Introduction
Colorectal cancer is the third most frequent
malignant neoplasm and the fourth leading cause of cancer-related
death worldwide (1). Colorectal
cancer-related death is mainly caused by metastasis. Based on
accumulating data obtained over the past few years, the EMT plays a
crucial role in tumor metastasis (2,3). The
EMT is the process by which epithelial cells lose their epithelial
characteristics and gain mesenchymal characteristics. In this way,
the EMT endows cells with invasive and migratory properties,
prevents apoptosis and senescence, induces stem cell properties,
and contributes to immunosuppression (4–7).
Furthermore, several signaling pathways are involved in the EMT,
such as Wnt/β-catenin (8),
transforming growth factor-β (TGF-β) (9), Notch (10), and nuclear factor-κB (NF-κB)
signaling pathway (11). The
Wnt/β-catenin signaling pathway has a significant impact on the
maintenance of stem cell properties and cancer metastasis (12,13).
Additionally, the nuclear accumulation of β-catenin is a crucial
step in the activation of the Wnt signaling pathway (14). Lithium chloride (LiCl), an inhibitor
of glycogen synthase kinase 3β (GSK3β), promotes the nuclear
accumulation of β-catenin and activates the Wnt/β-catenin signaling
pathway (15,16).
G protein nucleolar 3 (GNL3) was initially
identified in the nucleoli of neural stem cells, embryonic stem
cells and several cancer cells, and it is preferentially expressed
by other stem cell-enriched populations, but is not expressed in
differentiated somatic cells (17,18).
GNL3 was initially named nucleostemin (NS) (17). GNL3 was subsequently shown to play
significant roles in regulating cell proliferation and the cell
cycle in various human cancers, such as gastric cancer, colorectal
cancer, liver cancer and other malignancies (19,20).
Furthermore, GNL3 also plays a crucial role in many other
physiological and pathological processes, such as the resistance to
apoptosis and cellular aging, cellular self-renewal, the
maintenance of stemness, the inhibition of differentiation,
ribosome biogenesis, the maintenance of genome stability and
telomere integrity, radioresistance, the maintenance of
tumor-initiating cells, and the induction of pluripotent stem cells
(21–27). Moreover, GNL3 is involved in cancer
invasion and metastasis (22,28,29).
However, the role of GNL3 in the invasion and metastasis of colon
cancer remains unclear.
This study is the first to report that GNL3 plays a
crucial role in the invasion and metastasis of colon cancer.
Moreover, GNL3 promotes the proliferation, invasion, migration, and
EMT in colon cancer cells by activating the Wnt/β-catenin signaling
pathway.
Materials and methods
Ethics statement
No human participant was involved in this study, and
the use of human tissue samples and all experimental procedures and
protocols were approved by the medical ethics review committee of
the First Affiliated Hospital of Chongqing Medical University,
Chongqing, China. Written informed consent was obtained from all
subjects prior to the publication of this study. All procedures
involving animals were performed in accordance with the guidelines
of the National Institutes of Health (NIH) regarding animal care
(Guide for the Care and Use of Laboratory Animals, Department of
Health and Human Services, NIH Publication No. 86-23, revised
1985). We made every effort to minimize the animals' suffering and
the number of animals experimented on.
Tissue specimens and
immunohistochemistry assays
In total, 130 colon cancer samples and their
corresponding normal, tumor-adjacent tissues were randomly
collected from the Department of Pathology, Chongqing Medical
University. An additional 60 fresh surgical specimens (30 colon
cancer tissues and corresponding normal, tumor-adjacent tissues)
were collected from the Department of Gastrointestinal Surgery, the
First Affiliated Hospital of Chongqing Medical University. The
fresh specimens were immediately frozen and stored in liquid
nitrogen after resection. None of the patients had been treated
before the operation.
Immunohistochemical staining was performed as
previously described (30).
Briefly, tissue sections were deparaffinized in xylene, and
antigens were retrieved by heating the tissue sections (95°C, 15
min). Then, 3% hydrogen peroxide was added to the slides and
incubated for 15 min before the slides were permeabilized with 0.5%
Triton-X-100 (Beyotime, Jiangshu, China) for 15 min. Subsequently,
5% bovine serum albumin (BSA) was used to block the slides for 30
min. Then, the slides were incubated with mouse primary antibodies
against human GNL3 (1:200; Santa Cruz Biotechnology, Dallas, TX,
USA) overnight at 4°C. Negative controls were incubated without
primary antibodies. The next day, the tissue sections were washed
three times with PBS for 5 min each and incubated with appropriate
HRP-conjugated secondary antibodies (anti-mouse IgG; Zhongshan
Jinqiao, Beijing, China). The slides were developed in
diaminobenzidine and counterstained with hematoxylin. The
percentage of positive cells was graded as follows: 0, <10%; 1,
10–30%; 2, 30–50%; and 3, >50% (31). The patients with colon cancer were
subsequently divided into either a negative expression group (a
score of 0) or a positive expression group (scores of 1–3).
Cell culture and transfection
The Caco-2, HCT-116, HT-29, LoVo, and SW620 human
colon cancer cell lines were obtained from the Type Culture
Collection of the Chinese Academy of Sciences, Shanghai, China.
HCT-116 and HT-29 cells were both maintained in RPMI-1640 medium
(RPMI-1640, Gibco, Gaithersburg, MD, USA) supplemented with 10%
fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin
(Beyotime) in a humidified atmosphere containing 95% air and 5%
CO2 at 37°C.
GNL3 knockdown plasmids were purchased from
Sigma-Aldrich (St. Louis, MO, USA; TRCN0000293740) and GNL3
expression plasmids were purchased from Sino Biological Inc.
(Beijing, China; HG12415-UT). The sequences of the GNL3-specific
shRNAs used in this study were: GNL3-shRNA,
5′-TTCATCGTATCTCCACTTAAT-3′ and control-shRNA,
5′-AAGCAATACCAGCAATTCATA-3′. Cells were transfected with plasmids
using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). HCT-116
cells were transfected with the GNL3 knockdown plasmids and HT-29
cells were transfected with the GNL3 expression plasmids according
to the manufacturer's protocols. Briefly, cells (at densities of
3.0×105 and 2.0×105 cells/well, respectively)
were seeded in 6-well plates in 2 ml of antibiotic-free growth
medium. The cells were transfected on the day after plating using
10 µl of Lipofectamine and 4 µg of plasmid DNA; the cells were
incubated with the transfection mixture for 6 h. Then, the cells
were transferred to 2 ml of complete growth medium at 37°C in a 5%
CO2 incubator. After 48 h of incubation, the cells were
selected with the corresponding concentrations of puromycin (Qcbio
S&T Co., Shanghai, China) and hygromycin B (Qcbio S&T Co.).
The cells were collected for protein analysis and other analyses
after a reasonable time.
Cell proliferation and colony
formation assays
HCT-116 and HT-29 cells were seeded in triplicate in
96-well plates containing 100 µl of medium at densities of
3×103 cells and collected for the MTT assay at time
points ranging from 1–5 days. The cells were treated with 20 µl of
MTT (5 mg/ml; Sigma-Aldrich) for 4 h, after which the MTT solution
was removed. The crystals were dissolved in 200 µl of DMSO
(Sigma-Aldrich). The absorbance of the solution was measured at 570
nm using a spectrophotometer.
HCT-116 and HT-29 cells were treated with 0.25%
trypsin and dispersed into a single cell suspension in medium
supplemented with 10% FBS (Gibco) prior to use. Cells were seeded
into culture dishes at a density of 200 cells/dish and cultured in
a 5% CO2 atmosphere at 37°C for 2–3 weeks. When cell
clones appeared, they were fixed with 4% paraformaldehyde for 10–30
min. Then, the cells were stained with Giemsa (Beyotime) for 10–30
min and counted under a light microscope.
Cell invasion and migration
assays
Three hundred microliters of preheated serum-free
medium was added to a cell invasion chamber (Millipore, Billerica,
MA, USA) to moisten the extracellular matrix (ECM) membrane. The
serum-free medium was removed after 1–2 h. Five hundred microliters
of medium supplemented with 10% FBS was added to each well of a
24-well plate that did not contain a chamber. Then, the chamber was
placed in the well and 2×105 cells were seeded in the
chamber (suspended in serum-free medium). After the cells were
incubated in a 5% CO2 atmosphere at 37°C for 24 h, they
were stained with 0.1% crystal violet for 10 min. Cells were
counted under a light microscope, and images were captured.
HCT-116 and HT-29 cells were separately seeded in a
6-well plate in serum-free medium. Cells grew overnight to
confluence. A pipette tip was dragged through the cells to scratch
the monolayer, creating a wound with an even width. Then, PBS was
used to wash and remove the cellular debris. Cells were allowed to
migrate for 24 h. An inverted microscope was used to capture images
at 0 and 24 h.
Western blot analysis
Western blot analyses were performed as previously
described (32). Briefly, the
proteins were extracted and their concentrations were determined
using a protein extraction kit (Beyotime) and an enhanced
bicinchoninic acid (BCA) protein assay kit (Beyotime),
respectively, according to the manufacturer's instructions. Protein
samples were separated by SDS-PAGE and transferred to PVDF
membranes (Millipore). The transferred PVDF membranes were blocked
with 5% non-fat dry milk in PBST for 1.5 h and incubated with
primary antibodies against GNL3 (1:200 dilution; Santa Cruz
Biotechnology), E-cadherin (1:1,000 dilution; Santa Cruz
Biotechnology), N-cadherin (1:1,000 dilution; Epitomics,
Burlingame, CA, USA), vimentin (1:1,000 dilution; eBioscience, San
Diego, CA, USA), and β-catenin (1:5,000 dilution; Epitomics)
overnight at 4°C. Then, the membranes were incubated with the
appropriate secondary antibodies for 1 h. Bands were visualized
with an enhanced chemiluminescence kit (Beyotime) and quantitated
with Quantity One 4.6.2 software. Protein band intensities were
normalized to the intensity of GAPDH (1:500 dilution;
Beyotime).
Immunofluorescence staining
Cells were seeded in plates and fixed with 4%
paraformaldehyde for 15 min. The cells were permeabilized with 0.5%
TritonX-100 (Beyotime) for 10 min at room temperature. Then, the
cells were blocked with 1% bovine serum albumin (BSA) for 2 h.
Afterwards, the cells were incubated with primary antibodies
overnight at 4°C. The next day, the appropriate secondary
antibodies were incubated with the cells for 2 h in the dark at
4°C, and 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich) was
added to stain the nucleus. Finally, the cells were visualized, and
images were captured using a fluorescence microscope (Leica,
Wetzlar, Germany).
In vivo experiments
Five-week-old male nude BALB/c mice were purchased
from the National Biological Industry Base, Laboratory Animal
Center of Chongqing Medical University. Approximately
5×106 HCT-116 and HT-29 cells that had been transfected
with the GNL3 knockdown plasmids and GNL3 expression plasmids,
respectively, were subcutaneously injected into the axillae of nude
mice. The sizes of the xenograft tumors were measured weekly. Four
weeks later, the nude mice were euthanized and tumor samples were
collected.
Statistical analysis
The data are presented as the means ± standard
deviations (SD). The expression levels of GNL3 and
clinicopathological features were compared using the Chi-square
test. The differences between groups were analyzed using Student's
t-test using SPSS 13.0 (SPSS, Inc., Chicago, IL, USA). A
Kaplan-Meier analysis was used to obtain the survival curves, and
differences were assessed with the log-rank test. P<0.05 was
considered to indicate a statistically significant difference.
Results
The correlation between GNL3
expression and clinicopathological characteristics in colon
cancer
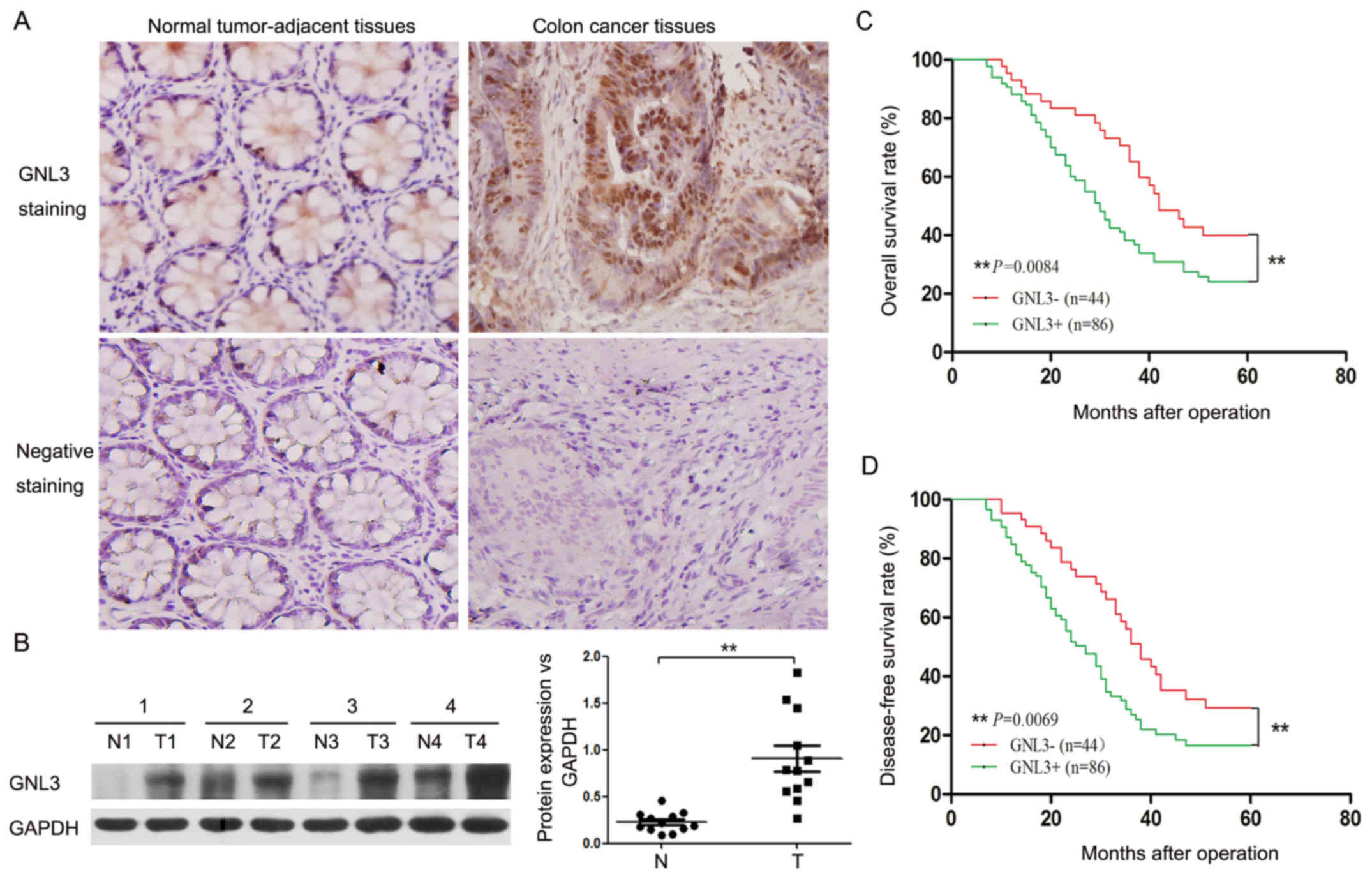
We collected tissue specimens from 130 patients and
performed immunohistochemistry assays on these specimens to
determine the GNL3 expression level in colon cancer. The rates of
GNL3-positive expression in colon cancer tissues and normal,
tumor-adjacent tissues were 66.2% (86/130) and 31.5% (41/130),
respectively. As expected, GNL3 was expressed at much higher levels
in colon cancer tissues than in normal, tumor-adjacent tissues
(Fig. 1A). Furthermore, we examined
the expression levels of GNL3 in the fresh samples using western
blotting. GNL3 was expressed at significantly higher levels in
colon cancer tissues than in normal, tumor-adjacent tissues
(P<0.01; Fig. 1B). The
correlations between the GNL3 expression levels and the
clinicopathological features of patients with colon cancer are
presented in Table I. The GNL3
expression levels were significantly associated with tumor
differentiation (P<0.01), serosal invasion (P<0.05), lymph
node metastasis (P<0.05), and vascular invasion (P<0.05) in
patients with colon cancer (P<0.05) (Table I).
 | Table I.The correlations between the GNL3
expression levels and the clinicopathological features of patients
with colon cancer. |
Table I.
The correlations between the GNL3
expression levels and the clinicopathological features of patients
with colon cancer.
|
|
| GNL3 |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | Cases n=130 | Negative n=44 | Positive n=86 | P-value |
|---|
| Sex |
|
|
| 0.5169 |
|
Male | 76 | 24 | 52 |
|
|
Female | 54 | 20 | 34 |
|
| Age (years) |
|
|
| 0.4647 |
|
<60 | 53 | 16 | 37 |
|
|
≥60 | 77 | 28 | 49 |
|
| Tumor size
(cm) |
|
|
| 0.1059 |
| ≤3
cm | 61 | 25 | 36 |
|
| >3
cm | 69 | 19 | 50 |
|
|
Differentiation |
|
|
| 0.0043b |
|
Poor | 58 | 11 | 47 |
|
|
Moderate | 36 | 18 | 18 |
|
|
Well | 36 | 15 | 21 |
|
| Serosal
invasion |
|
|
| 0.0176a |
|
Absent | 58 | 26 | 32 |
|
|
Present | 72 | 18 | 54 |
|
| Vascular
invasion |
|
|
| 0.0468a |
|
Absent | 61 | 26 | 35 |
|
|
Present | 69 | 18 | 51 |
|
| Lymph node
metastasis |
|
|
| 0.0353a |
|
Yes | 84 | 23 | 61 |
|
| No | 46 | 21 | 25 |
|
| Liver
metastasis |
|
|
| 0.0830 |
|
Absent | 94 | 36 | 58 |
|
|
Present | 36 | 8 | 28 |
|
| Stage |
|
|
| 0.1559 |
| I +
II | 51 | 21 | 30 |
|
| III +
IV | 79 | 23 | 56 |
|
A Kaplan-Meier survival analysis was used to
determine the correlation between the GNL3 expression levels and
the survival of patients with colon cancer as a measure of the
prognostic significance of GNL3 expression in these patients. As
shown in Fig. 1C and D, the 5-year
survival rate of the GNL3-positive groups was markedly reduced
compared with that in the GNL3-negative groups (P<0.01; Fig. 1C). The median disease-free survival
times were 27.0 and 38.0 months for the GNL3-positive and
GNL3-negative groups, respectively (P<0.01; Fig. 1D). Based on these results, the
upregulation of GNL3 expression was closely related to a shorter
disease-free survival (P<0.01; Fig.
1D).
GNL3 expression levels in colon cancer
cell lines
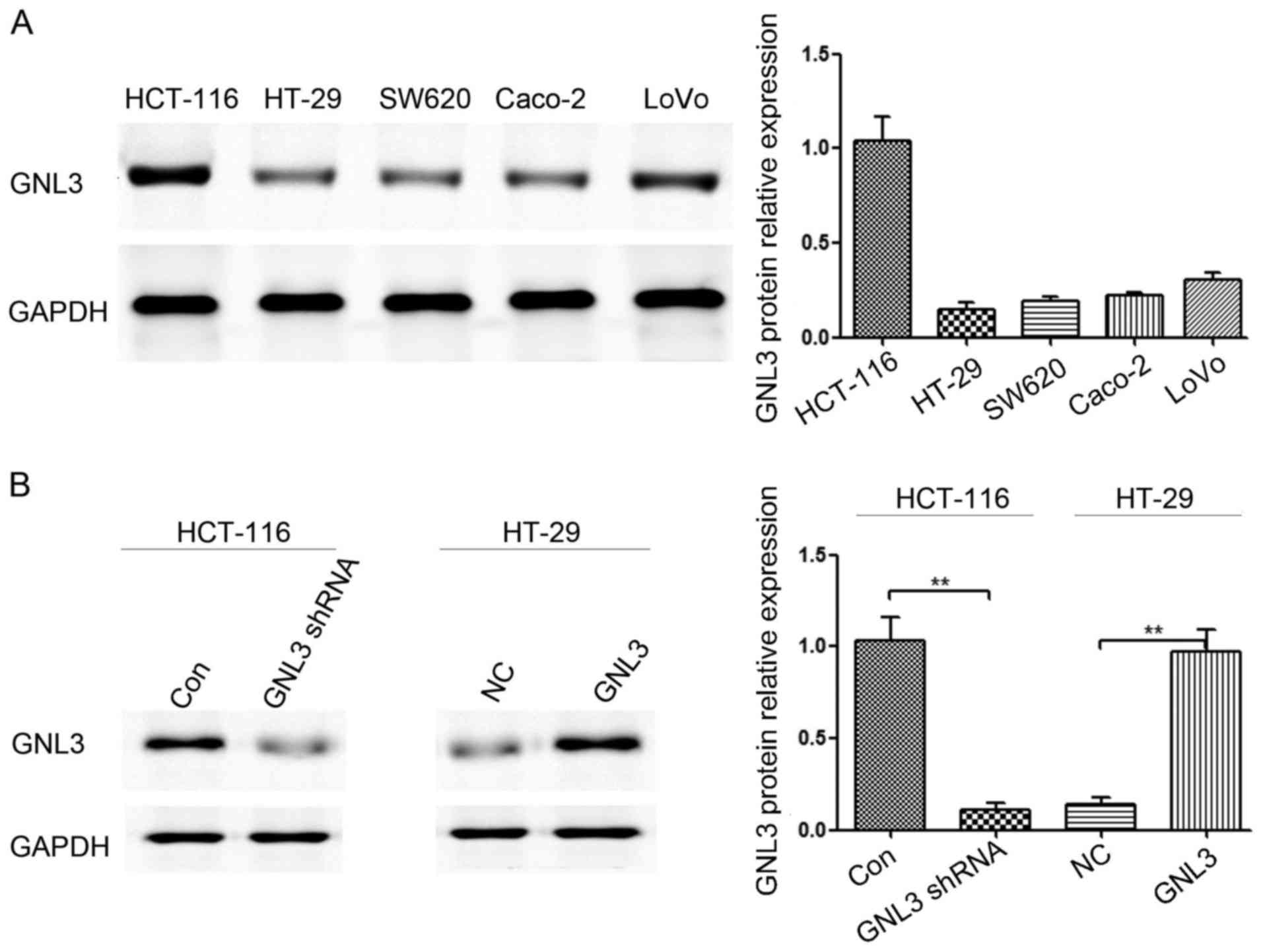
Western blot assays were used to detect the GNL3
levels in five colon cancer cell lines, and the results showed
different levels of GNL3 expression in Caco-2 (0.22±0.02), HCT-116
(1.04±0.13), LoVo (0.31±0.05), HT-29 (0.15±0.04), and SW620
(0.19±0.02) cells (Fig. 2A).
Therefore, we selected HCT-116 and HT-29 cells for transfection. We
used western blot assays to test the GNL3 expression levels in
HCT-116 and HT-29 cells that had been transfected with GNL3
knockdown and expression plasmids, respectively, as well as cells
transfected with the corresponding control plasmids. GNL3
expression was decreased in HCT-116 cells transfected with GNL3
knockdown plasmids (P<0.01; Fig.
2B) and increased in HT-29 cells transfected with GNL3
expression plasmids (P<0.01; Fig.
2B).
GNL3 promotes colon cancer cell
proliferation and colony formation
As previously reported, GNL3 plays an important role
in regulating the proliferation of various stem cells and cancer
cells. Therefore, we postulated that GNL3 plays a similar role in
regulating the proliferation of colon cancer cells. We performed
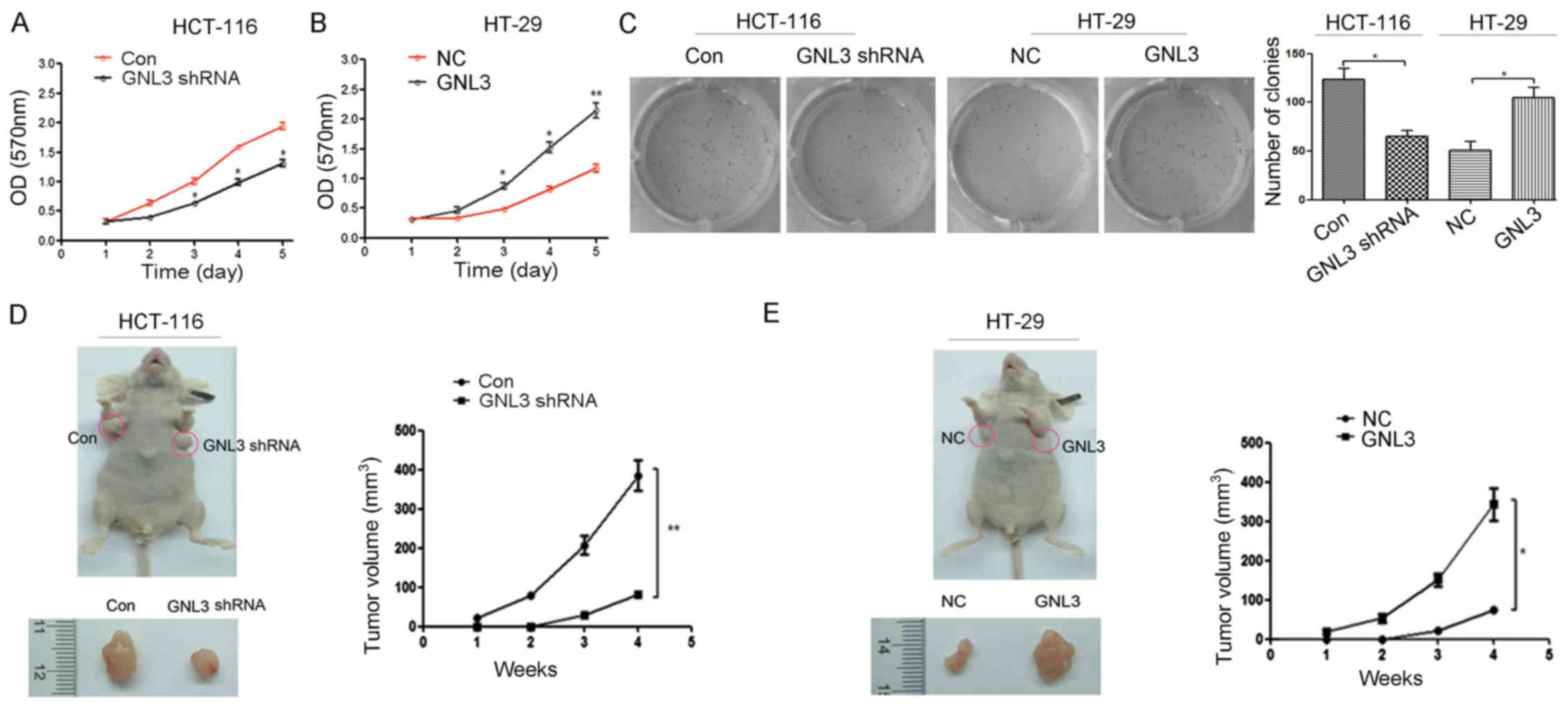
MTT cell proliferation assays to assess their proliferative
capacity. GNL3 knockdown inhibited the proliferation of HCT-116
cells (P<0.05; Fig. 3A) and GNL3
overexpression promoted the proliferation of HT-29 cells (P<0.05
or P<0.01; Fig. 3B). Colony
formation assays were performed to further examine the effects of
GNL3 on the formation of colon cancer cell colonies. Cells were
transfected with the vectors described above. As shown in Fig. 3C, GNL3 knockdown significantly
inhibited the ability of HCT-116 cells to form colonies (P<0.05;
Fig. 3C), and GNL3 overexpression
remarkably enhanced the capacity of HT-29 cells to form colonies
(P<0.05; Fig. 3C). In
conclusion, GNL3 overexpression promoted colon cancer cell
proliferation and colony formation in vitro.
GNL3 promotes tumor growth in
vivo
We further tested whether GNL3 affected
tumorigenesis in vivo using a xenograft model in nude mice.
Nude mice were transplanted with HCT-116 and HT-29 cells that had
been transfected with the vectors described above. Four weeks
later, the xenograft tumors in the HCT-116 knockdown groups were
smaller than the tumors in the control groups (Fig. 3D), and the tumors in the HT-29
overexpression groups were larger than the tumors in the
corresponding controls (Fig. 3E).
Moreover, the average tumor volume exhibited similar trends in the
GNL3 knockdown groups (P<0.01; Fig.
3D) and GNL3 overexpression groups (P<0.05; Fig. 3E) as those in the corresponding
control groups. Thus, GNL3 promoted tumor growth in
vivo.
GNL3 promotes the invasion and
migration of colon cancer cells
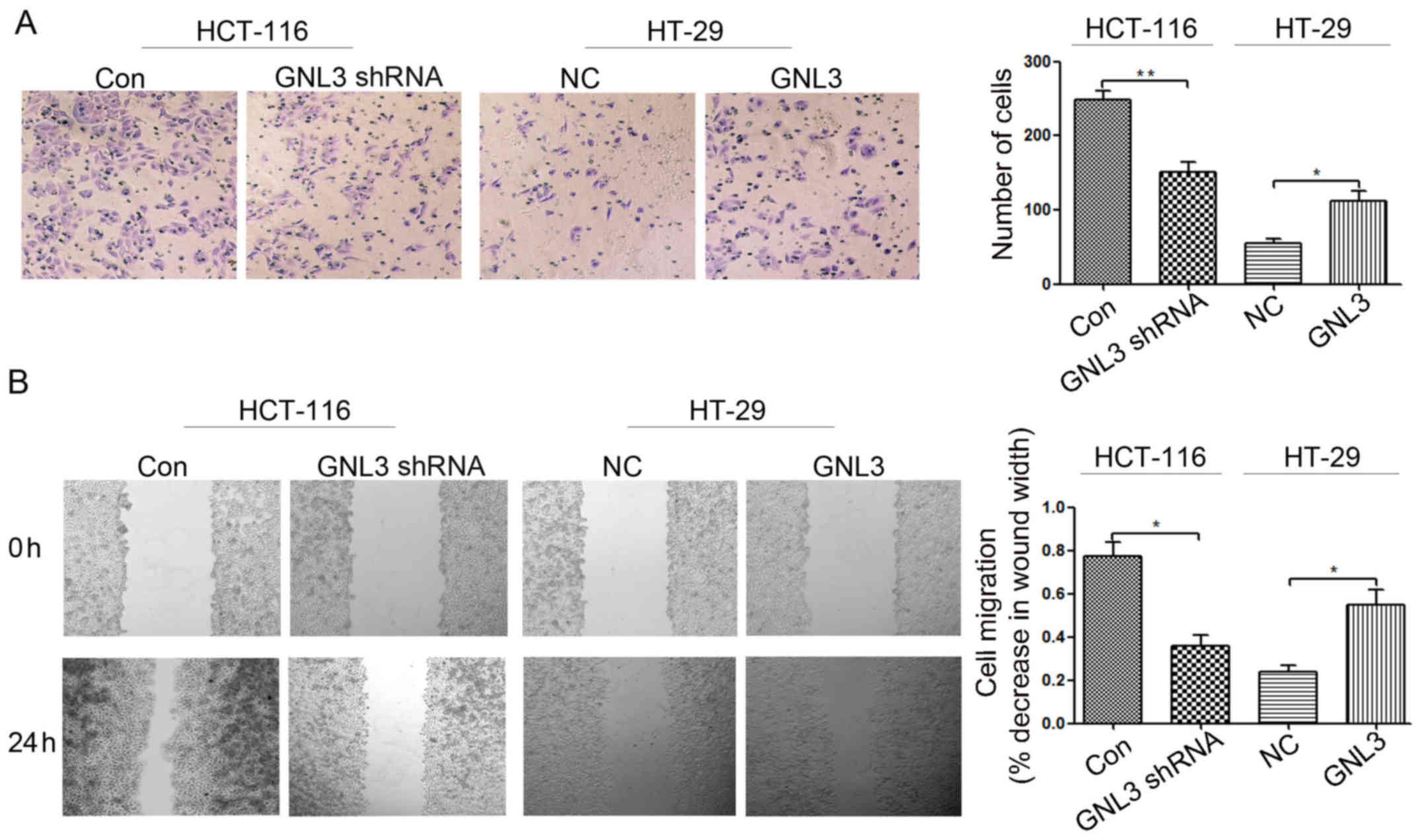
Cell invasion assays were performed using cell
invasion chambers to assess the effects of GNL3 on colon cancer
cell invasion. Cells that migrated through the ECM membrane are
shown in Fig. 4A. Following GNL3
knockdown, the invasion of HCT-116 cells was significantly
inhibited compared with the paired control groups (P<0.01;
Fig. 4A). However, GNL3
overexpression significantly increased the invasion of HT-29 cells
compared with the corresponding control groups (P<0.05; Fig. 4A). In vitro scratch
wound-healing assays were performed to assess the effects of GNL3
on colon cancer cell migration. As shown in Fig. 4B, cells in which GNL3 expression was
knocked down migrated significantly slower than the paired control
groups (P<0.05; Fig. 4B).
However, GNL3 overexpression induced significantly faster migration
in the transfected cells than in the corresponding control groups
(P<0.05; Fig. 4B). Based on
these data, GNL3 promotes the invasion and migration of colon
cancer cells.
GNL3 promotes EMT in colon cancer
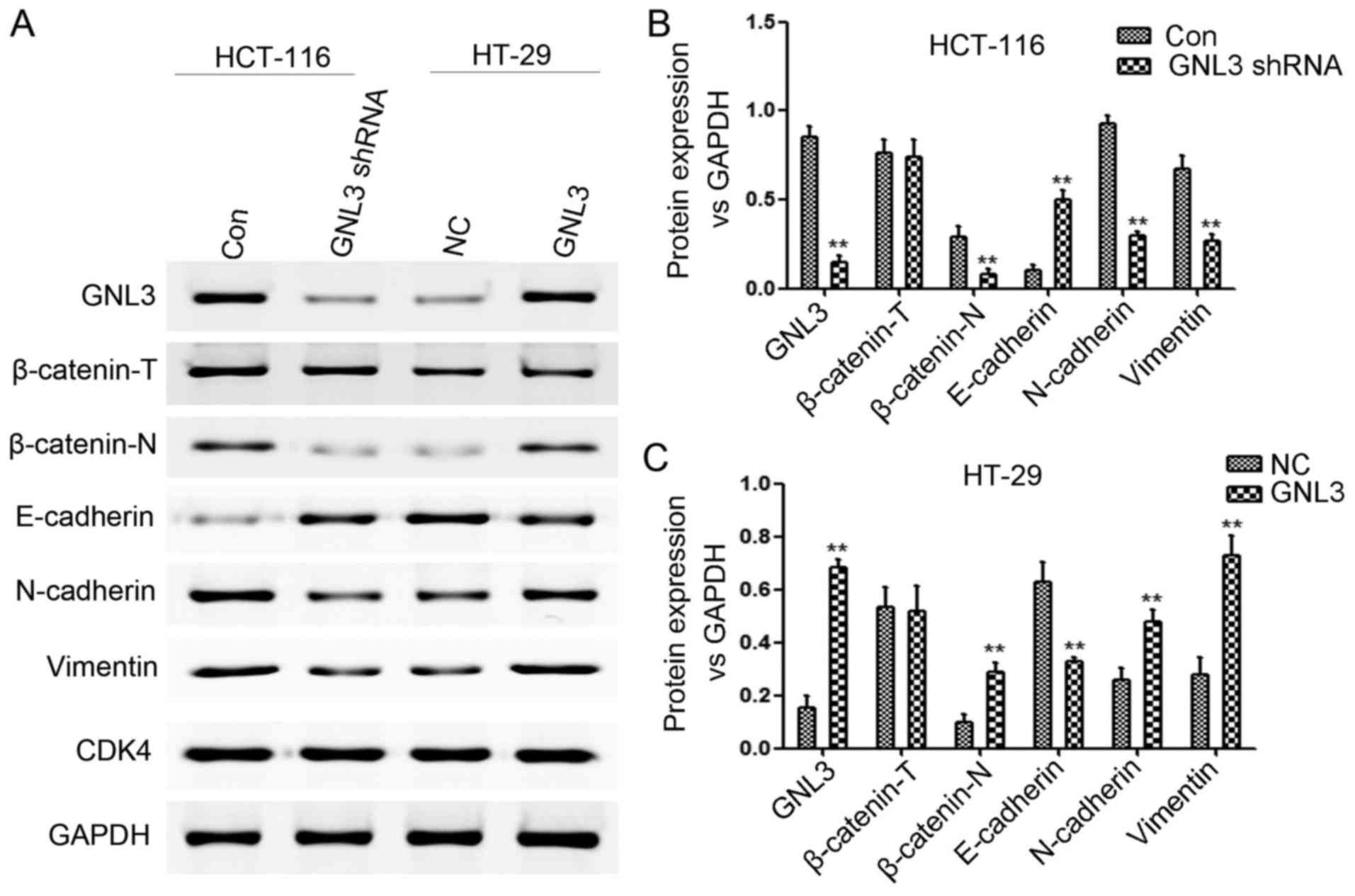
cells by activating the Wnt/β-catenin signaling pathway
We performed western blot assays to quantitate the
expression levels of total β-catenin (β-catenin-T), nuclear
β-catenin (β-catenin-N), GNL3, E-cadherin, N-cadherin, and vimentin
in HCT-116 and HT-29 cells transfected with GNL3 knockdown vectors
or GNL3 expression vectors, respectively, or paired control vectors
to confirm that GNL3 promoted EMT in colon cancer cells by
activating the Wnt/β-catenin signaling pathway. GNL3 knockdown in
HCT-116 cells increased E-cadherin expression (P<0.01) and
decreased β-catenin-N (P<0.01), GNL3 (P<0.01), N-cadherin
(P<0.01) and vimentin expression (P<0.01) (Fig. 5A and B). However, GNL3
overexpression in HT-29 cells decreased E-cadherin expression
(P<0.01) and increased β-catenin-N (P<0.01), GNL3
(P<0.01), N-cadherin (P<0.01) and vimentin expression
(P<0.01) (Fig. 5A and C). No
significant differences in the β-catenin-T levels were observed in
each group (P>0.05; Fig.
5A-C).
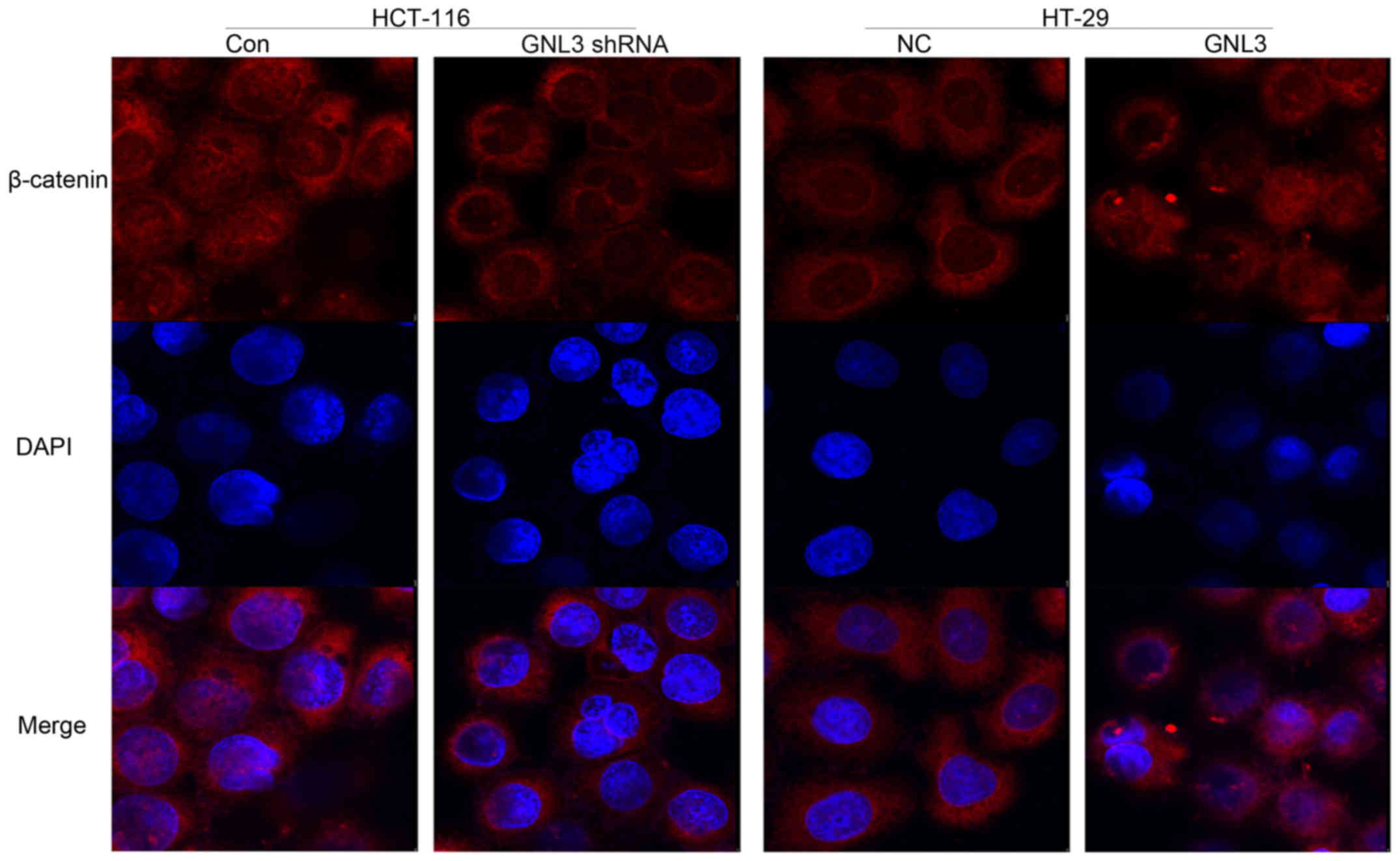
Moreover, nuclear β-catenin immunofluorescence
staining was reduced upon GNL3 knockdown in HCT-116 cells and
increased upon GNL3 overexpression in HT-29 cells (Fig. 6).
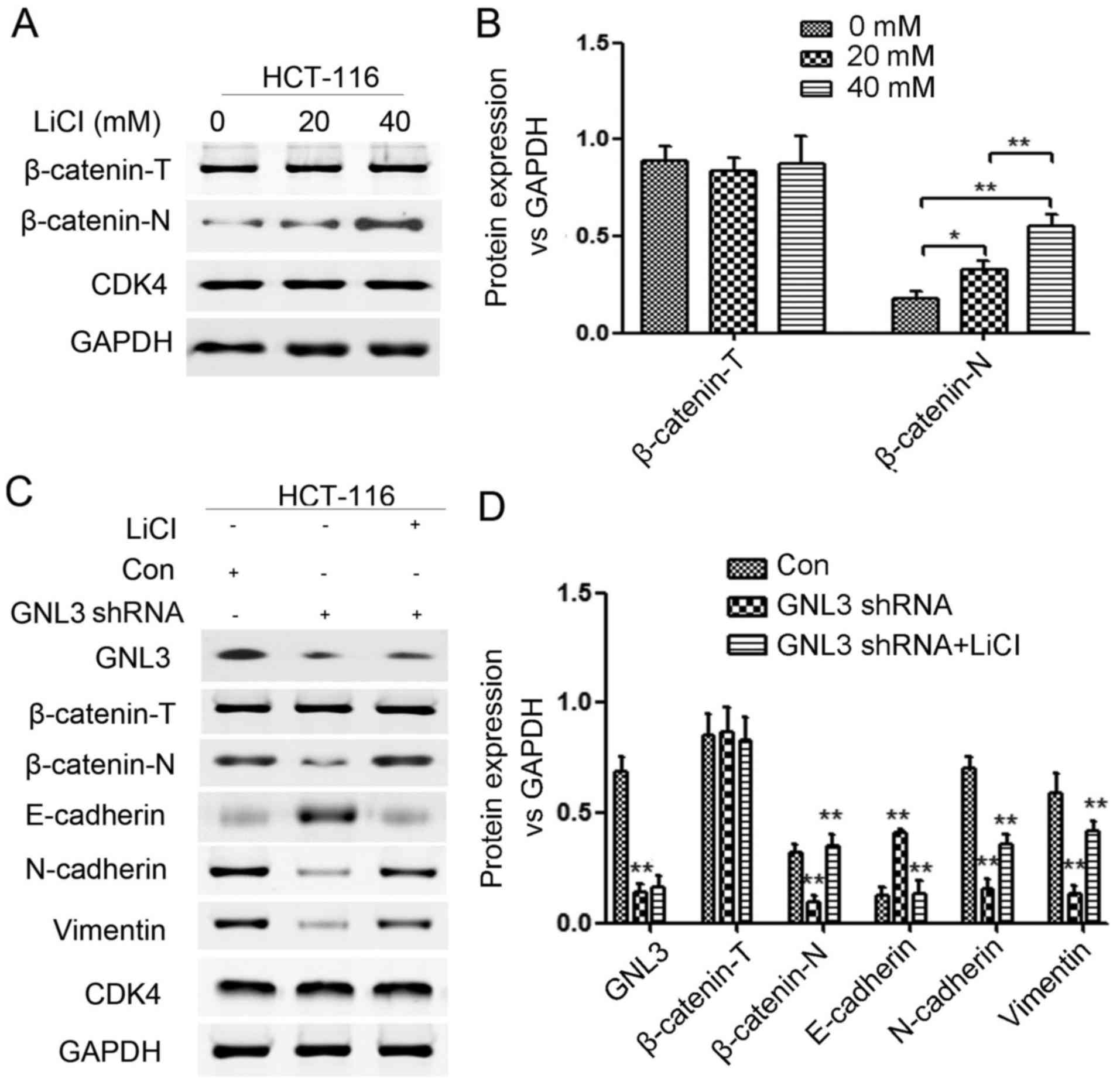
We further evaluated whether the modulatory effects
of GNL3 on EMT in colon cancer cells were attributable to the
Wnt/β-catenin signaling pathway. LiCl was used to promote the
nuclear accumulation of β-catenin (β-catenin-N) and activate the
Wnt/β-catenin signaling pathway in HCT-116 cells transfected with
GNL3 knockdown plasmids. The control group (Con), GNL3 knockdown
group (GNL3 shRNA), and LiCl-treated GNL3 knockdown group (GNL3
shRNA+LiCl) were evaluated. Western blot assays were performed to
examine the expression levels of proteins related to the EMT and
Wnt/β-catenin signaling pathway. First, we determined the
appropriate concentration of LiCl needed to activate the
Wnt/β-catenin signaling pathway and found that 20 mM (P<0.05)
and 40 mM (P<0.01) were suitable concentrations (Fig. 7A and B). We found that β-catenin-N,
GNL3, N-cadherin and vimentin were all remarkably downregulated and
E-cadherin was upregulated in GNL3 knockdown groups compared with
the levels in the control groups (P<0.01; Fig. 7C and D). We also found that
β-catenin-N, N-cadherin and vimentin expression were all
upregulated and E-cadherin expression was downregulated in the GNL3
shRNA+LiCl groups compared with the levels in the GNL3 shRNA groups
(P<0.01; Fig. 7C and D). The
inhibition of the EMT and Wnt/β-catenin signaling pathway caused by
GNL3 knockdown was partially reversed by LiCl. In other words, GNL3
actually regulated EMT in colon cancer cells by activating the
Wnt/β-catenin signaling pathway; otherwise, LiCl would not have
partially restored the activity of the suppressed Wnt/β-catenin
signaling pathway and EMT. In summary, GNL3 promotes the EMT in
colon cancer cells by activating Wnt/β-catenin signaling
pathway.
Discussion
Metastasis is the main cause of death in patients
with colon cancer. The EMT plays a crucial role in tumor metastasis
(33). Although GNL3 has been shown
to be involved in cancer metastasis, the role of GNL3 in colon
cancer metastasis remains unclear (22,29,34).
Herein, GNL3 was expressed at much higher levels in colon cancer
tissues than in normal, tumor-adjacent tissues, and GNL3 levels
were related to the prognosis of patients with colon cancer. GNL3
promotes the proliferation, invasion, migration and EMT in colon
cancer cells. Furthermore, GNL3 promotes EMT in colon cancer by
activating the Wnt/β-catenin signaling pathway. These results are
consistent with our previous hypotheses.
We used immunohistochemistry and western blot assays
to determine the GNL3 expression levels in 130 colon cancer tissue
specimens as a first step in elucidating the function and mechanism
by which GNL3 regulates colon cancer metastasis. GNL3 was expressed
at significantly higher levels in tumor tissues than in normal,
tumor-adjacent tissues. The GNL3 expression levels were closely
related to tumor differentiation, serosal invasion, lymph node
metastasis and vascular invasion. Moreover, high GNL3 expression
levels resulted in lower 5-year and disease-free survival rates for
patients with colon cancer. These results are consistent with the
findings of a previous study (29).
In hepatocellular carcinoma (HCC), GNL3 expression levels are also
closely related to tumor differentiation and metastasis, and high
GNL3 expression levels predict a worse prognosis for patients with
HCC. Based on these results, high GNL3 expression levels show
significant associations with the clinicopathological
characteristics and predict a poorer prognosis for patients with
colon cancer.
Next, we tested the effects of GNL3 on colon cancer
cell proliferation and colony formation. GNL3 overexpression
increased cell proliferation and colony formation; GNL3 knockdown
exerted the opposite effect. Furthermore, we used a xenograft model
in nude mice and found that the xenograft tumors in the
GNL3-overexpressing groups were larger than the tumors in the
control groups. In contrast, GNL3 knockdown groups exhibited the
opposite trend. These data were similar to those in previously
published studies. As shown in previous studies (17,29),
GNL3 regulates cell proliferation in p53-dependent and
p53-independent manner. The Wnt/β-catenin signaling pathway is
involved in regulating cell proliferation (35). Moreover, GNL3 was recently shown to
promote the proliferation of human glioma through the Wnt/β-catenin
pathway (36).
Then, we examined the effects of GNL3 on the
invasion and migration of colon cancer cells using cell invasion
assays and scratch wound-healing assays. GNL3 overexpression
promoted the invasion and migration of colon cancer cells, whereas
GNL3 knockdown inhibited invasion and migration. Then, we tested
the expression levels of EMT-related markers using western blot
assays and showed that GNL3 overexpression reduced E-cadherin
expression and increased N-cadherin and vimentin expression. In
contrast, GNL3 knockdown exerted the opposite effects. These data
are similar to the results from a previous study (22). GNL3 overexpression activates EMT via
the TGF-β signaling pathway to induce HeLa cell metastasis
(22). However, the TGF-β signaling
pathway is different from the pathway examined in our study.
Because of the complexity of the EMT, the EMT utilizes several
signaling pathways, as we previously described. Different tumor
cells may utilize different signaling pathways to induce EMT.
Finally, we performed western blot analyses and
immunofluorescence staining to evaluate the levels and localization
of β-catenin in colon cancer cells and to clarify the mechanism by
which GNL3 regulates EMT in colon cancer cells. Furthermore, we
used LiCl to reverse the GNL3 knockdown-induced suppression of the
EMT and Wnt/β-catenin signaling pathway. Then, GNL3 over-expression
increased the β-catenin-N levels, and GNL3 knockdown reduced the
β-catenin-N levels. Moreover, the suppression of EMT and
Wnt/β-catenin signaling pathway in the GNL3 shRNA groups was
partially restored by the LiCl treatment. Based on these results,
GNL3 induces the nuclear translocation of β-catenin, thereby
activating the Wnt/β-catenin signaling pathway and inducing EMT in
colon cancer cells. β-catenin may be located downstream of GNL3,
and we conclude that GNL3 promotes the EMT in colon cancer by
activating the Wnt/β-catenin signaling pathway.
In conclusion, GNL3 promotes EMT by activating the
Wnt/β-catenin signaling pathway and thus plays an important role in
the invasion and metastasis of colon cancer. Our findings not only
expand our understanding of the molecular mechanisms underlying
GNL3 function and the invasion and metastasis of colon cancer but
also suggest a potential target or strategy for treating colon
cancer.
Acknowledgements
This study was supported by the National Key
Clinical Specialties Construction Program of China ([No. 2012]649)
and the National Natural Science Foundation of China (No.
81272753).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nieto MA: Epithelial-mesenchymal
transitions in development and disease: Old views and new
perspectives. Int J Dev Biol. 53:1541–1547. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang Y, Liu Q, Zhang H, Zhao H, Mao R, Li
Z, Ya S, Jia C and Bao Y: Silencing of GP73 inhibits invasion and
metastasis via suppression of epithelial-mesenchymal transition in
hepatocellular carcinoma. Oncol Rep. 37:1182–1188. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chaudhury A, Hussey GS, Ray PS, Jin G, Fox
PL and Howe PH: TGF-beta-mediated phosphorylation of hnRNP E1
induces EMT via transcript-selective translational induction of
Dab2 and ILEI. Nat Cell Biol. 12:286–293. 2010.PubMed/NCBI
|
|
10
|
Timmerman LA, Grego-Bessa J, Raya A,
Bertrán E, Pérez-Pomares JM, Díez J, Aranda S, Palomo S, McCormick
F, Izpisúa-Belmonte JC, et al: Notch promotes
epithelial-mesenchymal transition during cardiac development and
oncogenic transformation. Genes Dev. 18:99–115. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huber MA, Azoitei N, Baumann B, Grünert S,
Sommer A, Pehamberger H, Kraut N, Beug H and Wirth T: NF-kappaB is
essential for epithelial-mesenchymal transition and metastasis in a
model of breast cancer progression. J Clin Invest. 114:569–581.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leung CO, Mak WN, Kai AK, Chan KS, Lee TK,
Ng IO and Lo RC: Sox9 confers stemness properties in hepatocellular
carcinoma through Frizzled-7 mediated Wnt/β-catenin signaling.
Oncotarget. 7:29371–29386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yuan X, Sun X, Shi X, Wang H, Wu G, Jiang
C, Yu D, Zhang W, Xue B and Ding Y: USP39 promotes colorectal
cancer growth and metastasis through the Wnt/β-catenin pathway.
Oncol Rep. 37:2398–2404. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mao Y, Xu J, Li Z, Zhang N, Yin H and Liu
Z: The role of nuclear β-catenin accumulation in the Twist2-induced
ovarian cancer EMT. PLoS One. 8:e782002013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Azzolin L, Panciera T, Soligo S, Enzo E,
Bicciato S, Dupont S, Bresolin S, Frasson C, Basso G, Guzzardo V,
et al: YAP/TAZ incorporation in the β-catenin destruction complex
orchestrates the Wnt response. Cell. 158:157–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stambolic V, Ruel L and Woodgett JR:
Lithium inhibits glycogen synthase kinase-3 activity and mimics
wingless signalling in intact cells. Curr Biol. 6:1664–1668. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsai RY and McKay RD: A nucleolar
mechanism controlling cell proliferation in stem cells and cancer
cells. Genes Dev. 16:2991–3003. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tsai RY: Turning a new page on
nucleostemin and self-renewal. J Cell Sci. 127:3885–3891. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu SJ, Cai ZW, Liu YJ, Dong MY, Sun LQ,
Hu GF, Wei YY and Lao WD: Role of nucleostemin in growth regulation
of gastric cancer, liver cancer and other malignancies. World J
Gastroenterol. 10:1246–1249. 2004.PubMed/NCBI
|
|
20
|
Zia-Jahromi N, Hejazi SH, Panjepour M,
Parivar K and Gharagozloo M: Comparison of nucleostemin gene
expression in CD133+ and CD133− cell
population in colon cancer cell line HT29. J Cancer Res Ther.
10:68–72. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meng L, Lin T, Peng G, Hsu JK, Lee S, Lin
SY and Tsai RY: Nucleostemin deletion reveals an essential
mechanism that maintains the genomic stability of stem and
progenitor cells. Proc Natl Acad Sci USA. 110:11415–11420. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Okamoto N, Yasukawa M, Nguyen C, Kasim V,
Maida Y, Possemato R, Shibata T, Ligon KL, Fukami K, Hahn WC, et
al: Maintenance of tumor initiating cells of defined genetic
composition by nucleostemin. Proc Natl Acad Sci USA.
108:20388–20393. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin T, Ibrahim W, Peng CY, Finegold MJ and
Tsai RY: A novel role of nucleostemin in maintaining the genome
integrity of dividing hepatocytes during mouse liver development
and regeneration. Hepatology. 58:2176–2187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meng L, Lin T and Tsai RY: Nucleoplasmic
mobilization of nucleostemin stabilizes MDM2 and promotes G2-M
progression and cell survival. J Cell Sci. 121:4037–4046. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tsai RY and McKay RD: A multistep,
GTP-driven mechanism controlling the dynamic cycling of
nucleostemin. J Cell Biol. 168:179–184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamashita M, Nitta E, Nagamatsu G,
Ikushima YM, Hosokawa K, Arai F and Suda T: Nucleostemin is
indispensable for the maintenance and genetic stability of
hematopoietic stem cells. Biochem Biophys Res Commun. 441:196–201.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu Q, Yasumoto H and Tsai RY:
Nucleostemin delays cellular senescence and negatively regulates
TRF1 protein stability. Mol Cell Biol. 26:9279–9290. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang G, Zhang Q, Zhang Q, Yin L, Li S,
Cheng K, Zhang Y, Xu H and Wu W: Expression of nucleostemin,
epidermal growth factor and epidermal growth factor receptor in
human esophageal squamous cell carcinoma tissues. J Cancer Res Clin
Oncol. 136:587–594. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu B, Hua L, Ni W, Wu M, Yan D, Chen Y, Lu
C, Chen B and Wan C: Nucleostemin/GNL3 promotes nucleolar
polyubiquitylation of p27(kip1) to drive hepatocellular carcinoma
progression. Cancer Lett. 388:220–229. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li W, Wang Z, Zha L, Kong D, Liao G and Li
H: HMGA2 regulates epithelial-mesenchymal transition and the
acquisition of tumor stem cell properties through TWIST1 in gastric
cancer. Oncol Rep. 37:185–192. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu MZ, Yao TJ, Lee NPY, Ng IO, Chan YT,
Zender L, Lowe SW, Poon RT and Luk JM: Yes-associated protein is an
independent prognostic marker in hepatocellular carcinoma. Cancer.
115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zha L, Zhang J, Tang W, Zhang N, He M, Guo
Y and Wang Z: HMGA2 elicits EMT by activating the Wnt/β-catenin
pathway in gastric cancer. Dig Dis Sci. 58:724–733. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee M, Williams KA, Hu Y, Andreas J, Patel
SJ, Zhang S and Crawford NP: GNL3 and SKA3 are novel prostate
cancer metastasis susceptibility genes. Clin Exp Metastasis.
32:769–782. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xie Y and Wang B: Downregulation of
TNFAIP2 suppresses proliferation and metastasis in esophageal
squamous cell carcinoma through activation of the Wnt/β-catenin
signaling pathway. Oncol Rep. 37:2920–2928. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bao Z, Wang Y, Yang L, Wang L, Zhu L, Ban
N, Fan S, Chen W, Sun J, Shen C, et al: Nucleostemin promotes the
proliferation of human glioma via Wnt/beta-Catenin pathway.
Neuropathology. 36:237–249. 2016. View Article : Google Scholar : PubMed/NCBI
|