Introduction
Breast cancer (BC) is one of the leading cancers in
women accounting for more than one million new cases being
diagnosed and >400,000 deaths in the world every year (1,2).
Approximately 1.4 million women were diagnosed with BC in 2008
(3). In 2016, US Breast Cancer
Statistics estimated the diagnosis of 246,660 new cases of invasive
BC in women in the US (4). The
probability for a woman to be diagnosed with BC has increased at an
alarming rate from 1 to 11 (1975) to 1 to 8 (2013) (5). In 2010, 438,000 deaths were observed
globally due to BC (2). Tumor
metastasis is one of the primary reasons for its high mortality
rate. Despite the 5-year disease-free survival rate for patients
diagnosed with and treated for localized BC is an appreciable 89%;
the incidence of metastasis always results in poor prognosis
(6). Patients diagnosed with
distant metastasis have <2-year survival rate; while only 25% of
those patients survive longer than five years (7). Such critical statistics urge for
immediate action to limit metastasis and drastically improve
survivability of BC patients.
Triple negative breast cancer (TNBC) subtype of BC
have been defined as cells which lack the expression of estrogen
receptor (ER), progesterone receptor (PR) and human epidermal
growth factor receptor 2 (HER2), have an aggressive phenotype when
compared to other BC subtypes and comprises of 10–20% of BCs
(8,9). They are generally found in younger
women, have a high rate of metastasis, poor prognosis and most
importantly, lack clinically beneficial target molecules (10). Also, our previous study identified
that the occurrence of TNBC is especially high in the
African-American and Hispanic women residing in the South Los
Angeles area (11). Treatment
options for women with TNBC are very limited. While Trastuzumab and
endocrine therapy indicate negative beneficiary effects, cytotoxic
chemotherapy and molecular-targeted therapies are a few that have
demonstrated affirmative efficacy towards TNBC metastasis treatment
(12–14). Thus, it is imperative to identify
specific unique markers overexpressed in TNBC that may assist in
inhibiting metastasis and also be used as a potential targeted
therapy for treatment of women diagnosed with TNBC subtype of
breast cancer.
Recent studies have demonstrated that chemokines
regulate paracrine survival networks that could be beneficial for
cancer cells to thrive, metastasize and cause chemoresistance
(6,15–17).
GROα/CXCL1 (C-X-C motif ligand 1) is a chemotactic cytokine
composed of small peptides that act as a chemoattractant for
leukocytes during homeostasis and inflammatory responses (18,19).
GROα specifically binds to a seven-transmembrane G protein-coupled
CXCR2 chemokine receptor to trigger downstream NF-κB signaling
pathway and PI-3K pathway (20–25).
Reports also illustrate that GROα plays a vital role in tumor cell
transformation, angiogenesis and metastasis in various forms of
cancer (15,18,26–29). A
study has identified numerous inflammation-related cytokines to be
differentially regulated in TNBC cells (30).
Here, we report our findings on the importance of
GROα expression in TNBC metastasis. We investigated the GROα
expression levels in different BC subtypes along with a panel of
additional TNBC cell lines and performed functional and mechanism
studies on high-GROα expressing TNBC cell lines versus low-GROα
expressing BC cell line in vitro. GROα stimulation or
inhibition demonstrated significant differences in migration and
invasion abilities indicating the role of GROα in BC metastasis.
Also, these results were substantiated by differential alterations
in the expression of EMT markers. In this study we also identify
that GROα regulates BC migration/invasion via MAPK pathway and that
treatment with MAPK inhibitor PD98059 significantly reverses all
the effects induced by GROα on BC cells.
Materials and methods
Cell cultures
MDA-MB-231, MCF-7, SKBR3 and BT474 cells were
purchased from ATCC (American Type Culture Collection) and grown in
DMEM/F12 medium containing 10% FBS, 1% glutamine and 0.5%
antibiotics (penicillin/streptomycin) in 5% CO2
incubator at 37°C. Triple-Negative Breast Cancer Panel 3 (TCP 1003)
cell lines mentioned below were purchased from ATCC and cultured as
per the Company's instructions. BT20 and HCC2157 were grown in
DMEM/F12 medium containing 10% FBS, 1% glutamine, 0.5% antibiotics
in 5% CO2 incubator at 37°C; HCC2157 was additionally
supplemented with 0.02 mg/ml insulin, 0.01 mg/ml transferrin, 25 nM
sodium selenite, 50 nM hydrocortisone, 1 ng/ml EGF, 0.01 mM
ethanolamine, 0.01 mM O-phosphorylethanolamine, 100 pM
3,3′,5-triiodo-L-thyronine (T3), 0.5% bovine serum albumin, and 0.5
mM sodium pyruvate. DU4475, HCC1187, HCC1937, HCC1143, HCC1599,
HCC1806, HCC1395, HCC38 and HCC70 were cultured in RPMI-1640 medium
with 10% FBS and 1% glutamine in 5% CO2 incubator at
37°C. BT-549 and HS578T were cultured in DMEM/F12 medium with 10%
FBS, 1% glutamine and 0.01 mg/ml Insulin in 5% CO2
incubator at 37°C. MDA-MB-157, MDA-MB-453, MDA-MB-468, MDA-MB-436
were grown in Leibovitz's L-15 medium with 10% FBS, 1% glutamine
and 0.5% antibiotics in a free gas exchange incubator with
atmospheric air; MDA-MB-436 was additionally supplemented with 16
µg/ml glutathione.
Materials
Materials were purchased from various manufacturers:
recombinant GROα (R&D Systems), GROα siRNA (Santa Cruz
Biotechnology), control siRNA (Santa Cruz Biotechnology),
Lipofectamine ltx (Invitrogen), Opti-MEM Reduced Serum medium
(Invitrogen), GROα ELISA kit (R&D Systems), GROα primers (IDT),
ThermoScript™ RT-PCR systems (Invitrogen), SYBR Green (Qiagen), EMT
marker primers (IDT), PD98059 (Cell Signaling), E-cadherin antibody
(Santa Cruz Biotechnology), N-cadherin antibody (Santa Cruz
Biotechnology), Snail antibody (Abcam), Slug antibody (Abcam),
Twist antibody (Santa Cruz Biotechnology), Vimentin antibody (Santa
Cruz Biotechnology), VEGF antibody (Santa Cruz Biotechnology),
phospho-MAPK antibody (Cell Signaling), MAPK antibody (Cell
Signaling), β-actin mouse antibody (Santa Cruz Biotechnology).
Enzyme-linked immunosorbent assay
(ELISA)
Medium containing 10% FBS was used for performing
all ELISA experiments since preliminary studies to analyze baseline
GROα concentration in the complete culture medium used by all the
cell lines in this study by ELISA demonstrated no presence of GROα
(data not shown). ELISA was performed by following the
manufacturer's instructions. Briefly, cells were plated in the
6-well plate until reaching 70–80% confluency. The medium was then
replaced with fresh 10% FBS containing medium for 24 h. The culture
medium was collected and spun momentarily to remove all
particulates; the cells were detached and counted and later used
for protein extraction. Cell culture supernatants/lysates were then
immediately assayed for GROα expression levels. Briefly, standards
or samples were added to each well of the GROα specific ELISA strip
and incubated for 1.5 h at room temperature (RT). The wells were
washed thrice with wash buffer, and re-incubated with GROα
conjugate for 1 h at 4°C. The wells were washed again and incubated
with the substrate solution, protected from light for 15 min at RT.
The reaction or the color production was ceased by adding stop
solution which turned the solution yellow. The absorbance was read
colorimetrically (Promega Glomax Multidetection System) at 450 nm.
Standard curve was generated to calibrate the concentration of GROα
in each supernatant sample and expressed as concentration of GROα
(pg/ml) per µg of the protein. The samples were run in duplicates
and the experiment repeated thrice for consistent results.
RNA isolation
Cells were washed with cold 1X PBS before lysing
them with RNA Bee (Tel-Test) for ~15 min at RT. To the cell lysate,
chloroform was added, shaken vigorously and incubated for 15 min on
ice. The samples were then centrifuged at 13,000 rpm for 15 min at
4°C resulting in two phases, lower blue chloroform-phenol phase and
the color-less upper aqueous phase. The aqueous phase was then
transferred to a new tube adding equal volume of isopropanol. The
samples were stored on ice for 30 min and then centrifuged at
13,000 rpm for 15 min at 4°C to form a white-yellow pellet of
precipitated RNA. The RNA pellet was then washed in 75% ethanol for
10 min at 7,500 rpm at 4°C. The isolated RNA was air-dried and
dissolved in DEPC treated water. The concentration and purity of
the isolated RNA was calibrated spectrophotometrically using
NanoDrop (Thermo Fisher Scientific 2000C).
Quantitative PCR (Q-PCR)
RNA (2 µg) from each sample was used to generate
cDNAs using ThermoScript RT-PCR systems kit (Invitrogen). Briefly,
RNA along with the first master mix (oligonucleotide, random
hexamerase, 10 mM dnTPs) was incubated at 65°C for 5 min. Second
master mix (5X buffer, DTT, DEPC water, RNAse Out, Thermoscript)
was then added to the solution and the samples were loaded on to
the mastercycler (Eppendorf) for converting mRNA to cDNA. The cDNAs
were then treated with RNAse H and incubated at 37°C for 15 min.
cDNA (1 µl) from each sample was later used along with SYBR Green
in a thermocycler (Bio-Rad) to perform Q-PCR. The expression levels
of a gene of interest was tested by using specific primers along
with an endogenous control 18S by subjecting the cDNA to 58°C
primer annealing and 72°C of elongation process for 40 cycles. The
Ct values were noted, normalized with an endogenous
control (18S) and the results were analyzed by the
2−∆∆Ct method (31).
Experiments were performed in triplicates and repeated at least
twice.
Transfections
MDA-MB-231 and HCC1937 cells (2×106) were
transfected with GROα siRNA using Lipofectamine LTX (Invitrogen) in
Opti-MEM (Invitrogen) reduced serum medium following the
manufacturer's instructions for 4 h. Scrambled siRNA served as the
control. The cells were then incubated at 37°C in a CO2
incubator. Transfected cells were then used for functional
studies.
Cell proliferation assay
Cells/well (1×105) of MDA-MB-231 and
HCC1937 or serum-starved MCF7 and SKBR3 either transfected with
GROα siRNA or induced with recombinant GROα respectively, along
with their respective controls were placed in 96-well plates. In
another experiment, HCC1937 cells were subjected to increasing
concentrations of Olaparib (PARP inhibitor, - 0, 50, 100 and 200
µM) to determine the IC50 value for the drug. After 72 h
of treatment, cell proliferation rates were determined using MTT
assay (Sigma) as per the manufacturer's protocol. Briefly, the
plates were flipped to empty out the media and 50 µl of MTT buffer
reagent was added to each well. The setup was incubated for 4 h at
37°C wrapped in an aluminum foil. After incubation, the MTT reagent
was replaced with 100 µl of DMSO and incubated for 15 min at RT on
a rocker. The absorbance was read at 560 nm. Experiments were
performed in triplicates and repeated thrice. The results are
presented as percentage for cell proliferation.
Scratch assay
MDA-MB-231 and HCC1937 cells along with
serum-starved MCF7 and SKBR3 cells were grown up to absolute
confluency in a culture-insert (Ibidi) with 500 µM cell-free gap
overnight. At 0 hour, the insert was removed and the cell patches
were rinsed with 1X PBS to take away all the detached cells. It was
then treated with different conditions for 24 h. Images were
captured at 0 and 24 h after treatment to mark the changes in
migration of phenotypically modified BC cells. The distance
remaining in the gap after 24 h was captured again. The experiment
was performed in triplicates and repeated at least twice.
Cell invasion assay
The invasion assay was performed in 24-well Boyden
chambers comprising of inserts with 8-µM pore membranes which were
coated with Matrigel (28 µg/insert; Sigma). Briefly, equal no. of
phenotypically modified BC cells in reduced medium along with
control (1×105/well) were placed in the upper wells and
10% FBS containing DMEM/F12 culture medium was filled in the lower
wells to act as a chemo-attractant. The cells were cultured for 24
h and then fixed with 0.5% glutaraldehyde and stained with 0.5%
toluidine blue. The number of invaded cells was counted with 20X
objective of a microscope from 3 different fields per membrane. The
results are presented as percentage for cell invasion. Each
experiment was performed at least twice.
Western blotting
Total protein was extracted using standard RIPA
buffer (Thermo Fisher Scientific) with proteinase inhibitors
(Thermo Fisher Scientific) and the concentration was measured by
BCA protein assay (Thermo Fisher Scientific). Protein (30 µg) from
each sample was loaded on to the well and separated using NuPAGE
pre-cast gels (Invitrogen). The proteins were then transferred onto
the nitrocellulose membrane (0.45 µM, Bio-Rad). The membrane was
initially blocked using 5% milk blocking buffer, washed with 1X
TBST and later incubated with specific primary antibody overnight
at 4°C. The membrane was washed again and then incubated with
secondary antibody for 2 h at RT. The results were visualized using
SuperSignal® West Pico Chemiluminescence Substrate
(Thermo Fisher Scientific) (Bio-Rad ChemiDoc). β-actin was used as
the loading control. The densitometric analysis was performed by
calculating the volume density ratio of bands of genes of interest
over loading control. Experiments were repeated at least twice for
consistent results.
Statistical analysis
All the experiments were performed at least in 2
separate sets of experiments with 3 technical replicates in each.
Data are presented as mean values ± standard deviation. Differences
between 2 groups were analyzed using Student's t-test. p<0.05
was considered statistically significant.
Results
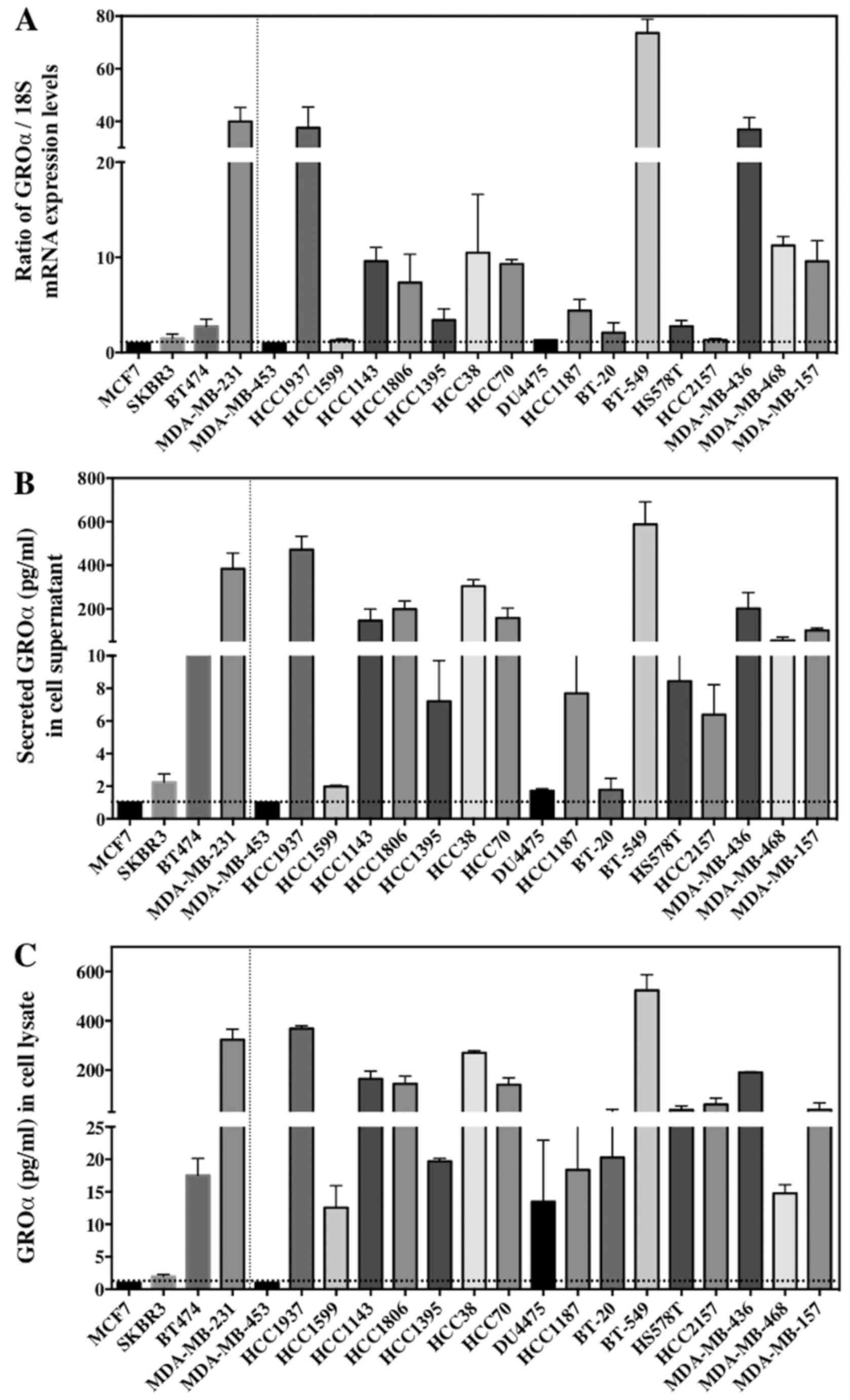
GROα is overexpressed in TNBC
cells
BC comprises of different subtypes. Four BC cell
lines were chosen for this study to represent each subtype - SKBR3
(ER−/PR−/HER2+, HER2 subtype),
MCF7 (ER+/PR+/HER2−, luminal A
subtype), BT474 (ER+/PR+/HER2+,
luminal B subtype), and MDA-MB-231
(ER−/PR−/HER2−, basal/TNBC
subtype), and cultured for further analysis. Our first approach was
to investigate the baseline GROα expression in each subtype.
Results from q-PCR and ELISA demonstrated that GROα is
significantly overexpressed in the MDA-MB-231 cells with TNBC
subtype when compared to other BC cell lines/subtypes. The GROα
mRNA levels from q-PCR and protein concentration levels from ELISA
were analyzed. Interestingly, expression levels of GROα
specifically in MDA-MB-231 cells both in mRNA (30.3, expressed as
ratio of GROα/18S) and protein concentration levels (383.92 pg/ml
in supernatant) were significantly upregulated when compared to
other cell lines expressing <2 and 30 pg/ml (in supernatant) of
mRNA and protein respectively (Fig.
1A). Thus, indicating differential GROα expression in BC
subtypes.
To further confirm our observation that GROα is
overexpressed specifically in TNBC cells which might suggest the
aggressive nature of TNBCs, we examined additional 17 TNBC cell
lines (BT20, HCC2157, DU475, HCC1187, HCC1937, HCC1143, HCC1599,
HCC1806, HCC1395, HCC38, HCC70, BT-549, HS 578T, MDA-MB-157,
MDA-MB-453, MDA-MB-468 and MDA-MB-436), for baseline GROα
expression in these cells as a proof-of-principle. All the cell
lines were cultured as per the ATCC guidelines. Cells were
harvested for total RNA and supernatants were collected for
secreted protein analysis by q-PCR and ELISA respectively.
Remarkably, >50% of the TNBC cell lines (HCC1937, HCC1143,
HCC1806, HCC38, HCC70, BT-549, MDA-MB-436 and MDA-MB-468) expressed
significantly elevated levels of GROα mRNA and protein while other
cell lines demonstrated lower GROα expression levels (Fig. 1). Cell lines expressing higher
levels of GROα such as HCC1937, BT549 and MDA-MB-436 are metastatic
in nature and have an aggressive phenotype. Hence, from these
results we hypothesized that GROα is an important molecule
associated with TNBC metastasis and that upregulation of GROα
expression aids BC cell migration and invasion.
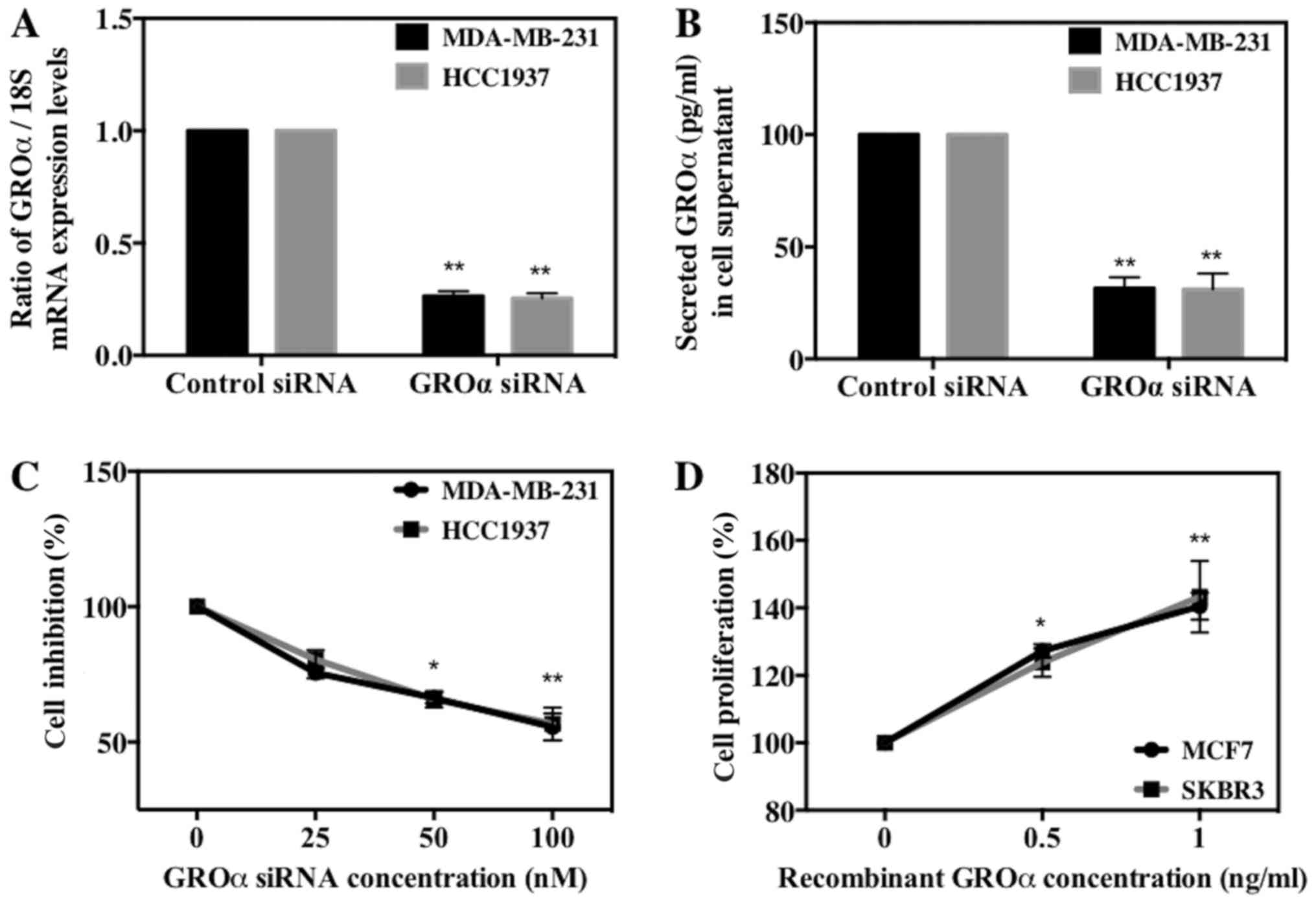
GROα effects on BC cell
proliferation
We next investigated the role of GROα in BC cell
proliferation in vitro by either knocking down or
stimulating BC cells with GROα. To achieve this we used four cell
lines, MDA-MB-231 and HCC1937 (high-GROα) and MCF7 and SKBR3
(low-GROα). We then transfected MDA-MB-231 and HCC1937 cells with
GROα specific siRNA (100 nM) using scrambled siRNA as control and
induced MCF7 and SKBR3 cells with recombinant GROα (1 ng/ml) using
water as control for 72 h. The effect of GROα knockdown using GROα
specific siRNA on respective cell lines was tested both by qPCR and
ELISA (Fig. 2A and B). After 72 h,
the treated cells were subjected to MTT assay to assess the effect
of GROα on BC cell proliferation. Results obtained demonstrated a
gradual decrease (~35–40%) in cell proliferation in GROα-knocked
down MDA-MB-231 and HCC1937 cells when compared to control cells
(without siRNA treatment) (Fig.
2C). Similarly, a gradual increase (38–42%) in cell
proliferation was observed in GROα stimulated MCF7 and SKBR3 cells
when compared to control cells (untreated with GROα) (Fig. 2D). These results indicate that GROα
induces positive effects on BC cell proliferation over 72-h time
period.
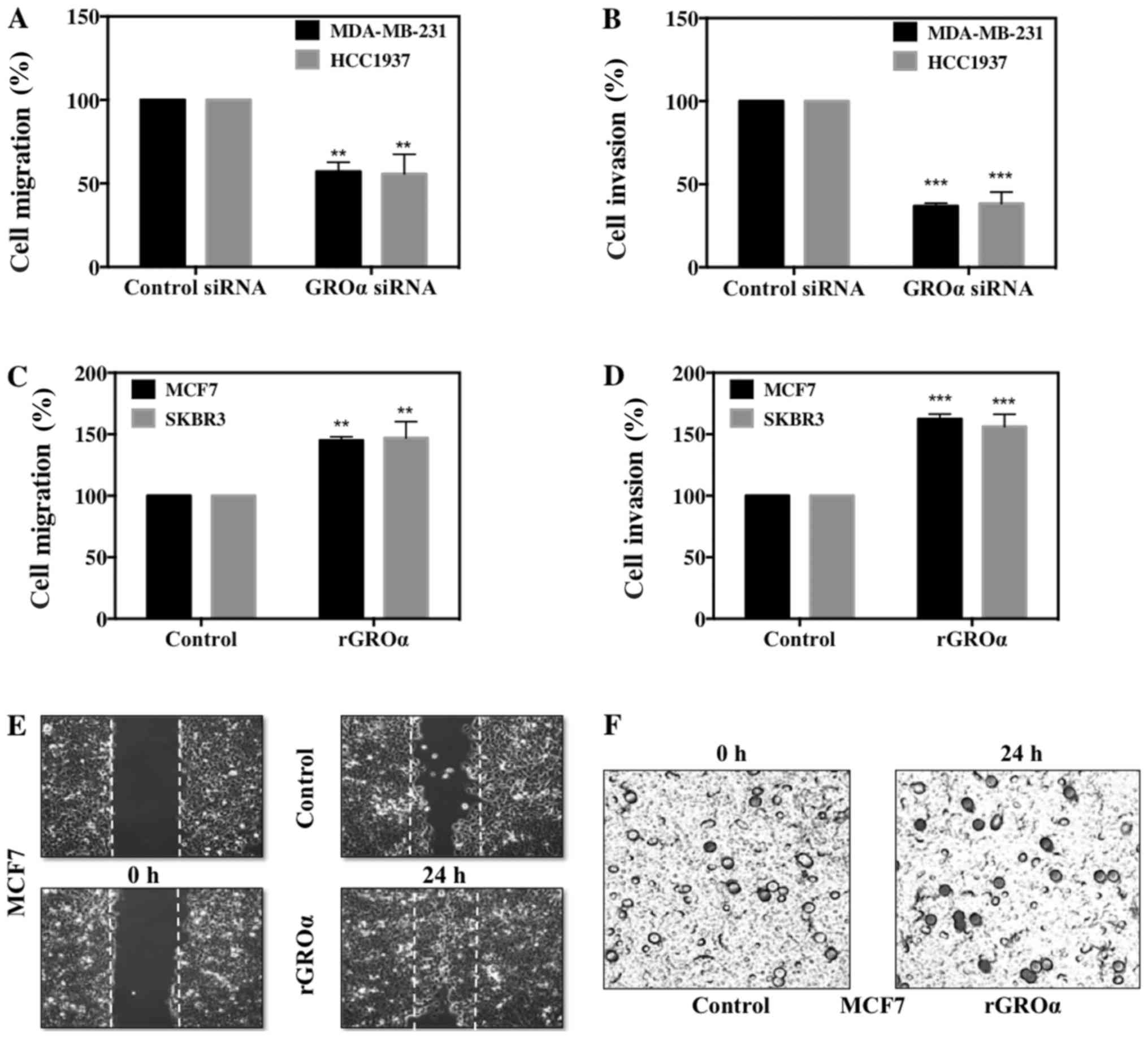
GROα promotes BC cell migration and
invasion
We further evaluated the significance of GROα on BC
cell migration and invasion in vitro. For this, we used the
phenotypically modified GROα-knocked down MDA-MB-231 and HCC1937
cells and GROα-stimulated MCF7 and SKBR3 cells along with their
respective untreated controls and subjected them to scratch
assay/wound healing assay and Boyden chamber Matrigel invasion
assay. Images were captured at 0 and 24 h for migration and at 24 h
for invasion assays to mark the changes in treatment conditions.
Results from both migration and invasion assay demonstrated
complementary data; while GROα-knocked down MDA-MB-231 and HCC1937
cells displayed significant decrease after 24 h with as low as
43–47% reduced migratory and 59–60% reduced invasive abilities
(Fig. 3A and B). In contrast,
GROα-stimulated MCF7 and SKBR3 cells showed increase in both
migration and invasion, with as high as 45–47% enhanced migratory
and 56–69% invasive abilities with reference to their untreated
controls (Fig. 3C and D).
Representative images of migration and invasion assays in MCF7
cells with GROα-stimulation are shown in Fig. 3E and F. Results from the in
vitro functional studies clearly demonstrate that GROα plays a
crucial modulatory role in BC cell migration and invasion;
suggesting that GROα could be an important target molecule in the
treatment for TNBC metastasis.
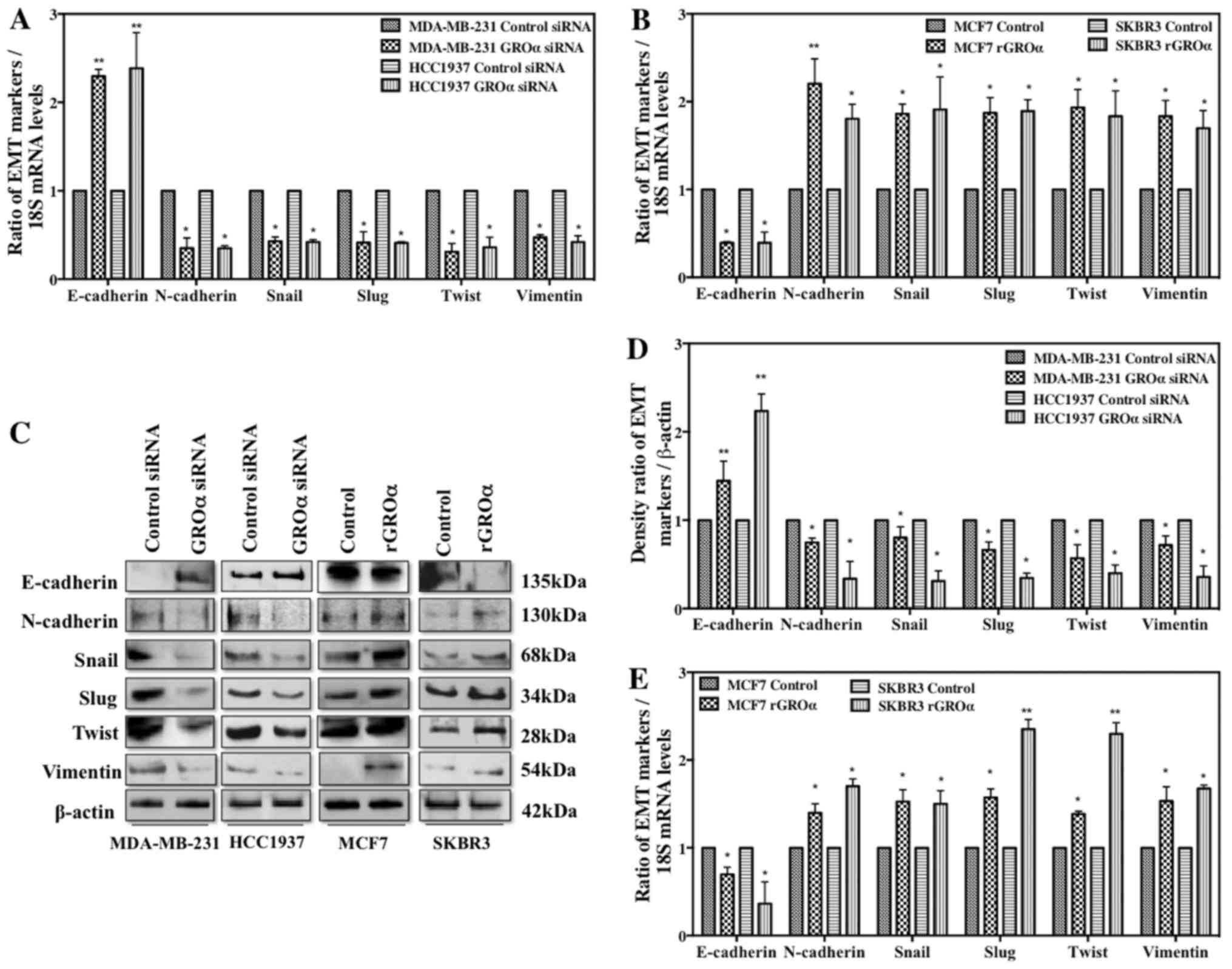
GROα stimulation/knockdown induces
phenotypic changes in EMT markers
Thus far, our studies imply that GROα is a critical
modulator for BC cell metastasis. To further understand the
molecular mechanisms that regulate the phenotypically modified
MDA-MB 231/HCC1937 and MCF7/SKBR3 cells to inhibit/initiate
metastasis process, we evaluated the change in expression of
various EMT markers in the presence or absence of GROα in BC cells
by q-PCR and western blotting after 48-h treatment. Interestingly,
the results from both q-PCR (Fig. 4A
and B) and western blotting (Fig.
4C) demonstrated that in the presence of GROα (GROα induced
MCF7/SKBR3 cells), a significant downregulation of E-cadherin was
accompanied by strong upregulation of all the other mesenchymal
markers such as N-cadherin, Snail, Slug, Twist and Vimentin. In
contrast, in the absence of GROα (GROα-knocked down
MDA-MB-231/HCC1937 cells), an elevated expression of E-cadherin was
accompanied with decrease in mesenchymal markers. Densitometric
analysis of the bands obtained from western blotting method clearly
indicated loss of E-cadherin and gain of other EMT markers in
GROα-knocked down cells versus the contrary effects in
GROα-stimulated cells (Fig. 4D and
E). These results further emphasize the role and the molecular
mechanisms regulated by GROα to influence and trigger metastasis
process in TNBC cells.
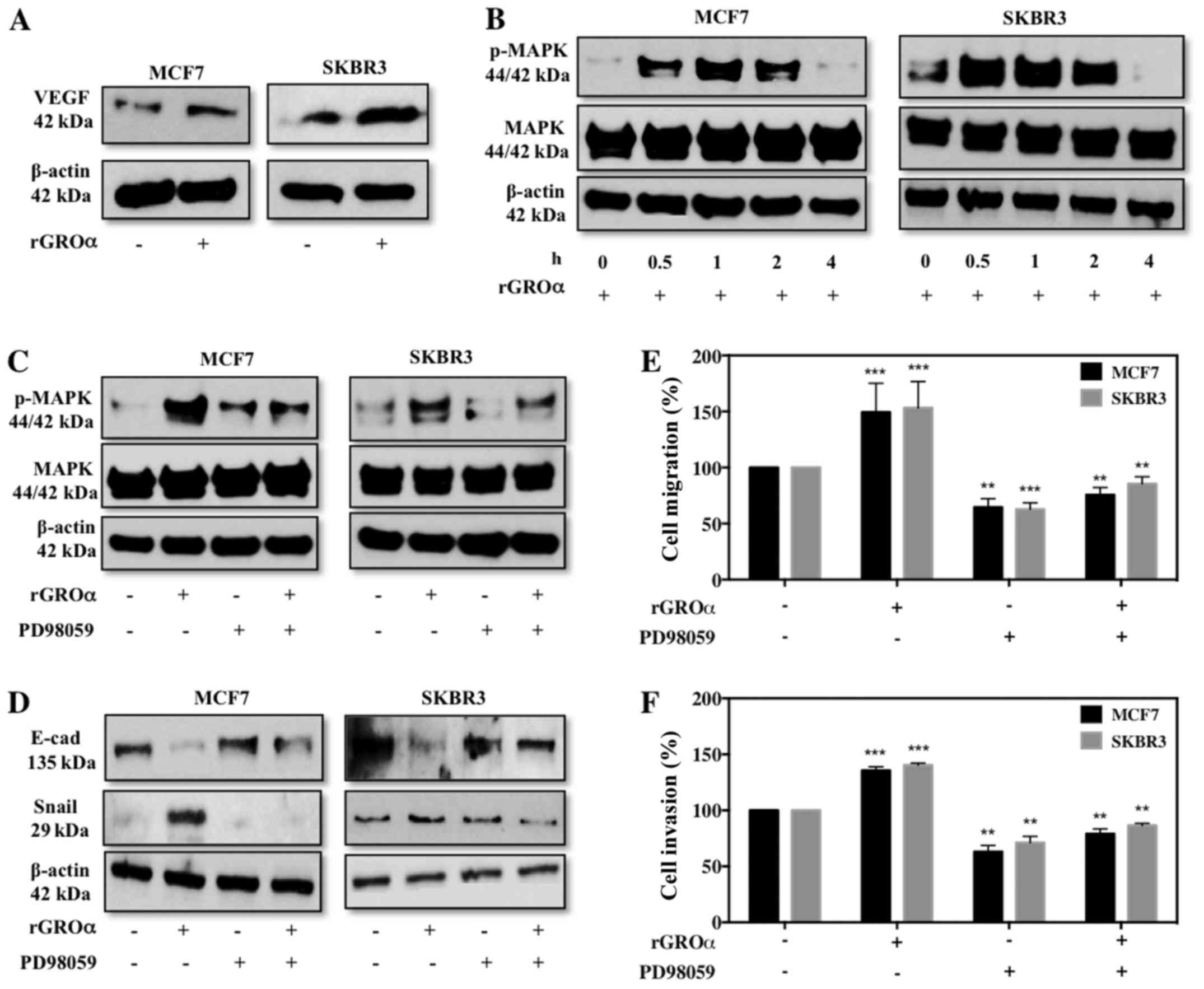
GROα stimulation activates VEGF and
MAPK targets
To further investigate the specific pathway through
which GROα might induce migration/invasion in BC cells, we
evaluated VEGF (angiogenic) and MAPK targets. To our surprise we
observed that MCF7 and SKBR3 cells induced with GROα demonstrated
an enhanced expression level of VEGF (Fig. 5A). Additionally, GROα induction at
different time-points gradually increased p-MAPK with maximum
activation at 1 h time-point and then gradually decreased (Fig. 5B). Next we performed experiments to
verify specificity of MAPK pathway with regards to GROα. For this,
we pre-treated MCF7 and SKBR3 cells with PD98059 (MAPK inhibitor,
50 µM) for 2 h and then induced with GROα (1 ng/ml) to re-confirm
whether the effects of GROα were specific through MAPK pathway in
BC cells. Interestingly, we observed that while GROα-alone
induction activated MAPK, in the presence of PD98059 GROα induction
had reduced effect on MAPK activation, clearly demonstrating that
GROα induces migration/invasion via MAPK pathway in BC cells
(Fig. 5C). To further validate our
findings, we performed functional studies in vitro and
identified that PD98059 significantly reduces migratory/invasion
abilities induced by GROα induction both in MCF7 and SKBR3 cells
(Fig. 5D and E). Our immediate
follow through experiment was to assess if these changes affected
EMT markers; thus we performed western blotting on MCF7 and SKBR3
cells pre-treated with PD98059 and induced with GROα to evaluate
changes in EMT markers. As expected, while GROα alone decreased
E-cadherin and increased Snail expression, pre-treatment with
PD98089 reversed these effects in BC cells (Fig. 5F); suggesting a definitive role of
GROα in regulating BC metastasis.
Discussion
Our study mainly focuses on identifying a specific
target molecule for the treatment of TNBC metastasis. Numerous
studies have reported that chemokines such as IL-6 and IL-8 play a
crucial role in tumor metastasis and chemoresistance (6,30).
Acharyya et al have put forth a study highlighting a network
of paracrine signaling pathway involving inflammatory regulators
such as TNF-α, CXCL1/2 and S100A8/9 that assist tumor cells to
evade stress and survive severe conditions (6). Literature also signifies the
importance of proinflammatory cytokine expression for the growth of
TNBC cells (30). Based on
literature and our data we hypothesized that GROα is an important
inflammatory cytokine regulating TNBC migration/invasion. To prove
the hypothesis, we initially performed a baseline GROα expression
studies using four BC cell lines - SKBR3, MCF7, BT474 and
MDA-MB-231, each representing a subtype of BC. Interestingly, we
noted that expression of GROα was upregulated specifically in
MDA-MB-231/TNBC cell line while negligible amounts were found in
other cell lines from different subtypes (Fig. 1).
Intrigued by these results, we analyzed additional
17 TNBC cell lines as a proof-of-principle to further confirm the
concept that TNBCs express high levels of GROα thus making them
aggressive in nature. Remarkably, 8 cell lines (50%, HCC1937,
HCC1143, HCC1806, HCC38, HCC70, BT-549, MDA-MB-436 and MDA-MB-468)
of the 17 cell lines tested expressed significantly elevated levels
of GROα (Fig. 1). Thus, our results
clearly suggest that GROα might play a vital modulatory role in
TNBC metastasis.
Furthermore, we investigated the role of GROα in BC
cell proliferation, migration and invasion by performing in
vitro functional studies such as MTT assay, scratch assay and
Boyden chamber Matrigel invasion assay respectively. Numerous
cytokine have been demonstrated as an inducer of invasion in
cancers such as breast, gastric, bladder and colorectal (6,26–28).
Here we report that GROα, an inflammatory cytokine, plays a crucial
modulatory role to trigger phenotypic changes in BC cells that
induce the cells to acquire aggressive characteristics to regulate
BC migration/invasion. For this we either knocked down GROα using
GROα specific siRNA in MDA-MB-231 and HCC1937 cells or stimulated
MCF7 and SKBR3 cells with recombinant GROα along with their
respective controls. These phenotypically modified cells were then
tested for the change in proliferation, migration and invasion.
Data from Fig. 2C and D
demonstrated a reduced proliferation (45%) in GROα-knocked down
MDA-MB-231/HCC1937 cells and an enhanced proliferation (40%) in
GROα-stimulated MCF7/SKBR3 cells after 72 h. Even more profound
results were observed from migration and invasion studies,
migration (43%) and invasion (60%) in the GROα siRNA-treated
MDA-MB-231/HCC1937 cells (Fig. 3A and
B) versus migration (45%) and invasion (69%) in the
GROα-stimulated MCF7/SKBR3 cells (Fig.
3C-F) when compared to their respective control cells after 24
h. These results strongly suggest a positive correlation between
GROα expression levels and TNBC migration/invasion.
Our next aim was to decipher the molecular
mechanisms induced by GROα that trigger the metastasis process in
TNBCs. Last decade has evidenced an influx of reports suggesting a
phenomenon called epithelial-mesenchymal transition (EMT) to
initiate the metastasis process.
EMT is a succession of various events that initially
leads to alterations in cell-cell and cell-extracellular matrix
(ECM) communications resulting in the conferring of epithelial
cells the ability to move from its primary site and later resulting
in additional changes for the maintenance of the newly transformed
mesenchymal phenotype (32–35). Specific EMT markers have been
identified that are unique to either epithelial cells (E-cadherin)
or mesenchymal cells (N-cadherin, Vimentin) and others which are
EMT regulators (Snail, Slug, Twist) (35–37).
Data from our experiments clearly showed an upregulation of EMT
regulators such as Snail, Slug, Twist and mesenchymal marker genes
such as Vimentin and N-cadherin along with a notable downregulation
of the epithelial marker gene E-cadherin in GROα-stimulated
MCF7/SKBR3 cells versus contrary results in GROα-knocked down
MDA-MB-231/HCC1937 cells (Fig. 4)
either increasing or decreasing the motility of BC cells.
Investigation to identify a specific pathway through
which GROα influences migration/invasion in BC cells led us to
study the effect of GROα stimulation on VEGF and phosphorylation
status of MAPK in MCF7 and SKBR3 cells. In the presence of GROα, we
observed an increase in both VEGF and pMAPK proteins in BC cells
(Fig. 5A and B). To further confirm
the specificity of MAPK pathway, we pre-treated cells with PD98059
(MEK inhibitor) and observed for changes in pMAPK levels, effect on
EMT markers and eventually migration/invasion of BC cells. With
decrease in pMAPK, we detected a significant reversal of EMT marker
expression and migration/invasion abilities in MCF7 and SKBR3
GROα-stimulated cells (Fig. 5C-F).
These results clearly indicate the effect of GROα-MAPK pathway on
BC cell migration/invasion. In conclusion, results from this study
for the first time define a strong association between GROα and
TNBC subtype while elucidating the modulatory role of GROα in TNBC
cell migration/invasion via MAPK pathway and epithelial to
mesenchymal transition. We also demonstrated that targeting GROα
via targeting MAPK pathway using MAPK inhibitor PD98059 can limit
the effects induced by GROα on BC cells. Thus, a promising
therapeutic strategy can be postulated by targeting GROα in
aggressive breast cancer cells to limit metastasis specifically in
TNBC subtype of cancers.
Acknowledgements
This study was supported by grants from NIH/NCI
1U54- CA14393; U56 CA101599-01; CA15083-25S3; R25DK067015-01;
Department of Defense Breast Cancer Research Program grant
BC043180, NIH-NIMHD U54MD007598, NIH/NCATS CTSI UL1TR000124 to J.V.
Vadgama; and NIH/NIMHD CRECD R25 MD007610, U54MD007598-pilot and
bridge support, and NIMHD 5S21MD 000103-Faculty Retention Award to
Y. Wu. The cell lines used in this study were provided by
Integrated Clinical, Tissue, and Biomarker Database Shared Resource
Core (ICTBD) funded by NIH/NCI NIH/NCI 1U54CA14393.
References
|
1
|
Gonzalez-Angulo AM, Morales-Vasquez F and
Hortobagyi GN: Overview of resistance to systemic therapy in
patients with breast cancer. Adv Exp Med Biol. 608:1–22. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lozano R, Naghavi M, Foreman K, Lim S,
Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, et
al: Global and regional mortality from 235 causes of death for 20
age groups in 1990 and 2010: A systematic analysis for the Global
Burden of Disease Study 2010. Lancet. 380:2095–2128. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008 GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
National Cancer Institute; Altekruse SF,
Kosary CL, Krapcho M, Neyman N, Aminou R, Waldron W, Ruhl J,
Howlader N, Tatalovich Z, Cho H, et al: SEER Cancer Statistics
Review, 1975–2007. National Cancer Institute; Bethesda, MD:
http://seer.cancer.gov/csr/1975_2007/
|
|
6
|
Acharyya S, Oskarsson T, Vanharanta S,
Malladi S, Kim J, Morris PG, Manova-Todorova K, Leversha M, Hogg N,
Seshan VE, et al: A CXCL1 paracrine network links cancer
chemoresistance and metastasis. Cell. 150:165–178. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jones SE: Metastatic breast cancer: The
treatment challenge. Clin Breast Cancer. 8:224–233. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suba Z: Triple-negative breast cancer risk
in women is defined by the defect of estrogen signaling: Preventive
and therapeutic implications. Onco Targets Ther. 7:147–164. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clark O, Botrel TE, Paladini L and
Ferreira MB: Targeted therapy in triple-negative metastatic breast
cancer: A systematic review and meta-analysis. Core Evid. 9:1–11.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
André F and Zielinski CC: Optimal
strategies for the treatment of metastatic triple-negative breast
cancer with currently approved agents. Ann Oncol. 23 Suppl
6:vi46–vi51. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Y, Sarkissyan M, Elshimali Y and
Vadgama JV: Triple negative breast tumors in African-American and
Hispanic/Latina women are high in CD44+, low in CD24+, and have
loss of PTEN. PLoS One. 8:e782592013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Elias AD: Triple-negative breast cancer: A
short review. Am J Clin Oncol. 33:637–645. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Miles DW, Chan A, Dirix LY, Cortés J,
Pivot X, Tomczak P, Delozier T, Sohn JH, Provencher L, Puglisi F,
et al: Phase III study of bevacizumab plus docetaxel compared with
placebo plus docetaxel for the first-line treatment of human
epidermal growth factor receptor 2-negative metastatic breast
cancer. J Clin Oncol. 28:3239–3247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Miller K, Wang M, Gralow J, Dickler M,
Cobleigh M, Perez EA, Shenkier T, Cella D and Davidson NE:
Paclitaxel plus bevacizumab versus paclitaxel alone for metastatic
breast cancer. N Engl J Med. 357:2666–2676. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feliciano P: CXCL1 and CXCL2 link
metastasis and chemoresistance. Nat Genet. 44:8402012. View Article : Google Scholar
|
|
16
|
Opdenakker G and Van Damme J: The
countercurrent principle in invasion and metastasis of cancer
cells. Recent insights on the roles of chemokines. Int J Dev Biol.
48:519–527. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Van der Cappellen J, Van Damme J and
Struyf S: The role of CXC chemokines and their receptors in cancer.
Cancer Lett. 267:226–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dhawan P and Richmond A: Role of CXCL1 in
tumorigenesis of melanoma. J Leukoc Biol. 72:9–18. 2002.PubMed/NCBI
|
|
19
|
Strieter RM, Burdick MD, Mestas J,
Gomperts B, Keane MP and Belperio JA: Cancer CXC chemokine networks
and tumour angiogenesis. Eur J Cancer. 42:768–778. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Richmond A and Thomas HG: Purification of
melanoma growth stimulatory activity. J Cell Physiol. 129:375–384.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Luan J, Shattuck-Brandt R, Haghnegahdar H,
Owen JD, Strieter R, Burdick M, Nirodi C, Beauchamp D, Johnson KN
and Richmond A: Mechanism and biological significance of
constitutive expression of MGSA/GRO chemokines in malignant
melanoma tumor progression. J Leukoc Biol. 62:588–597.
1997.PubMed/NCBI
|
|
22
|
Wang D and Richmond A: Nuclear
factor-kappa B activation by the CXC chemokine melanoma
growth-stimulatory activity/growth-regulated protein involves the
MEKK1/p38 mitogen-activated protein kinase pathway. J Biol Chem.
276:3650–3659. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wilson C, Purcell C, Seaton A, Oladipo O,
Maxwell PJ, O'Sullivan JM, Wilson RH, Johnston PG and Waugh DJ:
Chemotherapy-induced CXC-chemokine/CXC-chemokine receptor signaling
in metastatic prostate cancer cells confers resistance to
oxaliplatin through potentiation of nuclear factor-kappaB
transcription and evasion of apoptosis. J Pharmacol Exp Ther.
327:746–759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Richmond A, Lawson DH, Nixon DW and Chawla
RK: Characterization of autostimulatory and transforming growth
factors from human melanoma cells. Cancer Res. 45:6390–6394.
1985.PubMed/NCBI
|
|
25
|
Xia M and Hyman BT: GROalpha/KC, a
chemokine receptor CXCR2 ligand, can be a potent trigger for
neuronal ERK1/2 and PI-3 kinase pathways and for tau
hyperphosphorylation - a role in Alzheimer's disease? J
Neuroimmunol. 122:55–64. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kawanishi H, Matsui Y, Ito M, Watanabe J,
Takahashi T, Nishizawa K, Nishiyama H, Kamoto T, Mikami Y, Tanaka
Y, et al: Secreted CXCL1 is a potential mediator and marker of the
tumor invasion of bladder cancer. Clin Cancer Res. 14:2579–2587.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bandapalli OR, Ehrmann F, Ehemann V, Gaida
M, Macher-Goeppinger S, Wente M, Schirmacher P and Brand K:
Down-regulation of CXCL1 inhibits tumor growth in colorectal liver
metastasis. Cytokine. 57:46–53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheng WL, Wang CS, Huang YH, Tsai MM,
Liang Y and Lin KH: Overexpression of CXCL1 and its receptor CXCR2
promote tumor invasion in gastric cancer. Ann Oncol. 22:2267–2276.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Keeley EC, Mehrad B and Strieter RM: CXC
chemokines in cancer angiogenesis and metastases. Adv Cancer Res.
106:91–111. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hartman ZC, Poage GM, den Hollander P,
Tsimelzon A, Hill J, Panupinthu N, Zhang Y, Mazumdar A, Hilsenbeck
SG, Mills GB, et al: Growth of triple-negative breast cancer cells
relies upon coordinate autocrine expression of the proinflammatory
cytokines IL-6 and IL-8. Cancer Res. 73:3470–3480. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Klymkowsky MW and Savagner P:
Epithelial-mesenchymal transition: A cancer researcher's conceptual
friend and foe. Am J Pathol. 174:1588–1593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Radisky DC: Epithelial-mesenchymal
transition. J Cell Sci. 118:4325–4326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: New insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cavallaro U and Christofori G: Cell
adhesion and signalling by cadherins and Ig-CAMs in cancer. Nat Rev
Cancer. 4:118–132. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|