Introduction
Ovarian cancer is the leading cause of death among
all gynecological malignancies. Despite advancements in the
surgical and systemic treatment of ovarian cancer, the 5-year
survival rate remains low (1). The
etiology of ovarian cancer also remains largely unclear. Some
evidence has indicated that endocrine factors may influence the
tumorigenesis of ovarian cancer (1,2).
Ovarian cancer is more common in patients with elevation of the
gonadotropins follicle stimulating hormone (FSH) and luteinizing
hormone (LH), such as postmenopausal women or women who have
received treatment for the induction of ovulation (3–5).
Additionally, reduced risk for ovarian cancer is associated with
multiple pregnancies, breastfeeding, and oral contraceptive use,
which are all associated with lower levels of and reduced exposure
to gonadotropins (1). Moreover, the
concentrations of both FSH and LH in cysts and the tumor fluid of
ovarian cancer are greater than in borderline tumors, benign
tumors, and functional cysts of the ovary (6). The expression of FSH receptor (FSHR)
and LH receptor (LHR) and the growth-stimulating effect of their
ligands in normal and neoplastic ovarian surface epithelium (OSE)
and ovarian cancer cells have been demonstrated (2,4,7–9),
although some reports are controversial (10,11).
Recently, some studies reported that treatment of ovarian cancer
cells with gonadotropins in vitro enhances tumor
angiogenesis and cell adhesion (2,12–14).
Together, these observations suggest a crucial role for
gonadotropins in the development and progression of ovarian
cancer.
Cyclooxygenases (COXs) are a family of
myeloperoxidases that catalyze the biosynthesis of prostaglandins
(PGs) from arachidonic acid. COX1 is constitutively expressed in
most tissues and plays a role in various physiological functions,
whereas COX2 is transiently inducible by stimuli such as cytokines,
growth factors, mitogens, tumor promoters and hormones and
regulates inflammation, differentiation, mitogenesis, and
angiogenesis (15–18). A number of studies have revealed
that COX2 plays a critical role in the ovulatory process, whereas
repeated ovulations were suggested to cause neoplastic
transformation of the ovarian epithelium (4,9,19,20).
COX2 is rarely found in normal OSE but is present in premalignant
ovarian inclusion cysts and is expressed in most ovarian cancer
cells (21,22). Additionally, to some extent, COX2 is
an independent prognostic factor for overall survival in patients
with ovarian cancer (23,24).
However, the roles of gonadotropins and of the
concomitant elevation of COX2 in ovarian cancer development and
progression have not been fully characterized. Therefore, we
conducted the present study to examine the effect of gonadotropins
on metastasis-related proteases and the invasiveness of ovarian
cancer cells, thus to identify the key regulator of the
gonadotropin pathway in ovarian cancer cells.
Materials and methods
Cell lines, drugs, inhibitors, and
ovary tissues
The ovarian cancer cell lines SKOV3 and HO8910 were
obtained from the American Type Culture Collection (ATCC, Manassas,
VA, USA). The SKOV3 cell line was cultured in McCoy's 5A medium
(Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum
(FBS, Gibco), 100 U/ml penicillin and 100 µg/ml streptomycin
(Gibco) in a 5% CO2 humidified atmosphere at 37°C. The
HO8910 cell line was cultured in RPMI-1640 (Gibco) under the
above-mentioned culture conditions. Recombinant human FSH (75 IU)
and LH (75 IU) were purchased from Merck Serono (Darmstadt,
Germany). NS-398, a selective cyclooxygenase-2 (COX-2) inhibitor,
was purchased from Beyotime (Shanghai, China). In this study, 10
µmol/l of NS-398 was used, which specifically inhibits COX2 enzyme
activity but has no effect on COX1 activity (25).
The ovary tissues, used as positive controls for
FSHR and LHR detection, were obtained from patients of ovarian
cancer undergoing surgical excision. The investigation has been
conducted in accordance with the ethical standards and according to
the Declaration of Helsinki and has been approved by the ethics
committee of the China-Japan Friendship Hospital. Written informed
consent was obtained from each patient.
Immunocytochemical staining
SKOV3 and HO8910 cells were seeded on 18-mm glass
coverslips in a 6-well culture plate and cultured for 48 h.
Coverslips were washed twice with PBS and fixed in ice-cold acetone
for 5 min. After the cells were washed twice with PBS, they were
permeabilized with 0.1% Triton X-100 in PBS for 5 min at room
temperature. The coverslips were then washed with PBS and further
incubated with normal goat serum (Boster, Wuhan, China) for 20 min
at room temperature to block non-specific binding. The cells were
then incubated with FSHR antibody (cat. PB1120; 1:200, Boster) or
LHR antibody (cat. A01120; 1:500, Boster) at 4°C overnight. Rabbit
IgG (cat. ab27478, Abcam, Cambridge, UK) was used as isotype
control instead of primary antibody. The binding of the primary
antibodies was visualized using a ChemMate Detection kit (cat.
SV0002; Boster). The coverslips were lightly counterstained with
Mayer's hematoxylin for 30 sec.
Cell viability assay
SKOV3 and HO8910 cells were seeded in 96-well plates
at a density of 1000 cells/well and incubated for 24 h. To examine
the effect of FSH and LH on the viability of SKOV3 and HO8910
cells, the cells were treated with 100 and 500 mIU/ml FSH and LH
alone or in combination. The levels of gonadotropins were chosen
because 100 mIU/ml are commonly achieved during ovarian
hyperstimulation (COH) and in 2–3 years after menopause (10,26),
while 500 mIU/ml is based on previous references (11,27).
Cell viability was measured using an MTS cell proliferation kit
(Promega, Madison, WI, USA) per the manufacturer's instructions.
Assays were performed in triplicate and repeated three times.
Annexin V-FITC staining
Cell apoptosis was evaluated using an Annexin V-FITC
apoptosis kit (eBioscience, San Diego, CA, USA) according to the
manufacturer's instruction. In brief, SKOV3 and HO8910 cells were
cultured in 10-cm plates, and treated with 500 mIU/ml FSH and LH
for 24 h. After that, cells were collected and washed with ice-cold
PBS, and then resuspended in 100 µl of binding buffer, containing 5
µl of each Annexin V-FITC and propidium iodide (PI) solution. The
samples were then examined using flow cytometer (FACSCalibur, BD
Biosciences, Franklin Lakes, NJ, USA).
Cell migration and invasion
assays
SKOV3 and HO8910 cells were grown in 6-well plates
at a density of 5×105 cells/well to nearly confluence
and replaced the growth medium with serum-free medium for 5-h
incubation to arrest cell growth. Then the cell monolayer was
scraped with a pipette tip to generate scratch wounds. After the
cells were washed three times to remove cell debris, they were
cultured in serum-free medium containing 500 mIU/ml FSH/LH and 10
µmol/l NS-398 alone or in combination. Images were obtained, and
the widths of the gaps were measured and analyzed at 24 h.
Cell invasion was assessed using 24-well Transwell
inserts (Corning Inc., Corning, NY, USA) with 8-µm pores coated
with 1 mg/ml Matrigel (BD Biosciences). Cells in the upper
compartment of the chamber were suspended in culture medium with 1%
FBS, and the lower chamber contained culture medium with 20% FBS.
After cell adhesion, the medium in the upper chamber was replaced
with fresh medium containing 1% FBS, 500 mIU/ml FSH and LH, FSH and
LH in combination with 10 µmol/l NS-398, or NS-398 alone. After a
24-h incubation, the upper surface of the filter was erased
carefully with a cotton swab, and then the cells that passed
through the Matrigel matrix were fixed and stained with 0.05%
crystal violet and counted in 5 random microscopic fields.
RT-PCR and qRT-PCR
Total RNA was extracted using TRIzol (Invitrogen)
and converted to cDNA using Superscript III reverse transcriptase
(Takara, Shiga, Japan). The cDNA was amplified by PCR using Premix
Ex Taq™ (Takara). Quantitative real-time PCR (qRT-PCR) was
performed using an iQ™ SYBR® Green Supermix kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) on a CFX96 Touch™ Real-time
PCR instrument (Bio-Rad Laboratories, Inc.). The primer sets used
in the RT-PCR and qRT-PCR are presented in Table I.
 | Table I.Specific primer sequences for RT-PCR
and qRT-PCR. |
Table I.
Specific primer sequences for RT-PCR
and qRT-PCR.
| Gene name | Primer sequences
(5′-3′) |
|---|
| FSHRa | F:
TATGACTCAGGCTAGGGGTC |
|
| R:
ATAAGGAACCTGGGGACTGT |
| LHRa | F:
CACCCCGATGTGCTCCTGAA |
|
| R:
TAGAGTGATGACGGTGAGGG |
| COX1 | F:
CGCTCCAACCTTATCCCCAGTCCCC |
|
| R:
CTTTAGGCACAGAGGGCAGAATACG |
| COX2 | F:
TCAAGTCCCTGAGCATCTACGGTT |
|
| R:
CTGTTGTGTTCCCGCAGCCAGATT |
| E-cadherin | F:
ACAATGCCGCCATCGCTTAC |
|
| R:
AACTCTCTCGGTCCAGCCCA |
| Slug | F:
GCCCCATTAGTGATGAAGAGGAAA |
|
| R:
AGCCCAGAAAAAGTTGAATAGGTC |
| Vimentin | F:
AAGCAGGAGTCCACTGAGTA |
|
| R:
GCTTCAACGGCAAAGTTCTC |
| MMP2 | F:
AACTACGATGATGACCGCAA |
|
| R:
CTCCTGAATGCCCTTGATGT |
| MMP9 | F:
GTCTTCCAGTACCGAGAGAAAGCCT |
|
| R:
CAGGATGTCATAGGTCACGTAGCCC |
| GAPDH | F:
CTTAGCACCCCTGGCCAAG |
|
| R:
GATGTTCTGGAGAGCCCCG |
Antibodies and western blots
Antibodies specific to FSHR (cat. PB1120; 1:200) and
LHR (cat. A01120; 1:200) were obtained from Boster. Antibodies
specific to COX1 (cat. ab109025, 1:2000), COX2 (cat. ab52237,
1:1000), E-cadherin (cat. ab76055, 1:500), slug (cat. ab51772,
1:500), and vimentin (cat. ab133260, 1:500) were purchased from
Abcam. Antibodies specific to MMP2 (cat. #4022, 1:1000), MMP9 (cat.
#3852, 1:1000), and GAPDH (cat. #2118, 1:2000) were obtained from
Cell Signaling Technology (Beverly, MA, USA). Protein was prepared
from the cells according to the kit manual (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). After electrophoresis, the
proteins were transferred to polyvinylidene fluoride (PVDF, Merck
Millipore, Darmstadt, Germany) membranes and probed with the
indicated primary antibodies. Incubation with secondary antibodies
of HRP-linked anti-rabbit IgG (cat. #7074, Cell Signaling
Technology) or HRP-linked anti-mouse IgG (cat. #7076, Cell
Signaling Technology) according to the species reactivity of
primary antibodies was performed at room temperature for 1 h. The
blots were developed with chemiluminescent substrate (Thermo Fisher
Scientific, Inc.), and autoradiography was performed with X-OMAT
film (Kodak, Rochester, NY, USA).
Statistical analyses
The data are presented as the mean ± SD. To assess
the statistical significance of differences, an analysis of
variance (ANOVA, SPSS software, version 13.0; SPSS Inc., USA) was
performed. A P-value of <0.05 was considered to indicate a
statistically significant difference.
Results
Gonadotropins do not induce
proliferation in ovarian cancer cells despite the expression of
FSHR and LHR in these cells
Gonadotropins are considered to play a pivotal role
in neoplastic conversion and growth potential via their receptors
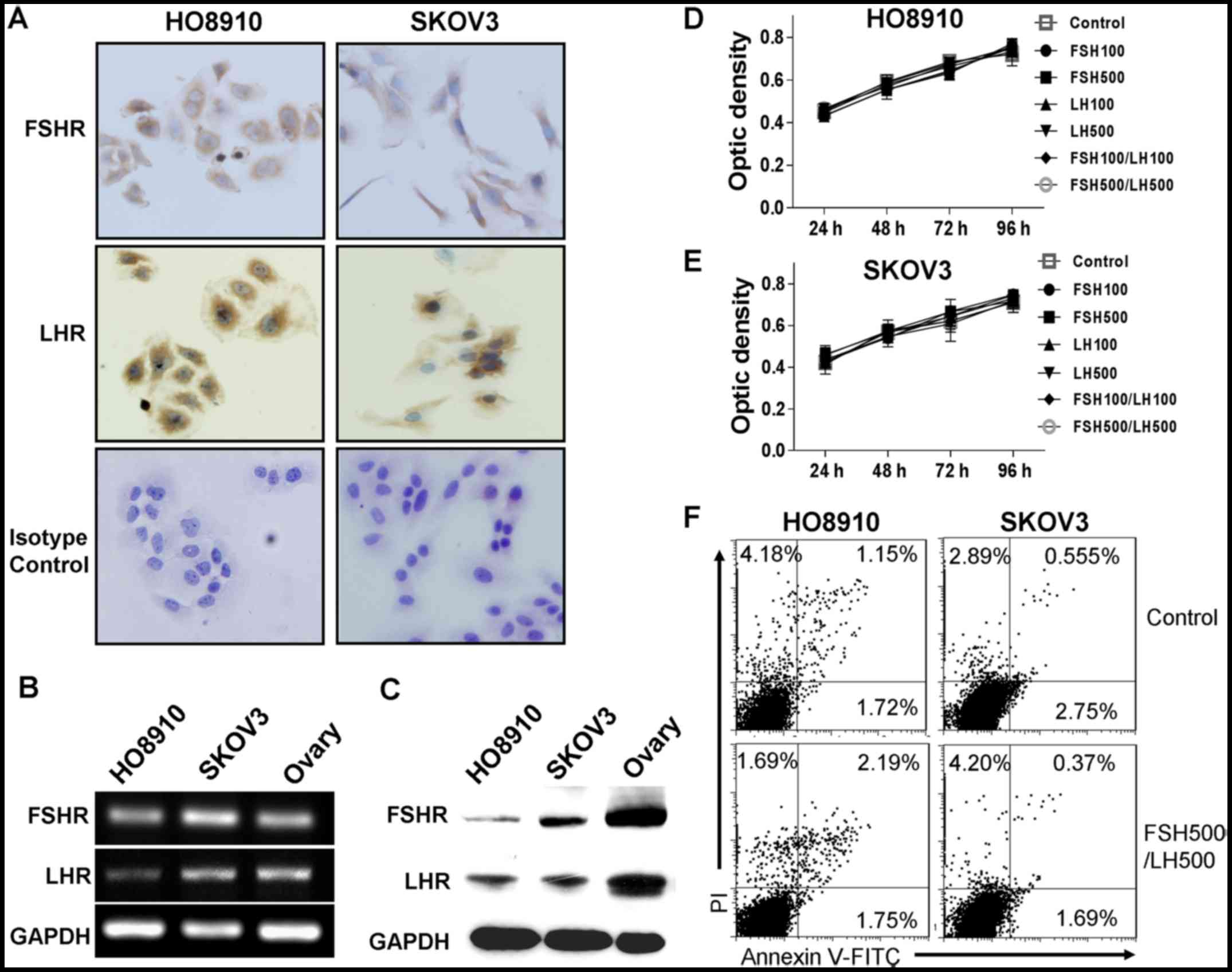
FSHR and LHR during the tumorigenesis of ovarian cancers (9). In this study, we first showed the
expression of FSHR and LHR in SKOV3 and HO8910 cells with
immunocytochemical staining, RT-PCR, and western blotting (Fig. 1A-C). Then, we determined the effect
of gonadotropins on the growth of SKOV3 and HO8910 cells. To our
surprise, neither FSH nor LH stimulated the growth of ovarian
cancer cells at either tested concentration (100 and 500 mIU/ml).
Furthermore, the combination of FSH and LH also failed to increase
the viable cell numbers as compared to prior treatment (Fig. 1D and E). In addition, no significant
variations in the ratio of cell apoptosis were observed in either
cell line after treatment with 500 mIU/ml FSH and LH for 24 h
(Fig. 1F).
Gonadotropins induce COX2 upregulation
in ovarian cancer cells
It is well documented that COX2 is an inducible,
immediate early gene that, in addition to inflammatory stimuli,
participates in the process of malignant cell transformation, while
COX1 is constitutively expressed in most tissues (15). In our study, COX2 was rapidly
induced after treatment with 500 mIU/ml FSH combined with 500
mIU/ml LH for 6 h and was steadily upregulated up to 24 h after
continued treatment in HO8910 cells (Fig. 2A and B). In SKOV3 cells treated with
FSH and LH, the expression of COX2 in SKOV3 cells increased slowly
and then elevated sharply at 24 h (Fig.
2C and D). In both cell lines, COX1 maintained stable
expression levels with or without FSH and LH treatment (Fig. 2). These results demonstrate that
COX2 is selectively upregulated by FSH and LH in SKOV3 and HO8910
cells.
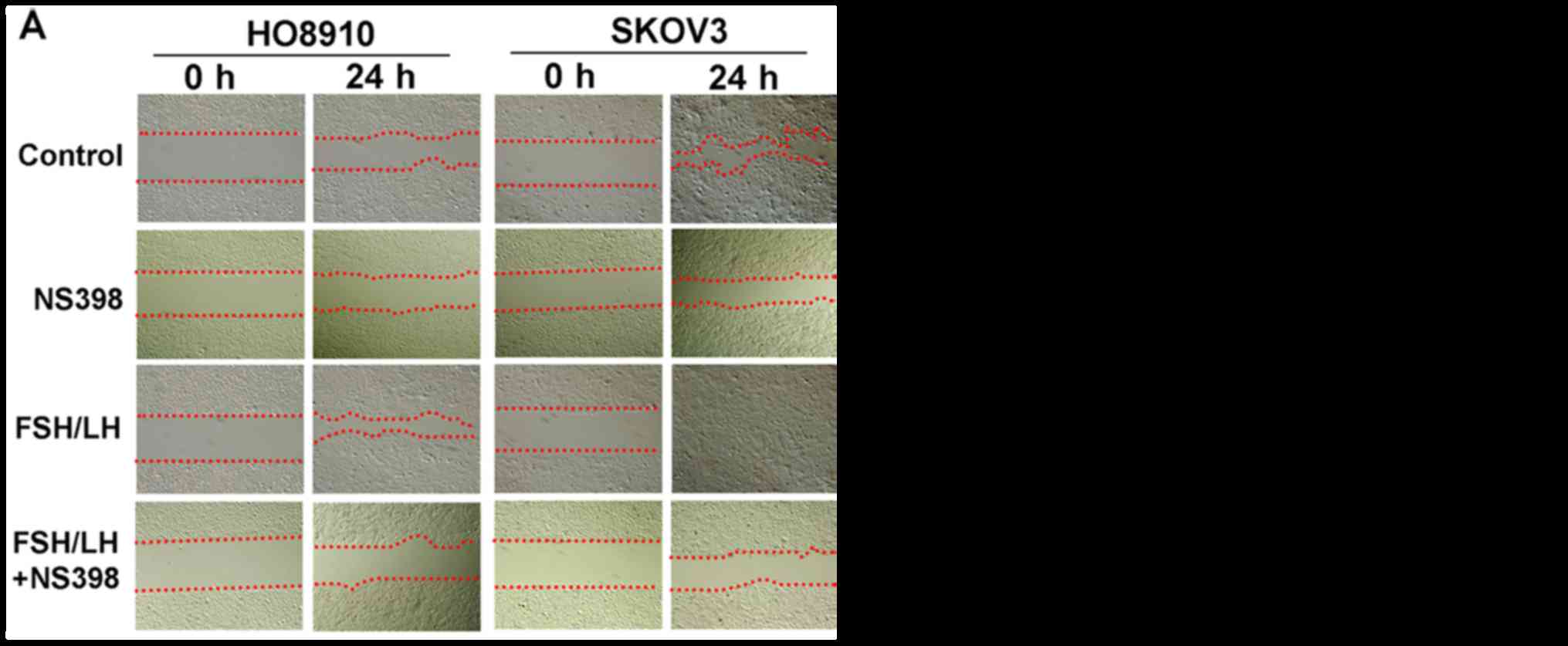
Gonadotropins promote ovarian cancer
cell migration and invasion
To further investigate the role of excessive
exposure to gonadotropins in the development and progression of
ovarian cancer, the migration and invasion activities of SKOV3 and
HO8910 cells with or without gonadotropin treatment was analyzed
using wound-healing and Transwell assays, respectively. At 24 h
after the scratch injury, the SKOV3 cells treated with 500 mIU/ml
FSH and LH were able to cover 95.3% of the scratch, whereas the
control cells only covered 54%. Importantly, NS-398, a specific
COX2 inhibitor, blocked SKOV3 cell migration even in the presence
of FSH and LH, which resulted in only 29% coverage of the scratch
(Fig. 3). Similarly, in HO8910
cells, 500 mIU/ml FSH and LH significantly promoted cell migration
compared to the control (73.3% compared with 38%), while NS-398
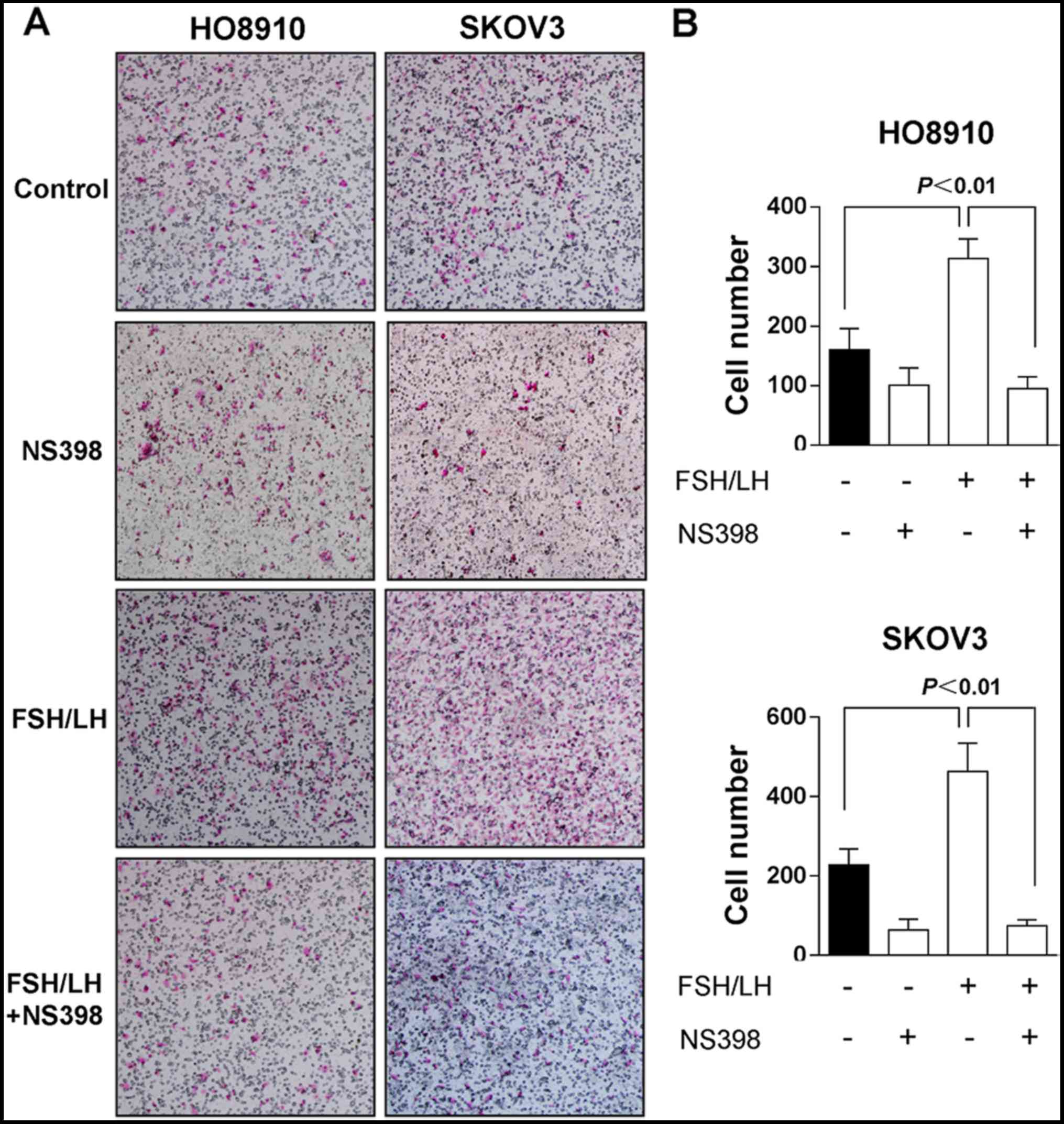
attenuated the effect of gonadotropins (18.3%) (Fig. 3). The invasion assays also showed
that treatment with 500 mIU/ml FSH and LH significantly promoted
ovarian cancer cell invasion, whereas cell invasion was effectively
blocked by NS-398 (Fig. 4). As low
level of COX2 expressed in ovarian cancer cells without FSH/LH
stimulation, NS-398 treatment alone also, to some extent, showed
inhibition effects in both cell migration and invasion.
Collectively, these results demonstrated that the combination of
FSH and LH notably enhanced ovarian cancer cell migration and
invasion in vitro, but these effects were markedly inhibited
by NS-398.
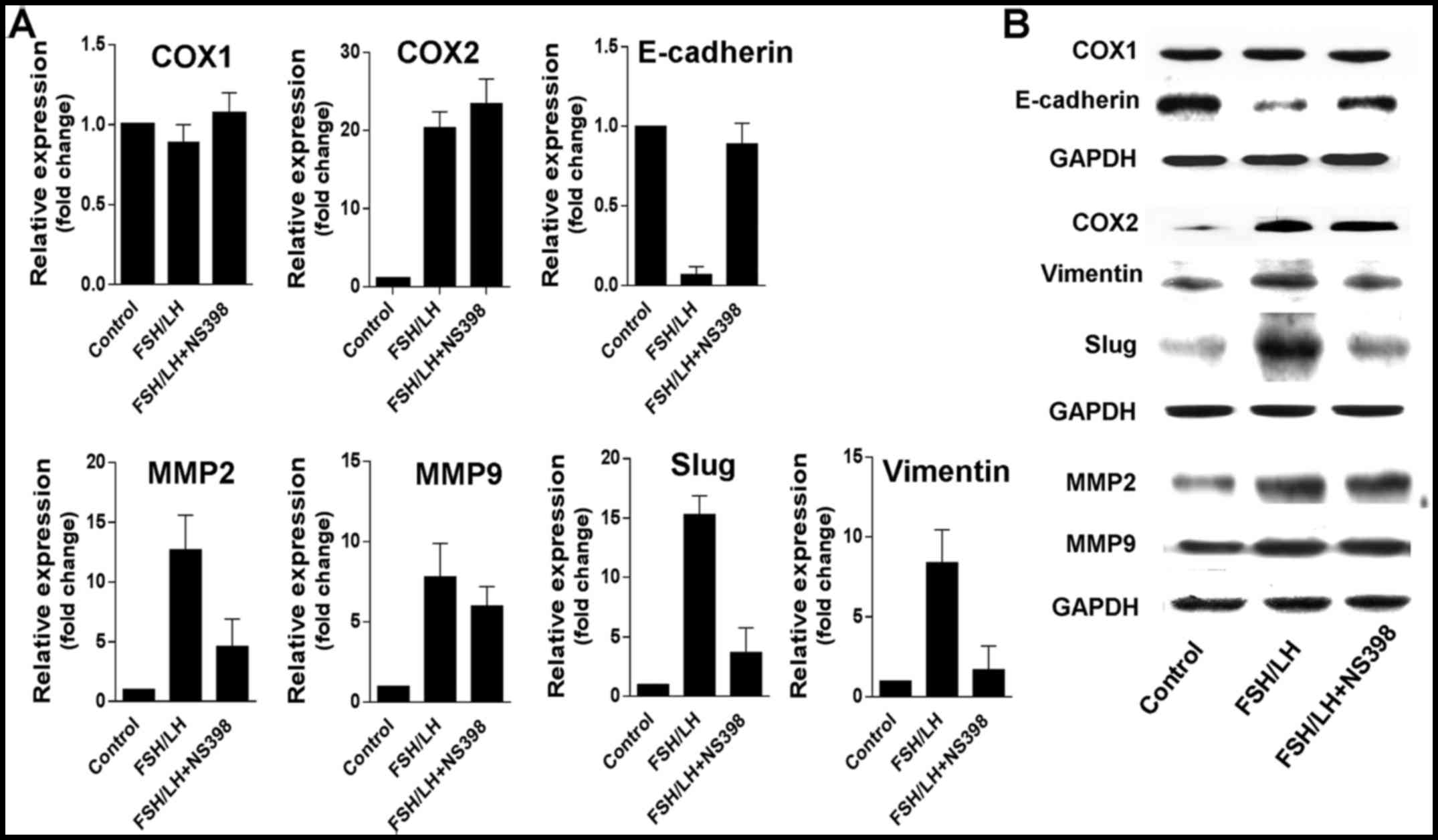
Gonadotropin-induced cell migration
and invasion occur due to COX2-dependent enhancement of
epithelial-mesenchymal transition (EMT) and upregulation of MMP2
and MMP9
EMT enables polarized epithelial cells to lose
adherence, thus enhancing the migratory capacity and acquirement of
invasive properties in these cells. In our study, the results
showed that treatment with FSH and LH led to ongoing downregulation
of E-cadherin and simultaneous upregulation of slug and vimentin in
SKOV3 cells (Fig. 5). Furthermore,
FSH and LH stimulated increased expression of MMP2 and MMP9, which
indicates that proteolysis, to some extent, plays a role in cell
invasion induced by gonadotropins (Fig.
5). Notably, the effects of EMT and MMP expression induced by
gonadotropins were specifically blocked by NS-398. However, neither
mRNA nor protein of COX2 expression was inhibited obviously by
NS-398 (Fig. 5), which indicated
that NS-398 just inhibited the enzyme activity but not the
expression of COX2. In addition, the same changes were observed in
HO8910 cells (data not shown). Taken together, the promotion of
cell migration and invasion by gonadotropins in ovarian cancer is
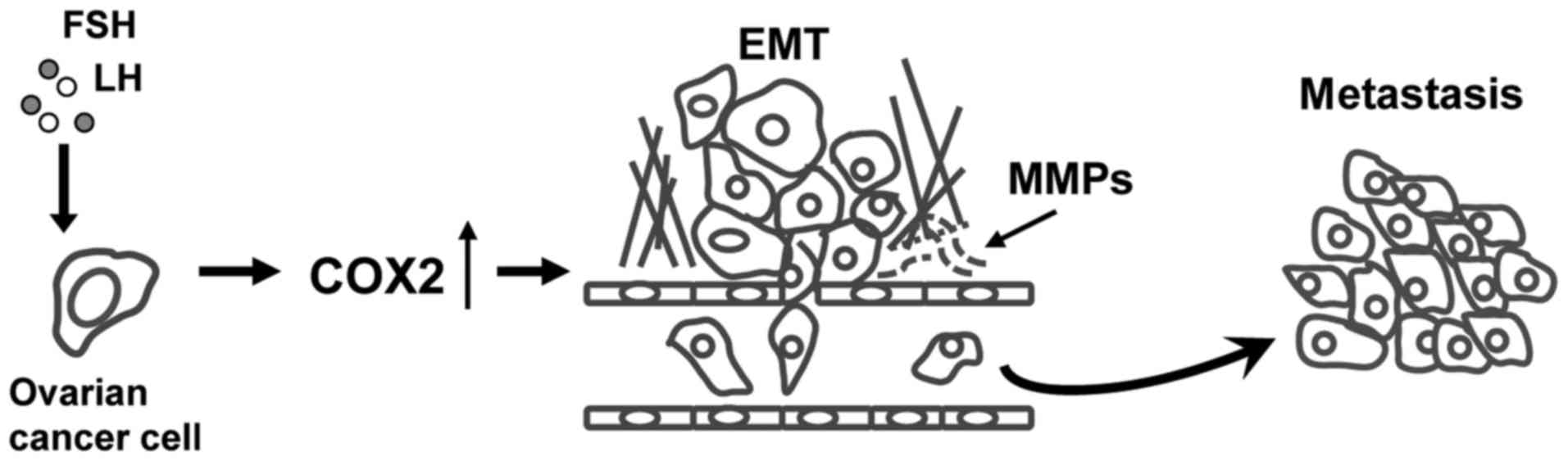
mostly dependent on the activation of COX2 pathway (Fig. 6).
Discussion
High levels of gonadotropins during ovulation, loss
of gonadal negative feedback during menopause, and premature
ovarian failure are all thought to be critical risk factors for
ovarian cancer (1–3). FSH and LH levels in the cyst fluid
from ovarian malignant tumors are significantly higher than those
from benign tumors (6,28). Furthermore, gonadotropins not only
induce both ovarian cancer cell proliferation and migration but
also stimulate normal OSE proliferation (4,9). In
this study, our data showed that the combination of 500 mIU/ml FSH
and LH significantly promotes ovarian cancer cell migration and
invasion within 24 h compared to cells not exposed to
gonadotropins. However, we did not observe proliferation in ovarian
cancer cells treated with 100 and 500 mIU/ml FSH and LH alone or in
combination. As previous studies have shown, the association of
gonadotropins with the proliferation of OSE and ovarian cancer
cells is inconsistent and has been shown to be stimulatory,
irrelevant, or even inhibitory (2,11,13).
Variations in the concentration of gonadotropin and density of
cultured cells and gonadotropin receptors may, to some extent,
contribute to these discrepancies.
To date, there is increasing evidence that the
expression of COX2 is upregulated in a variety of human cancers,
such as colorectal, skin, liver, breast, and lung cancers (29–33).
Elevated COX2 expression also has been detected in malignant
ovarian tumors, and high levels of COX2 expression are correlated
with poor prognosis and decreased overall survival in human ovarian
cancer (9,23). Our results showed that excessive
exposure to gonadotropins induced significant upregulation of COX2
expression in SKOV3 and HO8910 cells, whereas COX1 expression was
unchanged. Moreover, treatment with gonadotropins noticeably
promoted cell migration and invasion in SKOV3 and HO8910 cells, but
these effects can be sufficiently blocked by NS-398. Therefore,
these data indicate that COX2 plays an essential role in
gonadotropin-induced cell migration and invasion.
With further exploration of the mechanisms
underlying the above mentioned effects, we demonstrated that
treatment with FSH and LH enhanced the expression of MMPs and
epithelial-mesenchymal transition in both SKOV3 and HO8910 cells.
MMPs are a family of highly homologous, protein-degrading,
zinc-dependent enzyme endopeptidases; among the MMPs, MMP2 and MMP9
are notably correlated with tumor invasion and metastasis (34). EMT, a key event in the promotion of
neoplastic progression and the development of metastasis, is
characterized by multiple molecular changes, including
downregulation of E-cadherin, upregulation of its transcriptional
repressors snail and slug, and augmentation of the expression of
vimentin, fibronectin, and collagen (35). In our study, inhibition of COX2 by
NS-398 restored E-cadherin expression and repressed slug, vimentin,
MMP2 and MMP9 expression, which significantly blocked cell
migration and invasion induced by gonadotropins. Experimental
studies have also shown that COX2 inhibitors block tumor growth and
enhance anticancer activity when combined with chemotherapeutic
drugs, mainly through antiangiogenic and proapoptotic effects
(36,37). Recently, Majumder and his colleagues
reported that COX2 upregulation or overexpression in human breast
cancer cells induced breast cancer stem cells via
EP4/PI3K/AKT/NOTCH/WNT pathway and promoted tumor growth and
metastasis in immunodeficient mice, which can be reliably blocked
by COX2 inhibitor or EP4 antagonist (38). These data further suggest that COX2
can serve as not only a predictor of disease outcome but also a
target for the treatment of ovarian cancers.
In summary, the current study, to a certain extent,
demonstrates that gonadotropins promote cell migration and invasion
via induction of COX2 expression in ovarian cancer cells, whereas
specific inhibitors of COX2 significantly prevent these effects.
However, some limitations of this study must be highlighted, such
as incomplete elimination of endogenous COX2, an inappropriate
method for detection of cell proliferation, lack of enough evidence
from other ovarian cancer cell lines and primary cells to further
support the work. Thus, to get a more reliable conclusion, further
well-designed investigations are needed.
Acknowledgements
We would like to thank Dr Zhongjun Dong (School of
Medicine, Tsinghua University) for the helpful comments and for
critically reading of the manuscript. This work was supported by
the National Natural Science Foundation of China (81372779 and
81372777) and the Anhui Provincial Natural Science Foundation
(110406 M176).
Glossary
Abbreviations
Abbreviations:
|
COXs
|
cyclooxygenases
|
|
PGs
|
prostaglandins
|
|
FSH
|
follicle stimulating hormone
|
|
LH
|
luteinizing hormone
|
|
FSHR
|
FSH receptor
|
|
LHR
|
LH receptor
|
|
OSE
|
ovarian surface epithelium
|
|
MMPs
|
matrix metalloproteinases
|
|
EMT
|
epithelial-mesenchymal transition
|
References
|
1
|
Gharwan H, Bunch KP and Annunziata CM: The
role of reproductive hormones in epithelial ovarian carcinogenesis.
Endocr Relat Cancer. 22:R339–R363. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mertens-Walker I, Baxter RC and Marsh DJ:
Gonadotropin signalling in epithelial ovarian cancer. Cancer Lett.
324:152–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paulson RJ: Treatment of infertility and
the risk of ovarian cancer: Are the data reassuring? Fertil Steril.
105:605–606. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Burdette JE, Kurley SJ, Kilen SM, Mayo KE
and Woodruff TK: Gonadotropin-induced superovulation drives ovarian
surface epithelia proliferation in CD1 mice. Endocrinology.
147:2338–2345. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rodgers LH, Oh E, Young AN and Burdette
JE: Loss of PAX8 in high-grade serous ovarian cancer reduces cell
survival despite unique modes of action in the fallopian tube and
ovarian surface epithelium. Oncotarget. 7:32785–32795.
2016.PubMed/NCBI
|
|
6
|
Rzepka-Górska I, Chudecka-Głaz A and
Kosmowska B: FSH and LH serum/tumor fluid ratios and malignant
tumors of the ovary. Endocr Relat Cancer. 11:315–321. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saddick SY: In vitro regulation of sheep
ovarian surface epithelium (OSE) proliferation by local ovarian
factors. Saudi J Biol Sci. 19:285–290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mertens-Walker I, Bolitho C, Baxter RC and
Marsh DJ: Gonadotropin-induced ovarian cancer cell migration and
proliferation require extracellular signal-regulated kinase 1/2
activation regulated by calcium and protein kinase C{delta}. Endocr
Relat Cancer. 17:335–349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hilliard TS, Modi DA and Burdette JE:
Gonadotropins activate oncogenic pathways to enhance proliferation
in normal mouse ovarian surface epithelium. Int J Mol Sci.
14:4762–4782. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tourgeman DE, Lu JJ, Boostanfar R, Amezcua
C, Felix JC and Paulson RJ: Human chorionic gonadotropin suppresses
ovarian epithelial neoplastic cell proliferation in vitro. Fertil
Steril. 78:1096–1099. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ivarsson K, Sundfeldt K, Brännström M,
Hellberg P and Janson PO: Diverse effects of FSH and LH on
proliferation of human ovarian surface epithelial cells. Hum
Reprod. 16:18–23. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim D, Lee J and Johnson AL: Vascular
endothelial growth factor and angiopoietins during hen ovarian
follicle development. Gen Comp Endocrinol. 232:25–31. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tzuman YC, Sapoznik S, Granot D, Nevo N
and Neeman M: Peritoneal adhesion and angiogenesis in ovarian
carcinoma are inversely regulated by hyaluronan: The role of
gonadotropins. Neoplasia. 12:51–60. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lau M-T, Wong AST and Leung PCK:
Gonadotropins induce tumor cell migration and invasion by
increasing cyclooxygenases expression and prostaglandin E(2)
production in human ovarian cancer cells. Endocrinology.
151:2985–2993. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Harris RE: Cyclooxygenase-2 (cox-2) and
the inflammogenesis of cancer. Subcell Biochem. 42:93–126. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moran EM: Epidemiological and clinical
aspects of nonsteroidal anti-inflammatory drugs and cancer risks. J
Environ Pathol Toxicol Oncol. 21:193–201. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pang LY, Hurst EA and Argyle DJ:
Cyclooxygenase-2: A role in cancer stem cell survival and
repopulation of cancer cells during Therapy. Stem Cells Int.
2016:20487312016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Baek SJ and Eling T: COX
inhibitors directly alter gene expression: Role in cancer
prevention? Cancer Metastasis Rev. 30:641–657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Duffy DM: Novel contraceptive targets to
inhibit ovulation: The prostaglandin E2 pathway. Hum Reprod Update.
21:652–670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rask K, Zhu Y, Wang W, Hedin L and
Sundfeldt K: Ovarian epithelial cancer: A role for PGE2-synthesis
and signalling in malignant transformation and progression. Mol
Cancer. 5:622006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li S, Miner K, Fannin R, Barrett Carl J
and Davis BJ: Cyclooxygenase-1 and 2 in normal and malignant human
ovarian epithelium. Gynecol Oncol. 92:622–627. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Menczer J: Cox-2 expression in ovarian
malignancies: A review of the clinical aspects. Eur J Obstet
Gynecol Reprod Biol. 146:129–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee JY, Myung SK and Song YS: Prognostic
role of cyclooxygenase-2 in epithelial ovarian cancer: A
meta-analysis of observational studies. Gynecol Oncol. 129:613–619.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Whynott RM, Manahan P and Geisler JP:
Vascular endothelial growth factor (VEGF) and cyclooxygenase 2 (COX
2) immunostaining in ovarian cancer. Eur J Gynaecol Oncol.
37:164–166. 2016.PubMed/NCBI
|
|
25
|
Futaki N, Takahashi S, Yokoyama M, Arai I,
Higuchi S and Otomo S: NS-398, a new anti-inflammatory agent,
selectively inhibits prostaglandin G/H synthase/cyclooxygenase
(COX-2) activity in vitro. Prostaglandins. 47:55–59. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi JH, Wong AS, Huang HF and Leung PC:
Gonadotropins and ovarian cancer. Endocr Rev. 28:440–461. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wright JW, Toth-Fejel S, Stouffer RL and
Rodland KD: Proliferation of rhesus ovarian surface epithelial
cells in culture: lack of mitogenic response to steroid or
gonadotropic hormones. Endocrinology. 143:2198–2207. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barczyński B, Rogala E, Nowicka A,
Nurzyńska-Flak J and Kotarski J: Pelvic pain, free fluid in pelvis,
and human chorionic gonadotropin serum elevation: Recurrence of
malignant ovarian germ-cell tumor or early pregnancy? Eur J
Gynaecol Oncol. 34:489–492. 2013.PubMed/NCBI
|
|
29
|
Guo Z, Jiang J-H, Zhang J, Yang HJ, Yang
FQ, Qi YP, Zhong YP, Su J, Yang RR, Li LQ, et al: COX-2 promotes
migration and invasion by the side population of cancer stem
cell-like hepatocellular carcinoma cells. Medicine (Baltimore).
94:e18062015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Majumder M, Landman E, Liu L, Hess D and
Lala PK: COX-2 elevates oncogenic miR-526b in breast cancer by EP4
activation. Mol Cancer Res. 13:1022–1033. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu Y, Sun H, Hu M, Zhang Y, Chen S, Tighe
S and Zhu Y: The Role of cyclooxygenase-2 in colorectal
carcinogenesis. Clin Colorectal Cancer. Oct 6–2016.(Epub ahead of
print). View Article : Google Scholar :
|
|
32
|
Hua H-K, Jin C, Yang L-J, Tao S-Q and Zhu
X-H: Expression of cyclooxygenase-2 in squamous cell carcinoma and
keratoacanthoma and its clinical significance. Cell Biochem
Biophys. 72:475–480. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Maeng H-J, Lee W-J, Jin Q-R, Chang J-E and
Shim W-S: Upregulation of COX-2 in the lung cancer promotes
overexpression of multidrug resistance protein 4 (MRP4) via
PGE2-dependent pathway. Eur J Pharm Sci. 62:189–196. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tang Y, Lv P, Sun Z, Han L and Zhou W and
Zhou W: 14-3-3β Promotes migration and invasion of human
hepatocellular carcinoma cells by modulating expression of MMP2 and
MMP9 through PI3K/Akt/NF-κB pathway. PLoS One. 11:e01460702016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bottoni P, Isgrò MA and Scatena R: The
epithelial-mesenchymal transition in cancer: A potential critical
topic for translational proteomic research. Expert Rev Proteomics.
13:115–133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kurtova AV, Xiao J, Mo Q, Pazhanisamy S,
Krasnow R, Lerner SP, Chen F, Roh TT, Lay E, Ho PL, et al: Blocking
PGE2-induced tumour repopulation abrogates bladder cancer
chemoresistance. Nature. 517:209–213. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang G, Panigrahy D, Hwang SH, Yang J,
Mahakian LM, Wettersten HI, Liu JY, Wang Y, Ingham ES, Tam S, et
al: Dual inhibition of cyclooxygenase-2 and soluble epoxide
hydrolase synergistically suppresses primary tumor growth and
metastasis. Proc Natl Acad Sci USA. 111:11127–11132. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Majumder M, Xin X, Liu L, Tutunea-Fatan E,
Rodriguez-Torres M, Vincent K, Postovit LM, Hess D and Lala PK:
COX-2 induces breast cancer stem cells via EP4/PI3K/AKT/NOTCH/WNT
axis. Stem Cells. 34:2290–2305. 2016. View Article : Google Scholar : PubMed/NCBI
|