Introduction
Melanoma, a malignant tumor of skin cancer, is often
identified as one of the most deadly of tumors. The incidence of
melanoma has continued to increase in the past decades (1). Despite the variety of therapies used
to treat melanoma, the prognosis for melanoma remains very poor,
with a 5-year survival rate of <5% (2,3). Most
therapeutics such as BRAF inhibitors, cytotoxic T
lymphocyte-associated antigen 4 antibody and interleukin-2
biological therapy have failed in melanoma treatment because of the
low response rates and significant toxicities (4–6).
Therefore, new candidates for melanoma that possess potential
antitumor activity and low toxicity are urgently needed.
Many signaling pathways are involved in the
progression of melanoma, including signal transducer and activator
of transcription 3 (STAT3) signaling pathway (7). STAT3 is constitutively activated in
50–90% of melanoma and the high expression of phosphorylated STAT3
(p-STAT3) is associated with melanoma progression (8,9). STAT3
activity increased the invasive ability of melanoma and is required
for active melanogenesis by regulating tyrosinase gene expression
and enzyme activity (10). The
activation of STAT3 can stimulate brain metastasis of melanoma, and
upregulate the expression of vascular endothelial growth factor
(VEGF) and matrix metalloproteinase-2 (MMP-2) (9). Constitutive STAT3 activation promotes
melanoma growth and angiogenesis by increasing the expression of
VEGF (11). Moreover, cumulative
evidence shows that blockade of STAT3 by small molecule inhibitors
can suppress the growth, angiogenesis and metastasis of melanoma
cells (12–14). Therefore, STAT3 may be a critical
target for melanoma therapy.
Accumulating evidence demonstrates that natural
compounds present in the human diet may significantly alter the
natural history of a malignant tumor (15). Recently, there has been growing
interest in the role of nutrition in melanoma chemoprevention
(16). Olive oil is the main source
of dietary fat in the Mediterranean diet, the consumers of which
have a low incidence of cardiovascular disease, age related
cognitive disease, and cancer (17,18).
(−)-Oleocanthal (OC), a main ingredient contained in virgin olive
oil (VOO), has been reported to possess various biological
activities such as anti-oxidative, anti-bacterial and
anti-inflammatory activities (19–23).
In addition, OC exhibits potent antitumor abilities in many
malignant tumors, including breast carcinoma, prostate carcinoma
and multiple myeloma (24–27). Moreover, OC has also been identified
as a STAT3 inhibitor in human hepatocellular carcinoma (HCC)
(28). However, the role of OC in
melanoma remains unclear. In the present study, we investigated
whether OC inhibited the growth and metastasis of melanoma through
the blockage of STAT3 signaling pathway.
Materials and methods
Cell lines, reagents and
antibodies
A375, A2058, HUVEC and HaCaT cell lines were
purchased from the Cell Resource Center of Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences (Shanghai, China).
All the cells were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco, Grand Island, NY, USA) supplemented with 10% fetal
bovine serum (FBS). Z-VAD-FMK were purchased from Sigma-Aldrich
(St. Louis, MO, USA). Primary antibodies against Bcl-xL, Mcl-1,
cleaved-caspase-3, cleaved-caspase-9, cleaved-PARP, p-STAT3 and
STAT3 were purchased from Cell Signaling Technology (Danvers, MA,
USA). GAPDH and secondary antibodies against mouse IgG-horseradish
peroxidase (HRP) and rabbit IgG-HRP were obtained from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). Antibodies against
Ki-67, CD31, VEGF, MMP-9 and MMP-2 were obtained from Abcam
(Cambridge, MA, USA).
Extraction and isolation of OC
Approximately 2 liters of n-hexane and 1 kg of EVOO
(Sigma-Aldrich) were mixed, and then CH3CN-MeOH (1
liter, 20:80) was added and shaken twice. Dried organic layer (24
g) was subjected to repeated medium pressure liquid chromatography
(MPLC) in a 50×3 cm column on lipophilic Sephadex LH20 (bead size
25–100 µm; Sigma-Aldrich) using
n-hexane-CH2Cl2 (1:9), isocratic elution,
followed by MPLC (10 g, 25×1 cm column) on C-18 reversed-phase
silica gel with Bakerbond octadecyl (40 µm; Mallinckrodt Baker,
Inc.) to afford 13.3 mg of OC with >97% purity (HPLC) along with
several other impure fractions. Identification and purity of 1 were
also based on comparison of its 1H and 13C
NMR data with the literature (29).
Generally, 1:100 ratios of mixtures to be chromatographed versus
the stationary phase were used in all liquid chromatographic
purifications.
Establishment of stable
A375-luciferase cell line
Lentiviral vectors encoding the human Firefly
Luciferase gene were constructed by GeneChem Corporation (Shanghai,
China). The empty vector was used as a negative control. The
lentiviral vectors were transfected into A375 cells with a
multiplicity of infection (MOI) of 60 in the presence of polybrene
(5 µg/ml). At 48 h after transfection, transfected cells were
selected for 2 weeks with 2.5 µg/ml puromycin (Sigma-Aldrich).
Cells, which were obtained 2 weeks after drug selection without
subcloning, were used for further experiments.
Cell viability assay
Cell viability was determined using CCK-8 assay
(Dojindo Laboratories, Kumamoto, Japan) according to the
manufacturer's instruction. Melanoma, HaCaT or HUVEC cells
(3×103) were seeded into a well of a 96-well plate and
cultured in 150 µl of medium supplemented with 10% FBS. After 24 h,
OC (0–60 µM) was added into the culture medium. Then, after the
cells were incubated at 37°C for different times (24 or 48 h), the
medium was changed to 100 µl of medium and 10 µl of CCK-8 reagent.
The cells were incubated for 2 h at 37°C. Finally, the optical
density was measured using an EnSpire™ 2300 Multilabel Reader
(PerkinElmer, Waltham, MA, USA) at 450 nm.
Colony formation assay
Ten thousand A375 cells were cultured in 6-well
plates for 12 days. Fresh medium with OC was added to the plates
every 3 days. At the end of treatment, colonies were stained with
0.05% crystal violet for 30 min and counted.
Cell cycle assay
The cell cycle analysis kit was purchased from BD
Biosciences (San Jose, CA, USA). Melanoma cells were seeded into
6-well plates at 4×105/dish. After incubation with OC
(20 or 40 µM) for 48 h, cells were trypsinized and washed twice
with PBS. Then cells were cultured with Reagents A-C according to
the manufacturer's protocol and subjected to flow cytometry.
Apoptosis assay
The Annexin V-FITC apoptosis kit was purchased from
BD Biosciences. Melanoma cells (4×105 cells/well) were
incubated with OC (20 or 40 µM) for 48 h, 1×104 cells
were collected and washed twice with cold PBS. Then apoptotic cells
were evaluated by double staining with propidium iodide (PI) and
Annexin V labeled with FITC according to the manufacturer's
instruction.
Wound-healing assay
Melanoma cells were seeded at a plating density of
4×105/well in 6-well plates and cultured overnight.
After reaching 100% confluence, the cells were gently scraped with
aplastic tip to produce a wound area. After wounding cells were
incubated with fresh DMEM medium with 10 µM of OC, and the cell
movement throughout the wound area was examined after 24 h.
Migration and invasion assays
Migration assay was performed using BioCoat Chambers
(BD Biosciences). DMEM containing 10% FBS was placed in the lower
chamber as a chemoattractant. Cells (4×105) were seeded
to the upper chamber in 200 µl of serum-free medium containing
different concentrations of OC, and allowed to migrate for 24 h.
Then, the cells on the upper surface of the membrane were removed.
Chambers were fixed in 4% paraformaldehyde for 5 min and stained
with crystal violet. The cells that penetrated the filter were
observed with a microscope and cells from 5 random fields were
counted. Invasion assay was carried out under the same conditions
as the migration assay, except that the chambers were coated with
Matrigel.
Capillary-like tube formation
assay
Tube formation was assessed as described previously
(30). Briefly, HUVECs were
pretreated with various dilutions of OC for 24 h and then seeded
onto the Matrigel layer in 96-well plates at a density of
1×104 cells. After 8–12 h, tubular structure of
endothelial cells was photographed using an inverted
microscope.
Luciferase reporter assay
Luciferase reporter assay was performed as described
previously (30). Luciferase
activity was determined with the Promega luciferase assay kit.
Western blotting
Melanoma cells were plated at a density of
4×105/well on 6-well plates. After incubation with
different concentrations of OC, the cells were washed twice with
ice-cold PBS and treated with 120 µl sample buffer on ice for 30
min. The cell lysate was centrifuged at 12,000 rpm for 10 min at
4°C. Protein lysates (30 µl) were electrophoresed on a 12 or 10%
SDS gel. Then the proteins were electrotransferred to a PVDF
membrane and the membrane was blocked for 30 min with blocking
solution (5% non-fat dry milk in PBS-0.5% Tween-20). The membrane
was then incubated overnight at 4°C with primary antibodies
(1:1,000). After that the membrane was washed in PBST for 30 min,
exposed to HRP-conjugated secondary antibody (diluted 1:2,000), and
washed again in PBST for 30 min. Final detection was performed
using enhanced chemi-luminescence solution for 5 min.
Histopathological analysis
The tumor tissues were isolated to perform
hematoxylin and eosin staining and immunohistochemical staining.
The tumor tissues were immediately fixed with 4% paraformaldehyde,
sectioned, and stained using hematoxylin and eosin for light
microscopy. For immunohistochemical staining, the non-specific
binding was blocked with 1% (w/v) bovine serum albumin at room
temperature for 30 min. The sections were then incubated with
anti-Ki-67 and anti-CD31 antibodies. The slides were incubated with
biotinylated secondary antibody for 1 h at room temperature.
Finally, the sections were developed with DAB color solution (50
µl/section) for 5 min at room temperature. The number of positive
cells were mounted and examined under a light microscope.
Animal experiments
Male BALB/c athymic nude mice (5 weeks old) were
obtained from the experimental animal center of Shanghai Institute
for Biological Sciences (Shanghai, China) and housed under standard
conditions and cared for according to the institutional guidelines
for animal care. All procedures and care administered to the
animals were approved by the Institutional Ethics Committee. We
used six mice in each group to establish subcutaneous xenograft
model. A375 cells (4×106) in 200 µl of PBS were
subcutaneously injected into the flanks of nude mice. After the
tumor volume reached 100 mm3, OC (10 mg/kg/day) was
administered i.p. for 3 weeks. At the end of treatment, tumors were
excised and tumor volume was calculated by using the following
equation: Tumor volume = length × (width)2 × π/6
For the lung metastasis model, A375 cells
(5×106) in 200 µl of PBS were injected into the nude
mice through the tail vein (n=6/group). Animals were randomized to
receive either OC (15 mg/kg/day) or DMSO at 1 week after injection.
Tumor metastasis was imaged and quantified by bioluminescence every
two weeks. After 6 weeks, the mice were sacrificed, and their lungs
were collected for further experiment.
Statistical analysis
Data are presented as mean values ± standard
deviation (SD). Comparisons between multiple groups were performed
using the one-way analysis of variance (ANOVA) followed by Dunnetts
t-test. A value of P<0.05 was considered statistically
significant.
Results
OC suppresses proliferation and colony
formation of melanoma cells
CCK-8 assay was first performed to detect the
anti-proliferative effect of OC on melanoma cells. Melanoma cells
(A375 and A2058 cells) were cultured in different concentrations
(0–60 µM) for 24 h, and then cell viability was measured. OC
significantly inhibited proliferation of melanoma cells in a
dose-dependent manner (Fig. 1A).
Extending drug exposure to 48 h resulted in additional
cytotoxicity, indicating that OC also suppressed proliferation of
melanoma cells in a time-dependent manner (Fig. 1B). In addition, OC treatment showed
lesser toxicity on HaCaT cells (a keratinocyte cell line from adult
human skin), which suggested that OC was more potent to cancer
cells than normal cells (Fig. 1A and
B). Colony formation ability was then investigated to determine
the long-term impact of OC on melanoma cell growth. As is shown in
Fig. 1C and D, the colony formation
ability of A375 cells was significantly inhibited by OC in a
dose-dependent manner.
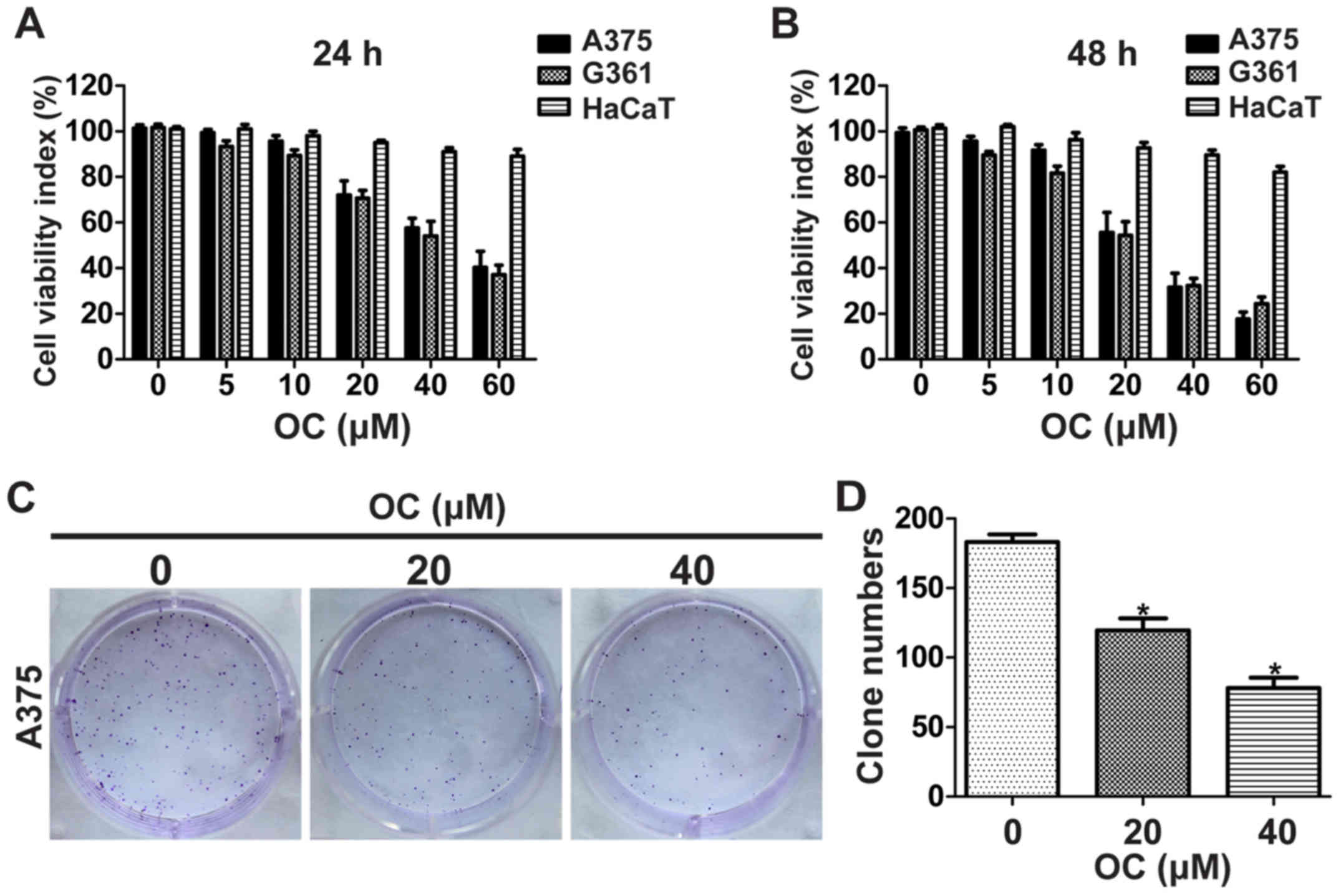 | Figure 1.OC suppresses the viability and
colony formation of melanoma cells. (A) Melanoma and HaCaT cells
were treated with 0, 5, 10, 20, 40 or 60 µM of OC for 24 h, and
cell viability was measured using the CCK-8 assay. (B) Melanoma and
HaCaT cells were treated with 0, 5, 10, 20, 40 or 60 µM of OC for
48 h, and cell viability was measured using the CCK-8 assay. (C)
Representative images of foci formation assay of A375 cells are
shown. (D) The number of foci was counted. Data are presented as
the means ± SD of three independent experiments. *P<0.05. |
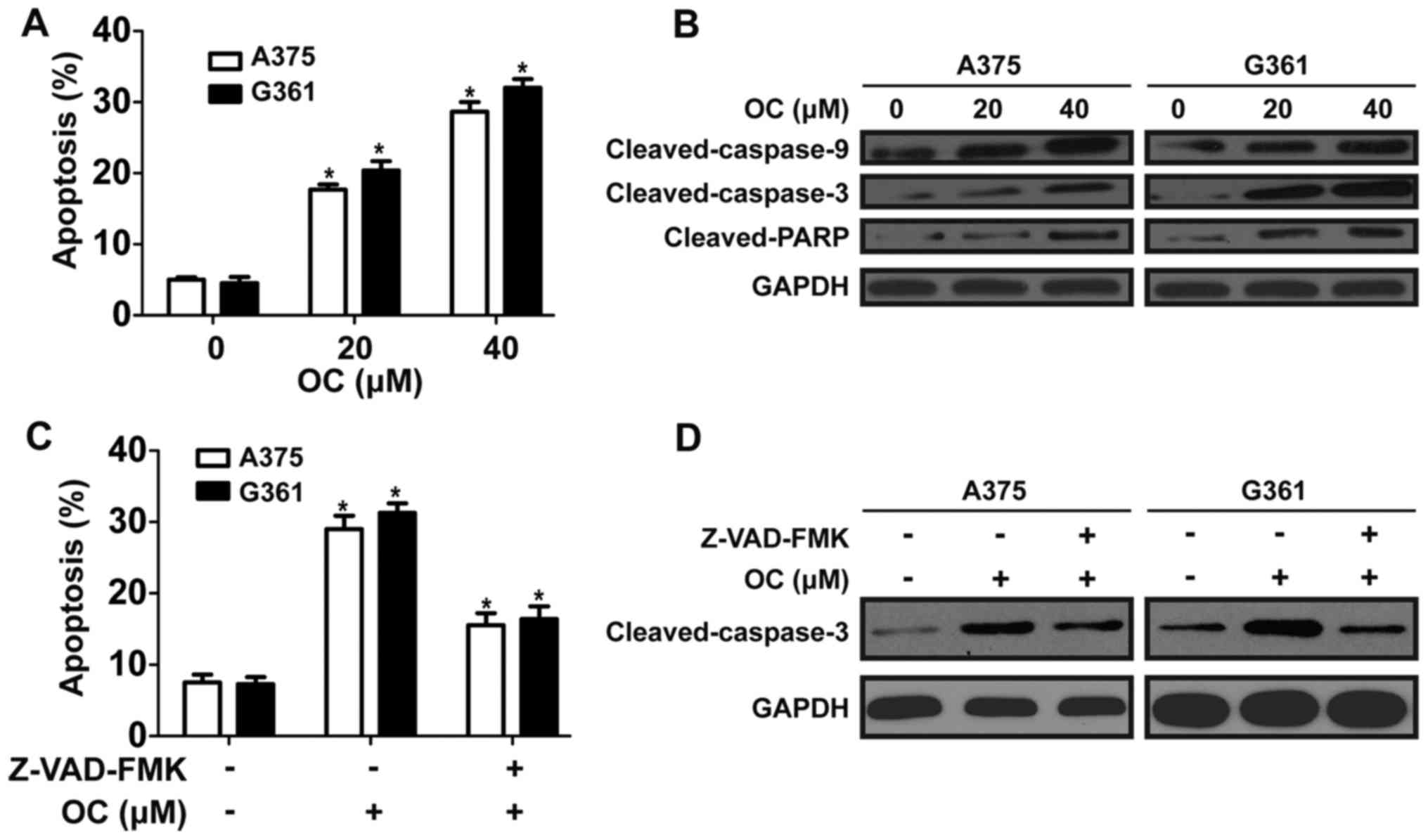
OC induces cell apoptosis in a
caspase-dependent manner
To determine whether anti-proliferation effect
induced by OC involved apoptosis, flow cytometric analysis with
Annexin V-PI staining was performed. OC induced apoptosis of A375
and A2058 cells in a dose-dependent manner after OC treatment for
48 h (Fig. 2A). To investigate the
signaling cascade which mediated OC-induced apoptosis, key proteins
of capase pathway were detected. Results showed that OC
significantly increased the expression of cleaved-caspase-9,
cleaved-caspase-3 and cleaved-PARP in a dose-dependent manner
(Fig. 2B). To explore the
dependence of OC-induced apoptosis on the caspase pathway, melanoma
cells were pretreated with a pan-caspase inhibitor, Z-VAD-FMK (10
mmol/l), before treatment with OC. The pretreatment with Z-VAD-FMK
partially reduced OC-induced apoptosis as determined by Annexin
V-PI staining (Fig. 2C). Western
blotting was performed to determin whether Z-VAD-FMK suppressed
OC-induced activation of caspase-3. Data showed that OC-induced
activation of caspase-3 was partially reversed by pretreatment of
Z-VAD-FMK (Fig. 2D). Then, flow
cytometric analysis was performed to evaluate the effect of OC on
cell cycle progression. However, there was no apparent change in
the distribution of the cell cycle after OC treatment (data not
shown). These data showed that OC induce apoptosis in melanoma
cells, partially in a caspase-dependent manner.
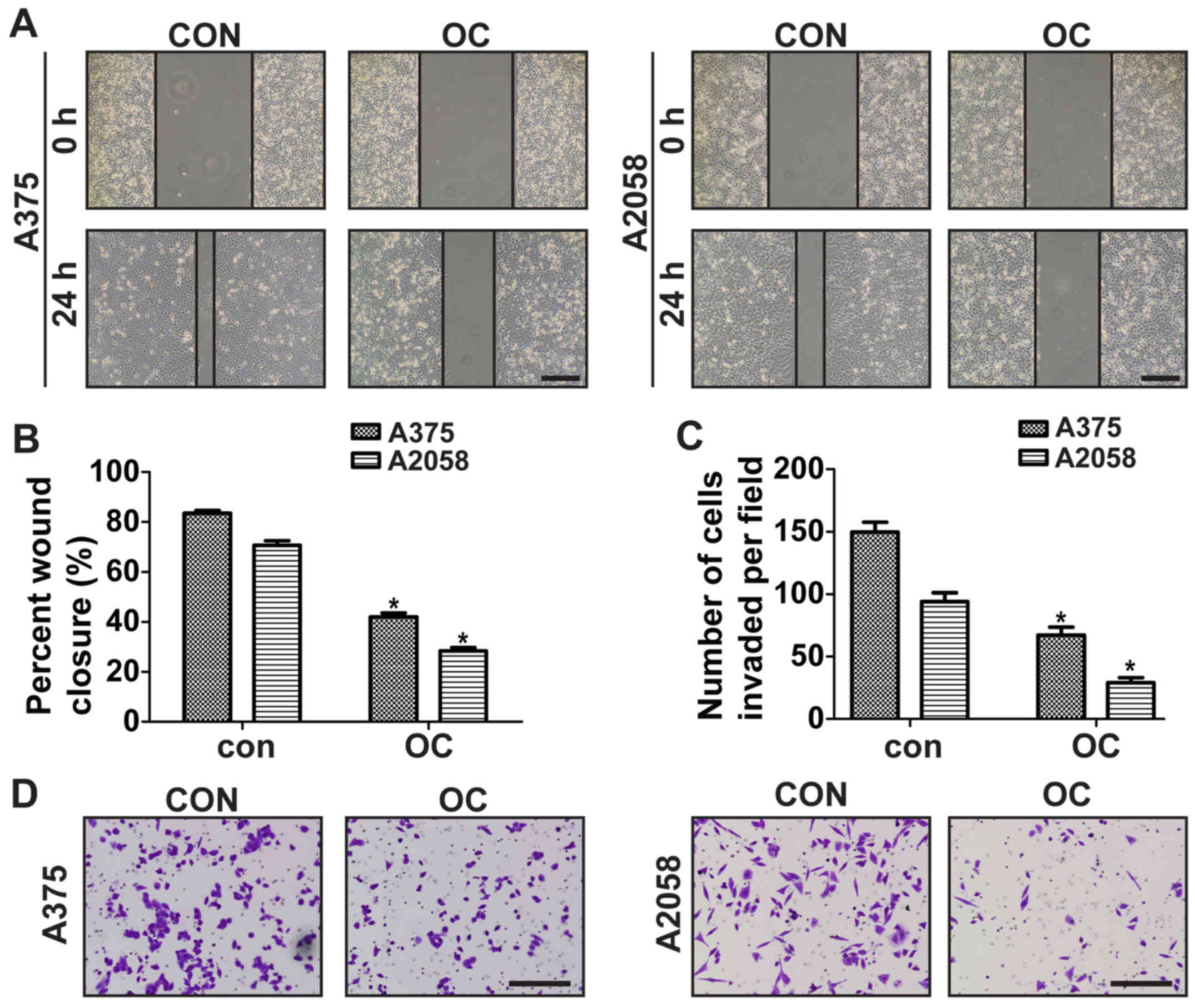
OC inhibits migration and invasion of
melanoma cells
To investigate the effect of OC on melanoma cell
migration, wound-healing assay was performed by using 10 µM of OC
to treat melanoma cells. OC at such doses did not affect cell
proliferation, so its anticancer metastasis effect can be further
studied unambiguously at noncytotoxic doses. Data showed that OC
treatment significantly decreased the migration of A375 and A2058
cells (Fig. 3A and B).
Matrigel-coated Transwell was then used to explore the effect of OC
on the invasion ability of melanoma cells. Data showed that the
invasive ability of A375 and A2058 cells was significantly
suppressed after OC treatment (Fig. 3C
and D).
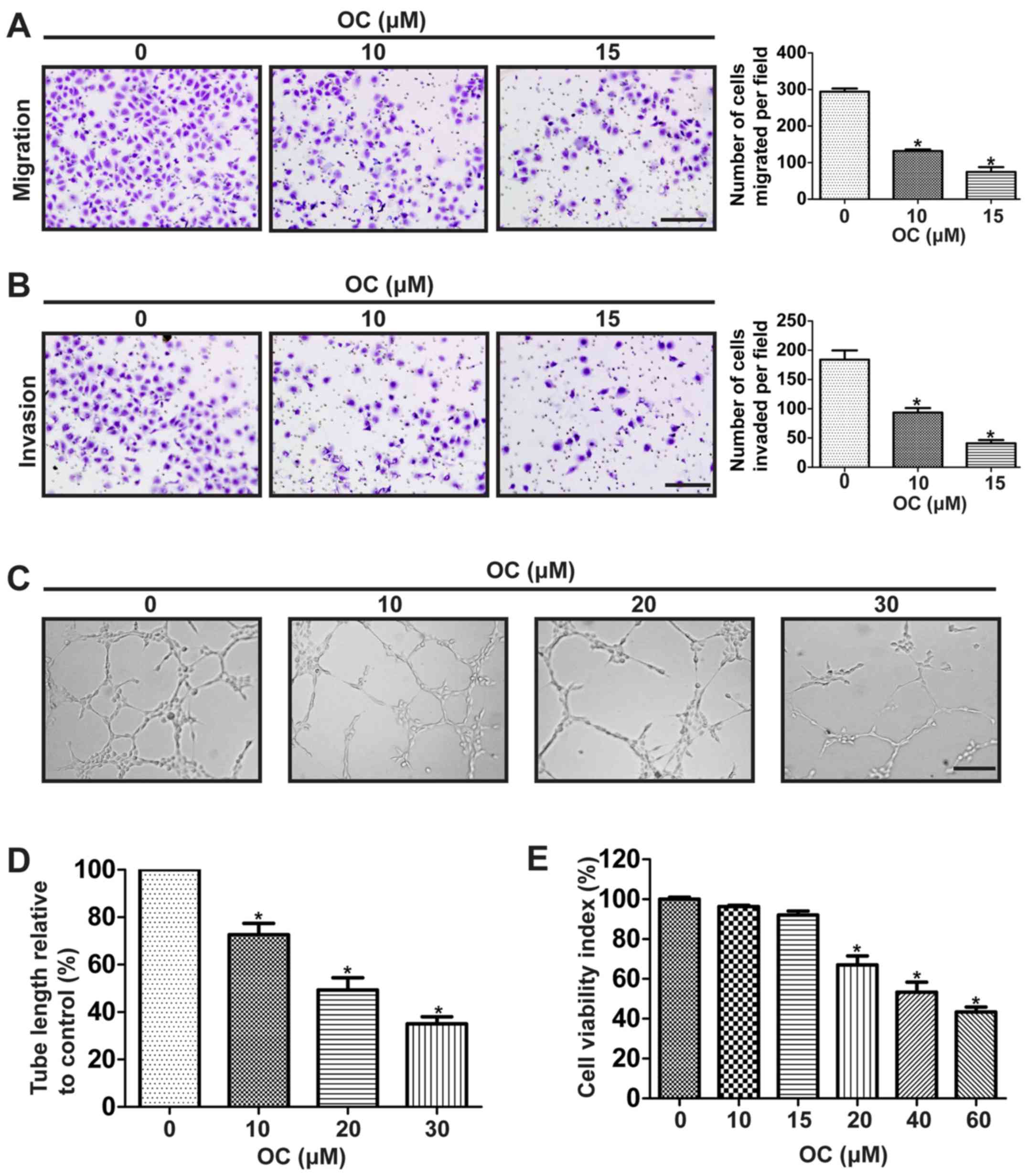
OC suppresses proliferation,
migration, invasion and tube formation of human umbilical vascular
endothelial cells (HUVECs)
Angiogenesis plays a critical role in the growth and
metastasis of malignant tumors as the newly formed tumor
vasculature serve initially as feeding tubes providing nutrients
and oxygen for the tumor mass (31). Therefore, we next investigated the
anti-angiogenic ability of OC. We firstly detected its effect on
the motility of HUVECs using the Transwell assay. Results showed
that OC significantly decreased the migration and invasion
abilities of HUVECs in a dose-dependent manner (Fig. 4A and B). For further insight into
the role of OC on angiogenesis, tube formation assay of HUVECs was
performed. We found that OC significantly suppressed tube formation
ability of HUVECs in a dose-dependent manner (Fig. 4C and D). The process of angiogenesis
also requires the proliferation of endothelial cells, so the effect
of OC on proliferation of HUVECs was also detected. Results showed
that OC suppressed the cell viability of HUVECs in a dose-dependent
manner (Fig. 4E).
OC inhibits constitutive STAT3
phosphorylation, and downregulates the expressions of apoptosis-,
invasion- and angiogenesis-related proteins in melanoma cells
Constitutive activation of STAT3 signaling pathway
has been reported to play a key role in the growth, angiogenesis,
and metastasis of melanoma (8).
Thus, we investigated if OC suppressed the constitutive
phosphorylation of STAT3. OC treatment inhibited the
phosphorylation of STAT3 at the tyrosine 705 (Tyr705) site in a
dose-dependent manner in A375 and A2058 cells, with little impact
on the expression of total STAT3 (Fig.
5A). STAT3 is recognized to be activated by upstream tyrosine
kinases such as JAK2 and Src in melanoma (32). Therefore, we explored if OC
suppressed the activation of JAK2 and Src. OC inhibited the
constitutive phosphorylation of JAK2 and Src in A375 and A2058
cells (Fig. 5A). We next
investigated whether OC could suppress the nuclear localization of
STAT3. After OC treatment, the cytoplasmic and nuclear proteins of
melanoma cells were extracted and expression of STAT3 in both
fractions were determined by western blotting. The levels of STAT3
in nuclear fractions were decreased in a dose-dependent manner
after OC treatment, while those in cytoplasmic fractions were
slightly increased (Fig. 5B). To
detect whether OC affected STAT3-mediated transcriptional activity,
a STAT3-luciferase reporter construct harboring four copies of
STAT3 binding sites was transfected into melanoma cells. Then,
melanoma cells were treated with indicated concentrations of OC for
24 h before testing the transcriptional activity by luciferase
assay. OC inhibited the STAT3-luciferase reporter activity in a
dose-dependent manner (Fig. 5D).
Furthermore, OC also downregulated the STAT3-regulated gene
products, including anti-apoptotic proteins (Bcl-xL and Mcl-1),
invasion-related proteins (MMP-2 and MMP-9), and angiogenesis
protein (VEGF) (Fig. 5C).
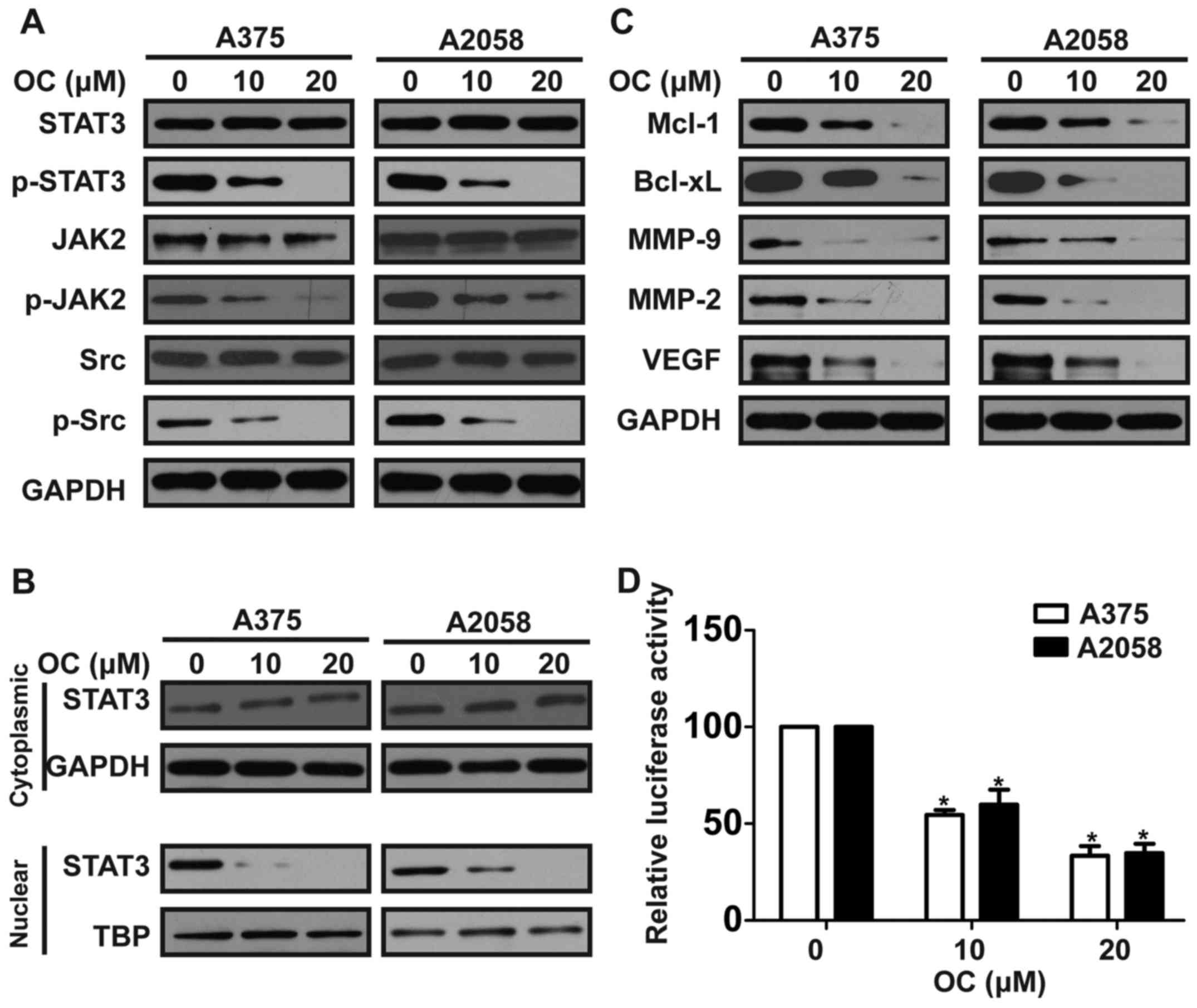 | Figure 5.OC inhibits constitutive STAT3
phosphorylation, and downregulates the expressions of apoptosis-,
invasion- and angiogenesis-related proteins in melanoma cells. (A)
Melanoma cells were treated with various concentrations of OC for
48 h, and then total cell lysates were extracted for western
blotting by using antibodies specific to p-STAT3 (Tyr705), STAT3,
p-Src (Tyr416), Src, p-JAK2 (Y1007/1008) and JAK2. (B) Melanoma
cells were treated with various concentrations of OC for 48 h,
expression levels of STAT3 in cytoplasmic and nuclear extracts were
examined by western blotting. (C) Melanoma cells were treated with
various concentrations of OC for 48 h, and then total cell lysates
were extracted for western blotting by using antibodies specific to
Mcl-1, Bcl-xL, MMP-9, MMP-2 and VEGF. (D) Melanoma cells were
cotransfected with STAT3-Luc reporter and Renilla luciferase
reporter plasmids for 48 h, and then cells were treated with
indicated concentrations of OC for 24 h and the luciferase
intensity was measured. Data are expressed as the mean ± SD of
three independent experiments. *P<0.05. |
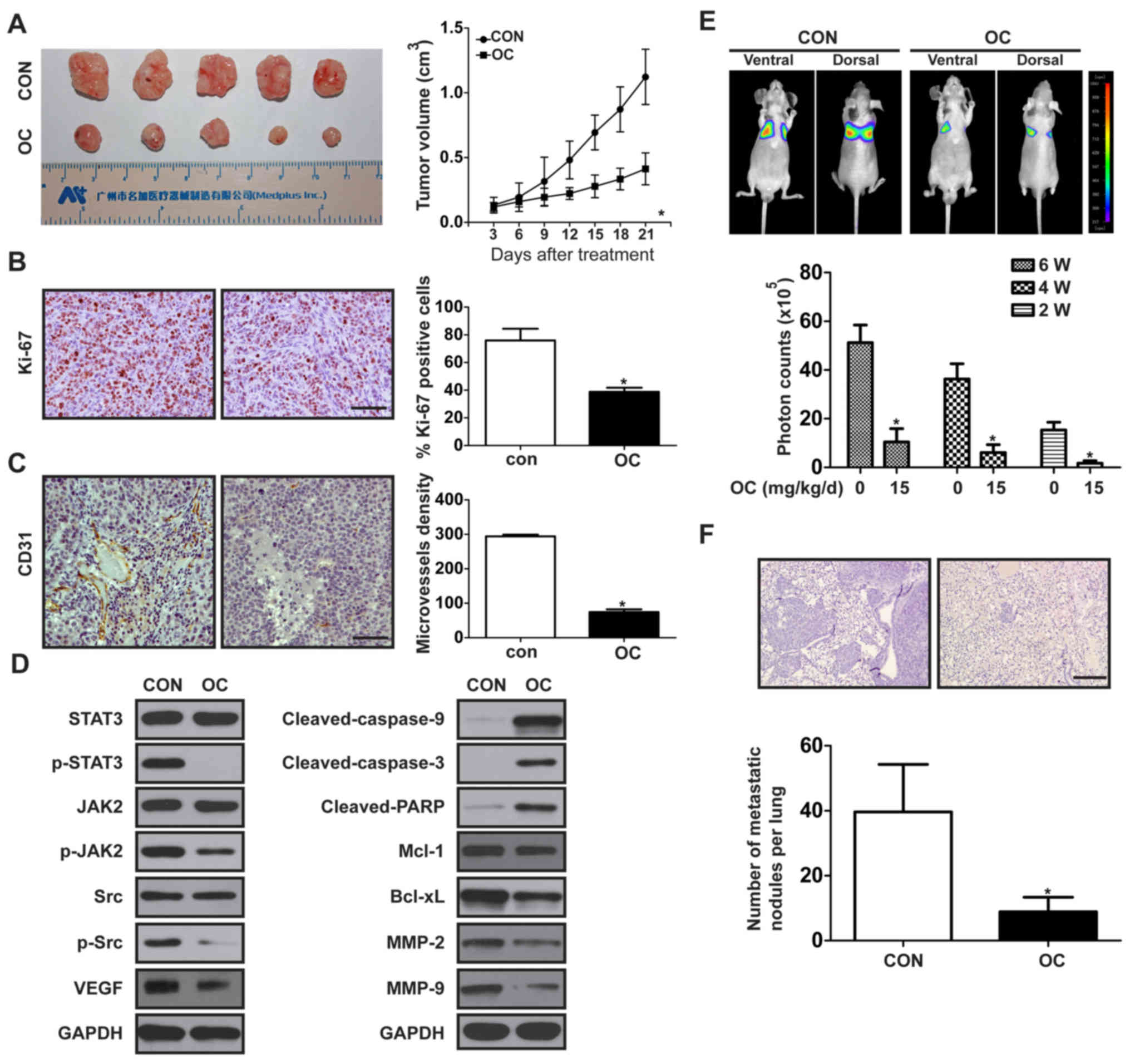
OC inhibits tumor growth and
metastasis of melanoma in vivo
We established a subcutaneous xenograft model in
nude mice using A375 cells to determine the effects of OC on
melanoma growth in vivo. Compared with the control group, OC
treatment resulted in a significant decrease of tumor size
(Fig. 6A). We further investigated
the effect of OC on the expression of Ki-67 (a marker of
proliferation) and CD31 (a marker of angiogenesis) in melanoma
tumor tissues by immunohistochemical analysis and found that OC
significantly decreased the number of Ki-67-positive tumor cells
and the microvessel density, compared with the control group
(Fig. 6B and C). Furthermore, we
investigated expression of tumor-related proteins in nude mice by
using western blotting, and the results were consistent with in
vitro data (Fig. 6D). The
effect of OC on the metastatic ability of melanoma was explored
in vivo by injecting luciferase-expressing A375 cells into
the tail vein of nude mice to establish a lung metastasis model. We
detected stronger illumination signals in control group than that
in the OC-treated group (Fig. 6E).
At the end of treatment, lungs were excised to perform hematoxylin
and eosin staining, and lung micrometastases were microscopically
evaluated. Fewer and smaller metastatic foci were detected in the
OC-treated group, compared with the control group (Fig. 6F).
Discussion
OC is a main phenolic compound contained in VOO, and
has a long history of consumption as part of the Mediterranean
diet. Many biological and pharmacological effects that may be
beneficial to human health have been attributed to OC, including
antitumor activity (21,23,24).
However, the role of OC in melanoma and the potential molecular
mechanisms are not clear. In the present study, we found that OC
suppressed proliferation, migration, invasion, angiogenesis and
induced apoptosis in melanoma in vitro and in vivo.
The underlying mechanisms may be, at least in part, due to the
inhibition of STAT3 activation and downregulation of its gene
products.
OC are thought to exhibit anti-proliferation effects
on cancer cells due to its ability to induce cell cycle arrest and
apoptosis. Akl et al reported that OC arrests breast cancer
cells at G1/M phase by modulating the expression of CDK6, cyclin
D1, p21 and p27 (25). Moreover, OC
induces cell apoptosis by activating caspase pathway in MDA-MB-231
cells (25). Based on our study, OC
suppressed proliferation and promoted apoptosis in melanoma cells
in vitro, but showed no apparent effect on the cell cycle.
Furthermore, the slight effect of OC on HaCaT cells in the
treatment suggests that it is a compound that specifically
suppresses the proliferation of melanoma with negligible side
effects. In addition, OC treatment increased the expression of
cleaved-caspase-3, cleaved-caspase-9 and cleaved-PARP, which are
critical factors in the caspase pathway. More importantly, a
pan-caspase inhibitor (Z-VAD-FMK) can reverse the effects of OC on
apoptosis and the expression of cleaved-caspase-3 in melanoma
cells. These data showed that OC induces apoptosis in melanoma
cells, partially in a caspase-dependent manner.
Angiogenesis, the formation of nascent blood vessels
from pre-existing vasculature, is vital for tumor growth and
metastasis of malignant tumors (31). This process supplies adequate oxygen
and nutrition to support the growth of tumor mass, and later aid in
the initiation of metastasis, which contributes to >90% of
deaths in various cancers, including melanoma. Angiogenesis is
initiated by endothelial cell proliferation, migration, invasion
and then tube formation (33). It
has been shown that inhibition at any step of the processes in
angiogenesis will suppress the formation of new blood vessels
(34). In the present study, we
found that OC significantly suppressed proliferation, migration,
invasion and tube formation of HUVECs in vitro, and
downregulated the expression of CD31 in the subcutaneous xenograft
model. These data suggest that OC may be a promising
anti-angiogenic drug with significant antitumor activity in
melanoma.
As a previous study reported that OC inhibited the
growth and metastasis of HCC through the block of STAT3 signaling
pathway (28), we detected the
effect of OC on the activation of STAT3. Our data showed that OC
significantly decreased the expression of p-STAT3, STAT3 nuclear
translocation and STAT3 transcriptional activity in melanoma cells.
STAT3 is reported to be activated by upstream tyrosine kinases such
as JAK2 and Src in melanoma (32),
thus we further explored the expression of JAK2, Src and their
phosphorylated forms after OC treatment. OC was found be able to
reduce the expression of p-JAK2 and p-Src in a dose-dependent
manner in melanoma cells, with little impact on the expression of
total JAK2 and Src. In addition, OC also reduced the expression of
the STAT3-regulated gene products, which are associated with
apoptosis, invasion and angiogenesis.
The anti-apoptotic genes Mcl-1 and Bcl-xL are
reported to be upregulated in the progression of melanoma (35). In patient samples, the expressions
of Bcl-xL and Mcl-1 were positively corrected with p-STAT3
expression (36). Suppression of
STAT3 can result in the decrease of Mcl-1 and Bcl-xL (37). In this study, we demonstrated that
OC suppressed the expressions of Mcl-1 and Bcl-xL in a
dose-dependent manner. This suggested that other than activating
the caspase pathway, OC-induced apoptosis in melanoma cells was at
least partly by inhibiting the expression of Bcl-xL and Mcl-1.
VEGF is a key pro-angiogenic factor that plays a key
role in tumor angiogenesis. High serum VEGF values are associated
with shorter disease-free survival of melanoma patient (38). STAT3 has been reported to directly
participate in regulating transcription of VEGF gene (11). According to our results, OC
treatment reduced the expression of VEGF in melanoma cells,
suggesting that the anti-angiogenesis effect of OC on melanoma is
associated with VEGF inhibition.
MMP-2 and MMP-9 are invasion-related proteins by
digesting the extracellular matrix surrounding the tumor tissue.
Higher expression of MMP-2 is observed in melanoma patients and
associated with metastasis of melanoma (39). High serum level of MMP-9 is
associated with rapid progression in patients with metastatic
melanoma (40). Moreover, it has
been reported that inactivation of STAT3 significantly impairs the
invasive ability of melanoma cells through decreasing MMP-2
expression (41). Therefore, it is
reasonable to deduce that OC suppressed the migration and invasion
of melanoma by reducing the expressions of MMP-2 and MMP9. In line
with the deduction, we found that OC also reduced the expressions
of MMP-9 and MMP2. These results indicated that anti-migration and
anti-invasion effects of OC on melanoma can be attributed to
suppression of MMP-9 and MMP-2.
In conclusion, we showed that OC significantly
suppress the constitutive STAT3 activation, which may lead to the
suppression of growth and metastasis of melanoma. Although the role
of OC on melanoma still needs to be investigated in clinical
trials, we propose that OC may be a potential agent for prevention
and therapy of melanoma.
Glossary
Abbreviations
Abbreviations:
|
OC
|
(−)-oleocanthal
|
|
VEGF
|
vascular endothelial growth factor
|
|
MMP-2
|
matrix metalloproteinase-2
|
|
MMP-9
|
matrix metalloproteinase-9
|
|
HCC
|
hepatocellular carcinoma
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cummins DL, Cummins JM, Pantle H,
Silverman MA, Leonard AL and Chanmugam A: Cutaneous malignant
melanoma. Mayo Clin Proc. 81:500–507. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chi Z, Li S, Sheng X, Si L, Cui C, Han M
and Guo J: Clinical presentation, histology, and prognoses of
malignant melanoma in ethnic Chinese: A study of 522 consecutive
cases. BMC Cancer. 11:852011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hauschild A, Grob JJ, Demidov LV, Jouary
T, Gutzmer R, Millward M, Rutkowski P, Blank CU, Miller WH Jr,
Kaempgen E, et al: Dabrafenib in BRAF-mutated metastatic melanoma:
A multicentre, open-label, phase 3 randomised controlled trial.
Lancet. 380:358–365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Callahan MK, Postow MA and Wolchok JD:
Immunomodulatory therapy for melanoma: Ipilimumab and beyond. Clin
Dermatol. 31:191–199. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Poust JC, Woolery JE and Green MR:
Management of toxicities associated with high-dose interleukin-2
and biochemotherapy. Anticancer Drugs. 24:1–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lopez-Bergami P, Fitchman B and Ronai Z:
Understanding signaling cascades in melanoma. Photochem Photobiol.
84:289–306. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kortylewski M, Jove R and Yu H: Targeting
STAT3 affects melanoma on multiple fronts. Cancer Metastasis Rev.
24:315–327. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie TX, Huang FJ, Aldape KD, Kang SH, Liu
M, Gershenwald JE, Xie K, Sawaya R and Huang S: Activation of stat3
in human melanoma promotes brain metastasis. Cancer Res.
66:3188–3196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Messina JL, Yu H, Riker AI, Munster PN,
Jove RL and Daud AI: Activated stat-3 in melanoma. Cancer control.
15:196–201. 2008.PubMed/NCBI
|
|
11
|
Niu G, Wright KL, Huang M, Song L, Haura
E, Turkson J, Zhang S, Wang T, Sinibaldi D, Coppola D, et al:
Constitutive Stat3 activity up-regulates VEGF expression and tumor
angiogenesis. Oncogene. 21:2000–2008. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu L, Nam S, Tian Y, Yang F, Wu J, Wang
Y, Scuto A, Polychronopoulos P, Magiatis P, Skaltsounis L, et al:
6-Bromoindirubin-3-oxime inhibits JAK/STAT3 signaling and induces
apoptosis of human melanoma cells. Cancer Res. 71:3972–3979. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu L, Kritsanida M, Magiatis P, Gaboriaud
N, Wang Y, Wu J, Buettner R, Yang F, Nam S, Skaltsounis L, et al: A
novel 7-bromoindirubin with potent anticancer activity suppresses
survival of human melanoma cells associated with inhibition of
STAT3 and Akt signaling. Cancer Biol Ther. 13:1255–1261. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kamran MZ and Gude RP: Pentoxifylline
inhibits melanoma tumor growth and angiogenesis by targeting STAT3
signaling pathway. Biomed Pharmacother. 67:399–405. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Martínez ME, Marshall JR and Giovannucci
E: Diet and cancer prevention: The roles of observation and
experimentation. Nat Rev Cancer. 8:694–703. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tong LX and Young LC: Nutrition: The
future of melanoma prevention? J Am Acad Dermatol. 71:151–160.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fortes C, Forastiere F, Farchi S, Mallone
S, Trequattrinni T, Anatra F, Schmid G and Perucci CA: The
protective effect of the Mediterranean diet on lung cancer. Nutr
Cancer. 46:30–37. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bosetti C, Negri E, Franceschi S, Talamini
R, Montella M, Conti E, Lagiou P, Parazzini F and La Vecchia C:
Olive oil, seed oils and other added fats in relation to ovarian
cancer (Italy). Cancer Causes Control. 13:465–470. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Monti MC, Margarucci L, Tosco A, Riccio R
and Casapullo A: New insights on the interaction mechanism between
tau protein and oleocanthal, an extra-virgin olive-oil bioactive
component. Food Funct. 2:423–428. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Monti MC, Margarucci L, Riccio R and
Casapullo A: Modulation of tau protein fibrillization by
oleocanthal. J Nat Prod. 75:1584–1588. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lucas L, Russell A and Keast R: Molecular
mechanisms of inflammation. Anti-inflammatory benefits of virgin
olive oil and the phenolic compound oleocanthal. Curr Pharm Des.
17:754–768. 2011.
|
|
22
|
Cicerale S, Breslin PA, Beauchamp GK and
Keast RS: Sensory characterization of the irritant properties of
oleocanthal, a natural anti-inflammatory agent in extra virgin
olive oils. Chem Senses. 34:333–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Romero C, Medina E, Vargas J, Brenes M and
De Castro A: In vitro activity of olive oil polyphenols against
Helicobacter pylori. J Agric Food Chem. 55:680–686. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Busnena BA, Foudah AI, Melancon T and El
Sayed KA: Olive secoiridoids and semisynthetic bioisostere
analogues for the control of metastatic breast cancer. Bioorg Med
Chem. 21:2117–2127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Akl MR, Ayoub NM, Mohyeldin MM, Busnena
BA, Foudah AI, Liu YY and Sayed KA: Olive phenolics as c-Met
inhibitors: (−)-Oleocanthal attenuates cell proliferation,
invasiveness, and tumor growth in breast cancer models. PLoS One.
9:e976222014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Elnagar AY, Sylvester PW and El Sayed KA:
(−)-Oleocanthal as a c-Met inhibitor for the control of metastatic
breast and prostate cancers. Planta Med. 77:1013–1019. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scotece M, Gómez R, Conde J, Lopez V,
Gómez-Reino JJ, Lago F, Smith AB III and Gualillo O: Oleocanthal
inhibits proliferation and MIP-1α expression in human multiple
myeloma cells. Curr Med Chem. 20:2467–2475. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pei T, Meng Q, Han J, Li HS, Song R, Sun
B, Pan S, Liang D and Liu L: (−)-Oleocanthal inhibits growth and
metastasis by blocking activation of STAT3 in human hepatocellular
carcinoma. Oncotarget. Jun 2–2016.(Epub ahead of print).
|
|
29
|
Smith AB III, Han Q, Breslin PA and
Beauchamp GK: Synthesis and assignment of absolute configuration of
(−)-oleocanthal: A potent, naturally occurring non-steroidal
anti-inflammatory and anti-oxidant agent derived from extra virgin
olive oils. Org Lett. 7:5075–5078. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song R, Song H, Liang Y, Yin D, Zhang H,
Zheng T, Wang J, Lu Z, Song X, Pei T, et al: Reciprocal activation
between ATPase inhibitory factor 1 and NF-κB drives hepatocellular
carcinoma angiogenesis and metastasis. Hepatology. 60:1659–1673.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ferrara N and Kerbel RS: Angiogenesis as a
therapeutic target. Nature. 438:967–974. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Al Zaid Siddiquee K and Turkson J: STAT3
as a target for inducing apoptosis in solid and hematological
tumors. Cell Res. 18:254–267. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fan TP, Yeh JC, Leung KW, Yue PY and Wong
RN: Angiogenesis: From plants to blood vessels. Trends Pharmacol
Sci. 27:297–309. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tournaire R, Simon MP, le Noble F,
Eichmann A, England P and Pouysségur J: A short synthetic peptide
inhibits signal transduction, migration and angiogenesis mediated
by Tie2 receptor. EMBO Rep. 5:262–267. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hartman ML and Czyz M: Anti-apoptotic
proteins on guard of melanoma cell survival. Cancer Lett.
331:24–34. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhuang L, Lee CS, Scolyer RA, McCarthy SW,
Zhang XD, Thompson JF and Hersey P: Mcl-1, Bcl-XL and Stat3
expression are associated with progression of melanoma whereas
Bcl-2, AP-2 and MITF levels decrease during progression of
melanoma. Mod Pathol. 20:416–426. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Niu G, Bowman T, Huang M, Shivers S,
Reintgen D, Daud A, Chang A, Kraker A, Jove R and Yu H: Roles of
activated Src and Stat3 signaling in melanoma tumor cell growth.
Oncogene. 21:7001–7010. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ascierto PA, Leonardi E, Ottaiano A,
Napolitano M, Scala S and Castello G: Prognostic value of serum
VEGF in melanoma patients: A pilot study. Anticancer Res.
24:4255–4258. 2004.PubMed/NCBI
|
|
39
|
Rotte A, Martinka M and Li G: MMP2
expression is a prognostic marker for primary melanoma patients.
Cell Oncol (Dordr). 35:207–216. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nikkola J, Vihinen P, Vuoristo MS,
Kellokumpu-Lehtinen P, Kähäri VM and Pyrhönen S: High serum levels
of matrix metalloproteinase-9 and matrix metalloproteinase-1 are
associated with rapid progression in patients with metastatic
melanoma. Clin Cancer Res. 11:5158–5166. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xie TX, Wei D, Liu M, Gao AC, Ali-Osman F,
Sawaya R and Huang S: Stat3 activation regulates the expression of
matrix metalloproteinase-2 and tumor invasion and metastasis.
Oncogene. 23:3550–3560. 2004. View Article : Google Scholar : PubMed/NCBI
|