Introduction
Nasopharyngeal carcinoma (NPC) is a common tumor
which originates from epithelial cells located in the nasopharynx
(1). NPC has a complex etiology and
specific world distribution, with most cases occuring in Southern
China and Southeast Asia (2).
Currently, radiotherapy is the standard therapeutic strategy for
NPC (3). NPC in its early stage is
highly radiosensitive. However, NPC usually shows radioresistance
in the more advanced stage due to local recurrence and distant
metastasis (4). Therefore,
investigation of the molecular mechanisms that enhance
radiosensitivity of NPC may be conducive to exploring novel
therapeutic strategies thereby consequently improving clinical
outcomes.
Transforming growth factor-β (TGF-β) is a
pluripotent cytokine that demonstrates distinct roles during
tumorigenesis. In normal epithelial cells and early-stage cancer,
TGF-β is a strong inhibitor of cell growth. However, in late-stage
cancer, TGF-β increases tumor invasiveness and metastasis, thus
contributing to tumor progression (5,6).
Recently, a growing number of studies suggest that TGF-β signaling
enhances the invasive and metastatic potential of various cancers
by inducing a cellular process called epithelial-to-mesenchymal
transition (EMT) (7). EMT is a
reversible biologic program changing polarized epithelial cells
into motile fibroblastoid cells. EMT is associated with increased
invasion, migration, and cell proliferation of epithelial cancers,
including NPC (8).
Berberine (2,3-methylenedioxy-9,
10-dimenthoxyprotoberberine chloride, BBR) (Fig. 1A) is a natural isoquinoline alkaloid
derived from Berberis species. Berberine exhibits a variety
of pharmacological effects with antibacterial, antifungal and
anti-inflammatory activities (9).
Recently, studies have shown that berberine exhibits anticancer
activity in various cancers including NPC. Berberine can inhibit
tumor metastasis, tumorigenicity and growth, and enhance
radiosensitivity of NPC via regulation of multiple pathways
(10–12). However, the molecular mechanisms
underlying the radiosensitizing effects of berberine in NPC remain
largely unclear.
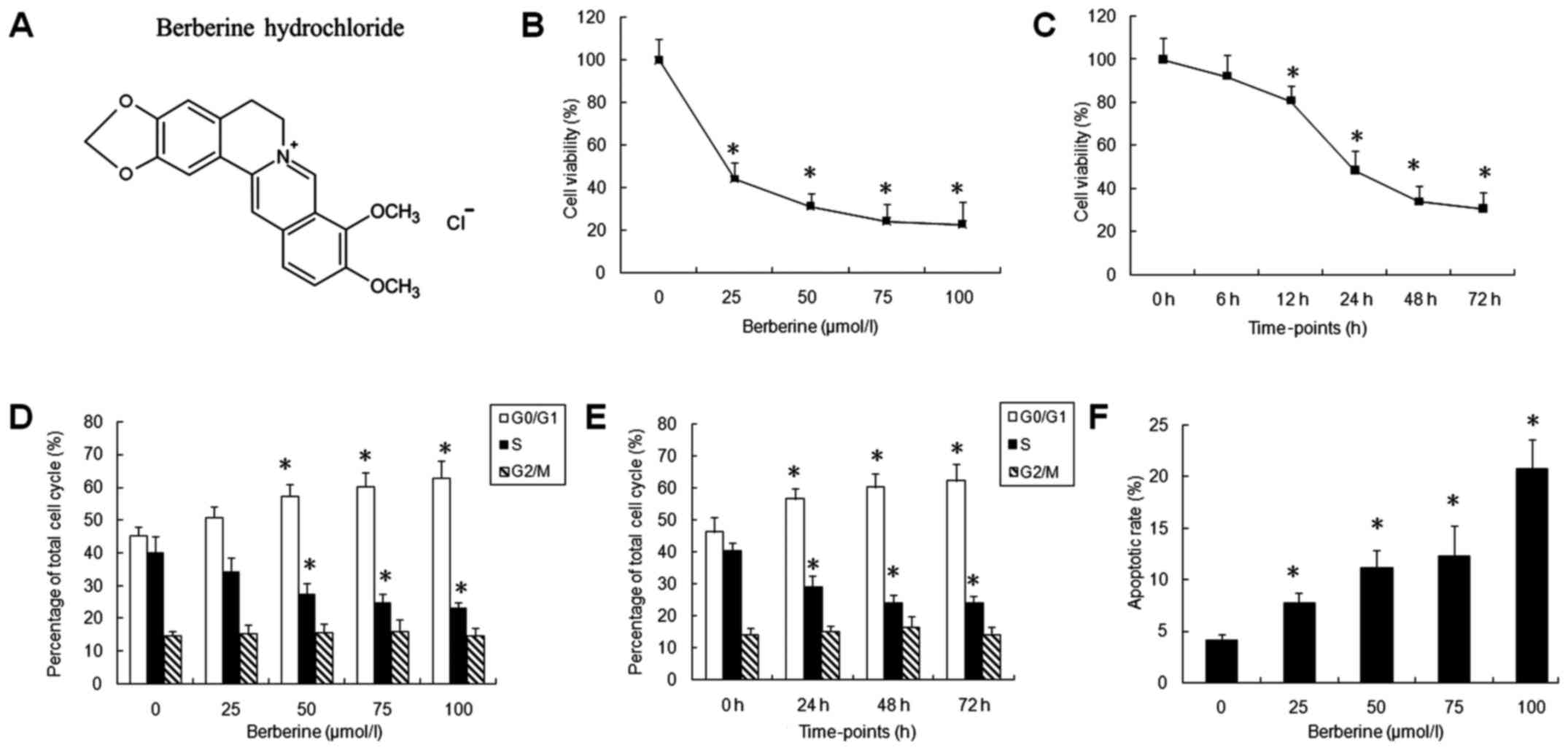 | Figure 1.Berberine inhibits cell proliferation,
induces cell cycle arrest and apoptosis of nasopharyngeal carcinoma
(NPC) cells. (A) Chemical structure of berberine hydrochloride. (B)
Berberine inhibits the viability of NPC cells in a
concentration-dependent manner. CNE-2 cells were treated with
berberine at the indicated concentrations (0, 25, 50, 75 and 100
µmol/l) for 72 h, and the cell viability was then determined by MTT
assay. (C) Berberine inhibits the viability of NPC cells in a
time-dependent manner. CNE-2 cells were treated with 100 µmol/l
berberine for 6, 12, 24, 48 or 72 h. All data are expressed as the
mean ± SD values from three independent experiments and were
compared using a two-tailed, unpaired t-test. Cell viability was
normalized to that of the untreated control cells. (D) Berberine
induced cell cycle arrest at the G0/G1 phase in NPC cells in a
concentration-dependent manner. CNE-2 cells were treated with
berberine at various concentrations (0, 25, 50, 75 and 100 µmol/l)
for 72 h, and then were stained with propidium iodide (PI; 20
µg/ml) to determine the percentages of cells in the G0/G1, S and
G2/M phases. (E) Berberine induced cell cycle arrest in the G0/G1
phase of NPC cells in a time-dependent manner. CNE-2 cells were
treated with 100 µmol/l berberine for 24, 48 or 72 h, followed by
PI staining. (F) Berberine induced cell apoptosis in NPC cells.
CNE-2 cells were treated with various concentrations of berberine
(0, 25, 50, 75 and 100 µmol/l) for 24 h. Significant difference
from the control group (berberine concentration is 0; or berberine
treatment time is 0) is denoted by *P<0.05. |
Our laboratory previously found that specificity
protein 1 (Sp1), a transcription factor, is overexpressed in NPC
tissues compared with their adjacent normal tissues. High
expression of Sp1 was found to be correlated with tumor invasion,
distant metastasis and radioresistance in NPC patients (13). This indicates that Sp1 protein may
mediate the signaling pathway of NPC radioresistance. However,
currently there is no study concerning the regulation of Sp1 in the
radiosensitizing effects of NPC.
In this study, we investigated the effects of
berberine on the proliferation, cell cycle and apoptosis of CNE-2
NPC cells, especially the effect on TGF-β-induced EMT which
promotes NPC cell invasion, migration and metastasis. Our study
also explored the roles of Sp1 in inhibition of TGF-β-induced EMT
by berberine in NPC cells.
Materials and methods
Cell culture
The undifferentiated human NPC cell line CNE-2 was
routinely maintained in RPMI-1640 medium (Invitrogen-Gibco,
Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS)
(Sijichun Bioengineering Materials Inc., Hangzhou, Zhejiang,
China), 2 mM sodium pyruvate, 2 mM L-glutamine (Invitrogen-Gibco),
100 U/ml penicillin and 100 U/ml streptomycin. Cells were cultured
in a humid atmosphere of 5% CO2 at 37°C. CNE-2 cells
were digested by trypsinization after reaching confluence and were
subcultured in new culture dishes at lower densities.
Irradiation condition
The NPC cells at the logarithmic growth phase were
randomly divided into 96-well culture plates and incubated with
berberine (BBR) (0, 25, 50, 75 and 100 µmol/l) or mithramycin A
(Mith) (100 nmol/l) for 24 h. Then the cells were exposed to (60)Co
gamma rays with various doses of irradiation (0, 4, 6 and 8 Gy)
using a linear accelerator (Elekta, Stockholm, Sweden). The
source-skin distance (SSD) technique was 80 cm and the depth was
set at 0.8 cm to the bottom of the culture plates. The irradiation
was performed for 72 h followed by MTT assay.
Cell proliferation assay
CNE-2 cells in the logarithmic growth phase were
trypsinized and seeded in a 96-well plate at a density of
1×104 cells/well. After 24 h, old medium was removed and
incubated with fresh RPMI-1640 medium supplemented with 10% FBS and
containing various concentrations of berberine (0, 25, 50, 75 and
100 µmol/l) for 6, 12, 24, 48 or 72 h. Cell viability was measured
using the MTT (Sigma Chemical Co., St. Louis, MO, USA) assay
following the manufacturer's instructions. To assess cell
viability, 10 µl of MTT solution (5 mg/ml) was added into each well
to incubate at 37°C for 4 h. Centrifugation at 1,000 × g was
performed for 10 min to remove the supernatant, and the obtained
formazan pellet was dissolved with 100 µl DMSO in each well. The
absorbance at 570 nm wavelength was determined using a ELISA plate
reader (Ricso RK201; Shenzhen Ricso Technology Co., Ltd., Shenzhen,
China) to evaluate the amount of pellet. The surviving cells from
each group were normalized to those of the controls and were
expressed as percentage of cell viability. All experiments were
repeated at least three times.
Cell cycle analysis
CNE-2 cells at the logarithmic growth phase were
randomly seeded in 60-mm culture dishes, and were incubated with
serum-free medium after reaching 50% confluence to induce cell
quiescence. After 24 h, cells were incubated with different
concentrations of berberine (0, 25, 50, 75 and 100 µmol/l) for 24,
48 or 72 h in complete medium. Cells were harvested by
trypsinization and centrifugation at 1,000 × g for 5 min, and then
cells were mixed with cold 70% ethanol for resuspension. Then the
suspension was incubated with 1 ml propidium iodide (PI) solution
(20 µg/ml PI; 100 µg/ml RNase A) for 30 min, and were analyzed on a
flow cytometry (FACScan; Becton-Dickinson, San Francisco, CA, USA).
Data were acquired from 10,000 cells and analyzed by Lysis II
software (Becton-Dickinson). The cell number in the G0/G1, S and
G2/M phases were calculated and the respective percentages were
calculated.
Cell apoptosis assay
CNE-2 cells were randomly seeded in 60-mm culture
dishes and incubated with various concentrations of berberine (0,
25, 50, 75 and 100 µmol/l) for 24 h. Then cells were harvested by
trypsinization from each group and underwent centrifugation at
1,000 × g for 5 min. The cell were incubated with 100 µl 1X binding
buffer containing 5 µl Annexin V and 5 µl PI (final concentration
of 10 µg/ml). After a 15-min incubation in the dark, apoptosis of
samples was determined by flow cytometry (FACScan;
Becton-Dickinson), and data were analyzed using CellQuest software
(Becton-Dickinson, Franklin Lakes, NJ, USA). At least 10,000 events
were analyzed for each sample.
Quantitative real-time polymerase
chain reaction (qRT-PCR)
CNE-2 cells were incubated with various
concentrations of berberine (0, 100 and 300 µmol/l) for 12, 24 or
48 h, and total RNA was extracted using TRIzol® reagent
(Life Technologies, Carlsbad, CA, USA) according to the
manufacturer's instructions. After treatment with DNase,
complementary DNA (cDNA) was synthesized from total RNA (2 µg) by
reverse transcription using the SuperScript III enzyme (both from
Life Technologies). The mRNA level of Sp1 was determined by qRT-PCR
based on cDNA as a template using SYBR-Green reagent (Takara,
Tokyo, Japan) in the StepOne Plus Real-Time PCR system (Applied
Biosystems, Foster City, CA, USA). The PCR conditions were as
follows: initial denaturation at 95°C for 10 min, followed by 40
amplification cycles of denaturation at 95°C for 15 sec and
annealing at 60°C for 60 sec. Housekeeping gene
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as an
internal control. Sp1 mRNA expression was calculated by the
2−ΔΔCt method. The primer sequences used in our study
were as follows: Sp1 forward, 5′-AGTTCCAGACCGTTGATGGG-3′ and
reverse, 5′-GTTTGCACCTGGTATGATCTGT-3′; GAPDH forward,
5′-GGAGTCCACTGGCGTCTTC-3′ and reverse,
5′-GCTGATGATCTTGAGGCTGTTG-3′. Relative expression of Sp1 mRNA was
normalized to GAPDH expression. All reactions were performed in
triplicate.
Western blotting
CNE-2 cells were incubated with different
concentrations of berberine (0, 25, 50, 75 and 100 µmol/l) for 24
h, and then the cells were harvested and washed with ice-cold PBS
three times. Cell lysates were lysed with RIPA buffer (150 mM NaCl,
1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS 50 mM Tris-Cl, pH 7.4)
containing 20% (v/v) protease inhibitor cocktail (Sigma-Aldrich).
Then cell lysates were sonicated and incubated on ice for 30 min,
followed by centrifugation at 12,000 × g for 30 min at 4°C for
removal of insoluble debris. Protein concentrations were determined
by bicinchoninic acid (BCA) protein concentration assay kit
(Beijing Biosea Biotechnology Co., Ltd., Beijing, China). Proteins
(50 µg) were separated on sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gels (polyacrylamide concentration 120
g/l), and then electrophoretically transferred to PVDF membranes
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The membranes were
blocked with 3% BSA in TBS-T buffer (3 g/l Tris base, 8 g/l NaCl,
0.2 g/l KCl, 0.1% Tween-20, pH 7.4) at 4°C. After that, the
membranes were incubated with specific mouse monoclonal antibodies
against human Sp1, E-cadherin or vimentin (Santa Cruz Biotechnology
Inc., Santa Cruz, CA, USA) (all 1:1,000 dilutions), and then
incubated with the horseradish peroxidase-conjugated rabbit
anti-mouse secondary antibodies (IgG) (1:1,000 dilution) in TBS-T
plus 3% BSA at room temperature for 1 h. The density of the
targeted bands in the membranes was visualized using enhanced
chemical luminescence (ECL; Pierce® ECL Plus Western
Blotting Substrate; Pierce Biotechnology, Inc., Rockford, IL, USA),
and the protein bands were detected by Bio-Rad ChemiDoc XRS
(Bio-Rad Laboratories, Inc.).
Cell invasion assay
The in vitro invasion capability of the NCE-2
cells was measure by the Boyden chamber invasion assay. In this
experiment, 24-well tissue culture plates were used with a
Transwell filter membrane. The lower side of the filters was coated
with type I collagen (0.5 mg/ml). Cells (5×104) were
seeded on the upper side of the filter membrane in 100 µl DMEM.
Cells were incubated with TGF-β1 (5 ng/ml), TGF-β1+BBR (50 µmol/l),
or TGF-β1+Mith (100 nmol/l). Cells treated with DMEM served as the
control. After 48 h, cells on the upper surface of the filter were
wiped off, and migrated cells on the lower part of filter membrane
were stained with crystal violet (Sigma-Aldrich), and counted using
light microscopy (x100 magnification) as the numbers of migrated
cells. The migrated cells of each group were normalized to those of
the untreated controls. Each sample was performed in three
independent experiments.
Statistical analysis
All quantitative data are expressed as the mean ±
standard deviation (SD) and were acquired from experiments that
were repeated more than three times. Statistical analysis was
performed using commercially available software (SPSS version 19.0)
(SPSS, Inc., Chicago, IL, USA). A two-tailed unpaired Student's
t-test or one-way analysis of variance (ANOVA) was performed to
measure the differences between the means of the different groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Berberine inhibits the proliferation
and induces cell cycle arrest and apoptosis in NPC cells
To evaluate the effects of berberine on the cell
proliferation of NPC cells, an MTT assay was performed in CNE-2
cells treated with different concentrations of berberine (25, 50,
75 and 100 µmol/l) for 72 h, or in CNE-2 cells treated with 50
µmol/l berberine for 6, 12, 24, 48 or 72 h. Increasing
concentrations of berberine and prolonged time from 12 to 72 h
resulted in significantly reduced cell viability in the CNE-2 cells
(Fig. 1B and C). This indicates
that berberine inhibited NPC cell proliferation in a concentration-
and time-dependent manner.
To investigate whether the berberine-induced
decrease in CNE-2 cell viability was associated with cell cycle
regulation, we analyzed the cell cycle of CNE-2 cells by PI
staining in flow cytometry. CNE-2 cells were treated with various
concentrations of berberine (0, 25, 50, 75 and 100 µmol/l) for 72
h, or treated with 50 µmol/l berberine for 24, 48 or 72 h.
Berberine significantly increased the percentage of cells in the
G0/G1 phase at concentrations of 50, 75 and 100 µmol/l for 72 h,
and at 50 µmol/l for 24, 48 and 72 h (P<0.05) (Fig. 1D and E). However, the percentage of
cells in the G2/M phase remained unchanged after berberine
treatment.
We further measured cell apoptosis in CNE-2 cells to
explore whether reduced cell viability was caused by cell death.
CNE-2 cells were treated with various concentrations of berberine
(0, 25, 50, 75 and 100 µmol/l) for 24 h. Berberine induced cell
apoptosis of the CNE-2 cells in a concentration-dependent manner
(P<0.05) (Fig. 1F).
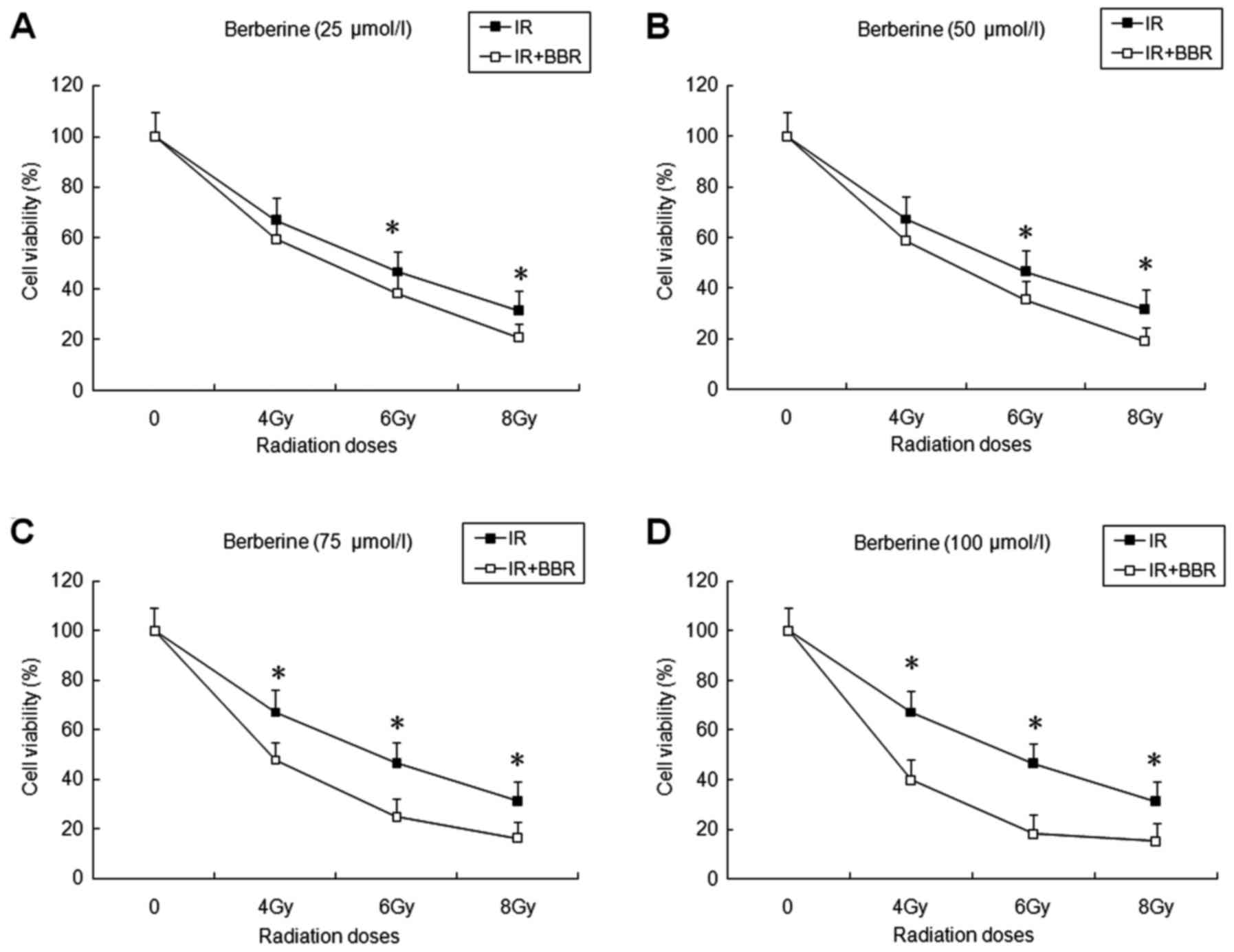
Berberine sensitizes NPC cells to
radiation
To evaluate the effects of berberine on the
radiosensitivity of NPC cells, an MTT assay was performed. CNE-2
cells were treated with irradiation (4, 6 and 8 Gy) alone or
irradiation with berberine (25, 50, 75 and 100 µmol/l) pretreatment
for 3 h. After irradiation for 72 h, cell viability was measured.
Berberine significantly reduced the proliferation and cell
viability of the NPC cells following various doses of irradiation
(P<0.05) (Fig. 2A-D). This
indicates that berberine markedly enhanced the radiosensitivity of
CNE-2 cells.
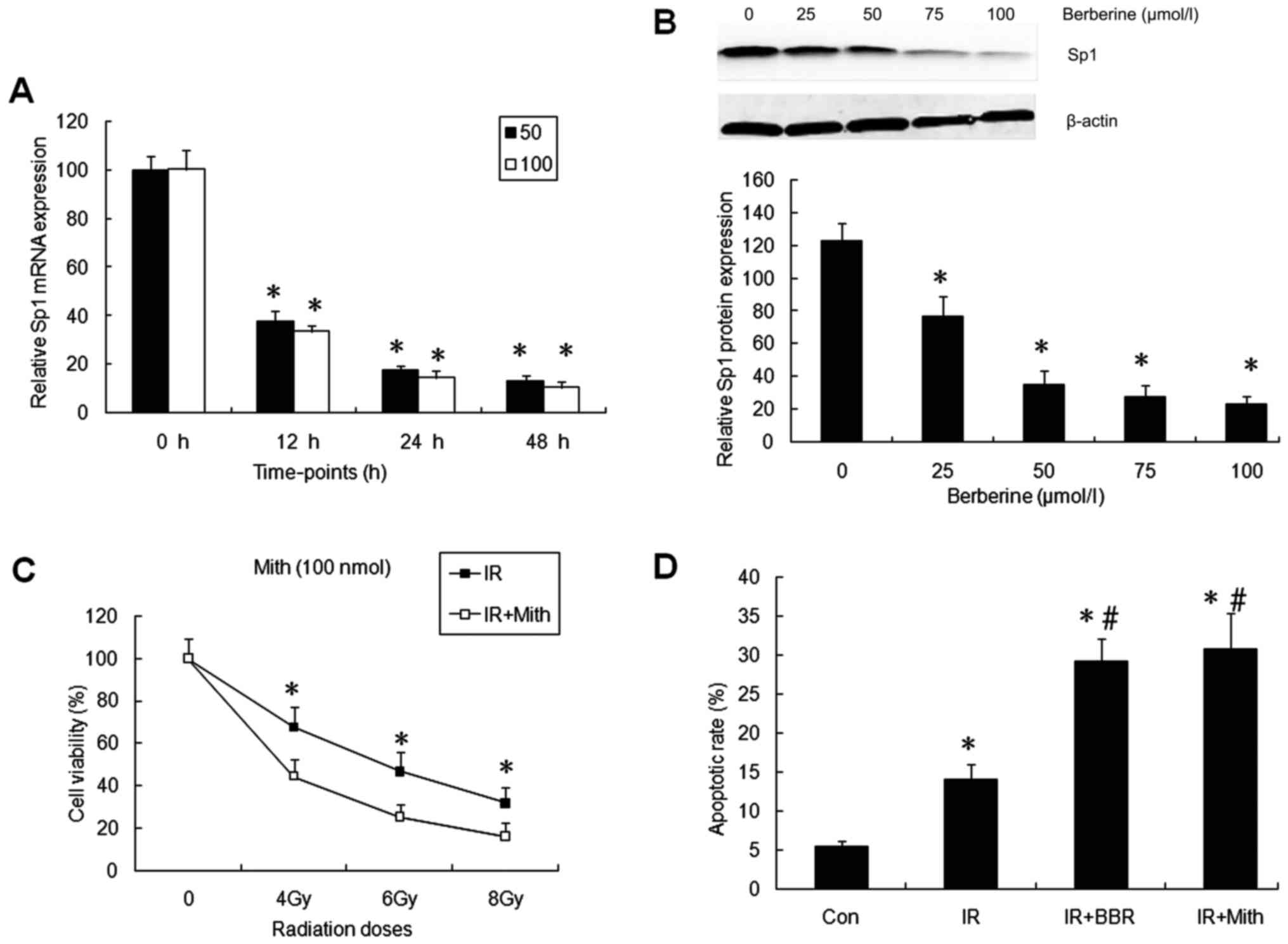
Sp1 is a direct target of berberine
and is involved in NPC-cell radioresistance
To investigate the molecular mechanism underlying
enhanced radiosensitivity of NPC cells by berberine, we performed
qRT-PCR and western blotting to measure the Sp1 mRNA and protein
expression in CNE-2 cells following incubation with berberine.
qRT-PCR showed that berberine decreased Sp1 mRNA in a
time-dependent manner (P<0.05) (Fig.
3A). However, treatment with berberine at two concentrations
(50 and 100 µmol/l) showed similar Sp1 mRNA levels in the NPC cells
at a 12-, 24- and 48-h incubation. Western blotting showed that
berberine decreased Sp1 protein in a concentration-dependent manner
after a 24-h incubation (P<0.05) (Fig. 3B).
To investigate whether decreased Sp1 expression
participates in the enhanced radiosensitivity of CNE-2 cells by
berberine, we analyzed the effect of mithramycin A, a Sp1 specific
inhibitor, on the radiosensitivity and apoptosis of NPC cells.
CNE-2 cells were treated with irradiation (4, 6 and 8 Gy) alone or
irradiation with a 3-h pretreatment of mithramycin A (100 nmol/l).
Mithramycin A significantly reduced the cell viability of the NPC
cells following various doses of irradiation (P<0.05) (Fig. 3C). Furthermore, mithramycin A
increased the apoptotic rate in the CNE-2 cells treated with
irradiation at 4 Gy (P<0.05) (Fig.
3D). This indicates that Sp1 is a molecule essential for the
radioresistance of NPC cells and decreased Sp1 expression may be
involved in the radiosensitivity induced by berberine.
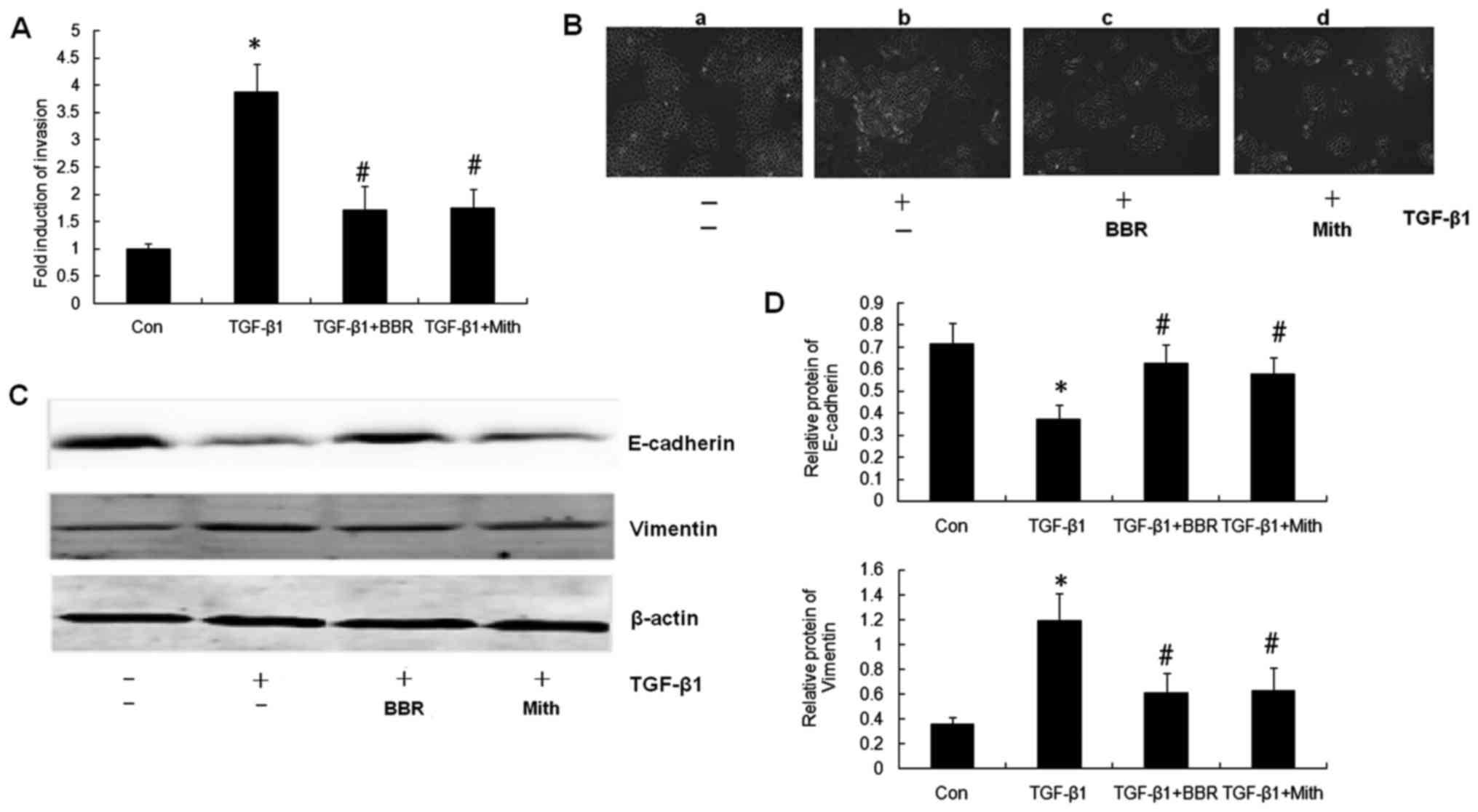
Berberine suppresses tumor invasion
and EMT by decreasing Sp1 expression in NPC cells
In order to explore the detailed pathway regulated
by Sp1 in the berberine-induced enhanced radiosensitivity of NPC
cells, we aimed to confirm whether berberine affects the tumor
invasion induced by TGF-β1. CNE-2 cells were treated with DMEM
(control group), TGF-β1 (5 ng/ml) (TGF-β group), TGF-β1+berberine
(50 µmol/l) (BBR group), or TGF-β1+mithramycin A (100 nmol/l) (Mith
group), and a Boyden chamber assay was used to determine the impact
of berberine on CNE-2 cell invasion. Tumor invasion was
significantly increased in the TGF-β1 group than that noted in the
control group (P<0.05) (Fig.
4A). This result showed that TGF-β1 promoted NPC cell
metastasis. Berberine and mithramycin A both inhibited CNE-2 cell
invasion induced by TGF-β1, and the differences between the TGF-β1
group and the BBR group or Mith group were significant
(P<0.05).
We further explored whether berberine could inhibit
TGF-β1-induced EMT. CNE-2 cells showed a mesenchymal phenotype
after treatment with TGF-β1 (5 ng/ml) for 48 h. However, after
adding berberine (50 µmol/l) or mithramycin A (100 nmol/l), the
cells changed from a mesenchymal back to an epithelial morphology
(Fig. 4B). To confirm the
inhibitory effect of berberine on EMT, we performed western
blotting to determine the protein expression of two EMT markers,
E-cadherin and vimentin. Compared with the control group, TGF-β1
decreased the expression of epithelial phenotype marker E-cadherin
and increased the expression of mesenchymal phenotype marker
vimentin (P<0.05) (Fig. 4C and
D). After treatment with berberine or mithramycin A, the
E-cadherin protein was increased and vimentin protein was decreased
significantly (P<0.05). These findings indicate that berberine
inhibited the EMT process by TGF-β1 through increased E-cadherin
and decreased vimentin expression in the NPC cells, and its
mechanism may be associated with decreased Sp1 expression.
Discussion
In the present study, we showed that berberine
inhibited proliferation and induced cell cycle arrest in the G0/G1
phase and apoptosis in CNE-2 cells. Berberine enhanced the
radiosensitivity of CNE-2 cells and this was associated with the
downregulation of Sp1 mRNA and protein expression. Furthermore,
berberine suppressed tumor invasion and EMT of CNE-2cells induced
by TGF-β1, as evidenced by increased expression of epithelial
marker E-cadherin and decreased expression of mesenchymal marker
vimentin. Selective inhibition of Sp1 by mithramycin A enhanced
radiosensitivity and inhibited tumor invasion and EMT of CNE-2
cells induced by TGF-β1. Therefore, Sp1 participates in the
enhanced radiosensitivity induced by berberine and is required for
TGF-β1-induced invasion and EMT of NPC cells.
Radioresistance is the primary cause of poor
prognosis of advanced-stage NPC. Therefore, biomarkers with
radiosensitizing effects have been widely explored to improve the
clinical outcome of NPC (14). We
found that berberine enhanced the radiosensitivity of NPC cells,
which may be associated with reduced proliferation, cell cycle
arrest and apoptosis of NPC cells. The results of our study are
consistent with other studies showing the radiosensitizing effects
of berberine in NPC, which was associated with decreased expression
of hypoxia-inducible factor-1α (12). In fact, berberine was found to
enhance the radiosensitivity of a variety of cancers, including
esophageal, breast, esophageal squamous and prostate cancers
(15–18). This indicates that the
radiosensitizing effects of berberine are universal in many types
of cancers, and its mechanism in cancers including NPC warrants
extensive investigation.
In the present study, we firstly found that
berberine can downregulate Sp1 expression in NPC cells, which is
involved in the radiosensitizing effects of berberine. Sp1 is one
transcription factor and can bind GC/GT-rich promoter elements and
regulate the promoter activity of multiple genes involved in cell
cycle, differentiation and oncogenesis (19). In NPC cells, Sp1 cooperates with
c-MYC to bind the promoter of the BMI1 gene and participates in the
pathogenesis of NPC (20). In fact,
Sp1 was overexpressed in advanced NPC tissues and downregulation of
Sp1 inhibited cell proliferation and clonogenicity of NPC cells
(21). Sp1 DNA binding was
increased within 30 min after ionizing radiation in radioresistant
human malignant melanoma (U1-Mel) cells (22), which indicates that Sp1 may
participate in the induction of genes in radioresistance. However,
currently there are few studies concerning the association between
Sp1 and the radioresistance of cancers. Our previous study found a
correlation between high expression of Sp1 and radioresistance of
NPC (13). Furthermore, this study
confirmed Sp1 as a therapeutic target of the radiosensitizing
effects on NPC induced by berberine.
EMT is a cellular process characterized by decreased
cell-cell adhesion and increased cell motility, and is involved in
cancer progression, metastasis and increased resistance to
radiotherapy (23). EMT not only
decreases the radiosensitivity of cancers but also can be induced
by radiation itself. Therefore, inhibition of EMT is an effective
therapeutic strategy for enhancing the radiosensitivity of cancers
(24). Berberine has been reported
to suppress EMT in lung cancer, cervical cancer, prostate cancer
and NPC (25–28). However, whether inhibition of EMT is
involved in the radiosensitizing effects of berberine remains
unclear. This study found that berberine inhibited TGF-β1-induced
EMT in NPC cells, and increased epithelial marker E-cadherin and
decreased mesenchymal marker vimentin. It has been reported that
E-cadherin loss in EMT promotes radioresistance in human tumor
cells (29). Therefore, in our
study increased E-cadherin and decreased EMT may also have induced
the radiosensitizing effects of berberine. In addition, EMT is an
inducer of tumor invasion. In the present study, berberine
inhibited CNE-2 cell invasion induced by TGF-β1, and suppression of
EMT may be an important cause of this decreased invasion. This
indicates that in NPC cells, berberine inhibits the EMT process,
and subsequently enhances radiosensitivity and decreases
invasion.
To further explore the detailed mechanism through
which berberine suppresses EMT, we treated NPC cells with
mithramycin A, a selective Sp1 inhibitor. We found that inhibition
of Sp1 enhanced radiosensitivity and suppressed invasion and EMT,
which was similar to the effects of berberine. This indicates that
Sp1 lies in the upstream of EMT and radioresistance in NPC cells.
The relationship between Sp1 and EMT is unclear. It was reported
that Sp1 is essential to maintain an epithelial state and inhibits
the EMT process (30). However,
other studies demonstrated a significant inhibitory effect on EMT
by the silencing Sp1 (31). Sp1 can
also cooperate with ZEB2 to activate mesenchymal genes and promote
EMT (32). The controversial effect
of Sp1 on EMT may be caused by the fact that Sp1 is a transcription
factor and can activate or repress various genes. In fact, Sp1
mainly acts as a promoter of EMT. High expression of Sp1 has been
reported to enhance invasion and migration in lung, breast, ovarian
and pancreatic cancer (33–36). Therefore, targeting and silencing of
Sp1 can inhibit the invasion and migration of cancers (37,38).
Our study presents berberine as a novel agent by inhibition of Sp1,
and suppressed EMT and invasion and enhanced radiosensitivity of
NPC. We also hypothesize a signaling pathway
Sp1-EMT-radioresistance in epithelial cancers, at least in NPC.
In conclusion, the present study confirmed berberine
as a radiosensitizing agent for NPC, and the underlying mechanism
is associated with inhibition of Sp1, EMT and invasion. Our study
demonstrated the regulation of Sp1 by berberine and suggests that
Sp1 may be one important promoter of radioresistance in epithelial
cancers. Our study provides new insights into the regulation of Sp1
and EMT in radioresistance and cancer migration and progression.
Further study is needed to explore the detailed mechanisms through
which Sp1 promotes EMT and radioresistance in NPC and other
epithelial cancers.
References
|
1
|
Wei WI and Sham JS: Nasopharyngeal
carcinoma. Lancet. 365:2041–2054. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lo KW, Chung GT and To KF: Deciphering the
molecular genetic basis of NPC through molecular, cytogenetic, and
epigenetic approaches. Semin Cancer Biol. 22:79–86. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee AW: Contribution of radiotherapy to
function preservation and cancer outcome in primary treatment of
nasopharyngeal carcinoma. World J Surg. 27:838–843. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen WC, McBride WH, Chen SM, Lee KF,
Hwang TZ, Jung SM, Shau H, Liao SK, Hong JH and Chen MF: Prediction
of poor survival by cyclooxygenase-2 in patients with T4
nasopharyngeal cancer treated by radiation therapy: clinical and in
vitro studies. Head Neck. 27:503–512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Loomans HA and Andl CD: Intertwining of
activin A and TGFβ signaling: dual roles in cancer progression and
cancer cell invasion. Cancers (Basel). 7:70–91. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roberts AB and Wakefield LM: The two faces
of transforming growth factor beta in carcinogenesis. Proc Natl
Acad Sci USA. 100:8621–8623. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Katsuno Y, Lamouille S and Derynck R:
TGF-β signaling and epithelial-mesenchymal transition in cancer
progression. Curr Opin Oncol. 25:76–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang P, Liu H, Xia F, Zhang QW, Zhang YY,
Zhao Q, Chao ZH, Jiang ZW and Jiang CC: Epithelial-mesenchymal
transition is necessary for acquired resistance to cisplatin and
increases the metastatic potential of nasopharyngeal carcinoma
cells. Int J Mol Med. 33:151–159. 2014.PubMed/NCBI
|
|
9
|
Kumar A, Chopra Ekavali K, Mukherjee M,
Pottabathini R and Dhull DK: Current knowledge and pharmacological
profile of berberine: an update. Eur J Pharmacol. 761:288–297.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang F, Wang D, Duan C, Huang D, Wu Y,
Chen Y, Wang W, Xie C, Meng J, Wang L, et al: Berberine inhibits
metastasis of nasopharyngeal carcinoma 5–8F cells by targeting Rho
kinase-mediated Ezrin phosphorylation at threonine 567. J Biol
Chem. 284:27456–27466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsang CM, Cheung YC, Lui VW, Yip YL, Zhang
G, Lin VW, Cheung KC, Feng Y and Tsao SW: Berberine suppresses
tumorigenicity and growth of nasopharyngeal carcinoma cells by
inhibiting STAT3 activation induced by tumor associated
fibroblasts. BMC Cancer. 13:6192013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang C, Yang X, Zhang Q, Yang B, Xu L,
Qin Q, Zhu H, Liu J, Cai J, Tao G, et al: Berberine radiosensitizes
human nasopharyngeal carcinoma by suppressing hypoxia-inducible
factor-1α expression. Acta Otolaryngol. 134:185–192. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Kang M, Qin YT, Wei ZX, Xiao JJ
and Wang RS: Sp1 is over-expressed in nasopharyngeal cancer and is
a poor prognostic indicator for patients receiving radiotherapy.
Int J Clin Exp Pathol. 8:6936–6943. 2015.PubMed/NCBI
|
|
14
|
Chen W and Hu GH: Biomarkers for enhancing
the radiosensitivity of nasopharyngeal carcinoma. Cancer Biol Med.
12:23–32. 2015.PubMed/NCBI
|
|
15
|
Liu Q, Jiang H, Liu Z, Wang Y, Zhao M, Hao
C, Feng S, Guo H, Xu B, Yang Q, et al: Berberine radiosensitizes
human esophageal cancer cells by downregulating homologous
recombination repair protein RAD51. PLoS One. 6:e234272011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Liu Q and Yang Q:
Radiosensitization effects of berberine on human breast cancer
cells. Int J Mol Med. 30:1166–1172. 2012.PubMed/NCBI
|
|
17
|
Yang X, Yang B, Cai J, Zhang C, Zhang Q,
Xu L, Qin Q, Zhu H, Ma J, Tao G, et al: Berberine enhances
radiosensitivity of esophageal squamous cancer by targeting HIF-1α
in vitro and in vivo. Cancer Biol Ther. 14:1068–1073. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Q, Zhang C, Yang X, Yang B, Wang J,
Kang Y, Wang Z, Li D, Huang G, Ma Z, et al: Berberine inhibits the
expression of hypoxia induction factor-1alpha and increases the
radiosensitivity of prostate cancer. Diagn Pathol. 9:982014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Davie JR, He S, Li L, Sekhavat A, Espino
P, Drobic B, Dunn KL, Sun JM, Chen HY, Yu J, et al: Nuclear
organization and chromatin dynamics - Sp1, Sp3 and histone
deacetylases. Adv Enzyme Regul. 48:189–208. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang HB, Liu GH, Zhang H, Xing S, Hu LJ,
Zhao WF, Xie B, Li MZ, Zeng BH, Li Y, et al: Sp1 and c-Myc regulate
transcription of BMI1 in nasopharyngeal carcinoma. FEBS J.
280:2929–2944. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang JP, Zhang H, Wang HB, Li YX, Liu GH,
Xing S, Li MZ and Zeng MS: Down-regulation of Sp1 suppresses cell
proliferation, clonogenicity and the expressions of stem cell
markers in nasopharyngeal carcinoma. J Transl Med. 12:2222014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang CR, Wilson-Van Patten C, Planchon SM,
Wuerzberger-Davis SM, Davis TW, Cuthill S, Miyamoto S and Boothman
DA: Coordinate modulation of Sp1, NF-kappa B, and p53 in confluent
human malignant melanoma cells after ionizing radiation. FASEB J.
14:379–390. 2000.PubMed/NCBI
|
|
23
|
Marie-Egyptienne DT, Lohse I and Hill RP:
Cancer stem cells, the epithelial to mesenchymal transition (EMT)
and radioresistance: potential role of hypoxia. Cancer Lett.
341:63–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Zheng L, Sun Y, Wang T and Wang
B: Tangeretin enhances radiosensitivity and inhibits the
radiation-induced epithelial-mesenchymal transition of gastric
cancer cells. Oncol Rep. 34:302–310. 2015.PubMed/NCBI
|
|
25
|
Qi HW, Xin LY, Xu X, Ji XX and Fan LH:
Epithelial-to-mesenchymal transition markers to predict response of
Berberine in suppressing lung cancer invasion and metastasis. J
Transl Med. 12:222014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chu SC, Yu CC, Hsu LS, Chen KS, Su MY and
Chen PN: Berberine reverses epithelial-to-mesenchymal transition
and inhibits metastasis and tumor-induced angiogenesis in human
cervical cancer cells. Mol Pharmacol. 86:609–623. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu CH, Tang WC, Sia P, Huang CC, Yang PM,
Wu MH, Lai IL and Lee KH: Berberine inhibits the metastatic ability
of prostate cancer cells by suppressing epithelial-to-mesenchymal
transition (EMT)-associated genes with predictive and prognostic
relevance. Int J Med Sci. 12:63–71. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li CH, Wu DF, Ding H, Zhao Y, Zhou KY and
Xu DF: Berberine hydrochloride impact on physiological processes
and modulation of twist levels in nasopharyngeal carcinoma CNE-1
cells. Asian Pac J Cancer Prev. 15:1851–1857. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Theys J, Jutten B, Habets R, Paesmans K,
Groot AJ, Lambin P, Wouters BG, Lammering G and Vooijs M:
E-cadherin loss associated with EMT promotes radioresistance in
human tumor cells. Radiother Oncol. 99:392–397. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kolesnikoff N, Attema JL, Roslan S, Bert
AG, Schwarz QP, Gregory PA and Goodall GJ: Specificity protein 1
(Sp1) maintains basal epithelial expression of the miR-200 family:
implications for epithelial-mesenchymal transition. J Biol Chem.
289:11194–11205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sung WJ, Kim KH, Kim YJ, Chang YC, Lee IH
and Park KK: Antifibrotic effect of synthetic Smad/Sp1 chimeric
decoy oligodeoxynucleotide through the regulation of epithelial
mesenchymal transition in unilateral ureteral obstruction model of
mice. Exp Mol Pathol. 95:136–143. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nam EH, Lee Y, Park YK, Lee JW and Kim S:
ZEB2 upregulates integrin α5 expression through cooperation with
Sp1 to induce invasion during epithelial-mesenchymal transition of
human cancer cells. Carcinogenesis. 33:563–571. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim WY, Jang JY, Jeon YK, Chung DH, Kim YG
and Kim CW: Syntenin increases the invasiveness of small cell lung
cancer cells by activating p38, AKT, focal adhesion kinase and SP1.
Exp Mol Med. 46:e902014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kong LM, Liao CG, Zhang Y, Xu J, Li Y,
Huang W, Zhang Y, Bian H and Chen ZN: A regulatory loop involving
miR-22, Sp1, and c-Myc modulates CD147 expression in breast cancer
invasion and metastasis. Cancer Res. 74:3764–3778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao J, Ye W, Wu J, Liu L, Yang L, Gao L,
Chen B, Zhang F, Yang H and Li Y: Sp1-CD147 positive feedback loop
promotes the invasion ability of ovarian cancer. Oncol Rep.
34:67–76. 2015.PubMed/NCBI
|
|
36
|
Tan Y, Yin H, Zhang H, Fang J, Zheng W, Li
D, Li Y, Cao W, Sun C, Liang Y, et al: Sp1-driven up-regulation of
miR-19a decreases RHOB and promotes pancreatic cancer. Oncotarget.
6:17391–17403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang J, Lv X, Fan L, Huang G, Zhan Y,
Wang M and Lu H: MicroRNA-27b suppresses growth and invasion of
NSCLC cells by targeting Sp1. Tumour Biol. 35:10019–10023. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang R, Luo H, Wang S, Chen W, Chen Z,
Wang HW, Chen Y, Yang J, Zhang X, Wu W, et al: MicroRNA-377
inhibited proliferation and invasion of human glioblastoma cells by
directly targeting specificity protein 1. Neuro-oncol.
16:1510–1522. 2014. View Article : Google Scholar : PubMed/NCBI
|