Introduction
Hepatocellular carcinoma (HCC) is one of the most
lethal malignant tumors in the world (1,2).
During the past decades, plenty of research regarding the genetic
markers (i.e. abnormal gene expression as well as the genomic
aberrations) have been accumulated for HCC (3–7). The
primary risk factors for the pathogenesis of HCC have been
elucidated, and the multiple steps involved in hepatocarcinogenesis
have been well defined in recent years (8). In spite of these associations between
the risk factors and the development of HCC being well-established
(4,9), no clear picture of the actual
mechanisms of how and what consequences these factors have leading
to malignant transformation has emerged yet.
Sox2 is a transcription factor that controls the
expression of a number of target genes through forming a trimeric
complex with Oct4 on DNA (10).
Until now, the relationship between Sox2 and tumorigenesis as well
as overall survival (OS) has been well elucidated in several solid
tumors, such as breast, esophageal and lung cancer (11–16).
In addition, Sox2 has also been implicated in the progression of
HCC. For instance, the expression levels of Sox2 and its co-factor
Oct4 correlate with an aggressive phenotype and low survival rate
of HCC (17). HCC cells
overexpressing Sox2 are characterized by increased ability of
epithelial-mesenchymal transition (EMT), invasion and clonal
formation (18). miR126, which is a
tumor suppressor, inhibits cell growth through reducing the
expression of Sox2 (19). These
studies indicate that Sox2 functions as an oncogene in HCC.
However, it is still unclear through which pathway Sox2 promotes
tumorigenesis.
PD-L1 (also known as B7-H1), a gene encoding an
immune inhibitory receptor ligand, is expressed in various types of
cancer and immune cells (i.e. B cells, T cells, and dendritic
cells) (20–22). Binding of PD-L1 to its receptor PD-1
(the programmed cell death-1 receptor) leads to tumor evasion from
host immune system (23), which is
achieved through suppressing T-cell response, migration,
proliferation and restricting the tumor cell killing ability of T
cells (21,24–26).
Immunotherapeutics targeting the PD-L1/PD-1 pathway are currently
in clinical trials and have shown impressive response rates in
patients, particularly for non-small cell lung cancer (NSCLC),
bladder cancer, renal cell carcinoma and melanoma (27,28).
It has been reported that the PD-L1 expression in HCC is
significantly associated with tumor malignancy and the risk of
postoperative recurrence. Therefore, PD-L1 might represent a
potential target for HCC immunotherapy as well as a biomarker that
aids in determining the prognosis both before and after therapy
(29).
In this study, we investigated the expression and
transcriptional regulation of Sox2 and PD-L1 in HCC. We report that
the expression level of Sox2 and PD-L1 is higher in HCC tissues and
cell lines compared with adjacent non-tumor tissues and normal
liver cell lines. Furthermore, the expression of Sox2 and PD-L1 in
HCC tissues are positively correlated with each other.
Functionally, we also demonstrate that knockdown of Sox2 reduces
the cell vitality, arrests the cell growth and induces the
apoptosis of HCC cell lines. Moreover, Sox2 regulates the
expression of PD-L1 through directly binding to the Sox2 consensus
binding site on PD-L1 promoter region and regulating the promoter
activity of PD-L1. This study describes the crucial role of Sox2 in
HCC, and for the first time we studied the relationship between
Sox2 and PD-L1.
Materials and methods
Cell culture
HL-7702, HepG2, SMMC7721, Huh7 and HEK293 cells were
cultured in DMEM (Gibco, Carlsbad, CA, USA) containing 10% fetal
bovine serum (Gibco).
RNA extraction and reverse
transcription-quantitive PCR (RT-qPCR)
Total RNA of cell lines (HL-7702, HepG2, SMMC7721,
and Huh7) and patient tissues (hepatocellular carcinoma tissue and
adjacent non-tumor tissue) were extracted using RNeasy Mini kit
(Qiagen, Valencia, CA, USA) according to the manufacturer's
protocol. RNase-free DNase I (Invitrogen, Carlsbad, CA, USA) were
used to remove genomic DNA. Reverse-transcription was carried out
with the Superscript III Reverse Transcriptase (Invitrogen). QPCR
assays were performed with the SYBE Premix Ex Taq™ (Takara Bio,
Otsu, Japan) on ABI ViiA 7 Real-time PCR System (Applied
Biosystems, Foster City, CA, USA). The relative expression level of
Sox2 and PD-L1 were calculated as 2−[Ct (Gene)-Ct
(GAPDH)] (30). Each assay
was performed three times. The primers used for RT-qPCR are shown
in Table I.
 | Table I.Primer sequences used for mRNA
expression level analysis through qPCR. |
Table I.
Primer sequences used for mRNA
expression level analysis through qPCR.
| Name | Primers sequence
(5′-3′) |
|---|
| Sox2-RT-F: |
TACAGCATGTCCTACTCGCAG |
| Sox2-RT-R: |
GAGGAAGAGGTAACCACAGGG |
| PD-L1-RT-F: |
TGGCATTTGCTGAACGCATTT |
| PD-L1-RT-R: |
TGCAGCCAGGTCTAATTGTTTT |
| GAPDH-RT-F: |
TGTTCGTCATGGGTGTGAAC |
| GAPDH-RT-R: |
ATGGCATGGACTGTGGTCAT |
RNA interference (RNAi)
Lipofectamine™ 2000 (Invitrogen) was used for siRNA
delivery. SMMC7721 and HepG2 cells (2.5×105) were seeded
in 6-well plate one day before transfection. A total amount of 100
pmol siRNA were diluted with 250 µl of DMEM without serum and mixed
gently. A total amount of 5 µl Lipofectamine 2000 were mixed with
250 µl of DMEM and incubated for 5 min at room temperature. Then
the diluted DNA and the diluted Lipofectamine 2000 were firstly
mixed gently and incubated for 20 min at room temperature. After 20
min of incubation, 500 µl of DMEM/Lipo/siRNA mix were added to each
well containing cells, and mixed gently. The cells were then
incubated in a CO2 incubator containing 5%
CO2 at 37°C. Cells were collected after 72 h of
transfection for mRNA and protein extraction. A Sox2 specific siRNA
sequence (siSox2: 5′-CGGCUCUGUAUUAUUUGAATTA-3′) was used for RNA
interference. A mismatch siRNA sequence (siNC:
5′-UUCUCCGAACGUGUCACGUTT-3′) was used as negative control.
Cell proliferation assay
Cell proliferation was detected by the Cell Counting
Kit-8 (CCK-8) (Dojindo Laboratories, Kumamoto, Japan) and BrdU
(Roche, Mannheim, Germany) assays. The cell viability of HepG2 and
SMMC7721 cells was detected by the CCK-8 (Dojindo Laboratories,
Kumamoto, Japan). Briefly, 10 µl of CCK-8 reagent was added at the
24, 48 and 72 h siRNA posttransfection. Then cells were incubated
at 37°C for 4 h, and the number of viable cells was evaluated
through measuring absorbance at 450 nm.
EdU staining
HepG2 and SMMC7721 cells transfected with siRNA were
used for EdU assay with Cell-Light™ EdU kit (RiboBio, Guangzhou,
China) according to the manufacturer's protocol. Images were
captured using a Leica DMI6000B microscope (Leica Microsystems,
Wetzlar, Germany).
Apoptosis analysis
siRNA-transfected HepG2 and SMMC7721 were collected
for apoptosis analysis. Annexin V-FITC and PI (BD Biosciences) were
used for detection of apoptotic cells.
Electrophoretic mobility shift assay
(EMSA)
EMSA was performed using Lightshift Chemiluminescent
EMSA kit (Pierce, Rockford, IL, USA) according to the
manufacturer's instructions. Biotin-labeled double-stranded
oligonucleotide (5′-TATGACACCATCGTCTGTCATC-3′) containing
the consensus Sox2 motif was used as EMSA probe. An unlabeled
double-stranded oligonucleotide was used as competitor probe. An
unlabeled-mutated oligonucleotide (5′-TATGACACGTACCACTGTCATC) was used as
negative competitor probe. Nuclear protein was extracted from HepG2
cells. Anti-Sox2 antibody (#5024S, Cell Signaling Technology, Inc.,
Danvers, MA, USA) was used to supershift the DNA-protein
complex.
Chromatin
immunoprecipitation-quantitative-PCR (ChIP-qPCR)
ChIP was performed according to the manufacturer
manual (Active Motif, Carlsbad, CA, USA) in HepG2 cells with the
anti-Sox2 antibody (#5024S). The qPCR assay was carried out with
the SYBE Premix Ex Taq (Takara Bio) on ABI ViiA 7 Real-time PCR
System. Normal rabbit IgG was used as negative control. ChIP-qPCR
assay was performed in triplicate. The primers used for ChIP-qPCR
are as follows: PD-L1 (for ChIP-qPCR)-F: AAGAAAAGGGAGCACACAGG,
PD-L1 (for ChIP-qPCR)-R: GCCCAAGATGACAGACGATG.
Plasmid
The PD-L1 promoter region was PCR amplified from
HepG2 cells and cloned into the pGL3-basic vector (PD-L1-WT).
Mutation of the Sox2 binding site on PD-L1-WT was performed by Quik
Change Site Mutagenesis kit (Stratagene, La Jolla, CA, USA)
according to the manufacturer's manual. Human Sox2 cDNA was
amplified from HepG2 cells by RT-PCR and cloned into the pSG5
vector. The primers used for cloning are as follows:
PD-L1-Promoter-F: GGGGTACCAGAAGGAAAGGCAAACAAC, PD-L1-Promoter-R:
CCGCTCGAGCTTTGGGTTAGTGAATGGG; PD-L1-Mutation-F:
TACTTAAGTAAATTATGACATGCAGACGTGTCATCTTGG, PD-L1-Mutation-R:
CCAAGATGACACGTCTGCATGTCATAATTTACTTAAGTA; Sox2-cDNA-F:
CGGGATCCTCTTCGCCTGATTTTCCTCG, Sox2-cDNA-R:
GGAATTCCCTCCCATTTCCCTCGTTTT.
Luciferase
HEK293T cells were transfected using Lipofectamine
2000 (Invitrogen). Luciferase activity was detected by the Dual
Luciferase Assay (Promega, Madison, WI, USA) after 24 h of
transfection. Co-transfection of Renilla luciferase plasmid
was used as the internal control for transfection efficiency.
Statistical analysis
Data in this research was presented as mean ±SD, and
t-test or one-way ANOVA was used among groups. p<0.05 was
considered statistically significant. Data were analyzed by using
GraphPad Prism 5 for Windows (IBM, USA).
Results
Sox2 and PD-L1 expression are
significantly higher in hepatocellular carcinoma (HCC) tissues than
in adjacent non-tumor tissues
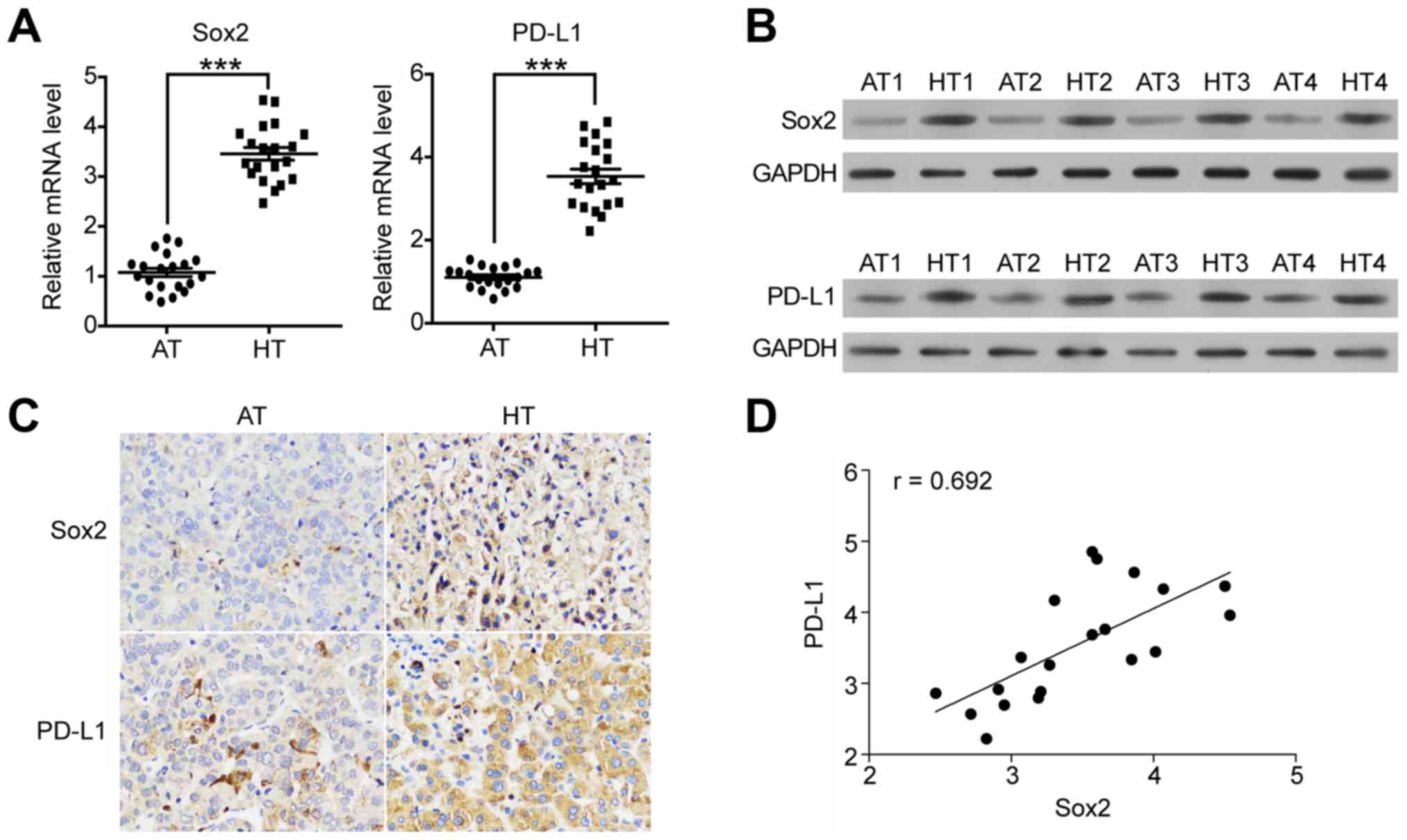
We initially compared Sox2 and PD-L1 expression in
HCC tissue samples and that in adjacent non-tumor tissue samples.
We performed RT-qPCR and western blotting to measure Sox2 and PD-L1
expression in HCC tissues versus adjacent non-tumor tissues from
HCC patients. As illustrated in Fig. 1A
and B, Sox2 expression in HCC tissues (HT) was significantly
higher than that in adjacent non-tumor tissues (AT) in both mRNA
and protein levels. PD-L1 showed similar expression pattern as Sox2
(Fig. 1A, right panel, Fig. 1B, lower panel). To confirm this
expression pattern, we conducted immunohistochemistry (IHC)
staining to detect the Sox2 and PD-L1 protein in the HCC tissue and
adjacent non-tumor tissue samples. As compared with normal tissue,
the Sox2 (Fig. 1C, upper panel) and
PD-L1 (Fig. 1C, lower panel) signal
is stronger in HCC tissue. Altogether, these analyses indicated
that the expression of Sox2 and PD-L1 might be correlated with each
other. Thus, we further investigated the relationship between the
expression level of Sox2 and PD-L1. As shown in Fig. 1D, the expression level of Sox2 was
positively correlated with that of PD-L1 (r=0.692). Collectively,
these results suggest that both Sox1 and PD-L1 are highly expressed
in HCC tissue, and their expression are positive co-related with
each other.
Sox2 and PD-L1 expression are
significantly higher in HCC cell lines than that of normal liver
cell line
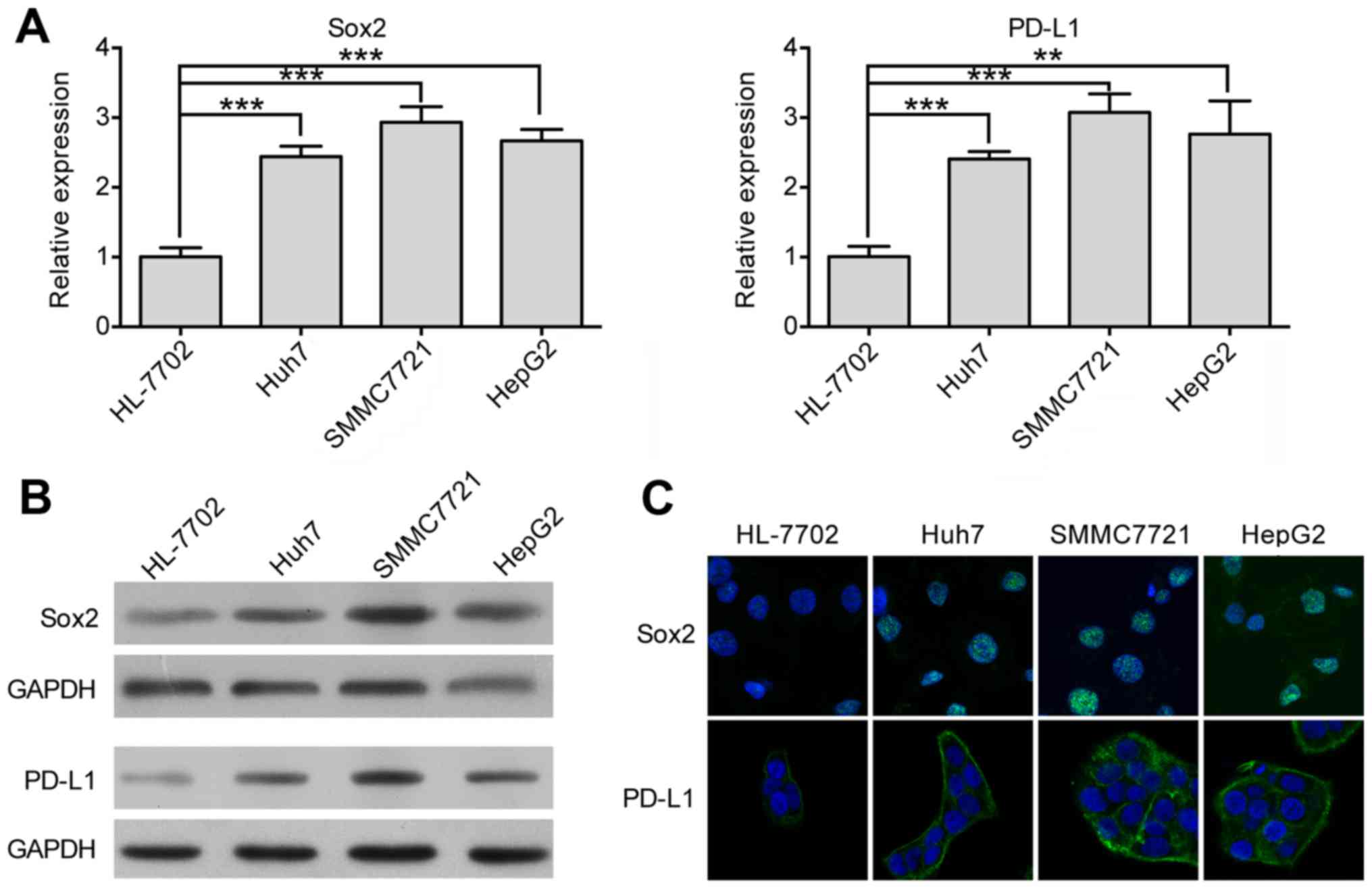
We performed RT-qPCR to measure the mRNA level of
Sox2 and PD-L1 in HCC cell lines (Huh7, SMMC7721, and HepG2) and
normal liver cell line (HL-7702). As shown in Fig. 2A, Sox2 and PD-L1 expression in HCC
cell lines were significantly higher than that in the normal liver
cell line. We also analyzed the protein level of both proteins
through western blot assays. As shown in Fig. 2B, the protein level of Sox2 and
PD-L1 were higher in HCC cell lines than that in the normal liver
cell line. Moreover, we performed immunofluorescence assays to
visualize the expression of Sox2 and PD-L1 globally. As illustrated
in Fig. 2C, both Sox2 and PD-L1,
even though they localized in the different cellular component,
showed higher fluorescence intensity in HCC cell lines than in
normal liver cell line. These data are consistent with the
aforementioned results shown in Fig.
1 and provide strong evidence that Sox2 and PD-L1 are both
highly expressed in HCC.
Knockdown of Sox2 represses the
proliferation growth and induces apoptosis of HCC cells
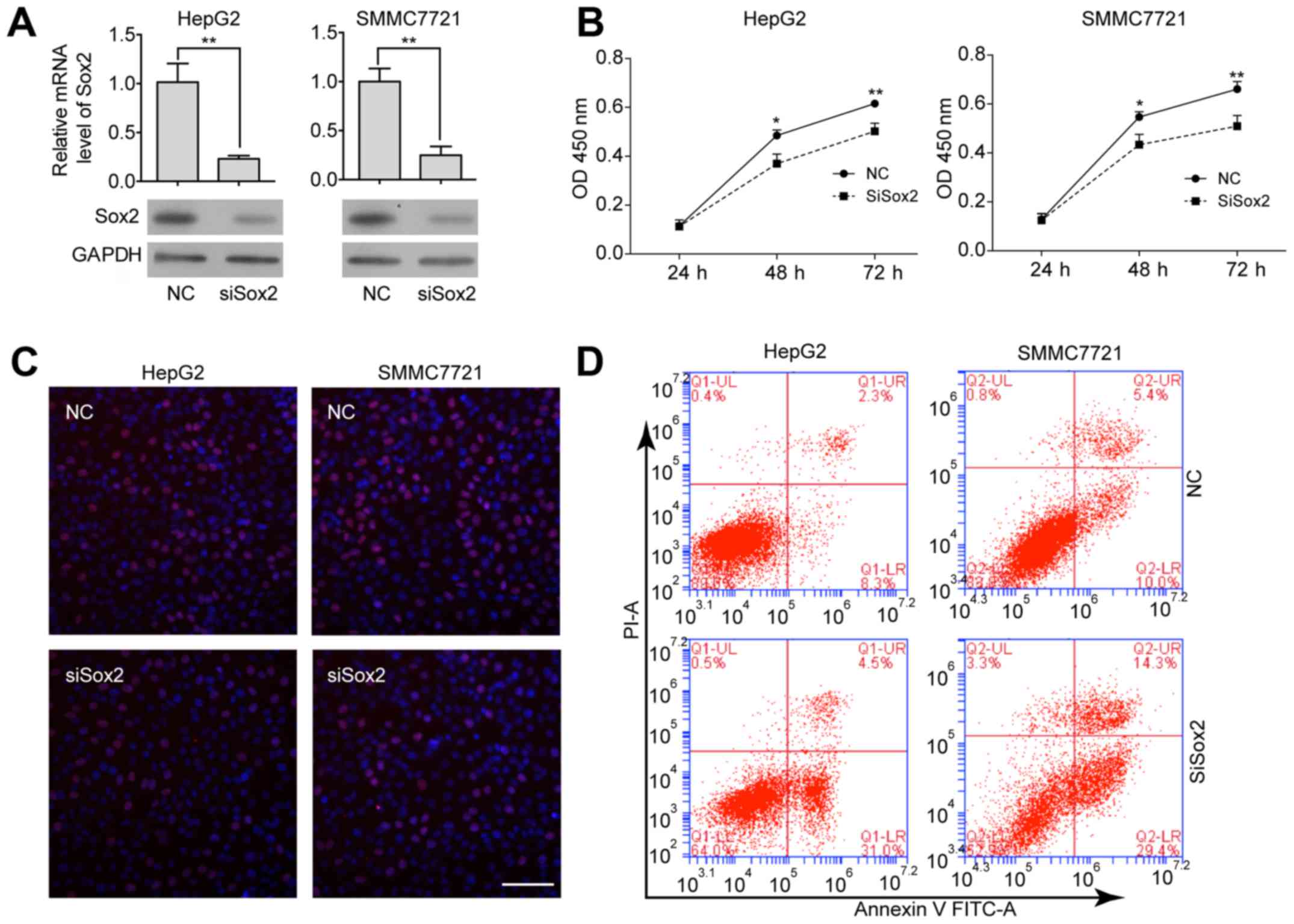
The finding that Sox2 is highly expressed in HCC
cells prompted us to elucidate the biological function of Sox2 in
HCC. We utilized an siRNA approach to knock down Sox2 expression in
Sox2 highly expressed HCC cell lines (HepG2 and SMMC7721), and
compared the cell proliferation ability and the status of cellular
apoptosis before and after Sox2 knockdown. The reduction of Sox2
expression in Sox2-specific siRNA transfected cells was confirmed
by qRT-PCR and western blot assays (Fig. 3A). The effect of Sox2 depletion on
cell proliferation was evaluated through using CCK-8 and EdU
assays. We observed a significant decrease in proliferation after
transfection of siSox2 (Fig. 3B and
C) in both HepG2 and SMMC7721 cells. Sox2 knockdown also led to
apoptosis of HCC cells, based on the observations of the percentage
of the Annexin V-positive and PI-negative cells (increased from 8.3
to 31.0% and 10.0 to 29.4%, in HepG2 and SMMC7221 after Sox2
knockdown, respectively) (Fig. 3D).
Taken together, these data indicate that the Sox2 oncogene is
required for the proliferation and growth of HCC cells.
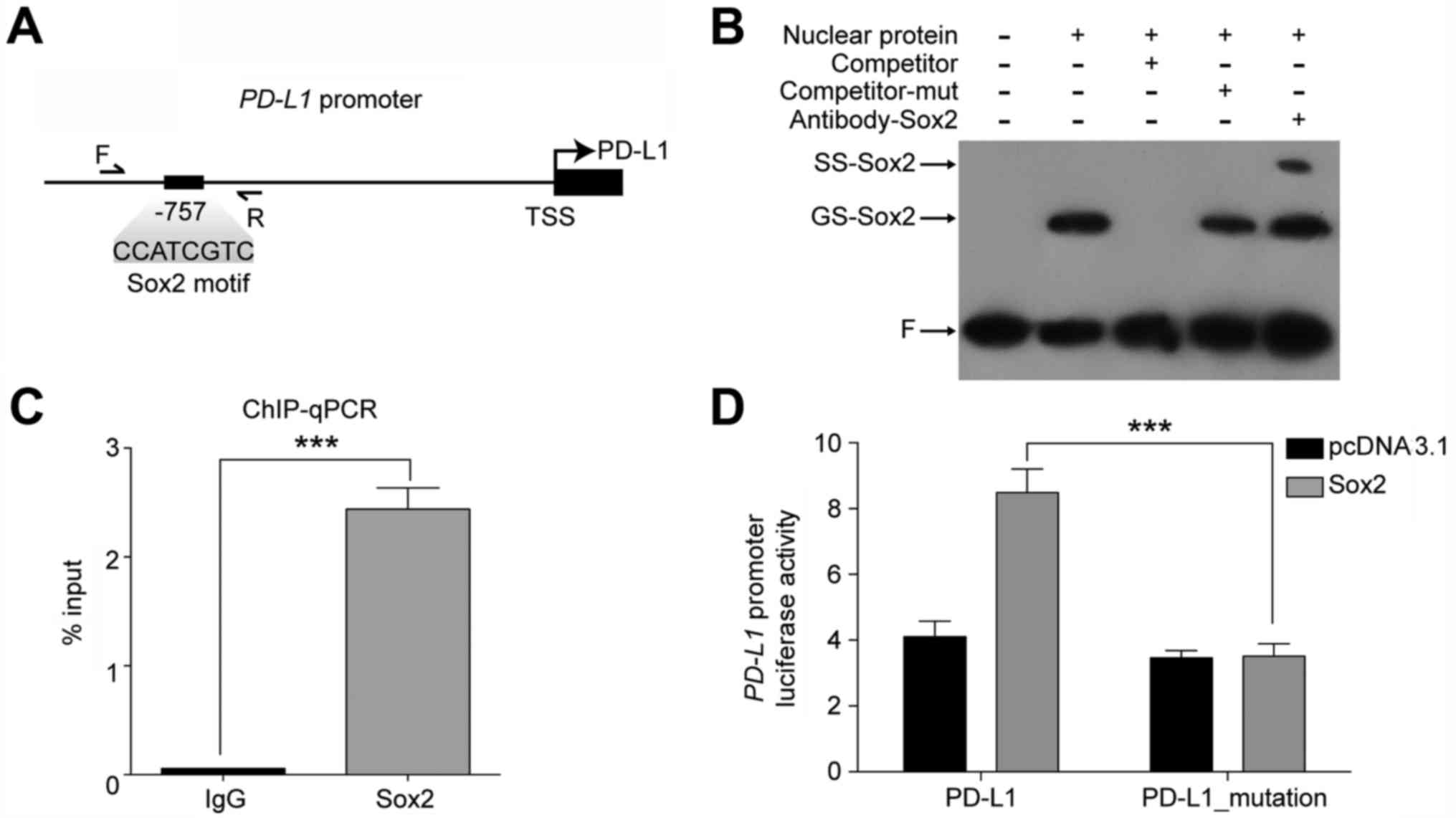
Sox2 transactivates PD-L1 through the
−757 region of the PD-L1 promoter
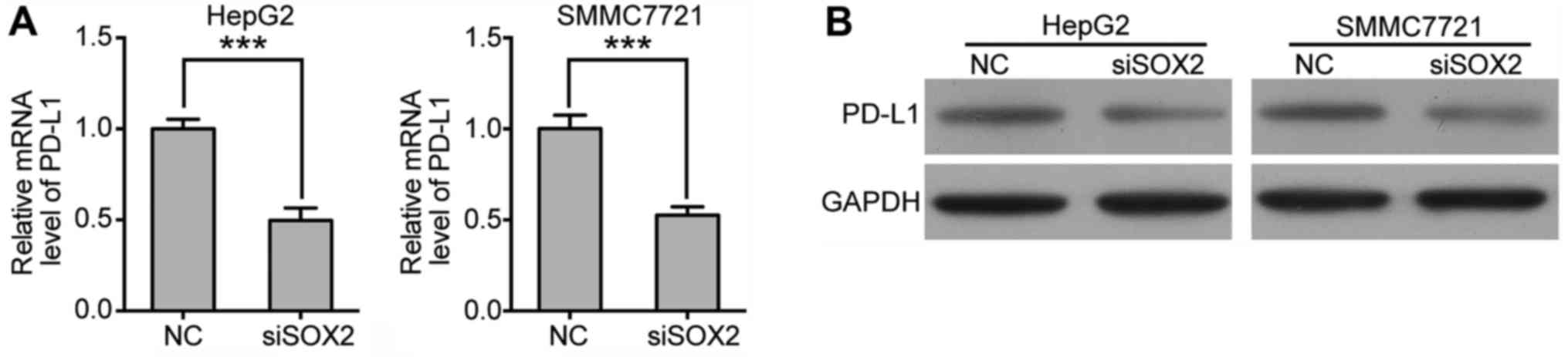
Next, we asked whether Sox2 regulates the expression
of PD-L1. Through RT-qPCR and western blotting, we found that the
expression level of PD-L1 was significantly decreased after Sox2
knockdown (Fig. 4A), which
indicated that the expression of PD-L1 might be regulated by Sox2.
One important problem that remains to be clarified is whether Sox2
directly target the PD-L1 promoter in HCC cells. To address this
question, we first investigated the transcription factor binding
sites within the promoter region of PD-L1 using the TRANSFAC
database (31). Of note, we found a
consensus Sox2 motif upstream of the transcription start site (TSS)
at position −757 (Fig. 5A). Then we
performed in vitro EMSA and in vivo ChIP-qPCR assay
on Sox2 motif containing promoter regions. As demonstrated in
Fig. 5B, when a 22 bp probe
contained the Sox2 motif was incubated with nuclear extracts from
HepG2, a specific DNA-protein complex was observed (lane 2,
GS-Sox2) and was supershifted by the anti-Sox2 antibody (lane 5,
SS-Sox2). This indicated that Sox2 bound to the promoter regions of
PD-L1 directly through the Sox2 motif. This was further verified by
ChIP-qPCR assays in HepG2 cells using primers marked in Fig. 5A and antibodies against Sox2.
To further investigate whether Sox2 transactivates
the regulatory region of PD-L1 through the Sox2 binding site
identified above, we performed luciferase reporter assays in
HEK-293T cells. We cloned the promoter region of PD-L1 that
contained Sox2 motif and TSS into a luciferase reporter plasmid,
which was defined as pGL3-PD-L1. We also constructed a
pGL3-PD-L1-mutant plasmid, in which the core sequence of Sox2 motif
around −757 bp was mutated from CCATCGTC to TGCAGACG. As shown in
Fig. 5D, co-expression of Sox2
resulted in significant activation of the PD-L1 promoter region. In
contrast, the transactivation ability of Sox2 on the PD-L1 promoter
region was significantly reduced as observed in the PD-L1 mutant
promoter. Collectively, the data indicate that Sox2 could
transactivate the PD-L1 promoter through the Sox2 motif located at
the −757 bp upstream of TSS.
Discussion
In the last few years, numerous studies have been
carried out to uncover the molecular markers during the development
of HCC (32,33); however, the actual mechanisms of how
these molecular markers lead to malignant transformation are not
yet well understood. Our data indicate that both Sox2 and PD-L1 are
highly expressed in hepatocellular carcinoma (HCC). Functional
analysis shows the expression of Sox2 is closely related to the
proliferation and growth of HCC cells. We also found that PD-L1, a
well-qualified immunotherapeutic target in various solid tumors, is
transactivated by Sox2. These findings provide novel insight into
the function and the interplay between Sox2 and PD-L1 in HCC, and
significantly enrich our understanding of the role of Sox2 in HCC
malignancy.
Sox2 is a potential biomarker for HCC prognosis
(18,34). Previous research including 75
patient samples concluded that high expression of Sox2 in HCC
tissues was positively correlated with malignancy and poor survival
(34). In our study, we show that
the expression of Sox2 is significantly higher in HCC tissue from
patient samples and HCC cell lines. This observation is consistent
with several previous reports. Moreover, we ivestigated the
function of Sox2 in HCC and found that Sox2 is needed for keeping
the proliferation ability and prevent HCC cells from apoptosis. Our
study not only provides direct evidence (in tissue samples and cell
lines) to support the expression pattern of Sox2 but also further
explores the Sox2 function. In summary, our finding supports the
fact that Sox2 may function as an oncogene in solid tumors
especially in HCC.
PD-L1 is an immunotherapeutic target in various
solid tumors, such as NSCLC, bladder cancer and renal cell
carcinoma (20,21,23,26).
We demonstrate here that the expression level of PD-L1 is higher in
HCC tissues/cells than that in normal tissues/cells. Recent
attention to the role of PD-L1 in the tumor progress of HCC has
provided worthy insights into the mechanism of HCC development and
associated therapeutic approaches. For example, the PD-L1
expression in HCC is associated with the tumor malignancy and the
postoperative recurrence risk (22,35).
Accordingly, we tentatively propose that PD-L1 might represent a
potential target for HCC immunotherapy.
We observed that the cellular location of PD-L1 and
Sox2 are different. In most instances, Sox2 localized only in the
nucleus (Fig. 2C). The distribution
pattern of Sox2 is due to Sox2 function as a transcriptional
factor, and it functions through binding to the promoter regions of
its target gene in the nucleus (10,36,37).
The location pattern we found here is different from a previous
report, which shows that Sox2 locates not only in the nucleus, but
also in the cytoplasm of HCC tissue and cell lines, this might be
caused by the different method we used for immunofluorescence. On
the other hand, as confirmed (38),
PD-L1 is located in the cytoplasm and on the cell membrane of tumor
cells and immune cells, we observed that PD-L1 indeed localized on
the cell membrane and partly in the cytoplasm. It seems that the
fluorescence single on the cell membrane is slightly stronger than
that in the cytoplasm.
In summary, our findings not only reveal the
relationship between Sox2 and PD-L1, but also enrich our
understanding of the potential role of Sox2 in HCC malignancy.
References
|
1
|
Farazi PA and DePinho RA: Hepatocellular
carcinoma pathogenesis: From genes to environment. Nat Rev Cancer.
6:674–687. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Simonetti RG, Liberati A, Angiolini C and
Pagliaro L: Treatment of hepatocellular carcinoma: A systematic
review of randomized controlled trials. Ann Oncol. 8:117–136. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mas VR, Fisher RA, Archer KJ, Yanek KC,
Williams B, Dumur CI and Maluf DG: Genes associated with
progression and recurrence of hepatocellular carcinoma in hepatitis
C patients waiting and undergoing liver transplantation:
Preliminary results. Transplantation. 83:973–981. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moradpour D and Blum HE: Pathogenesis of
hepatocellular carcinoma. Eur J Gastroenterol Hepatol. 17:477–483.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nam SW, Park JY, Ramasamy A, Shevade S,
Islam A, Long PM, Park CK, Park SE, Kim SY, Lee SH, et al:
Molecular changes from dysplastic nodule to hepatocellular
carcinoma through gene expression profiling. Hepatology.
42:809–818. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Poon TC, Wong N, Lai PB, Rattray M,
Johnson PJ and Sung JJ: A tumor progression model for
hepatocellular carcinoma: Bioinformatic analysis of genomic data.
Gastroenterology. 131:1262–1270. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Teufel A, Staib F, Kanzler S, Weinmann A,
Schulze-Bergkamen H and Galle PR: Genetics of hepatocellular
carcinoma. World J Gastroenterol. 13:2271–2282. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schafer DF and Sorrell MF: Hepatocellular
carcinoma. Lancet. 353:1253–1257. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mas VR, Maluf DG, Archer KJ, Yanek K, Kong
X, Kulik L, Freise CE, Olthoff KM, Ghobrial RM, McIver P, et al:
Genes involved in viral carcinogenesis and tumor initiation in
hepatitis C virus-induced hepatocellular carcinoma. Mol Med.
15:85–94. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rodda DJ, Chew JL, Lim LH, Loh YH, Wang B,
Ng HH and Robson P: Transcriptional regulation of nanog by OCT4 and
SOX2. J Biol Chem. 280:24731–24737. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bass AJ, Watanabe H, Mermel CH, Yu S,
Perner S, Verhaak RG, Kim SY, Wardwell L, Tamayo P, Gat-Viks I, et
al: SOX2 is an amplified lineage-survival oncogene in lung and
esophageal squamous cell carcinomas. Nat Genet. 41:1238–1242. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Y, Shi L, Zhang L, Li R, Liang J, Yu
W, Sun L, Yang X, Wang Y, Zhang Y, et al: The molecular mechanism
governing the oncogenic potential of SOX2 in breast cancer. J Biol
Chem. 283:17969–17978. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hussenet T, Dali S, Exinger J, Monga B,
Jost B, Dembelé D, Martinet N, Thibault C, Huelsken J, Brambilla E,
et al: SOX2 is an oncogene activated by recurrent 3q26.3
amplifications in human lung squamous cell carcinomas. PLoS One.
5:e89602010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hussenet T and du Manoir S: SOX2 in
squamous cell carcinoma: Amplifying a pleiotropic oncogene along
carcinogenesis. Cell Cycle. 9:1480–1486. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Justilien V, Walsh MP, Ali SA, Thompson
EA, Murray NR and Fields AP: The PRKCI and SOX2 oncogenes are
coamplified and cooperate to activate Hedgehog signaling in lung
squamous cell carcinoma. Cancer Cell. 25:139–151. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schoenhals M, Kassambara A, De Vos J, Hose
D, Moreaux J and Klein B: Embryonic stem cell markers expression in
cancers. Biochem Biophys Res Commun. 383:157–162. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang P, Qiu J, Li B, Hong J, Lu C, Wang
L, Wang J, Hu Y, Jia W and Yuan Y: Role of Sox2 and Oct4 in
predicting survival of hepatocellular carcinoma patients after
hepatectomy. Clin Biochem. 44:582–589. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun C, Sun L, Li Y, Kang X, Zhang S and
Liu Y: Sox2 expression predicts poor survival of hepatocellular
carcinoma patients and it promotes liver cancer cell invasion by
activating Slug. Med Oncol. 30:5032013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao C, Li Y, Zhang M, Yang Y and Chang L:
miR-126 inhibits cell proliferation and induces cell apoptosis of
hepatocellular carcinoma cells partially by targeting Sox2. Hum
Cell. 28:91–99. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Blank C, Kuball J, Voelkl S, Wiendl H,
Becker B, Walter B, Majdic O, Gajewski TF, Theobald M, Andreesen R,
et al: Blockade of PD-L1 (B7-H1) augments human tumor-specific T
cell responses in vitro. Int J Cancer. 119:317–327. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Herbst RS, Soria JC, Kowanetz M, Fine GD,
Hamid O, Gordon MS, Sosman JA, McDermott DF, Powderly JD, Gettinger
SN, et al: Predictive correlates of response to the anti-PD-L1
antibody MPDL3280A in cancer patients. Nature. 515:563–567. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zeng Z, Shi F, Zhou L, Zhang MN, Chen Y,
Chang XJ, Lu YY, Bai WL, Qu JH, Wang CP, et al: Upregulation of
circulating PD-L1/PD-1 is associated with poor post-cryoablation
prognosis in patients with HBV-related hepatocellular carcinoma.
PLoS One. 6:e236212011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nomi T, Sho M, Akahori T, Hamada K, Kubo
A, Kanehiro H, Nakamura S, Enomoto K, Yagita H, Azuma M, et al:
Clinical significance and therapeutic potential of the programmed
death-1 ligand/programmed death-1 pathway in human pancreatic
cancer. Clin Cancer Res. 13:2151–2157. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Butte MJ, Keir ME, Phamduy TB, Sharpe AH
and Freeman GJ: Programmed death-1 ligand 1 interacts specifically
with the B7-1 costimulatory molecule to inhibit T cell responses.
Immunity. 27:111–122. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park JJ, Omiya R, Matsumura Y, Sakoda Y,
Kuramasu A, Augustine MM, Yao S, Tsushima F, Narazaki H, Anand S,
et al: B7-H1/CD80 interaction is required for the induction and
maintenance of peripheral T-cell tolerance. Blood. 116:1291–1298.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Paterson AM, Brown KE, Keir ME, Vanguri
VK, Riella LV, Chandraker A, Sayegh MH, Blazar BR, Freeman GJ and
Sharpe AH: The programmed death-1 ligand 1:B7-1 pathway restrains
diabetogenic effector T cells in vivo. J Immunol. 187:1097–1105.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Motzer RJ, Rini BI, McDermott DF, Redman
BG, Kuzel TM, Harrison MR, Vaishampayan UN, Drabkin HA, George S,
Logan TF, et al: Nivolumab for metastatic renal cell carcinoma:
results of a randomized phase II trial. J Clin Oncol. 33:1430–1437.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Robert C, Ribas A, Wolchok JD, Hodi FS,
Hamid O, Kefford R, Weber JS, Joshua AM, Hwu WJ, Gangadhar TC, et
al: Anti-programmed-death-receptor-1 treatment with pembrolizumab
in ipilimumab-refractory advanced melanoma: A randomised
dose-comparison cohort of a phase 1 trial. Lancet. 384:1109–1117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gao Q, Wang XY, Qiu SJ, Yamato I, Sho M,
Nakajima Y, Zhou J, Li BZ, Shi YH, Xiao YS, et al: Overexpression
of PD-L1 significantly associates with tumor aggressiveness and
postoperative recurrence in human hepatocellular carcinoma. Clin
Cancer Res. 15:971–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Wang H, Wang X, Jin W, Tan Y, Fang
H, Chen S, Chen Z and Wang K: Genome-wide studies identify a novel
interplay between AML1 and AML1/ETO in t(8;21) acute myeloid
leukemia. Blood. 127:233–242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matys V, Kel-Margoulis OV, Fricke E,
Liebich I, Land S, Barre-Dirrie A, Reuter I, Chekmenev D, Krull M,
Hornischer K, et al: TRANSFAC and its module TRANSCompel:
Transcriptional gene regulation in eukaryotes. Nucleic Acids Res.
34:D108–D110. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin R, Maeda S, Liu C, Karin M and
Edgington TS: A large noncoding RNA is a marker for murine
hepatocellular carcinomas and a spectrum of human carcinomas.
Oncogene. 26:851–858. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sakamoto M: Early HCC: Diagnosis and
molecular markers. J Gastroenterol. 44:(Suppl 19). 108–111. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu X, Liu RF, Zhang X, Huang LY, Chen F,
Fei QL and Han ZG: DLK1 as a potential target against cancer
stem/progenitor cells of hepatocellular carcinoma. Mol Cancer Ther.
11:629–638. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi F, Shi M, Zeng Z, Qi RZ, Liu ZW, Zhang
JY, Yang YP, Tien P and Wang FS: PD-1 and PD-L1 upregulation
promotes CD8(+) T-cell apoptosis and postoperative recurrence in
hepatocellular carcinoma patients. Int J Cancer. 128:887–896. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pan X, Cang X, Dan S, Li J, Cheng J, Kang
B, Duan X, Shen B and Wang YJ: Site-specific disruption of the
Oct4/Sox2 protein interaction reveals coordinated mesendodermal
differentiation and the epithelial-mesenchymal transition. J Biol
Chem. 291:18353–18369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Scaffidi P and Bianchi ME: Spatially
precise DNA bending is an essential activity of the sox2
transcription factor. J Biol Chem. 276:47296–47302. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu C, Zhu Y, Jiang J, Zhao J, Zhang XG and
Xu N: Immunohistochemical localization of programmed death-1
ligand-1 (PD-L1) in gastric carcinoma and its clinical
significance. Acta Histochem. 108:19–24. 2006. View Article : Google Scholar : PubMed/NCBI
|