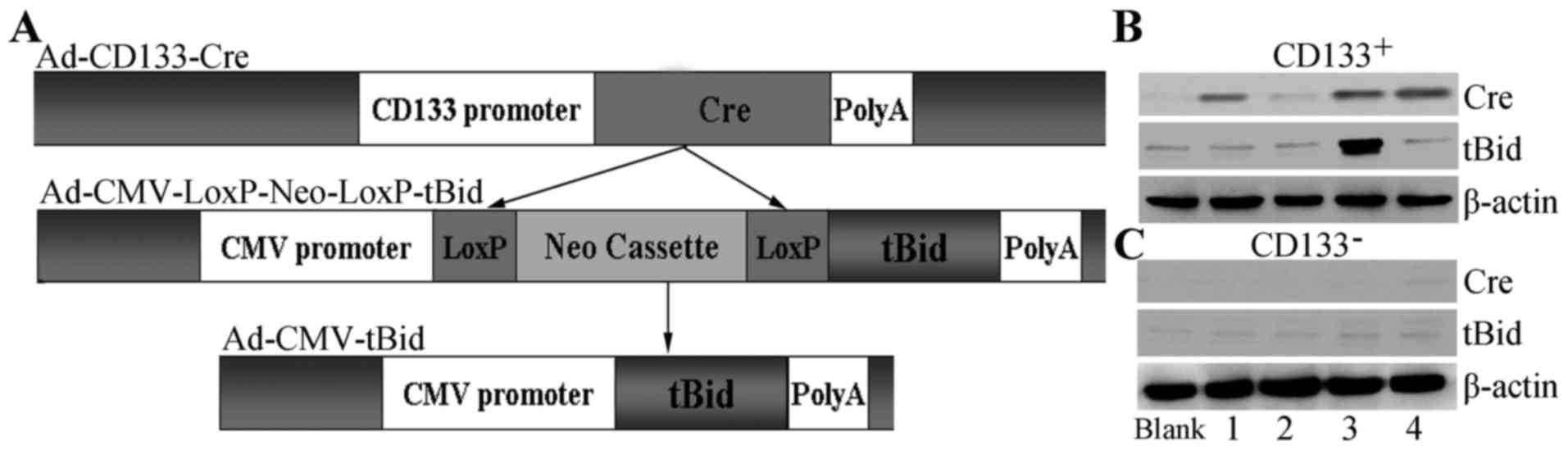
introduced a Cre/LoxP-mediated expression system (Fig. 1A). Briefly, we cloned the CD133
promoter into an adenovirus and inserted the open reading frame of
Cre downstream from the CD133 promoter. Therefore, Cre was
expressed only when the CD133 promoter was activated. Meanwhile, we
constructed another recombinant adenovirus containing a flanked
LoxP cassette upstream of tBid. Thus, tBid was expressed only when
the LoxP cassette was deleted. To test whether this Cre/LoxP system
was successful, we infected these recombinant adenoviruses into
CD133+ ovarian cancer stem cells. The results showed
that Cre was expressed in the Ad-CD133-Cre-infected
CD133+ ovarian cancer stem cells (Fig. 1B). The single infection of
Ad-CMV-LoxP-Neo-LoxP-tBid did not result in tBid overexpression
whereas co-infection of Ad-CD133-Cre with Ad-CMV-LoxP-Neo-LoxP-tBid
into CD133+ ovarian cancer stem cells resulted in tBid
overexpression (Fig. 2B). In
contrast, in CD133− cells, infection of Ad-CD133-Cre
or/and Ad-CMV-LoxP-Neo-LoxP-tBid did not produce Cre or tBid
(Fig. 1C). These results indicated
that the Cre/LoxP-mediated tBid overexpression system was
successfully established in the CD133+ ovarian cancer
stem cells.
Cre/LoxP system-mediated tBid
overexpression promotes cell apoptosis of CD133+ ovarian
cancer stem cells
To determine the efficiency of Cre/LoxP
system-mediated tBid overexpression in gene therapy against ovarian
cancer, we assessed its biological effect on cell growth and
apoptosis in CD133+ ovarian cancer stem cells in
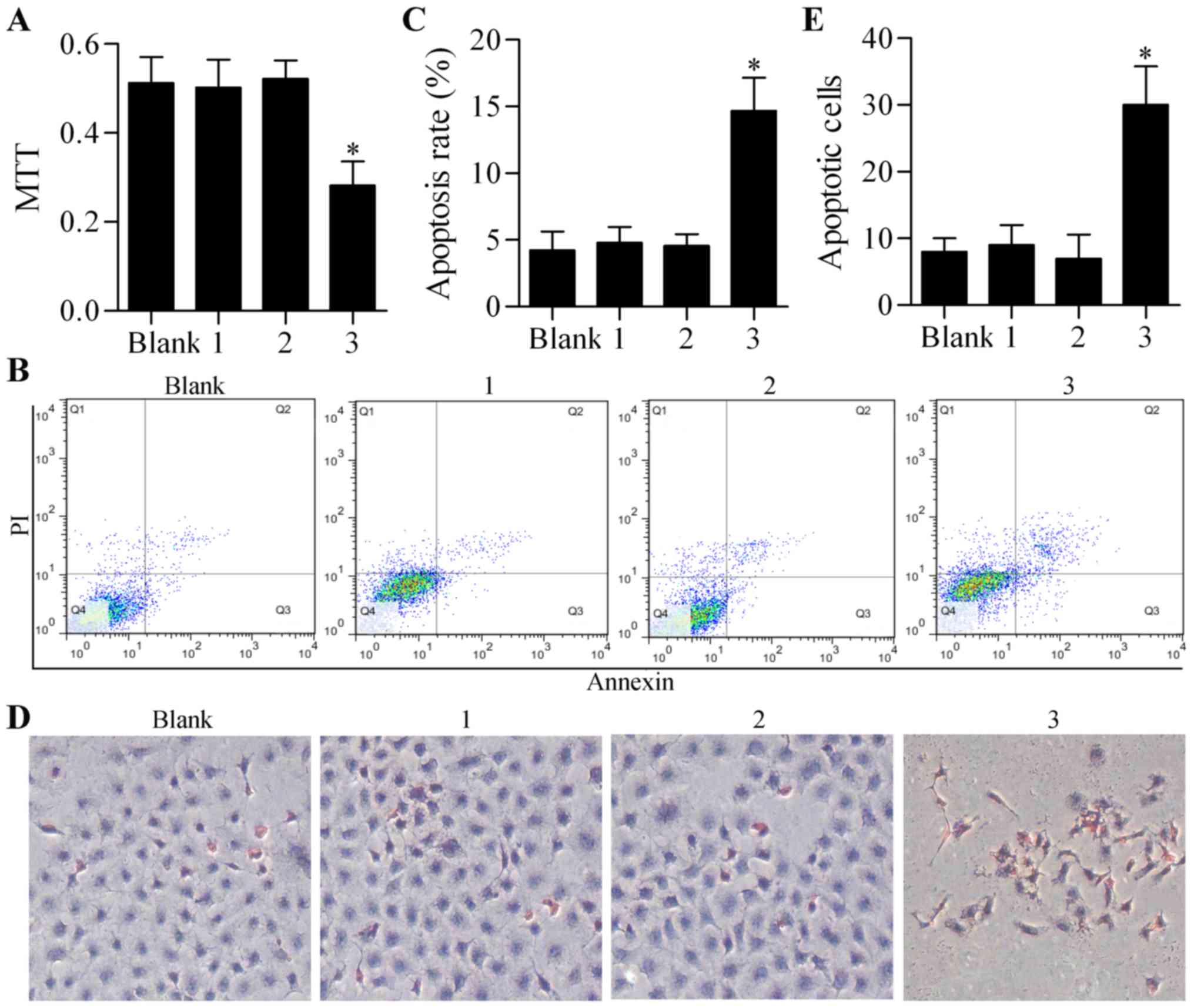
vitro. The results showed that the single infection of
Ad-CD133-Cre or Ad-CMV-LoxP-Neo-LoxP-tBid had no obvious effect on
cell growth or viability, as detected by MTT assay (Fig. 2A). However, the co-infection of
Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid significantly inhibited
the growth of CD133+ ovarian cancer stem cells (Fig. 2A). Furthermore, Annexin V-FITC/PI
assay also showed a significantly high apoptosis rate in the
CD133+ ovarian cancer stem cells co-infected with
Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid (Fig. 2B and C). Similarly, TUNEL assay
showed that the co-infection of Ad-CD133-Cre and
Ad-CMV-LoxP-Neo-LoxP-tBid markedly increased cell apoptosis of the
CD133+ ovarian cancer stem cells compared with single
infection of Ad-CD133-Cre or Ad-CMV-LoxP-Neo-LoxP-tBid (Fig. 2D and E). Taken together, these
results suggested that Cre/LoxP system-mediated tBid overexpression
effectively promoted the apoptosis of CD133+ ovarian
cancer stem cells.
Cre/LoxP system-mediated tBid
overexpression activates the pro-apoptotic signaling pathway
To further verify the effect of Cre/LoxP
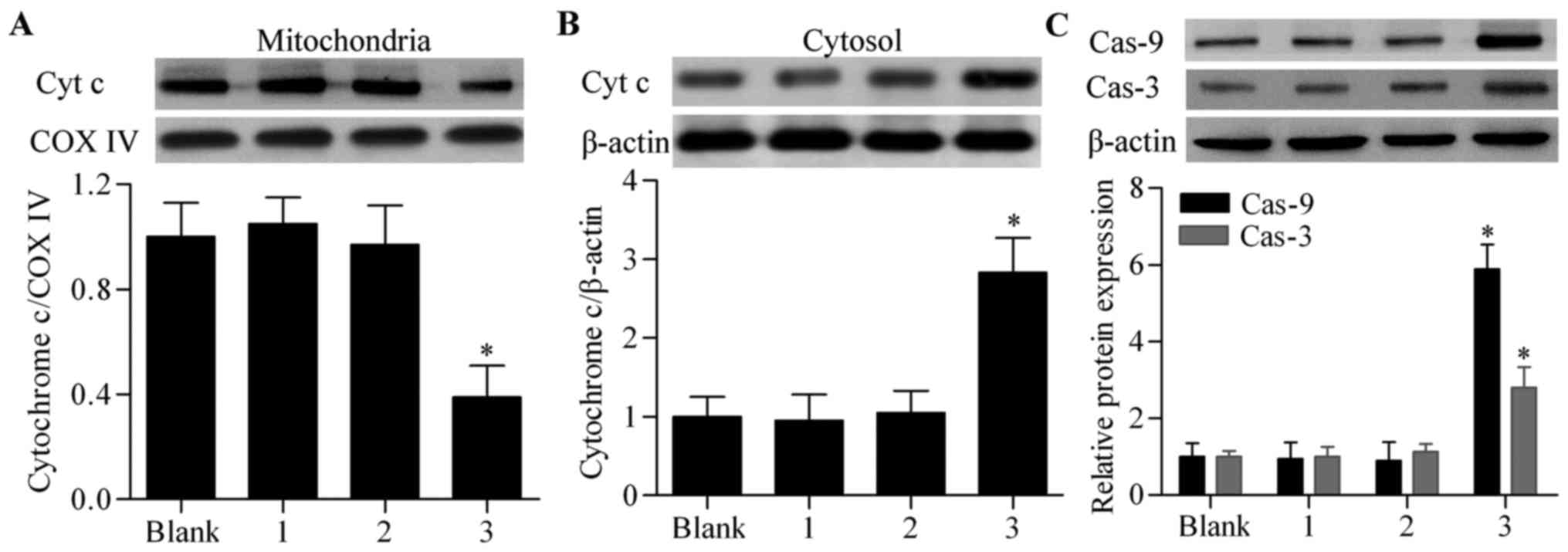
system-mediated tBid overexpression on induction of cell apoptosis,
we assessed its effect on the pro-apoptotic signaling pathway. The
results showed that the co-infection of Ad-CD133-Cre and
Ad-CMV-LoxP-Neo-LoxP-tBid significantly upregulated the release of
cytochrome c from mitochondria to the cytosol compared with
the other groups (Fig. 3A and B).
Furthermore, the activated forms of caspase-9 and caspase-3
(Fig. 3C) were also significantly
upregulated by co-infection of Ad-CD133-Cre and
Ad-CMV-LoxP-Neo-LoxP-tBid in the CD133+ ovarian cancer
stem cells. Taken together, these results indicated that the
Cre/LoxP system-mediated tBid overexpression significantly
activated the pro-apoptotic signaling pathway in the
CD133+ ovarian cancer stem cells.
Cre/LoxP system-mediated tBid
overexpression augments the cytotoxic effect of cisplatin in
CD133+ ovarian cancer stem cells
To further determine the efficiency of Cre/LoxP
system-mediated tBid overexpression in gene therapy against ovarian
cancer, we investigated the effect of Cre/LoxP system-mediated tBid
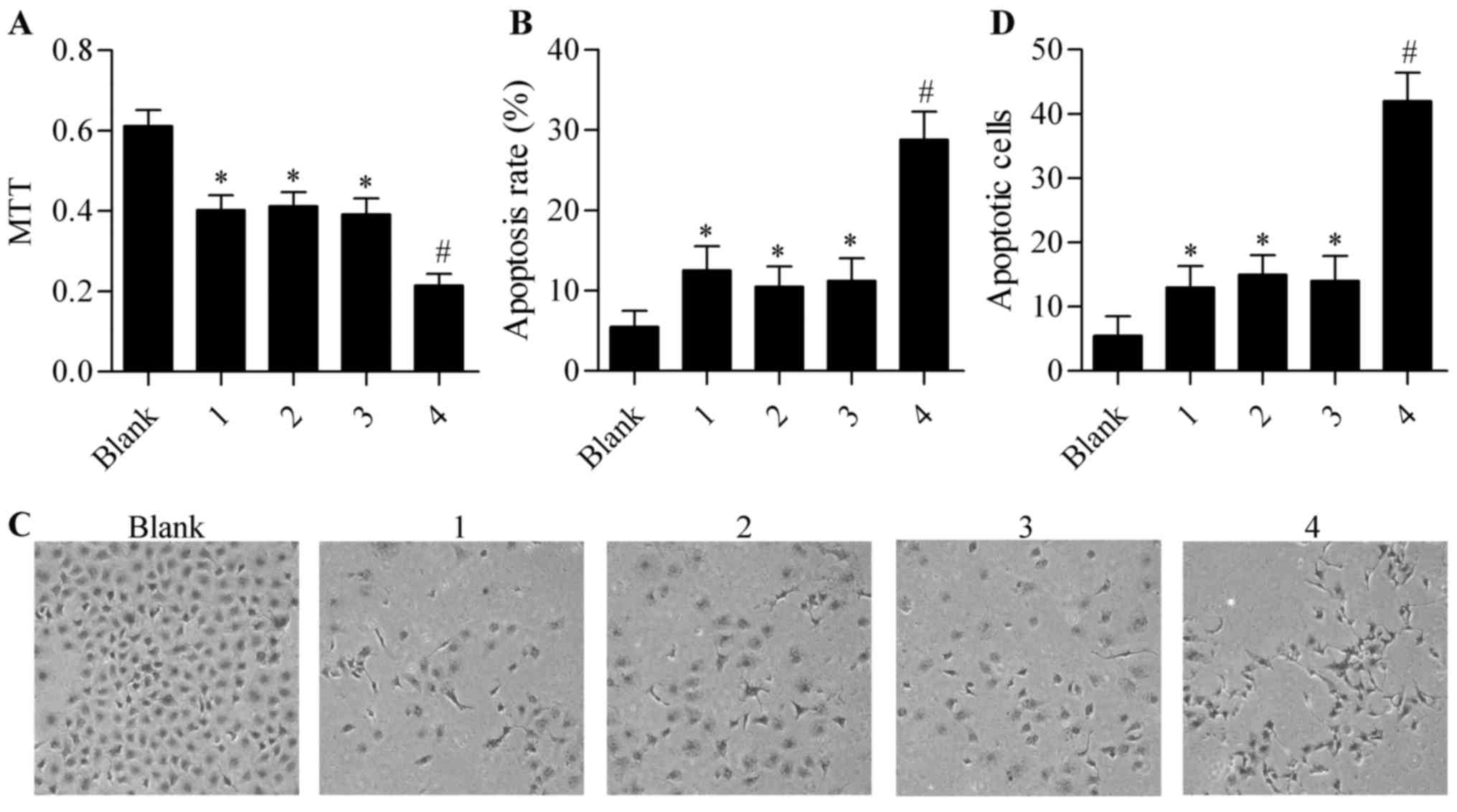
overexpression on cisplatin-induced cytotoxicity. The results
showed that cisplatin treatment inhibited the growth and viability
of CD133+ ovarian cancer stem cells, and that the single
infection of Ad-CD133-Cre or Ad-CMV-LoxP-Neo-LoxP-tBid had no
obvious effect on cisplatin-induced cytotoxicity (Fig. 4A). However, co-infection with
Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid significantly augmented
the cytotoxic effect of cisplatin in the CD133+ ovarian
cancer stem cells (Fig. 4A).
Furthermore, cisplatin-induced cell apoptosis was also enhanced by
co-infection with Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid in
comparison with single infection of Ad-CD133-Cre or
Ad-CMV-LoxP-Neo-LoxP-tBid (Fig.
4B-E). Taken together, these results suggested that Cre/LoxP
system-mediated tBid overexpression increased the sensitivity of
CD133+ ovarian cancer stem cells to cisplatin.
Cre/LoxP system-mediated tBid
overexpression inhibits tumor growth in vivo
To further confirm the antitumor effect of Cre/LoxP
system-mediated tBid overexpression against CD133+
ovarian cancer stem cells, we subcutaneously inoculated the tumor
cells into SCID mice. The recombinant adenoviruses were
intratumorally injected into the mice and the tumor volume was
monitored. The results showed that the single infection of
Ad-CD133-Cre or Ad-CMV-LoxP-Neo-LoxP-tBid had no obvious effect on
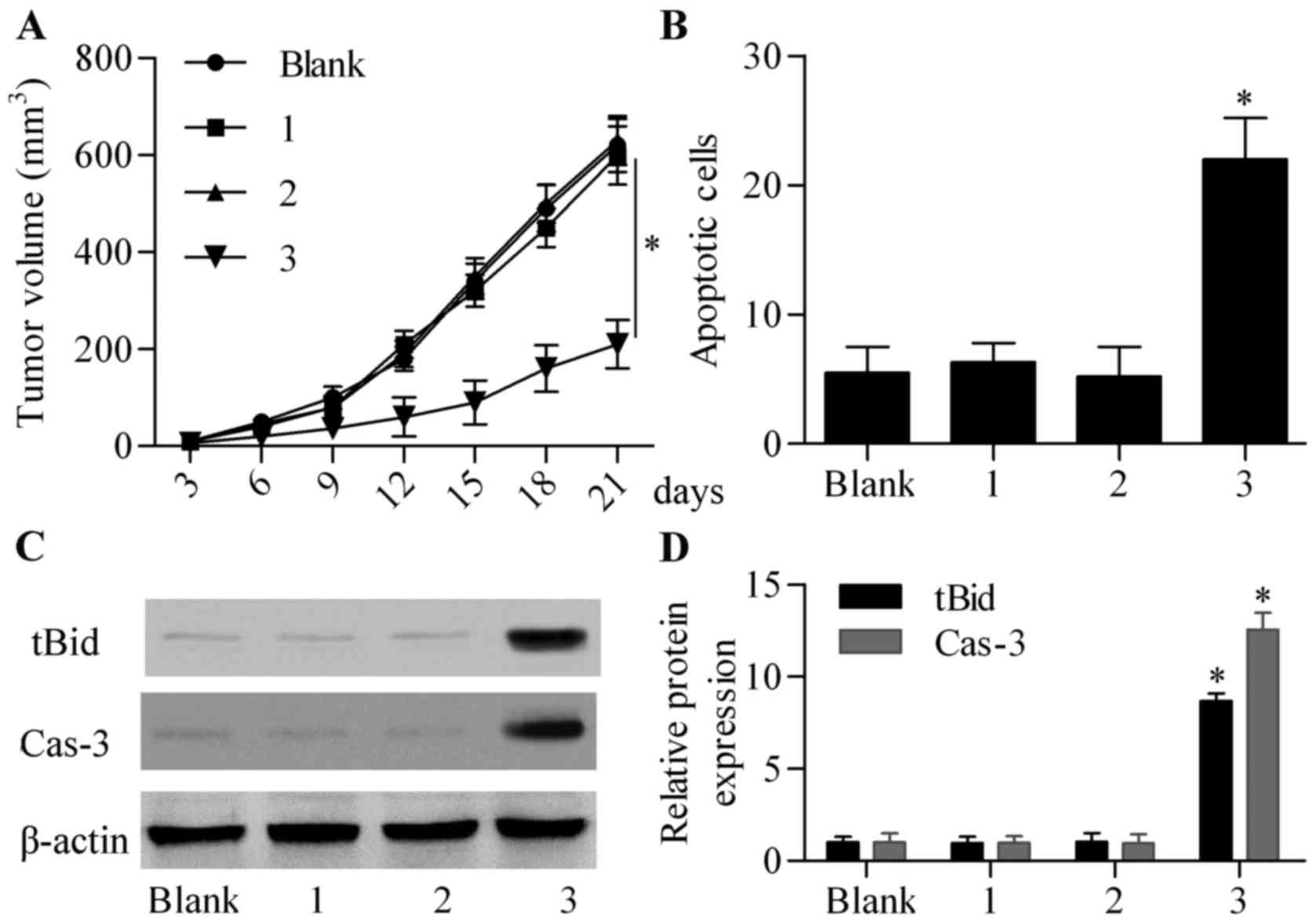
tumor growth compared with the non-infected control group (Fig. 5A). As expected, the co-infection of
Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid significantly suppressed
tumor growth compared with the other groups (Fig. 5A). TUNEL assay showed that the
apoptotic cells were not significantly altered in the Ad-CD133-Cre
or Ad-CMV-LoxP-Neo-LoxP-tBid single-infected group, but markedly
increased in the Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid
co-infected group (Fig. 5B). By
analysis of xenograft tumor tissues, we found that tBid was
overexpressed in the Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid
co-infected group (Fig. 5C and D).
Moreover, caspase-3 activity was also markedly upregulated by
co-infection with Ad-CD133-Cre and Ad-CMV-LoxP-Neo-LoxP-tBid
(Fig. 5C and D). Taken together,
these results revealed that Cre/LoxP system-mediated tBid
overexpression triggered cell apoptosis and suppressed tumor growth
in vivo.
Discussion
In the present study, we successfully established
conditional truncated Bid (tBid) overexpression induced by the
Cre/LoxP system. tBid overexpression mediated by the Cre/LoxP
system was triggered in CD133+ ovarian cancer stem
cells, significantly inducing cell apoptosis and inhibiting
CD133+ ovarian cancer stem cell growth in vitro
and in vivo. The present study suggests that
adenovirus-mediated tBid overexpression induced by the Cre/LoxP
system may represent a potential therapeutic tool for anti-ovarian
cancer therapy.
CD133 has been suggested as a stem cell marker in
many types of stem cells including hematopoietic, prostate
epithelial and renal stem cells, and endothelial progenitor cells
(24–27), implying that CD133 is associated
with the original cell differentiation state. In malignant tumors,
the promoter region of CD133 is in a hypomethylated state and CD133
is highly expressed in various types of cancers, including ovarian
(6), breast (28), brain (5), prostate (29), colon (30) and pancreatic cancer (31). Increasing evidence suggests that
CD133+ cells isolated from ovarian cancer cells or
primary ovarian tumor tissues have self-renewal, multi-lineage
differentiation and tumor initiation capabilities and are thus
defined as ovarian cancer stem cells (10–12). A
very small amount of CD133+ ovarian cancer stem cells
can form tumors in SCID (12).
Therefore, CD133 serves as a potential molecular target for
eliminating CD133+ cancer stem cells. One approach for
molecular-targeted therapy is monoclonal antibody therapy, for
example using monoclonal antibodies targeting CD44 and CD123 in the
removal of human acute myeloid leukemic stem cells (32,33).
Smith et al reported that the CD133 single-chain antibody
conjugated with cytotoxic drugs inhibited hepatocellular and
gastric cancer cell growth (34).
However, the effect of CD133 antibody targeted therapy is still
limited due to the complex molecular structure of CD133 (35,36).
Utilizing specific transcriptional regulation is
another strategy. The carcinoembryonic antigen (CEA)
promoter-mediated Cre/LoxP system was found to induce the
expression of the cytosine deaminase and specifically inhibit
CEA-producing gastric cancer cells in vitro and in
vivo (37). Similarly, in
CD133+ cells, the CD133 promoter can be used to control
selective gene expression. In the present study, we wanted to
introduce tBid, a potent suicide gene, into CD133+
ovarian cancer stem cells in order to kill the cells. However, the
recombinant adenovirus carrying tBid driven by the CMV potent
promoter is difficult to obtain since the tBid expression may kill
packaging cells during the production of the recombinant
adenovirus. Therefore, a conditional expression system was needed.
Miao et al constructed a recombinant adenovirus containing a
tBid gene driven by an α-fetoprotein (AFP) promoter and tBid was
overexpressed in AFP-producing hepatocellular carcinoma cells
(38). Similarly, a recent study
demonstrated that a selected fragment of the AFP promoter
designated as EA4D exhibited the highest reporter activity and that
the tBid driven by EA4D was highly and specifically expressed in
AFP-producing hepatocellular carcinoma cells (17). In CD133+ cancer stem
cells, we used the CD133 promoter to drive tBid expression.
However, the expression level of tumor-specific promoters is
generally low and the activity of the CD133 promoter is weaker than
that of the CMV promoter (39). The
tBid expression driven directly by the CD133 promoter is so low
that it cannot effectively kill CD133+ ovarian cancer
stem cells. Therefore, a way of augmenting the selective expression
of the tBid gene was needed.
In the present study, we used the Cre/LoxP
regulation system to selectively augment tBid expression. We
constructed two recombinant adenoviruses, Ad-CMV-LoxP-Neo-LoxP-tBid
and Ad-CD133-Cre. The tBid cannot be expressed in
Ad-CMV-LoxP-Neo-LoxP-tBid, and thus we were able to obtain a high
titer of recombinant adenovirus. With regards to Ad-CD133-Cre, the
Cre is expressed in CD133+ cancer stem cells. Even
though the CD133 promoter was a weaker promoter, a very small
amount of Cre was able to cut-off LoxP sequences effectively
(19,21). After LoxP was cut-off in
Ad-CMV-LoxP-Neo-LoxP-tBid, the tBid was highly overexpressed as
driven by the CMV promoter in CD133+ cancer stem cells.
Therefore, a selective and augmented tBid expression system was
established. Co-infection with Ad-CMV-LoxP-Neo-LoxP-tBid and
Ad-CD133-Cre effectively inhibited cell growth and induced cell
apoptosis in the CD133+ ovarian cancer stem cells. tBid
is a potent apoptosis inducer and sensitizes apoptosis induced by
chemotherapeutic drugs in cancer cells (40). The present study also revealed that
Cre/LoxP system-mediated tBid overexpression augmented the
cytotoxic effect of cisplatin in CD133+ ovarian cancer
stem cells. The data indicate that Cre/LoxP system mediated-tBid
overexpression represents a potential and promising adjuvant
therapy to prevent ovarian cancer metastasis and recurrence.
In conclusion, the present study applied the
Cre/loxP system to augment the selective expression of the tBid
gene in CD133+ ovarian cancer stem cells. The Cre/LoxP
system mediated-tBid overexpression significantly induced the
apoptosis of CD133+ ovarian cancer stem cells in
vitro and in vivo, representing a future clinical
approach for preventing ovarian cancer metastasis and recurrence.
Furthermore, the adenovirus-mediated tBid overexpression induced by
the Cre/LoxP system could be applicable to other CD133+
cancer stem cells.
Acknowledgements
The present study was supported by grants from the
Natural Science Foundation of Jiangsu Province (BK2012599), the
Suzhou Municipal Science and Technology Development Project
(SYSD2012088), and the Suzhou Key Medical Center (SZZX201506).
Glossary
Abbreviations
Abbreviations:
|
tBid
|
truncated Bid
|
|
Cre
|
causes recombination
|
|
LoxP
|
locus of crossover P1
|
|
SCID
|
severe combined immune-deficient
|
|
AFP
|
α-fetoprotein
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chekerov R, Braicu I, Castillo-Tong DC,
Richter R, Cadron I, Mahner S, Woelber L, Marth C, Van Gorp T,
Speiser P, et al: Outcome and clinical management of 275 patients
with advanced ovarian cancer International Federation of Obstetrics
and Gynecology II to IV inside the European Ovarian Cancer
Translational Research Consortium-OVCAD. Int J Gynecol Cancer.
23:268–275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peiretti M, Bristow RE, Zapardiel I,
Gerardi M, Zanagnolo V, Biffi R, Landoni F, Bocciolone L, Aletti GD
and Maggioni A: Rectosigmoid resection at the time of primary
cytoreduction for advanced ovarian cancer. A multi-center analysis
of surgical and oncological outcomes. Gynecol Oncol. 126:220–223.
2012.
|
|
4
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
6
|
Zhang S, Balch C, Chan MW, Lai HC, Matei
D, Schilder JM, Yan PS, Huang TH and Nephew KP: Identification and
characterization of ovarian cancer-initiating cells from primary
human tumors. Cancer Res. 68:4311–4320. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iwasaki H and Suda T: Cancer stem cells
and their niche. Cancer Sci. 100:1166–1172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao J: Cancer stem cells and
chemoresistance: The smartest survives the raid. Pharmacol Ther.
160:145–158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gao MQ, Choi YP, Kang S, Youn JH and Cho
NH: CD24+ cells from hierarchically organized ovarian
cancer are enriched in cancer stem cells. Oncogene. 29:2672–2680.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Curley MD, Therrien VA, Cummings CL,
Sergent PA, Koulouris CR, Friel AM, Roberts DJ, Seiden MV, Scadden
DT, Rueda BR, et al: CD133 expression defines a tumor initiating
cell population in primary human ovarian cancer. Stem Cells.
27:2875–2883. 2009.PubMed/NCBI
|
|
11
|
Moserle L, Indraccolo S, Ghisi M, Frasson
C, Fortunato E, Canevari S, Miotti S, Tosello V, Zamarchi R,
Corradin A, et al: The side population of ovarian cancer cells is a
primary target of IFN-alpha antitumor effects. Cancer Res.
68:5658–5668. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baba T, Convery PA, Matsumura N, Whitaker
RS, Kondoh E, Perry T, Huang Z, Bentley RC, Mori S, Fujii S, et al:
Epigenetic regulation of CD133 and tumorigenicity of
CD133+ ovarian cancer cells. Oncogene. 28:209–218. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Esposti MD: The roles of Bid. Apoptosis.
7:433–440. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo X, Budihardjo I, Zou H, Slaughter C
and Wang X: Bid, a Bcl2 interacting protein, mediates cytochrome c
release from mitochondria in response to activation of cell surface
death receptors. Cell. 94:481–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu X, Kim CN, Yang J, Jemmerson R and
Wang X: Induction of apoptotic program in cell-free extracts:
Requirement for dATP and cytochrome c. Cell. 86:147–157. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Zhu H, Xu CJ and Yuan J: Cleavage of
BID by caspase 8 mediates the mitochondrial damage in the Fas
pathway of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu BG, Liu LP, Chen GG, Ye CG, Leung KK,
Ho RL, Lin MC and Lai PB: Therapeutic efficacy of improved
α-fetoprotein promoter-mediated tBid delivered by
folate-PEI600-cyclodextrin nanopolymer vector in hepatocellular
carcinoma. Exp Cell Res. 324:183–191. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cristofanon S and Fulda S: ABT-737
promotes tBid mitochondrial accumulation to enhance TRAIL-induced
apoptosis in glioblastoma cells. Cell Death Dis. 3:e4322012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stricklett PK, Nelson RD and Kohan DE:
Site-specific recombination using an epitope tagged bacteriophage
P1 Cre recombinase. Gene. 215:415–423. 1998.PubMed/NCBI
|
|
20
|
Kanegae Y, Lee G, Sato Y, Tanaka M, Nakai
M, Sakaki T, Sugano S and Saito I: Efficient gene activation in
mammalian cells by using recombinant adenovirus expressing
site-specific Cre recombinase. Nucleic Acids Res. 23:3816–3821.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou L, Wei X, Cheng L, Tian J and Jiang
JJ: CD133, one of the markers of cancer stem cells in Hep-2 cell
line. Laryngoscope. 117:455–460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Long H, Xie R, Xiang T, Zhao Z, Lin S,
Liang Z, Chen Z and Zhu B: Autocrine CCL5 signaling promotes
invasion and migration of CD133+ ovarian cancer
stem-like cells via NF-κB-mediated MMP-9 upregulation. Stem Cells.
30:2309–2319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kanegae Y, Makimura M and Saito I: A
simple and efficient method for purification of infectious
recombinant adenovirus. Jpn J Med Sci Biol. 47:157–166. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peichev M, Naiyer AJ, Pereira D, Zhu Z,
Lane WJ, Williams M, Oz MC, Hicklin DJ, Witte L, Moore MA, et al:
Expression of VEGFR-2 and AC133 by circulating human
CD34+ cells identifies a population of functional
endothelial precursors. Blood. 95:952–958. 2000.PubMed/NCBI
|
|
25
|
Uchida N, Buck DW, He D, Reitsma MJ, Masek
M, Phan TV, Tsukamoto AS, Gage FH and Weissman IL: Direct isolation
of human central nervous system stem cells. Proc Natl Acad Sci USA.
97:14720–14725. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Corbeil D, Röper K, Hellwig A, Tavian M,
Miraglia S, Watt SM, Simmons PJ, Peault B, Buck DW and Huttner WB:
The human AC133 hematopoietic stem cell antigen is also expressed
in epithelial cells and targeted to plasma membrane protrusions. J
Biol Chem. 275:5512–5520. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Weigmann A, Corbeil D, Hellwig A and
Huttner WB: Prominin, a novel microvilli-specific polytopic
membrane protein of the apical surface of epithelial cells, is
targeted to plasmalemmal protrusions of non-epithelial cells. Proc
Natl Acad Sci USA. 94:12425–12430. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Marsden CG, Wright MJ, Pochampally R and
Rowan BG: Breast tumor-initiating cells isolated from patient core
biopsies for study of hormone action. Methods Mol Biol.
590:363–375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maitland NJ and Collins AT: Prostate
cancer stem cells: A new target for therapy. J Clin Oncol.
26:2862–2870. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li C, Lee CJ and Simeone DM:
Identification of human pancreatic cancer stem cells. Methods Mol
Biol. 568:161–173. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jin L, Hope KJ, Zhai Q, Smadja-Joffe F and
Dick JE: Targeting of CD44 eradicates human acute myeloid leukemic
stem cells. Nat Med. 12:1167–1174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jin L, Lee EM, Ramshaw HS, Busfield SJ,
Peoppl AG, Wilkinson L, Guthridge MA, Thomas D, Barry EF, Boyd A,
et al: Monoclonal antibody-mediated targeting of CD123, IL-3
receptor alpha chain, eliminates human acute myeloid leukemic stem
cells. Cell Stem Cell. 5:31–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Smith LM, Nesterova A, Ryan MC, Duniho S,
Jonas M, Anderson M, Zabinski RF, Sutherland MK, Gerber HP, Van
Orden KL, et al: CD133/prominin-1 is a potential therapeutic target
for antibody-drug conjugates in hepatocellular and gastric cancers.
Br J Cancer. 99:100–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Deonarain MP, Kousparou CA and Epenetos
AA: Antibodies targeting cancer stem cells: A new paradigm in
immunotherapy? MAbs. 1:12–25. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bidlingmaier S, Zhu X and Liu B: The
utility and limitations of glycosylated human CD133 epitopes in
defining cancer stem cells. J Mol Med. 86:1025–1032. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ueda K, Iwahashi M, Nakamori M, Nakamura
M, Matsuura I, Yamaue H and Tanimura H: Carcinoembryonic
antigen-specific suicide gene therapy of cytosine
deaminase/5-fluorocytosine enhanced by the Cre/loxP system in the
orthotopic gastric carcinoma model. Cancer Res. 61:6158–6162.
2001.PubMed/NCBI
|
|
38
|
Miao J, Chen GG, Chun SY, Yun JP, Chak EC,
Ho RL and Lai PB: Adenovirus-mediated tBid overexpression results
in therapeutic effects on p53-resistant hepatocellular carcinoma.
Int J Cancer. 119:1985–1993. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sato Y, Tanaka K, Lee G, Kanegae Y, Sakai
Y, Kaneko S, Nakabayashi H, Tamaoki T and Saito I: Enhanced and
specific gene expression via tissue-specific production of Cre
recombinase using adenovirus vector. Biochem Biophys Res Commun.
244:455–462. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miao J, Chen GG, Chun SY, Chak EC and Lai
PB: Bid sensitizes apoptosis induced by chemotherapeutic drugs in
hepatocellular carcinoma. Int J Oncol. 25:651–659. 2004.PubMed/NCBI
|