Introduction
Thyroid cancer (TC) is the most common malignancy of
the endocrine system, and the incidence of TC rapidly increases
worldwide each year. Thyroid carcinoma can be divided into
papillary, follicular, medullary and anaplastic types according to
histological type (1). Papillary is
the most common thyroid carcinoma, accounting for >83% of all
such malignancies (2). In recent
years, the prognosis of TC patients has shown improvement due to
advances in chemotherapy and radiotherapy. However, growing
evidence demonstrates that patients with advanced TC are resistant
to uptake of radioiodine or surgical resection, which leads to the
disease recurrence or even death (3). Therefore, it is urgent to investigate
novel strategies for the diagnosis and treatment of TC in early
stage.
Tripartite motif-containing 29 (TRIM29), also known
as ataxia-telangiectasia group D-associated protein (ATDC), belongs
to the TRIM family, TRIM family is composed of evolutionary
conserved N-terminal tripartite motif, such as B-box, RING finger
and coiled-coil (RBCC) domain (4,5). The
TRIM family members have been proved to be involved in various
physiological and biological processes including cell apoptosis,
proliferation and oncogenesis, once altered, pathological
conditions will occur (6). The
structure of TRIM29 is dissimilar to other TRIM protein members, it
has no RING domain, indicating no E3 ubiquitin ligase activity of
TRIM29 functions. Recent studies have reported that overexpression
of TRIM 29 is associated with tumor progression and poor prognosis
in a variety of cancers, such as lung cancer, gastric cancer,
pancreatic adenocarcinoma, bladder cancer, ovarian cancer and
endometrial neoplasms (7–9). Liu et al have verified that
TRIM29 knockdown inhibited cell proliferation and invasion in human
squamous cancer cells (10).
Moreover, in colorectal cancer, upregulation of TRIM29 is
associated with advanced tumor stage, histological differentiation,
and predicts poor survival (11).
In contrast, the tumor suppressor role of TRIM29 is also reported
in prostate and breast cancer (12,13).
However, the biological function and molecular details in TC
progression has not been well established.
In the present study, to illustrate the underlying
molecular mechanisms of TRIM29 in TC development, TRIM29 expression
level was evaluated in TC tumor samples and cell lines. We found
that overexpression of TRIM29 was correlated with
clinicopathological parameters and poor prognosis in TC patients.
TRIM29 knocking down by siRNA method led to the inhibition of cell
proliferation by arresting the cell cycle and inducing cell
apoptosis, and further confirmed that the P13K/AKT signaling
pathway was involved in the mediation of TRIM29 in TC
development.
Materials and methods
Collection of tissue samples
In this study, 56 thyroid cancer (TC) tissues and
paired adjacent normal tissues were obtained from TC patients who
underwent total thyroidectomy at the Department of General Surgery,
the Second Affiliated Hospital of Xi'an Jiaotong University, Xi'an,
China, between November 2009 and September 2014. All TC samples
were separately diagnosed and confirmed by two pathologists. Tumor
stage was performed according to the sixth edition of the TNM
(tumor, node, and metastasis) classification of the International
Union Against Cancer. The clinical characteristics of patients
including gender, age, TNM stage, tumor size, extrathyroidal
extension, regional lymph nodes and distant metastasis were
collected. This study was approved by the Ethics Committee of the
Second Affiliated Hospital of Xi'an Jiaotong University and
accordance with the Declaration of Helsinki, and the informed
consents were obtained from all patients. The expression level of
TRIM29 in tissue samples lower than the median level were
considered as the low TRIM29 expression group and vice versa. A
five-year survival time and outcomes were recorded in follow-up.
The median duration of follow-up was 38 months (range: 1–60
months).
Cell line and cell culture
The human TC cell lines TT, TPC-1, K1, and human
normal thyroid cell line Nthy-Ori-3-1 were purchased from the Cell
Bank of the Chinese Academy of Medical Science (Beijing, China).
All cells were cultured in DMEM medium (Invitrogen, Carlsbad, CA,
USA) supplemented with 10% fetal bovine serum (Gibco, Carlsbad, CA,
USA) and 1% penicillin/streptomycin under an atmosphere of 5%
CO2 at 37°C.
Silencing TRIM29 expression in TC
cells by siRNA
The siRNA (small interfence RNA) method was employed
to knock down TRIM29 expression in TC cell lines. The sequences for
siRNA were design as follows: sense strand
5′-AGUAGUUGGAGUUCUUGUCGU-3′, antisense strand
5′-GACAAGAACUCCAACUACUUC-3′. The scramble siRNA was random
sequenced by Blast website. Then the recombinant plasmids
(si-TRIM29) or scramble siRNA (si-NC) was transfected into TC cells
using Lipofectamine 2000 (Invitrogen) according to the
manufacturer's instructions. The efficacy of siRNA knockdown was
determined by RT-qPCR and western blotting.
Real-time quantitative PCR
analysis
Total RNA was extracted from either tissue samples
or cells using TRIzol (Invitrogen) according to the manufacturer's
protocol, and then the isolated mRNA were reversely transcribed
into cDNA by M-MLV Reverse transcriptase kit (Promega, Madison, WI,
USA) according to the manufacturer's protocol. RT-qPCR analysis was
performed by SYBR-Green Master Mix kit (Takara, Otsu, Japan). The
primer sequences were as follows: TRIM29 sense,
5′-ATGCTTGGTGGTCACTTTGG-3′ and antisense,
5′-GCACTTCCCTTACCAGCATAG-3′; β-actin sense,
5′-CAGCATCATGAAGTGCGACG-3′ and antisense,
5′-CTGCAGTGTGGTGCTTCAAC-3′. The mRNA expression of TRIM29 was
normalized to the housekeeping gene β-actin, which was used as an
internal control. In brief, the PCR conditions were as follows:
95°C for 3 min, 95°C for 15 sec, 65°C for 30 sec for 40 cycles. The
relative quantification referred as relative expression level was
calculated by the value of 2−∆∆Ct. Each experiment was
performed in triplicate.
Western blot analysis
Tissue samples or cells were washed twice with
ice-cold PBS and lysed with RIPA lysis buffer (Beyotime, Haimen,
China). BCA assay was used to quantify the protein concentration of
cell lysates. Total proteins were separated by 15%
SDS-polyacrylamide (SDS-PAGE) and then transferred onto PVDF
membranes (Pierce, Thermo Scientific, Rockford, IL, USA). After
blocking with 5% BSA for 1 h, the membranes were incubated with
appropriate primary antibodies at 4°C overnight. Next, the
membranes were incubated with goat anti-rabbit HRP-conjugated
secondary antibody (diluted at 1:2000) (Santa Cruz Biotechnology)
at 37°C for 1 h. The protein bands were detected using the Enhanced
Chemiluminescence Detection System (Pierce, Thermo Scientific).
β-actin was used for normalization and the protein intensity was
quantified by using Image-Pro Plus 6.0 software (Media Cybernetics,
MD, USA).
Cell viability assay
Cell proliferation was determined by the MTT
[3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide]
(Beyotime) assay according to the manufacturer's protocol. Cells
were seeded in 96-well plates at the density of 5×103
cells/well after transfected with siRNA-TRIM29 or siRNA-NC for 48 h
and incubate at 37°C, 5% CO2 for 5 days. For
chemotherapy drug sensitivity test, the transfected cells were
incubated with cisplatin (Sigma-Aldrich, St. Louis, MO, USA) at the
doses of 0, 5, 10, 15, 20 µg/ml for 24 h. At indicated time points,
20 µl MTT solution (5 mg/ml) was added into each well and incubated
for 4 h at 37°C. Dimethyl sulfoxide (DMSO) (150 µl) was added into
the well to dissolve the crystals for 10 min on a low-speed shaker.
The OD (optical density) value was measured each day at 490 nm
using a microplate reader (BioTek, Winooski, VT, USA).
Colony formation assay
Cells were seeded at the density of 500 cells/well
in 6-well plate after transfected with siRNA-TRIM29 or siRNA-NC for
48 h, and then incubated at 37°C, 5% CO2 for 14 days.
Colonies were fixed with 95% ethanol for 10 min after washing with
PBS twice, and then stained with crystal violet for 20 min,
colonies with at least 50 cells were counted and photographed. Each
experiment was performed in triplicate.
Cell cycle analysis
Cells were harvested after transfected with
siRNA-TRIM29 or siRNA-NC for 48 h and fixed in 75% ethanol
overnight, and then washed with PBS, stained by propidium iodide (5
mg/ml) containing RNase A. Cell cycle distributions were analyzed
by flow cytometer FACSCalibur (Beckman Coulter, Inc., Fullerton,
CA, USA) according to the manufacturer's protocol. Each experiment
was performed in triplicate.
Cell apoptosis analysis
Cells were harvested after transfected with
siRNA-TRIM29 or siRNA-NC for 48 h, and then washed with cold PBS,
and stained with Annexin V-FITC double staining kit (BD Bioscience,
San Jose, CA, USA) and analyzed using flow cyctometry FACSCalibur
(Beckman Coulter, Inc.) according to the manufacturer's protocol.
Each experiment was performed in triplicate.
Cell migration and invasion assay
Cells were harvested after transfected with
siRNA-TRIM29 or siRNA-NC for 48 h, and then suspended with
serum-free medium, then seeded in the upper chamber of Transwell (8
µm pore size, Corning Inc., Coring, NY, USA) at the density of
5×104 cells per chamber for invasion assay and
1×105 cells per chamber for migration assay,
respectively. For invasion assay, the upper chambers were precoated
with Matrigel (BD Bioscience). The lower chambers were filled with
medium containing 10% FBS and incubated for 24 h. Cells remaining
on the upper chamber were removed carefully, the migrating cells in
the lower chamber were fixed with 100% methanol, and stained with
crystal violet. The stained cells were imaged with an inverted
microscope (Olympus), and counted at ten randomly selected visual
fields for each experimental condition. Each experiment was
performed in triplicate.
Statistical analysis
Statistical analyses were performed using SPSS-12.0
software. Each experiment was performed in triplicate, unless
otherwise indicated. All data are presented as mean ± standard
deviation (SD). Comparisons between groups were done using the
unpaired Student's t-test. A P-value of <0.05 was considered to
indicate a statistically significant difference.
Results
Elevated expression of TRIM29 in TC
tissue samples and cell lines
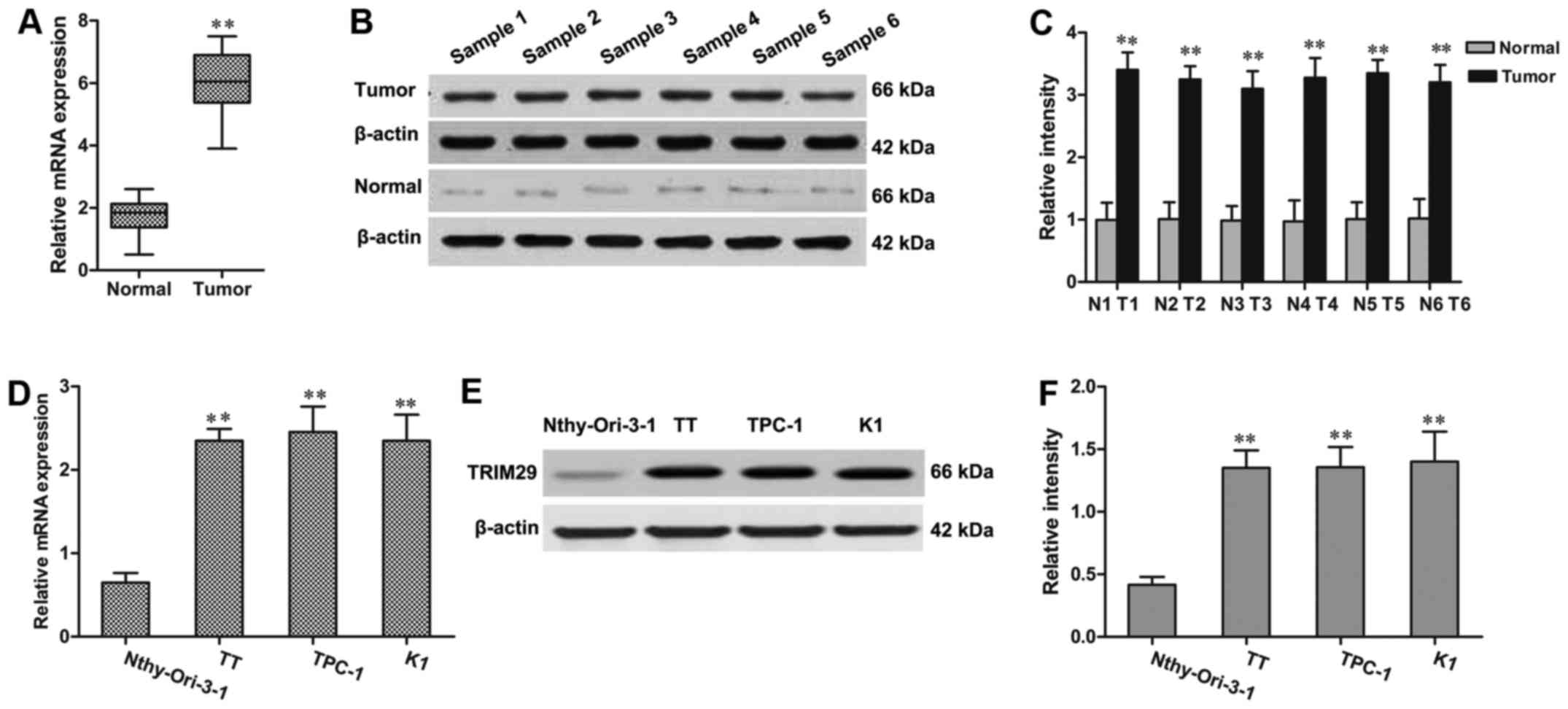
RT-qPCR and western blot assays were used to detect
the expression of TRIM29 in TC tissue samples and adjacent paired
normal tissues. As shown in Fig.
1A, the mRNA expression of TRIM29 was significantly upregulated
in TC tissue samples compared with the adjacent paired normal
tissues. Consistent with RT-qPCR results, the western blot also
revealed that TRIM29 was upregulated at protein levels in TC
samples (Fig. 1B and C).
Furthermore, the expression level of TRIM29 was evaluated among
three TC cell lines (TT, TPC-1, and K1) and normal thyroid cell
line Nthy-Ori-3-1. The result showed that TRIM29 expression was
remarkably increased in TT, TPC-1, and K1 cells when compared to
Nthy-Ori-3-1 cell line (Fig.
1D-F).
TRIM29 expression correlates with
clinicopathological parameters on the prognosis of TC patients
The correlation between TRIM29 expression and
clinicopathological para-meters of all patients were investigated
by Peason's Chi-square analysis. TC cases were subdivided into two
groups according to the mRNA expression level of TRIM29 as
described above. High TRIM29 expression was observed in 73.21%
(41/56) of cases, while 26.79% (15/56) had low TRIM29 expression.
The high TRIM29 expression was associated with TNM stage
(P<0.01), extrathyroidal extension (P<0.01), lymph nodes
metastasis (P<0.05), and distant metastasis (P<0.05), but had
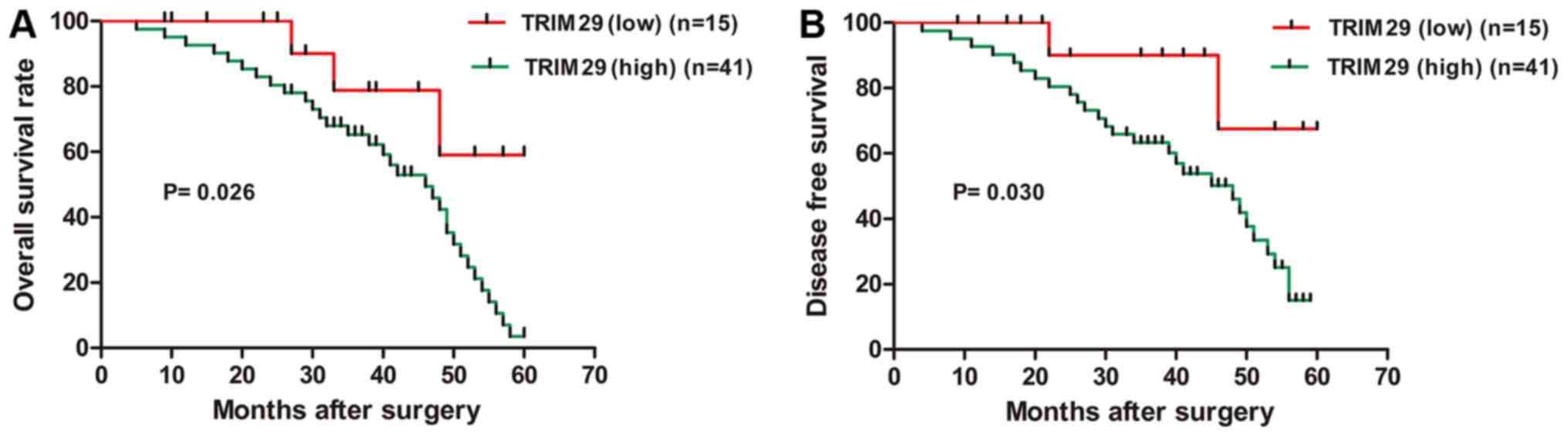
no correlation with patient gender, age or tumor size (Table I). In addition, the survival time of
the two groups was compared by Kaplan-Meier analysis, the
cumulative 5-year overall survival rate (OS) was 21.95% in the high
TRIM29 expression group whereas 80.00% in the low TRIM29 expression
group (P<0.05); while the disease-free survival rate (DFS) was
34.15% in the high TRIM29 expression group which was lower than
86.67% in the low TRIM29 expression group (P<0.05) (Fig. 2). These findings demonstrated that
the TRIM29 overexpression was correlated with poor prognosis in TC
patients.
 | Table I.TRIM29 expression and
clinicopathological factors in TC patients. |
Table I.
TRIM29 expression and
clinicopathological factors in TC patients.
|
|
| TRIM29
expression |
|
|---|
| Variables | Case | Low (n=15) | High (n=41) | P-value |
|---|
| Age | 56 | 15 | 41 | 0.431 |
|
>45 | 28 | 8 | 20 |
|
| ≤45 | 28 | 7 | 21 |
|
| Gender |
| 15 | 41 | 0.708 |
|
Male | 30 | 10 | 20 |
|
|
Female | 26 | 5 | 21 |
|
| Tumor size |
|
|
| 0.528 |
|
>2 | 25 | 3 | 22 |
|
| ≤2 | 31 | 12 | 19 |
|
| Extrathyroidal
extension |
|
|
<0.01a |
|
Negative | 23 | 10 | 13 |
|
Positive | 33 | 5 | 28 |
| Lymph
node metastasis |
|
|
<0.05a |
|
Absent | 26 | 5 | 21 |
|
|
Present | 30 | 10 | 20 |
|
| Distant
metastasis |
|
|
|
<0.05a |
|
Negative | 29 | 8 | 21 |
|
|
Positive | 27 | 7 | 20 |
|
| TNM stage |
|
|
|
<0.01a |
|
I/II | 27 | 6 | 21 |
|
|
III/IV | 29 | 9 | 20 |
|
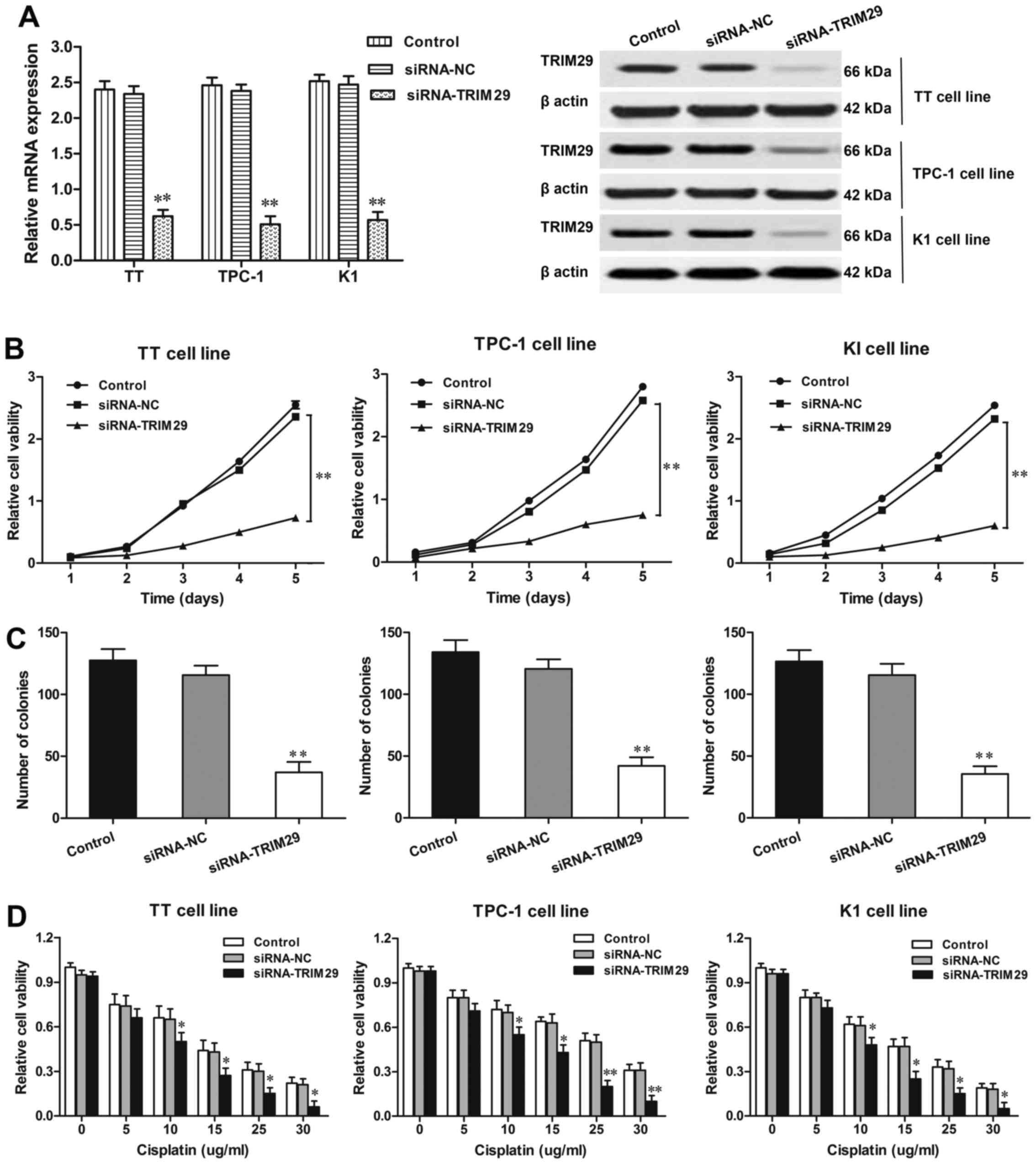
Knockdown of TRIM29 expression in TC
cells inhibits cell proliferation
The expression of TRIM29 was knocked down in TT,
TPC-1, and K1 cells by using siRNA method. The efficiency of TRIM29
knockdown was confirmed by RT-qPCR and western blotting (Fig. 3A). Next, the viability of cells with
TRIM29 knockdown was detected by MTT assay. As shown in Fig. 3B, TRIM29 knockdown significantly
decreased the viability of TT, TPC-1, and K1 cells (P<0.01,
respectively) when compared with NC group. Furthermore, in
agreement with MTT data, colony formation assays also illustrated
that the numbers of colonies were significantly reduced in TT,
TPC-1, and K1 cells with TRIM29 knockdown compared to corresponding
NC groups (P<0.01, respectively; Fig. 3C). These results suggested that
downregulated TRIM29 expression inhibited the proliferation of TC
cells.
TRIM29 knockdown enhanced the
sensitivity of TC cells to chemotherapy
The effect of TRIM29 knockdown on the sensitivity of
TC cells to cisplain was determined by MTT method. We transfected
TT, TPC-1, and K1 cells with siRNA-TRIM29 or siRNA-NC and treated
with cisplatin at various doses (0, 5, 10, 15, 20 µg/ml) for 24 h.
As shown in Fig. 3D, compared with
the control groups, TT, TPC-1, and K1 cells with siRNA-TRIM29
transfection significantly decreased cell viability by enhancing
cell sensitivity to cisplatin under different-dose treatments (10,
15, 20 µg/ml, respectively). Thus, we concluded that the
downregulation of TRIM29 by siRNA-TRIM29 enhanced cell sensitivity
to cisplatin in TC development.
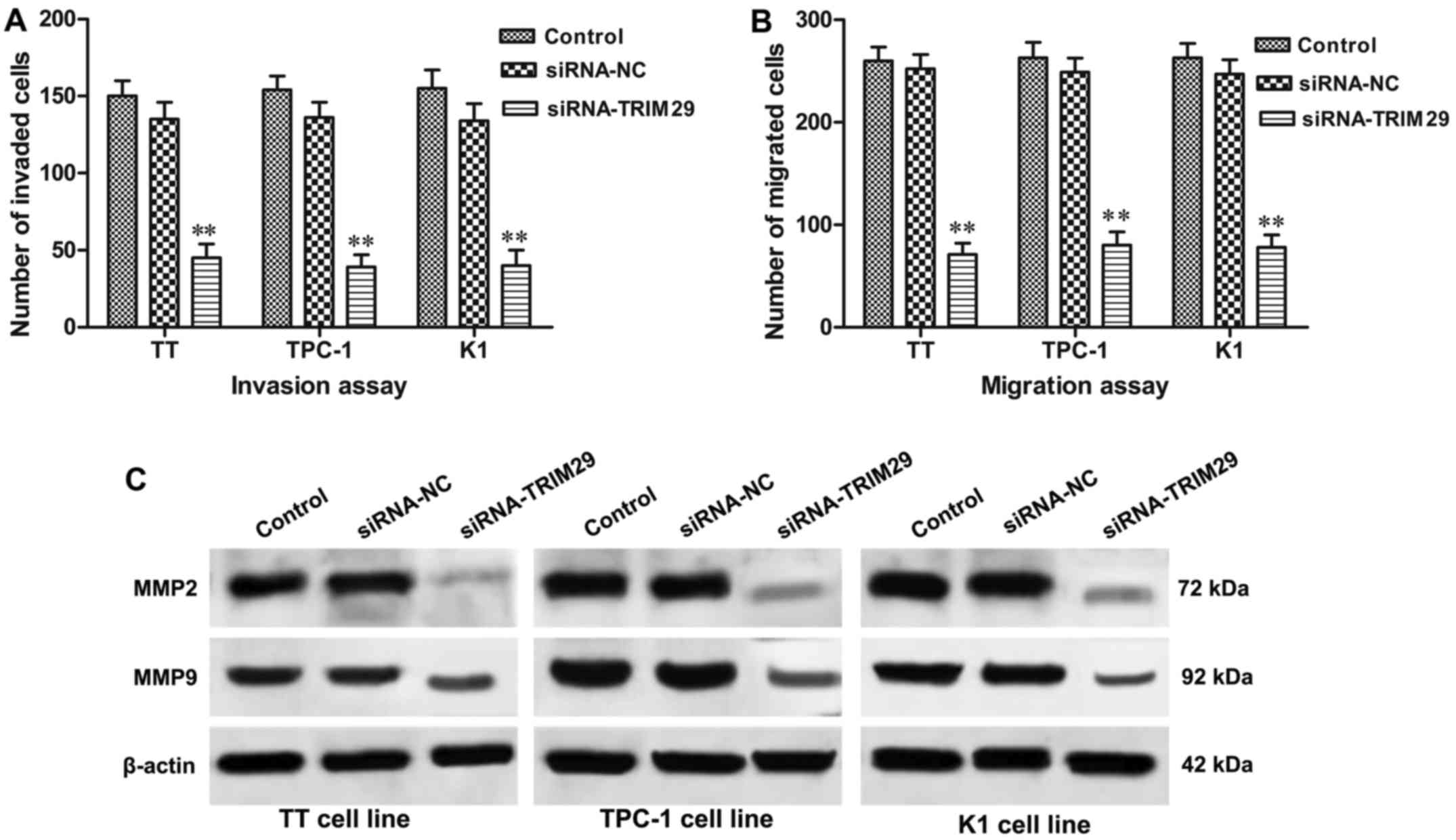
Silencing of TRIM29 expression reduces
invasion and migration of TC cells
To investigate whether the knockdown of TRIM29
influences invasion and migration of TC cells, Transwell invasion
and migration assays were conducted. As shown in Fig. 4A, the numbers of invading cells in
TT, TPC-1, and K1 cells with TRIM29 knockdown were obviously
reduced compared to control groups (P<0.01, respectively).
Consistent with invasion assay, migration assay (Fig. 4B) also revealed that, when compared
with control group, the number of migrated cells were significantly
decreased in TT, TPC-1, and K1 cells with TRIM29 knockdown
(P<0.01, respectively), which indicating that downregulation of
TRIM29 suppressed TC cell invasion and migration. To further
address the role of TRIM29 knockdown in cell invasion and
migration, western blot result revealed that TRIM29 knockdown
significantly decreased the hallmarks of cell invasion and
migration including matrix metallopeptidases (MMP2 and MMP9).
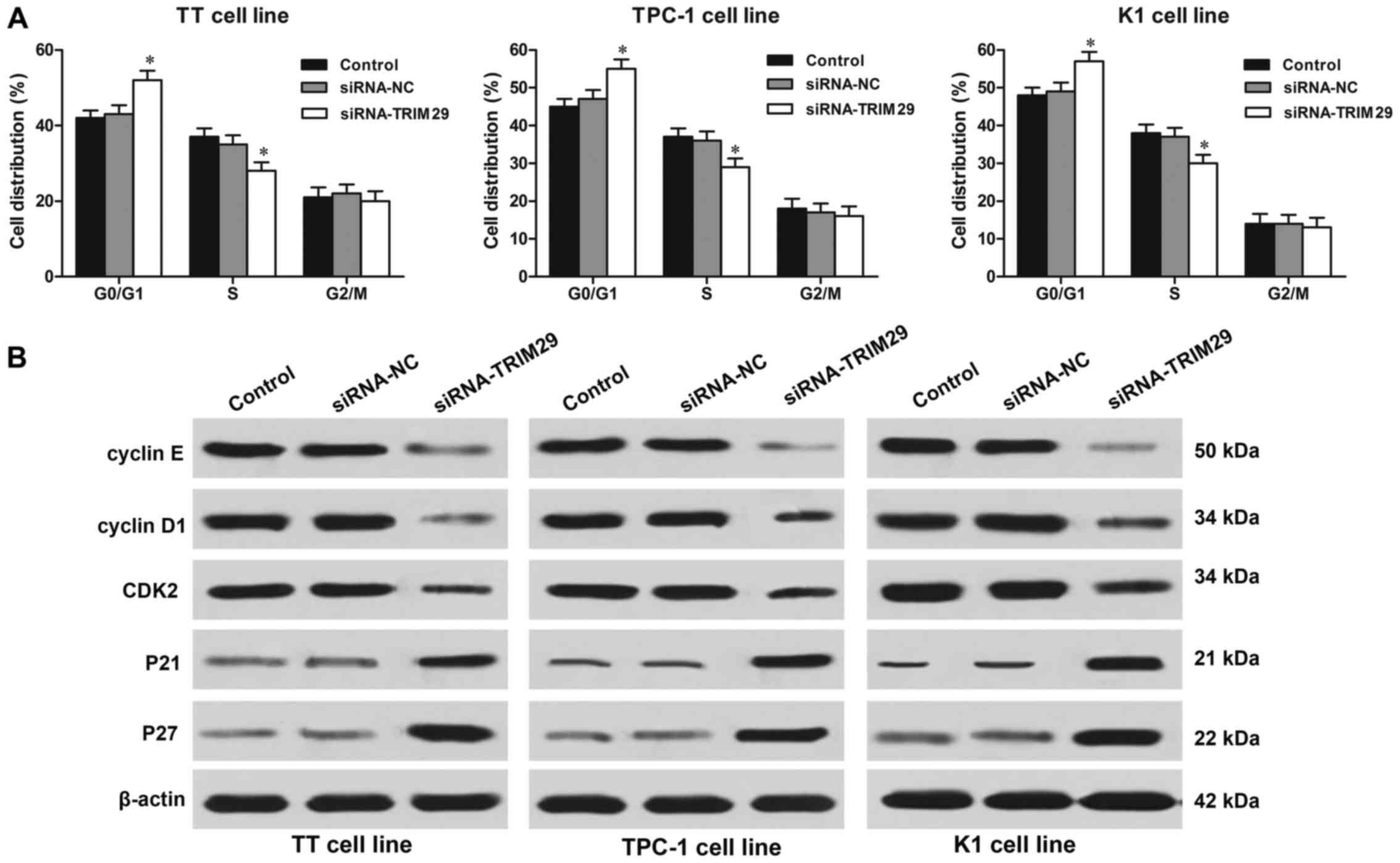
TRIM29 silencing causes cell cycle
arrest at G0/G1 phases in TC cells
The flow cytometry assay showed that knockdown of
TRIM29 contributed to a remarkably increased in the G0/G1 phase in
TT, TPC-1, and K1 cells compared to cells in control group
(P<0.01, respectively). While cells in S phase were reduced in
TRIM29 knockdown cells compared to cells in control group
(P<0.01, respectively; Fig. 5A).
Furthermore, to deeply investigate the mechanism underlying the
cell cycle arrest caused by TRIM29 knockdown in TC cells, the
central cell cycle regulatory proteins such as cyclin D1, cyclin E,
cyclin-dependent kinase 2 (CDK2), p21 and p27 were examined by
western blotting. As shown in Fig.
5B, the expression levels of cyclin D1, cyclin E and CDK2 were
significantly decreased; while the expression of cyclin-dependent
kinase inhibitors such as p21 and p27 were increased in TC cells
with TRIM29 knockdown. Thus, these findings indicated that
knockdown of TRIM29 resulted in cell cycle arrest in TC cells.
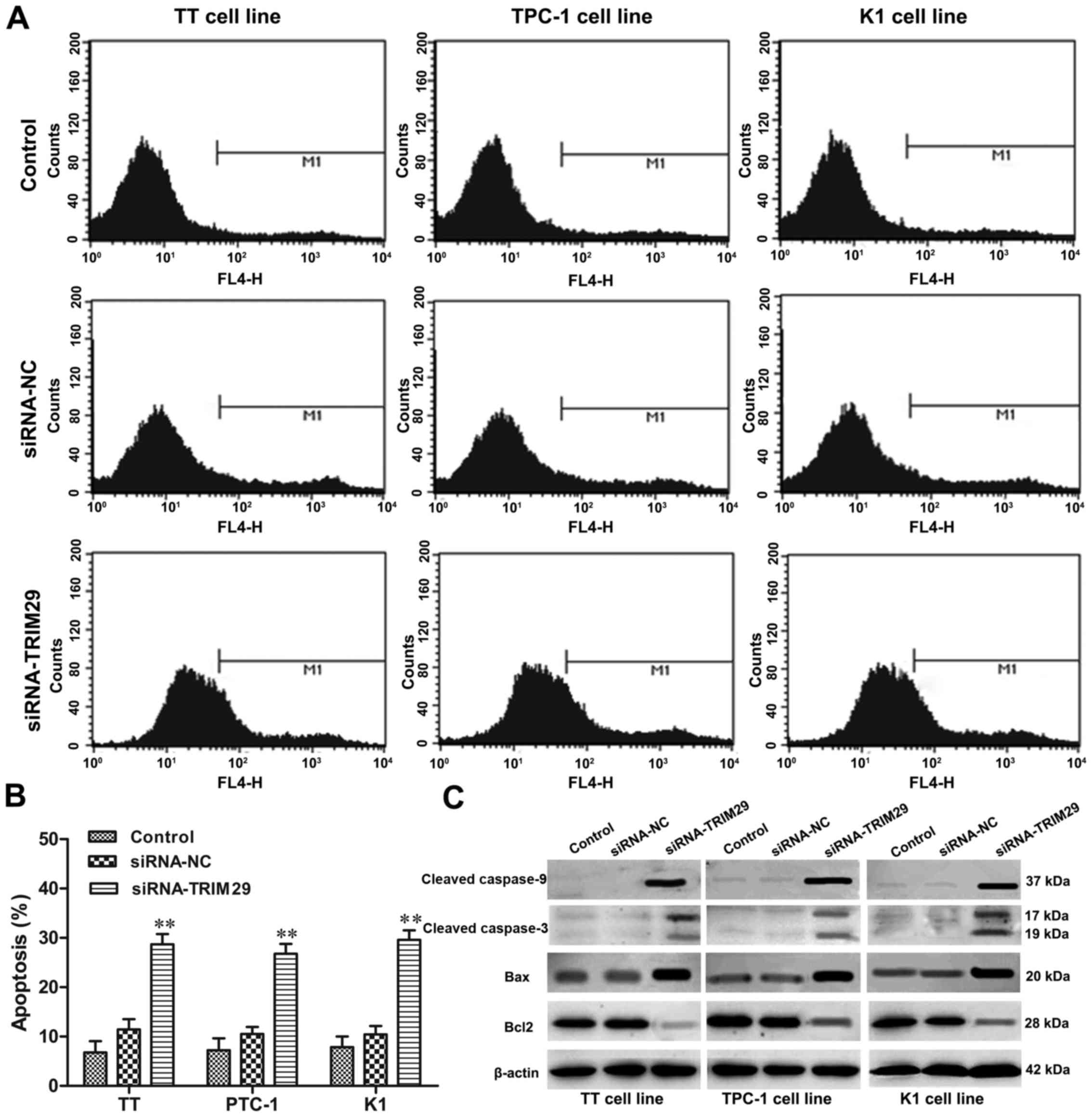
Knockdown of TRIM29 induces apoptosis
in TC cells
Flow cyctometry was used to clarify the effect of
TRIM29 knockdown on cell apoptosis of TC cells. TRIM29 knockdown
significantly increased the percentages of cell apoptosis in TT,
TPC-1, and K1 cells compared to control group (P<0.01,
respectively, Fig. 6A and B). In
addition, we also found that TRIM29 knockdown induced TC cell
apoptosis by activating apoptosis-related protein expression. As
shown in Fig. 6C, upregulation of
TRIM29 remarkably increased the levels of cleaved caspase-3,
cleaved caspase-9, Bax expression, while decreased the expression
of Bcl-2 in TT, TPC-1, and K1 cells compared to control groups.
Thus, these results suggested that inhibition of TRIM29 induced
apoptosis of TC cells.
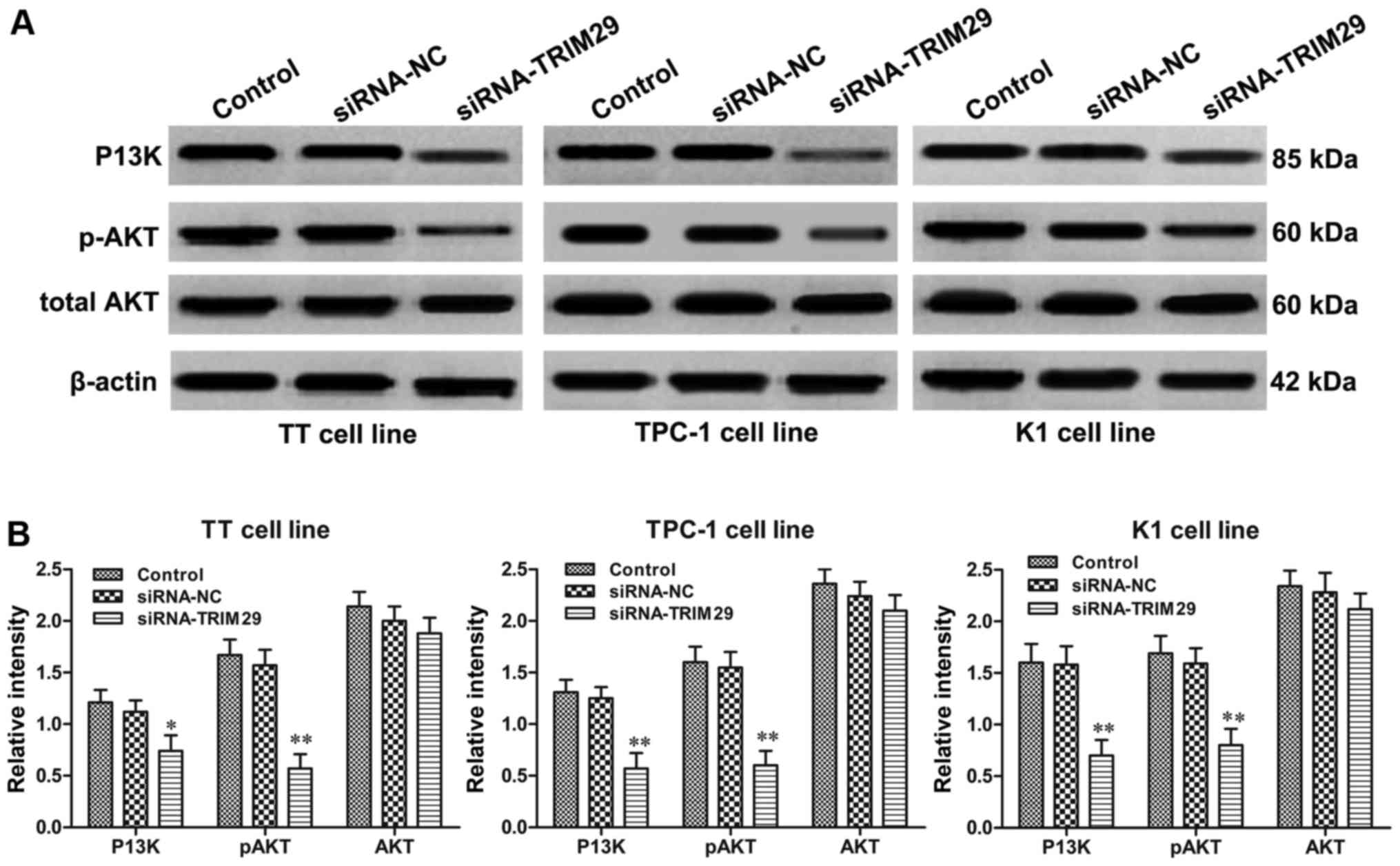
Knockdown of TRIM29 suppresses the
P13K/AKT signaling pathway
The phosphoinositide 3-kinase (P13K)/AKT signaling
pathway plays a crucial role in thyroid oncogenesis and affects
proliferation, metastasis, apoptosis of TC cells (14). Since the inhibitory effect of TRIM29
knockdown on TC cell cycle and apoptosis has been clarified, to
further determine whether P13K/AKT signaling pathway is involved in
TRIM29-mediated TC tumorigenesis, the key molecules of the P13K/AKT
pathway were detected in TT, TPC-1, and K1 cells with TRIM29
knockdown. Western blot results showed that the P13K, and
phosphorylation of AKT expression levels were prominently decreased
in TT, TPC-1, and K1 cells with downregulation of TRIM29 compared
to vehicle-treated cells, though the total AKT levels were similar
among different treated groups (Fig.
7). Our results illustrated that TRIM29 knockdown significantly
inhibited the P13K/AKT signaling pathway in TC cells.
Discussion
TC is the most common endocrine malignancy, with
increasing knowledge of its molecular pathogenesis (15,16).
In order to increase the efficacy of the therapeutic treatment and
to reduce side effects, the investigations on gene dysregulation
during TC progression are becoming the novel subject of research in
this field. The location of TRIM29 is at chromosome 11q23, and
functions as a sensitive component of a protein kinase mediated
signaling transduction pathway against ionizing radiation, which
might aggravate tumorigenesis and progression (17,18).
TRIM29 is involved in tumor proliferation and progression in a
variety of cancer cells. However, among different tumor cell types,
TRIM29 functioned either as tumor suppressor or oncogene according
to the origin of the neoplasm (4).
Recent analysis demonstrated that TRIM29 overexpression could
promote tumor cell survival and growth in various types of cancers
(19). To our knowledge, the
investigation of the relationship between TRIM29 expression and TC
tumorigenesis has not been previously reported.
In the present study, we found that TRIM29 mRNA and
protein were significantly enhanced in TC tumor samples and cell
lines, suggesting a pro-oncogenic role of TRIM29 in TC progression.
Moreover, the high expression of TRIM29 was tightly associated with
TNM staging, extrathyroidal extension, lymph nodes metastasis, and
distant metastasis. Notably, the overall survival rate and
disease-free rate of patients with high TRIM29 expression was much
lower than those with low TRIM29 expression, indicating that TRIM29
might be a potential predictor for the prognosis of TC patients.
These findings were in agreement with other reports on TRIM29 in
many cancers. Kosaka et al reported that TRIM29
overexpression was correlated with the metastatic process and poor
prognosis in gastric cancer patients (9).
A previous study emphasized that TRIM29 functioned
as a histone-binding protein to assemble DNA repair proteins into
chromatin and then activated DNA damage response (20). Wang et al also indicated that
TRIM29 played a crucial role in DNA damage signaling and
radioresistance in human pancreatic cancer cells (21). Xu et al demonstrated that
TRIM29 was a novel prognostic biomarker for prediction in
aggressive cervical cancer patients that need postoperative
adjuvant chemoradiotherapy (22).
Liu et al suggested that silencing of TRIM29 enhanced
chemosensitivity to cisplatin in human lung squamous cancer by
increasing cell apoptosis (10).
Cisplatin is a widely used chemotherapeutic drug against various
types of cancers through DNA damage mechanism (23). The present study stated the
association of TRIM29 expression with cisplatin cell resistance in
TC cells and proved knockdown of TRIM29 made TC cells more
sensitive to cisplatin treatment.
Studies have shown that TRIM29 is intimately
associated with cell proliferation, cell cycle, invasion and cell
apoptosis. Liu et al discovered that TRIM29 knockdown
suppressed proliferation and invasion in human lung squamous cancer
cells (10). Sun et al
reported that TRIM29 promoted tumor cell proliferation and
progression in pancreatic cancer (8). Yuan et al discovered that
TRIM29 promoted cancer cell proliferation by suppressing p53
nulcear activities (24). To
illustrate the function of TRIM29 on TC progression, siRNA approach
was employed to silence TRIM29 expression in TC cell lines. Our
data showed that TRIM29 knockdown significantly inhibited TC cell
proliferation and reduced colony formation, caused cell cycle to
halt at G0/G1 phase, accompanied by repressing the expression of
cyclin D1, cyclin B1, and CDK2; while increasing the expression of
p21 and p27, which resulted in cell growth inhibition. Cyclin D1
could phosphorylate Rb protein and accelerate cancer progression
via G1-S phase of cell cycle (25).
Furthermore, the activation of cyclin E-CDK2 complex has been
proved as another crucial demand for the G1/S phase transition in
cancer aggressiveness; conversely, p21 and p27 inhibit the
activation of CDK-cyclin complexes during tumor development
(26). Collectively, the
alternations of cell cycle regulatory molecules may contribute to
the phase distribution in TC cells.
In the present study, we found that the expression
of TRIM29 was significant in lymph nodes metastasis, and distant
metastasis in TC progression according to clinicopathological
factor analysis. The abilities of invasion and metastasis of cancer
cells are dependent on the degradation of the components of
extracellular matrix and basement membranes, which is regulated by
matrix metalloproteinase (MMP) family (27). Silencing of RTIM29 significantly
reduced the invasive and migratory abilities of TC cells by
inhibiting the activities of MMP-2 and MMP-9, which are implicated
in dysregulation of tumor invasion and metastasis in numerous human
malignancies (28). Furthermore,
the imbalance between apoptosis-induced and -inhibited molecule
expressions are responsible for the vulnerability of cells to
apoptosis (29). Thus, in this
study, the activation of caspase-3 and caspase-9, decreased ratio
of Bax/Bcl-2 indicated that TRIM29 knockdown induced TC cell
apoptosis in a caspase- and Bax/Bcl-2-dependent manner.
P13K/AKT pathway is a vital intracellular signaling
pathway for regulating various bioprocesses in mammalian cells,
including cell proliferation, metabolism, transformation, motility
and tumorigenesis (30). P13K/AKT
pathway could induce the development of tumor and promote tumor
metastasis via various pathways. In order to enhance cell survival,
activation of P13K/AKT pathway could in turn activate or inhibit a
series of downstream target proteins, such as caspase-9, p21, Bax,
MMP2, and mammalian target of rapamycin (mTOR) (14,31,32).
Tan et al proved that TRIM29 overexpression promoted cell
proliferation and survival through NF-κB signaling pathway in
bladder cancer cells (19). Xu
et al reported that TRIM29 overexpression enhanced tumor
progression by activating Wnt/β-actenin pathway in cervical cancer
(22). Furthermore, a recent study
demonstrated that elevated TRIM29 expression contributed to the
cell proliferation and metastasis of nasopharyngeal cancer by
activating PTEN/AKT/mTOR signaling pathway (33). On the basis of references above, in
our study, we found that the antitumor effect of TRIM29 silencing
was associated with the inhibition of P13K/AKT signaling pathway,
which might be the pivotal mechanism of TRIM29 knockdown-mediated
proliferation suppression, chemosensitivity enhancement, the
inhibition of invasion and migration, delayed cell cycle
transition, cell apoptosis in TC (34). However, the deep molecular details
still remained to be further elucidated.
In conclusion, this study highlighted the crucial
role of TRIM29 overexpression in TC prognosis and progression, and
subsequently demonstrated that TRIM29 knockdown resulted in the
alternations of cell proliferation, chemosensitivity to cisplatin,
invasion and migration, cell cycle arrest, and cell apoptosis via
blocking the activation of P13K/AKT signaling pathway and the
involved downstream target genes, which provide credible evidence
for novel therapeutic target combined with the P13K/AKT signaling
pathway for TC treatment.
Acknowledgements
We would like to thank Professor J.C. Ma for his
intelligent suggestions.
References
|
1
|
Zhang X, Wang Z, Tian L, Xie J, Zou G and
Jiang F: Increased expression of FGF19 contributes to tumor
progression and cell motility of human thyroid cancer. Otolaryngol
Head Neck Surg. 154:52–58. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kong LL, Man DM, Wang T, Zhang GA and Cui
W: siRNA targeting RBP2 inhibits expression, proliferation,
tumorigenicity and invasion in thyroid carcinoma cells. Oncol Lett.
10:3393–3398. 2015.PubMed/NCBI
|
|
3
|
Ye WC, Gao L, Huang J, Fang XM and Xie G:
Suppressed Krüppel-like factor 17 expression induces tumor
proliferation, metastasis and a poor prognosis in papillary thyroid
carcinoma. Mol Med Rep. 10:2087–2092. 2014.PubMed/NCBI
|
|
4
|
Hatakeyama S: TRIM proteins and cancer.
Nat Rev Cancer. 11:792–804. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Napolitano LM and Meroni G: TRIM family:
Pleiotropy and diversification through homomultimer and
heteromultimer formation. IUBMB Life. 64:64–71. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cambiaghi V, Giuliani V, Lombardi S,
Marinelli C, Toffalorio F and Pelicci PG: TRIM proteins in cancer.
Adv Exp Med Biol. 770:77–91. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Song X, Fu C, Yang X, Sun D, Zhang X and
Zhang J: Tripartite motif-containing 29 as a novel biomarker in
non-small cell lung cancer. Oncol Lett. 10:2283–2288.
2015.PubMed/NCBI
|
|
8
|
Sun H, Dai X and Han B: TRIM29 as a novel
biomarker in pancreatic adenocarcinoma. Dis Markers.
2014:3178172014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kosaka Y, Inoue H, Ohmachi T, Yokoe T,
Matsumoto T, Mimori K, Tanaka F, Watanabe M and Mori M: Tripartite
motif-containing 29 (TRIM29) is a novel marker for lymph node
metastasis in gastric cancer. Ann Surg Oncol. 14:2543–2549. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu C, Huang X, Hou S, Hu B and Li H:
Silencing of tripartite motif (TRIM) 29 inhibits proliferation and
invasion and increases chemosensitivity to cisplatin in human lung
squamous cancer NCI-H520 cells. Thorac Cancer. 6:31–37. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang T, Tang HM, Lu S, Yan DW, Yang YX
and Peng ZH: Up-regulation of tripartite motif-containing 29
promotes cancer cell proliferation and predicts poor survival in
colorectal cancer. Med Oncol. 30:7152013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ai L, Kim WJ, Alpay M, Tang M, Pardo CE,
Hatakeyama S, May WS, Kladde MP, Heldermon CD, Siegel EM, et al:
TRIM29 suppresses TWIST1 and invasive breast cancer behavior.
Cancer Res. 74:4875–4887. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kanno Y, Watanabe M, Kimura T, Nonomura K,
Tanaka S and Hatakeyama S: TRIM29 as a novel prostate basal cell
marker for diagnosis of prostate cancer. Acta Histochem.
116:708–712. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin S, Borkhuu O, Bao W and Yang YT:
Signaling pathways in thyroid cancer and their therapeutic
implications. J Clin Med Res. 8:284–296. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang X, Liu G, Xiao H, Yu F, Xiang X, Lu
Y, Li W, Liu X, Li S and Shi Y: TPX2 overexpression in medullary
thyroid carcinoma mediates TT cell proliferation. Pathol Oncol Res.
20:641–648. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Z, Yang L, Teng X, Zhang H and Guan H:
The involvement of CXCR7 in modulating the progression of papillary
thyroid carcinoma. J Surg Res. 191:379–388. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reymond A, Meroni G, Fantozzi A, Merla G,
Cairo S, Luzi L, Riganelli D, Zanaria E, Messali S, Cainarca S, et
al: The tripartite motif family identifies cell compartments. EMBO
J. 20:2140–2151. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kapp LN, Painter RB, Yu LC, van Loon N, CW
III Richard, James MR, Cox DR and Murnane JP: Cloning of a
candidate gene for ataxia-telangiectasia group D. Am J Hum Genet.
51:45–54. 1992.PubMed/NCBI
|
|
19
|
Tan ST, Liu SY and Wu B: TRIM29
Overexpression promotes proliferation and survival of bladder
cancer cells through NF-κB signaling. Cancer Res Treat.
48:1302–1312. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Masuda Y, Takahashi H, Sato S,
Tomomori-Sato C, Saraf A, Washburn MP, Florens L, Conaway RC,
Conaway JW and Hatakeyama S: TRIM29 regulates the assembly of DNA
repair proteins into damaged chromatin. Nat Commun. 6:72992015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang L, Yang H, Palmbos PL, Ney G, Detzler
TA, Coleman D, Leflein J, Davis M, Zhang M, Tang W, et al:
ATDC/TRIM29 phosphorylation by ATM/MAPKAP kinase 2 mediates
radioresistance in pancreatic cancer cells. Cancer Res.
74:1778–1788. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu R, Hu J, Zhang T, Jiang C and Wang HY:
TRIM29 overexpression is associated with poor prognosis and
promotes tumor progression by activating Wnt/β-catenin pathway in
cervical cancer. Oncotarget. 7:28579–28591. 2016.PubMed/NCBI
|
|
23
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan Z, Villagra A, Peng L, Coppola D,
Glozak M, Sotomayor EM, Chen J, Lane WS and Seto E: The ATDC
(TRIM29) protein binds p53 and antagonizes p53-mediated functions.
Mol Cell Biol. 30:3004–3015. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vermeulen K, Van Bockstaele DR and
Berneman ZN: The cell cycle: A review of regulation, deregulation
and therapeutic targets in cancer. Cell Prolif. 36:131–149. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Malumbres M: Cyclins and related kinases
in cancer cells. J BUON. 12 Suppl 1:S45–S52. 2007.PubMed/NCBI
|
|
27
|
Deryugina EI and Quigley JP: Tumor
angiogenesis: MMP-mediated induction of intravasation- and
metastasis-sustaining neovasculature. Matrix Biol. 44–46, 94–112.
2015.
|
|
28
|
Yoon SO, Park SJ, Yun CH and Chung AS:
Roles of matrix metalloproteinases in tumor metastasis and
angiogenesis. J Biochem Mol Biol. 36:128–137. 2003.PubMed/NCBI
|
|
29
|
Abarikwu SO and Farombi EO: Atrazine
induces apoptosis of SH-SY5Y human neuroblastoma cells via the
regulation of Bax/Bcl-2 ratio and caspase-3-dependent pathway.
Pestic Biochem Physiol. 118:90–98. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hsieh AC, Truitt ML and Ruggero D:
Oncogenic AKTivation of translation as a therapeutic target. Br J
Cancer. 105:329–336. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ávalos Y, Canales J, Bravo-Sagua R,
Criollo A, Lavandero S and Quest AF: Tumor suppression and
promotion by autophagy. BioMed Res Int. 2014:6039802014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xing M: Genetic alterations in the
phosphatidylinositol-3 kinase/Akt pathway in thyroid cancer.
Thyroid. 20:697–706. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou XM, Sun R, Luo DH, Sun J, Zhang MY,
Wang MH, Yang Y, Wang HY and Mai SJ: Upregulated TRIM29 promotes
proliferation and metastasis of nasopharyngeal carcinoma via
PTEN/AKT/mTOR signal pathway. Oncotarget. 7:13634–13650.
2016.PubMed/NCBI
|
|
34
|
Brzezianska E and Pastuszak-Lewandoska D:
A minireview: The role of MAPK/ERK and PI3K/Akt pathways in thyroid
follicular cell-derived neoplasm. Front Biosci (Landmark Ed).
16:422–439. 2011. View
Article : Google Scholar : PubMed/NCBI
|