Introduction
Lung cancer is the leading cause of cancer-related
deaths worldwide, and non-small cell lung cancer (NSCLC)
constitutes the predominant histologic type, representing 80% of
all lung neoplasias (1). The vast
majority of patients are diagnosed at a locally advanced stage or
with distant metastasis (2). For
these unresectable patients, the standard treatment modality is two
cycles of chemotherapy with etoposide and cisplatin followed by
radiation. However, the prognosis for a large portion of patients
remains poor. The median survival time for patients treated with
chemoradiotherapy is approximately 15 months, and the 5-year
survival rate is only 5–17% (3).
Therefore, it is urgent to find novel and more effective
therapeutic targets with which to improve the prognosis of advanced
NSCLC patients.
Tripartite motif containing 28 (TRIM28) is an
important member of the tripartite motif superfamily that regulates
embryonic development (4),
chromatin organization, erythroblast differentiation (5), and the DNA damage response (6–9),
acting as an essential corepressor for KRAB zinc finger proteins.
Resent studies have shown that it also plays a role in
carcinogenesis and metastasis of cervical cancer (10), breast cancer (11), colorectal cancer (12), gastric cancer (13) and Kaposi's sarcoma (14). Our previous study also demonstrated
that TRIM28 promoted the proliferation and metastatic progression
of NSCLC and may be a new marker with which to predict metastasis
and prognosis in early stage NSCLC patients (15). Wang et al (16) reported that TRIM28 stimulated
formation of the E2F1-HDAC1 complex and inhibited E2F1 acetylation,
acting as a partial backup to prevent E2F1-mediated apoptosis in
the absence of retinoblastoma protein (pRb). Yet, the underlying
mechanism of TRIM28 in NSCLC remains unclear.
Etoposide is one of the alternative first-line
chemotherapeutic agents for NSCLC. Moreover, the sensitivity of
tumor cells to etoposide is correlated with the induction of E2F1.
Overexpression of E2F1 protein in tumor cell lines was found to
lead to enhanced cytotoxicity following exposure to DNA damaging
agent etoposide (17). Hence, in
this study, whether the combination of TRIM28 siRNA and etoposide
can augment apoptosis in vitro and in vivo was
investigated, since TRIM28 contributes to the negative regulation
of E2F1 which has been shown to be critical for etoposide-induced
apoptosis. In addition, the possible underlying mechanisms involved
in the tumorigenesis of NSCLC regulated by TRIM28 were
examined.
Materials and methods
Cell culture and RNA interference
assay
Human NSCLC cell line PAa (BioVector NTCC Inc.,
Shanghai, China) was grown in monolayer in Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10% FCS and was maintained
at 37°C in an atmosphere of humidified air with 5% CO2.
Complementary oligonucleotides containing short hairpin RNA (shRNA)
targeting TRIM28 was dimerized and cloned into the pLVTHM
lentiviral vector (GeneChem, Shanghai, China). The targeting
sequence for TRIM28 shRNA construct was: 5′-CACTGAGGACTACAACCTT-3,
sense and 5′-GCGATCTGGTTATGTGCAA-3, antisense. A
replication-incompetent lentivirus was produced by co-transfection
of the pLVTHM/TRIM28-shRNA expression vector and ViraPower
packaging mix containing an optimized mixture of two packaging
plasmids: psPAX2 and pMD2.G into 293T cells. PAa cells were
infected with lentiviral particles containing specific or negative
control vectors, and the polyclonal cells with strong GFP
expression were selected by flow cytometric sorting using anti-GFP
monoclonal antibody (Abcam, Cambridge, UK) to culture and
expand.
Ten micrograms of E2F1 siRNA expression vector and
negative control vector (GeneChem) were transfected using 30 ml of
Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) into PAa cells.
The target sequences of the synthetic oligonucleotides for RNAi
were as follows: siRNA-E2F1 sense, 5′-GACGUGUCAGGACCUUCGU-3′ and
antisense, 5′-ACGAAGGUCCUGACACGUC-3′. The transfected cells were
cultured for 48 h, and mRNA expression and protein level in the
cells were confirmed by RT-PCR and western blot analysis,
respectively.
Real-time quantitative PCR assay
After infection for 1, 2, 3 and 4 days, total RNA
was extracted from blank control, PAa/control-siRNA (si-control)
and PAa/TRIM28-siRNA cells using TRIzol reagent (Invitrogen)
according to the manufacturer's protocol. cDNA was synthesized from
2 µg of total RNA using a Reverse transcription kit (Clontech
Laboratories, Inc., Palo Alto, CA, USA). The PCR reaction included
the following components: 100 nmol/l of each primer, diluted cDNA
templates, and SYBR-Green PCR Master Mix (Biosystems, Warrington,
UK), to a total volume of 20 µl. The real-time quantitative PCR
assays were conducted using an ABI Prism 7000 Sequence Detection
System instrument (Biosystems Life Technologies, Foster City, CA,
USA) under the following conditions: 94°C for 4 min followed by 40
cycles at 94°C for 15 sec, 58°C for 30 sec and 72°C for 35 sec. All
reactions were performed in triplicate and GAPDH mRNA was used as
the internal control. The relative quantity of mRNA, normalized
against GAPDH mRNA, was expressed in terms of cycle threshold (Ct)
using the following equations: 2−(Ct TRIM28 - Ct GAPDH).
The primer sequences for TRIM28 gene were: sense,
5′-ATGTGAGCGTGTACTGCTGG-3′ and antisense,
5′-ACGTCTGCCTTGTCCTCAGT-3′ (Shanghai Shenggong Co., Ltd., Shanghai,
China).
Western blot analysis
Blank control, PAa/control-siRNA and
PAa/TRIM28-siRNA cells were lysed in ice-cold lysis buffer: 50
mmol/l Tris-HCl (pH 8.0), 150 mmol/l NaCl, 0.5% NP40, 0.5% sodium
deoxycholate, 0.1% SDS, plus protease inhibitor (Calbiochem,
Darmstadt, Germany). The proteins were separated by sodium dodecyl
sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to polyvinylidene difluoride membranes (Millipore,
Bedford, MA, USA). The membranes were blocked with 0.5% milk in
phosphate-buffered saline (PBS) for 1 h at room temperature. After
being washed in Tris-buffered saline Tween (TBST), the membranes
were incubated for 1 h at room temperature with an appropriate
dilution of rabbit polyclonal TRIM28 antibody (ProteinTech Inc.,
Chicago, IL, USA). After being washed in TBS-T, the blots were
incubated with horseradish peroxidase-conjugated goat anti-rabbit
IgG for 1 h and exposed to X-ray film at room temperature. The
signal was detected by chemiluminescence using ECL detection kit
(Amersham, Amersham, UK).
Cytotoxicity assay
To determine the IC50 value of etoposide,
the PAa cells were treated with various concentrations of etoposide
(0.001, 0.2, 0.5, 1, 2 and 4 µM). The cell group treated with only
1% DMSO (solvent of etoposide) was considered as the etoposide
blank control. Twenty-four hours later, the cytotoxicity of the
treatments was assessed using 3-(4, 5-dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT) cell proliferation kit (Roche
Diagnostics GmbH, Mannheim, Germany) according to the
manufacturer's protocol. Absorbance at 570 nm was measured using an
ELISA plate reader (Bio-Tek Instruments, Menlo Park, CA, USA). The
survival rate (SR) was determined using the following equation: SR
(%) = (A treatment/A control) × 100%. The concentration that
produced 50% cytotoxicity (IC50) was determined by
GraphPad Prism 6.02 software (GraphPad Software Inc., San Diego,
CA, USA).
The effect of TRIM28 siRNA on the chemosensitivity
of NSCLC cells was also determined using MTT assay. In 96-well
plates, the four cell groups: PAa/control-siRNA, PAa/TRIM28-siRNA,
PAa and PAa/TRIM28-siRNA, were divided into 500 cells/well in 100
µl. After 16 h, the third and the fourth cell groups were treated
with etoposide (IC50 dose) in a 96-well plate and
incubated at 1, 2, 3 and 4 days. After using the MTT cell
proliferation kit, the amount of formazan dye was determined by
quantifying its absorbance (A) at 570 nm.
Soft agar assay
PAa cells were divided into four groups. The first
and second cell groups were transfected with TRIM28-siRNA and
control-siRNA, respectively. Then, the cells (1×104)
were resuspended in DMEM containing 10% fetal bovine serum (FBS)
with 0.3% agarose and layered on top of 0.6% agar in medium
supplemented with 20% FBS on 60-mm plates. The plates were
incubated at 37°C in a humid atmosphere of 5% CO2. The
third group of PAa cells and the fourth cell group of cells
transfected with TRIM28-siRNA were incubated in DMEM containing
etoposide (IC50 dose). After 3 weeks, cell colony
numbers were counted under a microscope and cell colonies were
photographed at an original magnification of ×100. Only cell
colonies containing more than 50 cells were counted. The experiment
was performed for three independent times for each group.
Tumorigenicity in nude mice
This study was conducted in 40 female BALB/c nude
mice (Vital River, Beijing, China) at 4 weeks of age, which were
divided into four groups of 10 animals. The first and the second
group of mice were subcutaneously injected with PAa/control-siRNA
and PAa/TRIM28-siRNA cells, respectively. The third group of mice
received a single i.v. injection of etoposide solution at 80 mg/kg.
The i.v. injections were administered via a tail vein at a rate of
~0.05 ml/sec, using a 27-gauge needle and syringe. The fourth group
of mice were subcutaneously injected with PAa/TRIM28-siRNA cells
and then received an i.v. injection of etoposide solution. Mice
were observed daily for viability and changes in general health and
behavior. Body weights were determined once a week. Tumor onset was
scored visually and by palpitation at the sight of injection by two
trained laboratory staff at different times on the same day.
Average tumor size was estimated by physical measurement of the
excised tumor at the time of sacrifice. With the exception of mice
with large tumor burdens, animals were sacrificed 4 weeks after
injection. Protocols were conducted under the supervision of an
authorised investigator following approval of the Institutional
Ethics Committee of Chengde Medical College (Chengde, Hebei,
China).
Flow cytometric analysis
Cells were washed twice with PBS, and incubated with
5 µl Annexin V-Fluos and 5 µl PI solution (Roche Diagnostics GmbH)
in 1X binding buffer (10 mM HEPES, pH 7.4, 140 mM NaCl, 2.5 mM
CaCl2), at a final concentration of 1×106
cells/100 µl solution for 15 min. After incubation, the cells were
analyzed by flow cytometry. A minimum of 10,000 cells were analyzed
using a FACSAria II flow cytometer (BD Biosciences, San Jose, CA,
USA). Data were analyzed using FlowJo (Tree Star, Inc., Ashland,
OR, USA) flow cytometry analysis software.
TUNEL assay
Apoptosis was quantitatively analyzed by detection
of DNA fragmentation via a fluorescence assay based on terminal
deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling
(TUNEL) technique (Beyotime Institute of Biotechnology, Beijing,
China). This method takes advantage of DNA fragmentation,
characteristic of apoptosis. The DNA breaking points (nicks) expose
the 3 ′OH ends of DNA, which are labeled, thus allowing the
identification of apoptotic cells. Briefly, terminal
deoxynucleotidyl transferase was used to incorporate residues of
digoxigenin nucleotide into the 3 ′OH ends of DNA fragments.
Immunohistochemical procedures for detecting apoptotic cells were
performed according to the manufacturer's instructions. For each
slide, 5 fields were randomly chosen under a microscope for each
section. The apoptosis index (AI) was determined as: AI = number of
positively stained apoptotic cells/total number of cells counted ×
100. Assays were performed in a blinded manner.
Immunohistochemistry
Immunohistochemical analyses for TRIM28 and E2F1
were performed on formalin-fixed, paraffin-embedded surgical
sections obtained from the 4 mouse groups by using the
streptavidin-peroxidase method (ZhongShan Golden Bridge Biotech
Co., Ltd., Beijing, China). Slides were deparaffinized and
rehydrated with xylene and graded alcohol. Optimal antigen
retrieval was carried out in citrate buffer (pH 6.0) for 10 min
with a steam oven to enhance the immunoreactivity. Primary rabbit
polyclonal antibodies against TRIM28 and E2F1 (ProteinTech Inc.,
Chicago, IL, USA) was used at a dilution of 1:100.
Statistical analysis
Data are presented as mean ± standard deviation
(SD). Analysis of variance (ANOVA) followed by Bonferroni's test
was used to determine significant differences between groups.
Values of P<0.05 were considered significant. All statistical
analyses were performed using GraphPad Prism software.
Results
TRIM28 siRNA in combination with
etoposide inhibits the growth of NSCLC cell line PAa
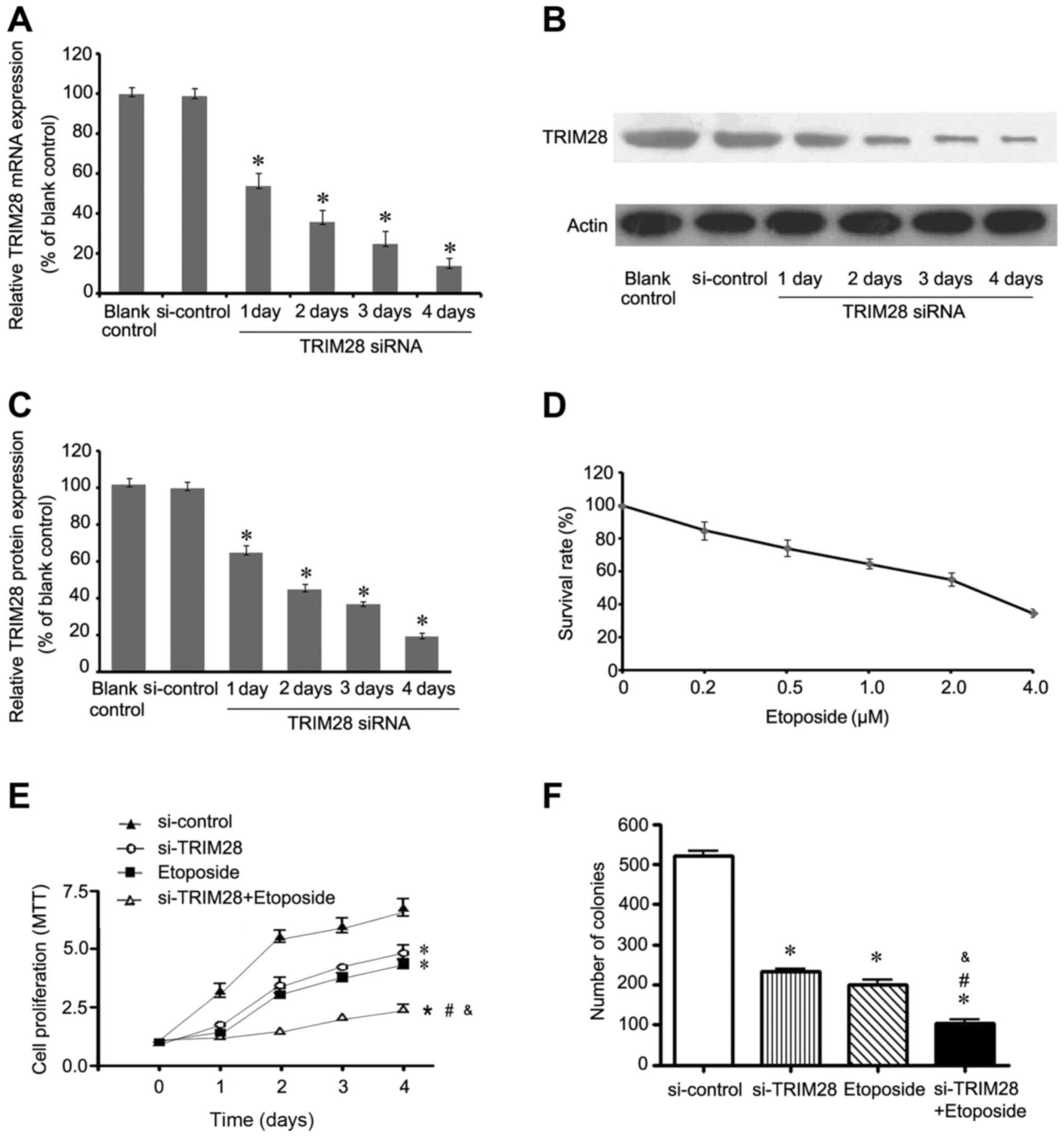
Our previous study indicated that the expression of
TRIM28 was significantly higher in 8 NSCLC cell lines in comparison
with bronchial epithelia BEAS2B cells (15). TRIM28-specific siRNA lentivirus
vector and a negative control vector (si-control) were designed and
constructed to silence TRIM28 in lung adenocarcinoma PAa cells,
which express higher TRIM28 than other NSCLC cell lines. Relative
TRIM28 gene expression was calculated in relation to the blank
control group, which was considered as 100%. Quantitative RT-PCR
analysis and western blot assay were used to confirm that TRIM28
siRNA resulted in a marked time-dependent reduction in both TRIM28
mRNA and protein levels (P<0.05, relative to the blank control).
At 1, 2, 3 and 4 days post-transfection, the relative expression of
TRIM28 mRNA was 53.67±6.51 (P=0.000), 36.00±6.08 (P=0.000),
24.67±6.43 (P=0.000) and 14.00±3.61% (P=0.000), respectively
(Fig. 1A), while the relative
TRIM28 protein expression levels were 64.67±4.16 (P=0.000),
45.00±3.00 (P=0.000), 36.67±1.53 (P=0.000) and 19.33%±2.08
(P=0.000), respectively (Fig. 1B and
C). Notably, treatment with siRNA control had no effect on
TRIM28 mRNA (P=0.829) and protein expression compared to the blank
control (P=0.291). To determine the IC50 dose of
etoposide, PAa cells treated with variable concentrations of
etoposide were analyzed using MTT assay. As shown in Fig. 1D, monotreatment with etoposide
induced cytotoxicity in a dose-dependent manner and the
IC50 value of etoposide was 2.30 µM.
To investigate whether the combination of TRIM28
siRNA and etoposide affects the proliferation of NSCLC cells, we
compared the proliferation of PAa cells treated with control siRNA,
TRIM28 siRNA, etoposide, and combination of TRIM28 siRNA and
etoposide using an MTT assay. As shown in Fig. 1E, PAa cells cotreated with TRIM28
siRNA and etoposide exhibited a significant growth inhibition when
compared with the cells treated with the other methods. Compared
with PAa/control-siRNA cells, the NSCLC cells treated with TRIM28
siRNA, etoposide and the combination of TRIM28 siRNA and etoposide
exhibited 33.78±1.67 (P=0.000), 37.84±2.84 (P=0.000) and
67.57±1.69% (P=0.000) decreased cell proliferation by day 4,
respectively. Furthermore, the combination group achieved a
statistically significant difference compared to the TRIM28 siRNA
group (P=0.000) or etoposide group (P=0.000).
The effect of growth inhibition was further examined
by a soft agar, a test for colony formation ability. As shown in
Fig. 1F, the ability of PAa cells
to grow in soft agar was significantly inhibited by TRIM28 siRNA,
etoposide, and cotreatment compared with control siRNA. In soft
agar, PAa cells treated with TRIM28 siRNA and etoposide formed
242±7 (P=0.000) and 221±19 colonies (P=0.000), respectively. The
cells treated with the combination of TRIM28 siRNA and etoposide
formed the least number of colonies (144±16) (P=0.000), while those
treated with control siRNA formed 520±15 colonies.
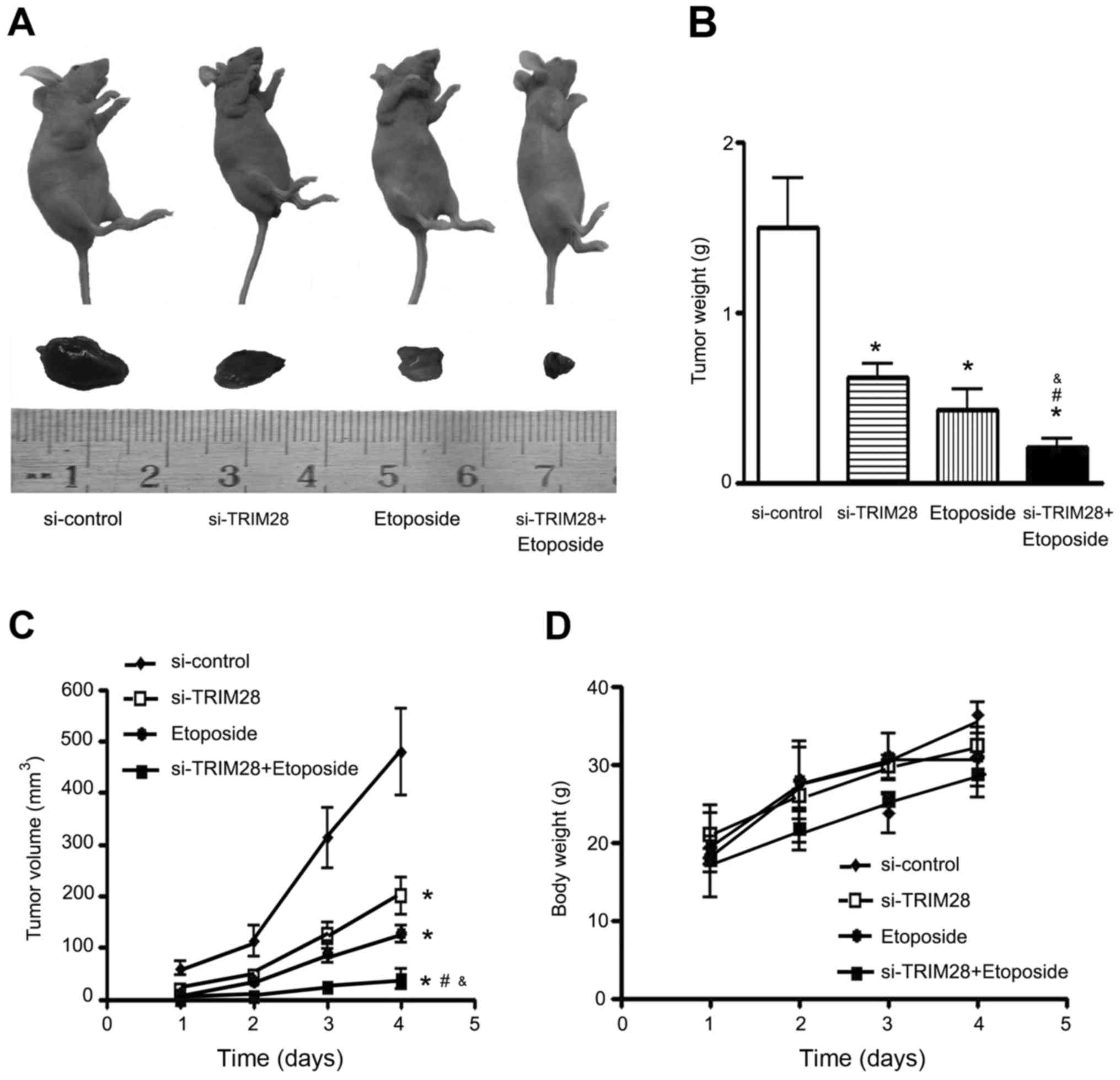
Antitumor effect of TRIM28 siRNA in
combination with etoposide on tumor growth in nude mice
Mice received an injection of PAa/TRIM28-siRNA cells
alone and in combination with 80 mg/kg etoposide for 4 weeks (5
times/week) and tumor growth was observed during the treatment
period. Growth inhibition was observed with TRIM28 siRNA alone or
etoposide alone, and this inhibition was statistically significant
when compared with that of the control (P=0.000). It appeared that
etoposide treatment was more effective than injection of
PAa/TRIM28-siRNA. Moreover, the combination of TRIM28 siRNA and
etoposide treatment was more effective than either method alone
(P=0.000; Fig. 2A-C). No loss of
body weight was observed in mice treated with PAa/TRIM28-siRNA
and/or etoposide during the experimental period (Fig. 2D).
TRIM28 siRNA in combination with
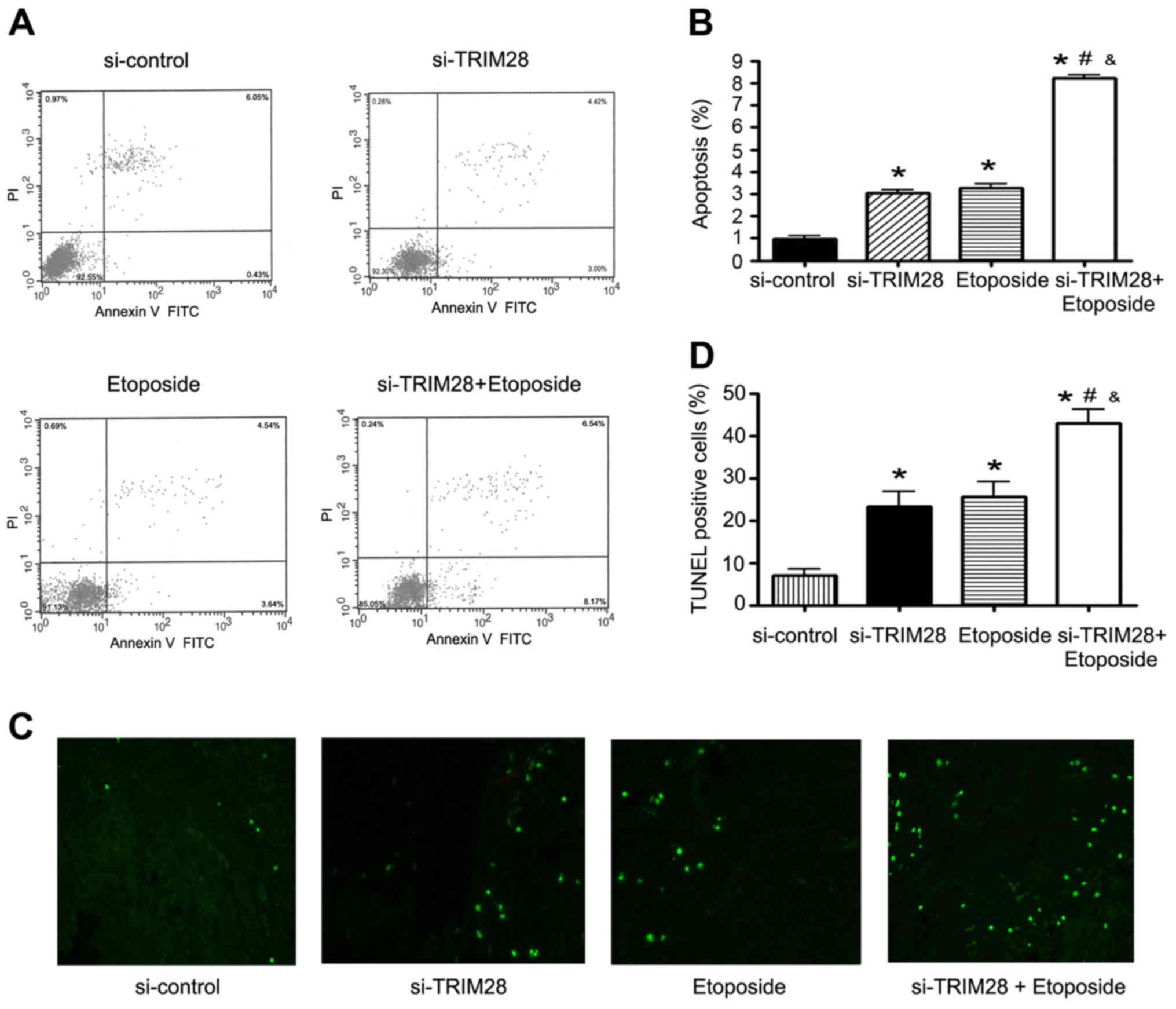
etoposide increases apoptosis in vitro and in vivo
After the NSCLC cell line PAa was cotreated with
TRIM28 siRNA and etoposide, we observed some abnormal changes. PAa
cells underwent a shrinkage in cell size and cells became round and
detached from the plates, which suggested that cell apoptosis had
occurred. To analyze the degree of apoptosis, flow cytometry was
performed. As shown in Fig. 3A and
B, apoptosis was observed in 0.97±0.27% of the PAa cells
transfected with control siRNA, 3.03±0.25% (P=0.001) of the PAa
cells transfected with TRIM28 siRNA and 3.26±0.31% (P=0.001) of the
cells treated with etoposide, while more apoptotic cells were
observed in the cells treated with the combination of TRIM28 siRNA
and etoposide (8.21±0.22%, P=0.000).
To further confirm whether the combination of TRIM28
siRNA and etoposide promoted tumor cell apoptosis, tumors were
removed from mice after treatment and the number of apoptotic cells
was quantified by TUNEL assay. The degree of apoptosis in the
tumors treated with either TRIM28 siRNA or etoposide alone was
significantly higher than that observed in the control group. The
largest number of apoptotic cells was observed in the tumors
cotreated with TRIM28 siRNA and etoposide. The combination
treatment significantly induced apoptosis compared to each
treatment alone as well as that noted in the control (Fig. 3C). The extent of apoptosis (the
percentage of total TUNEL-positive cells) in the groups were as
follows: control siRNA group, 8.20±2.11; TRIM28 siRNA group,
24.65±4.05 (P=0.004); etoposide group, 27.8±4.87 (P=0.004); TRIM28
siRNA plus etoposide group, 43.94±3.18 (P=0.000) (Fig. 3D).
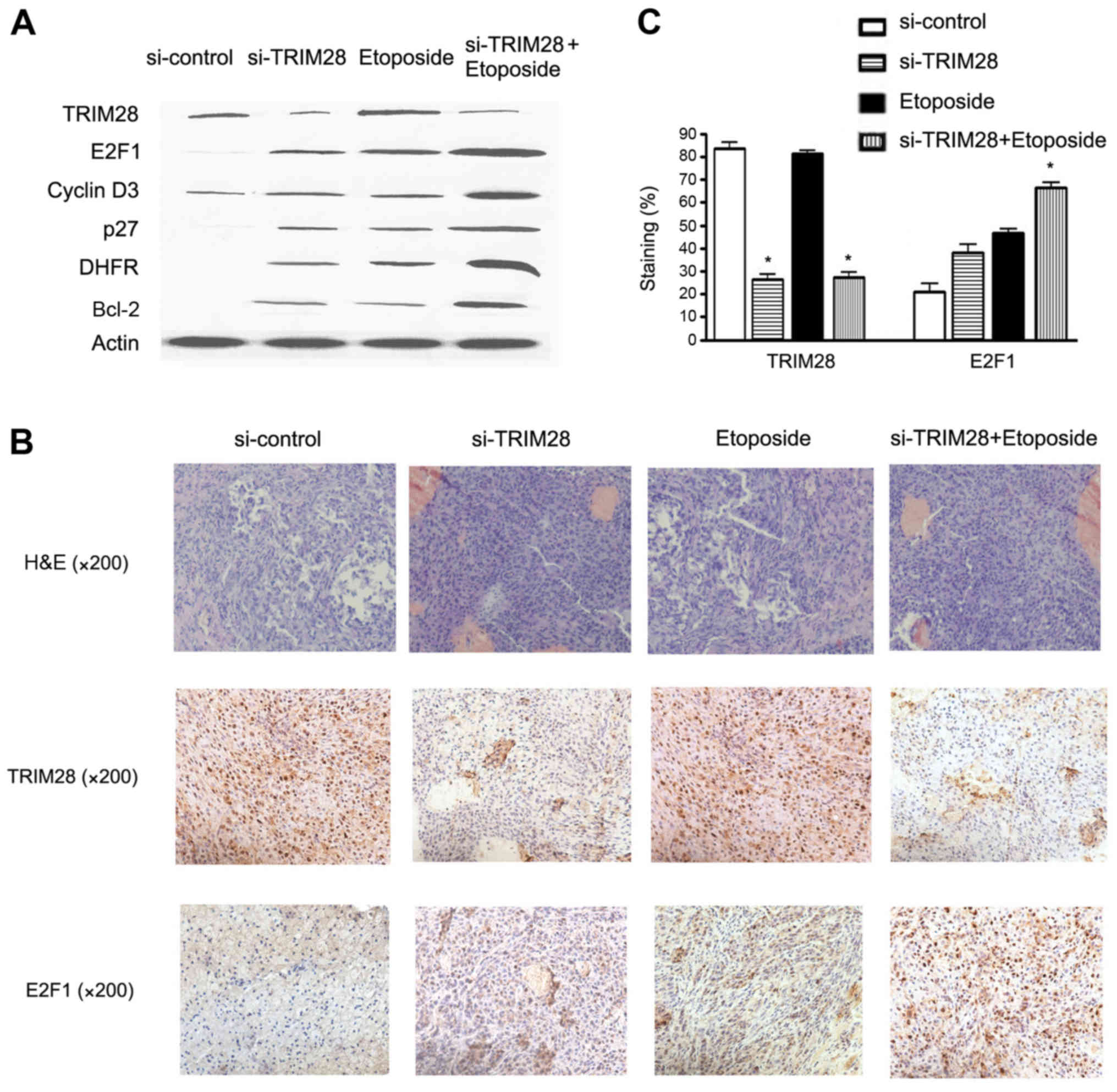
TRIM28 siRNA in combination with
etoposide increases the expression of E2F1
To test the hypothesis that E2F1 may play a role in
both TRIM28 siRNA and etoposide-induced cell death, the expression
of E2F1 protein and several E2F1 target genes was examined
following treatment of PAa with control siRNA, TRIM28 siRNA,
etoposide or the combination of TRIM28 siRNA and etoposide. As
shown in Fig. 4A, reduction in the
TRIM28 level or treatment with etoposide resulted in increased
expression of E2F1, cyclin D3, P27, dihydrofolate reductase (DHFR)
and Bcl-2. This combination treatment significantly induced the
expression of E2F1 and its target proteins, compared to each
treatment alone as well as to the control.
Tumors from mice were removed and processed for
further histologic and immunohistochemical analyses. We found that
E2F1 expression was upregulated in the TRIM28 siRNA and etoposide
treatment groups compared with that in the control siRNA group,
while the tumor cells in the cotreatment group had significantly
higher levels of E2F1 expression as compared with those in the
other treatment groups or the control group (Fig. 4B). As shown in Fig. 4C, the percentage of E2F1-positive
cells from 10 random fields in the combination group was increased
by 65.24±3.82% (P=0.003), compared to that in the control siRNA
group.
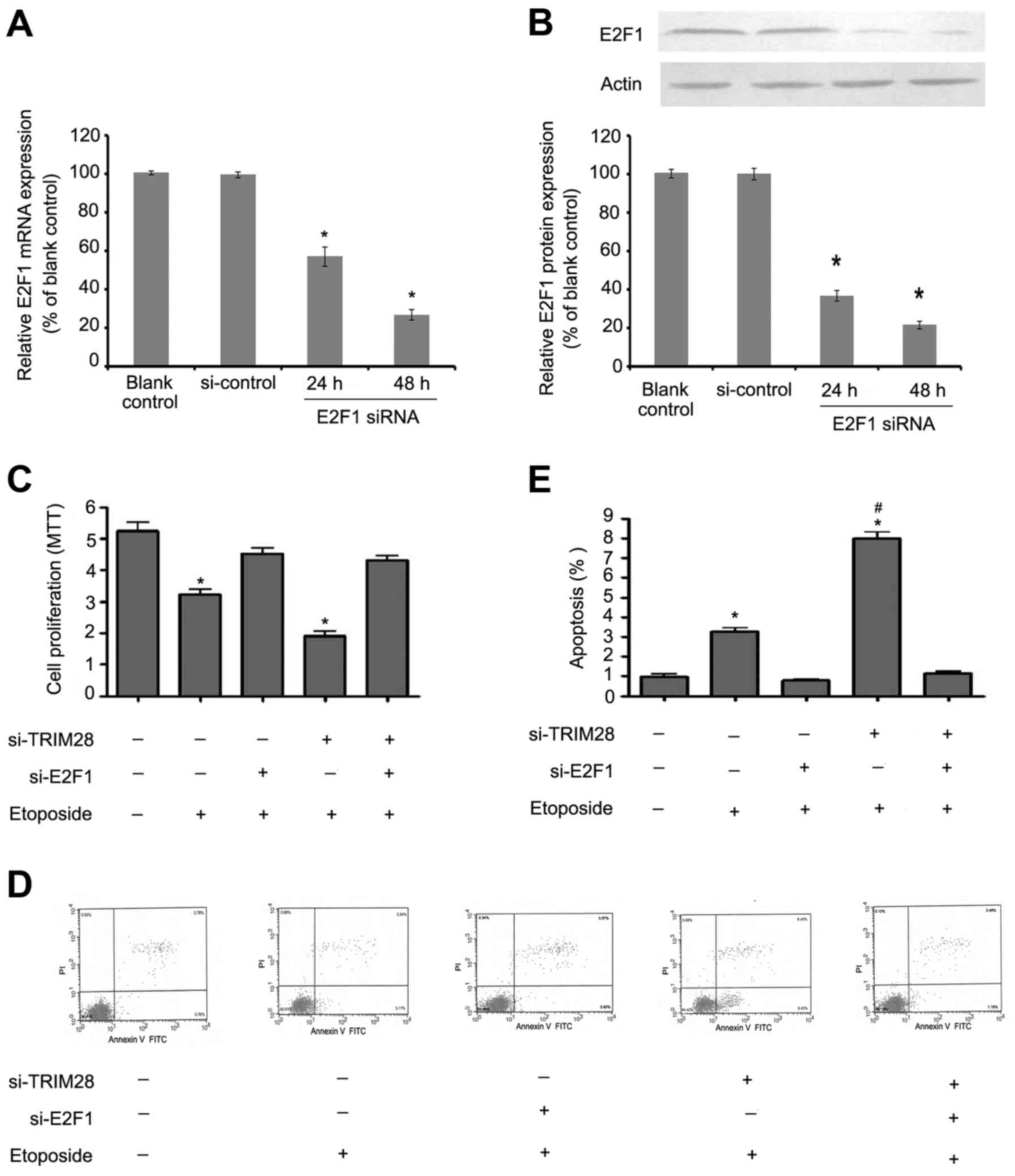
E2F1 mediates the sensitivity to
etoposide which is enhanced by TRIM28 siRNA
In order to determine whether the increased
sensitivity of PAa cells to etoposide after treatment with TRIM28
siRNA depends on E2F1, we examined the cell proliferation and
apoptosis in PAa cells transfected with TRIM28 siRNA alone or
together with E2F1 siRNA and then exposed to etoposide. In RNA
interference assay, >90% of the transfected cells expressed this
synthetic E2F1 siRNA, and endogenous expression of E2F1 was
effectively suppressed (Fig. 5A and
B, P<0.05). However, the inhibition of proliferation
(Fig. 5C) and induction of
apoptosis (Fig. 5D and E) were
almost completely abolished after E2F1 was knocked down in both the
TRIM28-silenced group as well as in the si-TRIM28 negative control
group which indicated that the promotion of sensitivity to
etoposide of TRIM28 siRNA is mediated by E2F1 activation.
Discussion
Resent studies indicate that TRIM28 is an important
regulator of carcinogenesis in several neoplasmas. Yokoe et
al (13) demonstrated that
TRIM28 provides a survival advantage to gastric cancer cells and is
an independent factor for peritoneal dissemination in gastric
cancer patients. An increase in TRIM28 expression which was induced
by loss of ZBRK1 enhanced the migration and invasion of cervical
cancer cells both in vitro and in vivo (10). Addison et al (11) showed that TRIM28 is overexpressed in
breast tumors and the depletion of TRIM28 in breast cancer cell
lines slowed cell proliferation and inhibited the growth and
metastasis of tumor xenografts. Multiple genes linked to tumor
progression and metastasis, including PTGS2/COX2, EREG, CD44, MMP1,
MMP2, were downregulated in breast cancer cells when TRIM28 was
silenced. Our previous study found that TRIM28 was upregulated in
NSCLC cell lines and tissues, promoting the growth and survival of
lung cancer cells. In this study, we further confirmed that stable
TRIM28 silencing significantly inhibited the growth of PAa cells in
DMEM and colony formation ability in soft agar. The proliferation
of PAa cells transfected with TRIM28 siRNA decreased by 33.78% when
compared to that noted in the PAa/control-siRNA cells after 4 days
by using an MTT assay. Our study also confirmed the antitumor
effect of TRIM28 siRNA in nude mice; the growth inhibition was
statistically significant compared with that of the controls.
Although a growing number of studies suggest that
TRIM28 plays a role in oncogenesis, the underlying mechanism of
TRIM28 remains unclear. Following DNA damage, TRIM28 is rapidly
phosphorylated and localized to sites of DNA strand breaks in the
nucleus (7). In addition,
HP1-dependent regulation of TRIM28 influenced DNA repair in
heterochromatin (18). TRIM28
stimulated formation of the p53-HDAC1 complex and inhibited p53
acetylation by interacting with MDM2. Depletion of endogenous
TRIM28 expression by RNAi stimulated p53 transcriptional activity,
sensitized p53 response to DNA damage, and increased apoptosis
(19). Kamitani et al
(20) found that TRIM28 associated
with endogenous STAT1 repressed the interferon (IFN)-mediated
signaling pathway. A siRNA-mediated reduction in TRIM28 expression
enhanced IFN-induced STAT1-dependent IRF-1 gene expression. A
recent study suggested that FIK acts as a bridging molecule to link
FOXP3 with TRIM28. Disruption of the FOXP3-FIK-TRIM28 complex in
Tregs restored expression of FOXP3-target genes and abrogated the
suppressor activity of Tregs (21).
These signaling pathways where TRIM28 plays a role were associated
with apoptosis in tumor genesis. The present study showed that
TRIM28 silencing increased apoptosis in vitro and in
vivo. We observed that cell apoptosis occurred after the NSCLC
cell line PAa was transfected with TRIM28 siRNA (apoptotic cells,
3.03±0.25%), significantly higher than the control group. The
degree of apoptosis in the tumors treated with TRIM28 siRNA
(24.65±4.05) was much higher than that in the control siRNA group
(8.20±2.11, P=0.004). Moreover, we observed that the expression of
E2F1 and several target genes was increased in the PAa cells
transfected with TRIM28 siRNA, and E2F1 expression was upregulated
in tumors from the TRIM28 siRNA treatment group mice. These results
suggest that TRIM28 regulates apoptosis induced by the E2F1
signaling pathway, in which TRIM28 binds the E2F1 transcription
factor in a PRb-independent fashion and inhibits E2F1 activity
(16). In our study, the E2F1
acetylation level increased, E2F1 transcription activity was
promoted, and apoptosis response was sensitized after the
endogenous TRIM28 was stably interfered.
Etoposide is a first-line cytotoxic drug for
advanced NSCLC, and belongs to the topoisomerase inhibitor drug
class. Etoposide formes a ternary complex with DNA and the
topoisomerase II enzyme (which aids in DNA unwinding), prevents
re-ligation of the DNA strands, and by doing so causes DNA strands
to break. Cancer cells rely on this enzyme more than healthy cells,
since they divide more rapidly. Therefore, this causes errors in
DNA synthesis and promotes apoptosis of the cancer cells. Numerous
studies have shown that E2F1 is critical for etoposide-induced
apoptosis. E2F1 protein levels were increased following treatment
with the DNA damaging agent etoposide. In contrast, overexpression
of E2F1 protein in tumor cell lines led to enhanced cytotoxicity of
etoposide, resulting from enhanced apoptosis (17). Nip et al (22) reported that E2F1 overexpression in
myeloid cells preferentially sensitized cells to apoptosis when
they were treated with etoposide, which was independent of p53
accumlation. Dong et al (23) used two human melanoma cell lines
treated with etoposide alone or in combination with adenoviral
vectors expressing E2F1 to evaluate the effect of E2F1 expression
on etoposide sensitivity. Cotreatment with Ad-E2F1 and low
concentrations of etoposide markedly sensitized melamoma cells to
apoptotic cell death. Topoisomerase II inhibitors also cooperated
with Ad-E2F1 to enhance antitumor activity in an in vivo
model using xenografts in nude mice. In the present study, we
observed that etoposide inhibited the growth and promoted the
apoptosis of NSCLC cells in vitro and in vivo, and
upregulated the expression of E2F1 and its target genes.
These results indicated that overexpression of E2F1
protein promoted apoptosis and enhanced the cytotoxicity of
etoposide. In addition, reduction of TRIM28 upregulated E2F1
transcription activity; thus we speculated that TRIM28 knockdown
would increase the sensitivity to etoposide in NSCLC cells. To
determine the hypothesis, we examined the antitumor effects of
TRIM28 siRNA in combination with etoposide in vitro and
in vivo. Cotreatment with TRIM28 siRNA and etoposide
inhibited the growth and proliferation more effectively in PAa
cells and nude mice. We further showed that the largest number of
apoptotic cells was observed in the PAa cells and mouse tumors
treated with the combination of TRIM28 siRNA and etoposide.
However, the inhibition of proliferation and induction of apoptosis
were almost completely abolished after E2F1 was knocked down in the
cells cotreated with TRIM28 siRNA and etoposide, which indicated
that the promotion of sensitivity to etoposide of TRIM28 siRNA is
mediated by E2F1 activation. A similar study was conducted by
Okamoto et al (24). The
authors examined the role of TRIM28 in p53 activation after the
treatment of cells with different types of chemotherapeutic agents.
TRIM28 reduction markedly enhanced the induction of p21, a product
of the p53 target gene, after treatment with actinomycin D or
γ-irradiation. Treatment with actinomycin D augmented the
interaction of p53 with Mdm2 and TRIM28. Furthermore, TRIM28
reduction in the actinomycin D-treated cells facilitated cell cycle
arrest and negatively affected clonal cell growth. Thus, the
reduction in TRIM28 levels promoted p53-dependent p21 induction and
inhibited cell proliferation in actinomycin D-treated cells. These
results suggest that TRIM28 recruits many proteins involved in gene
silencing and functions as an integral part of a co-repressor
complex. TRIM28 may serve as a therapeutic target against cancer in
combination with actinomycin D or etoposide.
Acknowledgements
This study was supported by the Hebei Natural
Science Foundation (H2015406014); the Hebei Talent Engineering
Training Funded Research Projects (A2016002085); the Heibei
University Science and Technology Research Project (QN20131010);
and the Hebei provincial focused discipline.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maghfoor I and Perry MC: Lung cancer. Ann
Saudi Med. 25:1–12. 2005.PubMed/NCBI
|
|
3
|
Hanna N, Neubauer M, Yiannoutsos C,
McGarry R, Arseneau J, Ansari R, Reynolds C, Govindan R, Melnyk A,
Fisher W, et al: Hoosier Oncology Group; US Oncology: Phase III
study of cisplatin, etoposide, and concurrent chest radiation with
or without consolidation docetaxel in patients with inoperable
stage III non-small-cell lung cancer: The Hoosier Oncology Group
and US Oncology. J Clin Oncol. 26:5755–5760. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Messerschmidt DM, de Vries W, Ito M,
Solter D, Ferguson-Smith A and Knowles BB: Trim28 is required for
epigenetic stability during mouse oocyte to embryo transition.
Science. 335:1499–1502. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barde I, Rauwel B, Marin-Florez RM,
Corsinotti A, Laurenti E, Verp S, Offner S, Marquis J, Kapopoulou
A, Vanicek J, et al: A KRAB/KAP1-miRNA cascade regulates
erythropoiesis through stage-specific control of mitophagy.
Science. 340:350–353. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pfeifer GP: Protein phosphatase PP4: Role
in dephosphorylation of KAP1 and DNA strand break repair. Cell
Cycle. 11:2590–2591. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
White DE, Negorev D, Peng H, Ivanov AV,
Maul GG and Rauscher FJ III: KAP1, a novel substrate for PIKK
family members, colocalizes with numerous damage response factors
at DNA lesions. Cancer Res. 66:11594–11599. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gilmore-Hebert M, Ramabhadran R and Stern
DF: Interactions of ErbB4 and Kap1 connect the growth factor and
DNA damage response pathways. Mol Cancer Res. 8:1388–1398. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu C, Zhang S, Gao X, Gao X, Xu X, Lv Y,
Zhang Y, Zhu Z, Zhang C, Li Q, et al: Roles of Kruppel-associated
box (KRAB)-associated co-repressor KAP1 Ser-473 phosphorylation in
DNA damage response. J Biol Chem. 287:18937–18952. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin LF, Li CF, Wang WJ, Yang WM, Wang DD,
Chang WC, Lee WH and Wang JM: Loss of ZBRK1 contributes to the
increase of KAP1 and promotes KAP1-mediated metastasis and invasion
in cervical cancer. PLoS One. 8:e730332013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Addison JB, Koontz C, Fugett JH, Creighton
CJ, Chen D, Farrugia MK, Padon RR, Voronkova MA, McLaughlin SL,
Livengood RH, et al: KAP1 promotes proliferation and metastatic
progression of breast cancer cells. Cancer Res. 75:344–355. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fitzgerald S, Sheehan KM, O'Grady A, Kenny
D, O'Kennedy R, Kay EW and Kijanka GS: Relationship between
epithelial and stromal TRIM28 expression predicts survival in
colorectal cancer patients. J Gastroenterol Hepatol. 28:967–974.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yokoe T, Toiyama Y, Okugawa Y, Tanaka K,
Ohi M, Inoue Y, Mohri Y, Miki C and Kusunoki M: KAP1 is associated
with peritoneal carcinomatosis in gastric cancer. Ann Surg Oncol.
17:821–828. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang L, Zhu C, Guo Y, Wei F, Lu J, Qin J,
Banerjee S, Wang J, Shang H, Verma SC, et al: Inhibition of KAP1
enhances hypoxia-induced Kaposi's sarcoma-associated herpesvirus
reactivation through RBP-Jκ. J Virol. 88:6873–6884. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu L, Zhao E, Li C, Huang L, Xiao L,
Cheng L, Huang X, Song Y and Xu D: TRIM28, a new molecular marker
predicting metastasis and survival in early-stage non-small cell
lung cancer. Cancer Epidemiol. 37:71–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang C, Rauscher FJ III, Cress WD and Chen
J: Regulation of E2F1 function by the nuclear corepressor KAP1. J
Biol Chem. 282:29902–29909. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng RD, Phillips P and El-Deiry WS:
p53-independent increase in E2F-1 expression enhances the cytotoxic
effects of etoposide and of adriamycin. Int J Oncol. 14:5–14.
1999.PubMed/NCBI
|
|
18
|
White D, Rafalska-Metcalf IU, Ivanov AV,
Corsinotti A, Peng H, Lee SC, Trono D, Janicki SM and Rauscher FJ
III: The ATM substrate KAP1 controls DNA repair in heterochromatin:
Regulation by HP1 proteins and serine 473/824 phosphorylation. Mol
Cancer Res. 10:401–414. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang C, Ivanov A, Chen L, Fredericks WJ,
Seto E, Rauscher FJ III and Chen J: MDM2 interaction with nuclear
corepressor KAP1 contributes to p53 inactivation. EMBO J.
24:3279–3290. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kamitani S, Ohbayashi N, Ikeda O, Togi S,
Muromoto R, Sekine Y, Ohta K, Ishiyama H and Matsuda T: KAP1
regulates type I interferon/STAT1-mediated IRF-1 gene expression.
Biochem Biophys Res Commun. 370:366–370. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang C, Martin S, Pfleger C, Du J,
Buckner JH, Bluestone JA, Riley JL and Ziegler SF: Cutting Edge: A
novel, human-specific interacting protein couples FOXP3 to a
chromatin-remodeling complex that contains KAP1/TRIM28. J Immunol.
190:4470–4473. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nip J, Strom DK, Fee BE, Zambetti G,
Cleveland JL and Hiebert SW: E2F-1 cooperates with topoisomerase II
inhibition and DNA damage to selectively augment p53-independent
apoptosis. Mol Cell Biol. 17:1049–1056. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong YB, Yang HL, Elliott MJ and McMasters
KM: Adenovirus-mediated E2F-1 gene transfer sensitizes melanoma
cells to apoptosis induced by topoisomerase II inhibitors. Cancer
Res. 62:1776–1783. 2002.PubMed/NCBI
|
|
24
|
Okamoto K, Kitabayashi I and Taya Y: KAP1
dictates p53 response induced by chemotherapeutic agents via Mdm2
interaction. Biochem Biophys Res Commun. 351:216–222. 2006.
View Article : Google Scholar : PubMed/NCBI
|