Introduction
Lung cancer is a common malignant tumor worldwide,
and its incidence and mortality rates have significantly increased
in recent years (1,2). Since the clinical manifestations of
early lung cancer are often hidden and lack specificity, most
patients are not diagnosed until the disease has reached an
advanced stage (3). At present, for
patients with late-stage lung cancer, the main treatment strategy
is chemotherapy or chemotherapy combined with radiotherapy
(4,5). Chemotherapy can kill tumor cells, but
it can also exert strong side effects on normal cells (6). Furthermore, after multiple cycles of
chemotherapy, tumor cells can develop resistance to
chemotherapeutic drugs. Multidrug resistance (MDR) is the acquired
resistance of cancer cells to structurally and functionally
different chemotherapeutic drugs (7), and remains a major obstacle to
chemotherapy efficacy, decreasing the effectiveness of treatment
for patients with lung cancer (8,9). Thus,
it is important to investigate the mechanisms related to MDR and to
improve the efficacy of chemotherapy for lung cancer.
Many studies have shown that the MDR mechanism of
tumors is mainly related to the ATP-binding cassette (ABC)
transporter superfamily of genes, such as P-glycoprotein (P-gp;
encoded by the MDR1 gene), which code for efflux pump proteins
(10,11). P-gp overexpression can increase the
rate at which drugs are pumped out of cells, which reduces the
chemotherapeutic effects of the drugs, inducing drug resistance
(12,13). Furthermore, tumor MDR mechanisms are
associated with detoxification, repair and various signal
transduction pathways (14).
Generally, the drug resistance mechanisms in lung cancer are
complex, involving many factors; therefore, it is considered highly
important to identify efficient, low-toxicity methods of reversing
lung cancer MDR.
Gene therapy has introduced new prospects for the
treatment of drug resistance in cancer. Interleukin 24 (IL-24), a
newly identified antitumor gene, can inhibit the growth of tumor
cells, including lung, breast and ovarian cancer cells, and is
considered to be combinable with radiotherapy or chemotherapy,
which could improve the effects of radiotherapy and chemotherapy on
tumor cells (15–17). The IL-24 gene has been widely
investigated in various cancers for its role as a tumor-suppressor
gene, particularly with regard to tumor gene therapy (18); however, its role in reversing MDR
has not been investigated in detail. In the present study, we used
adenovirus-mediated human IL-24 gene (Ad-hIL-24) transfection and
the cisplatin (DDP)-resistant human lung adenocarcinoma cell line
A549/DDP to study whether IL-24 can reverse the MDR of lung cancer
as well as investigate its mechanism. The results revealed that
Ad-hIL-24 could effectively increase the anticancer effect of DDP
on A549/DDP cells and induce A549/DDP cell apoptosis. These effects
were associated with decreases in the expression levels of
phosphorylated AKT (p-AKT) and P-gp.
Materials and methods
Adenoviral vectors, cell lines and
cell culture
The DDP-resistant human lung adenocarcinoma cell
line A549/DDP (19) was obtained
from the Central Laboratory of Xiangya Medical College of Central
South University (Hunan, China). Ad-hIL-24, an adenoviral vector
containing the hIL-24 gene, which is able to effectively express
hIL-24 (20), was acquired from the
Laboratory of Cell and Molecular Biology, Medicine College, Soochow
University (Suzhou, China). QBI-293A (a human embryonic kidney cell
line) was provided by Dr Jicheng Yang of Soochow University
(Suzhou, China). The cells were cultured in RPMI-1640 (Hyclone,
Nanjing, China), supplemented with 10% fetal bovine serum (FBS)
(Hyclone). An IL-24 enzyme-linked immunosorbent assay (ELISA) kit
was purchased from CusaBiol (Carlsbad, CA, USA; cat. #ELH-IL-24-1).
Rabbit anti-human P-gp (#129450), p-Akt (#4060), and Akt (#9272)
were purchased from Cell Signaling Technology (Shanghai, China).
Antibodies against GAPDH (#ab153802) and IL-24 (#ab182567) were
purchased from Abcam (Shanghai, China). The Cell Counting Kit-8
(CCK-8) assay was purchased from Dojindo (Nanjing, China).
Amplification of Ad-hIL-24 and
determination of the rate of infection
Ad-hIL-24 or Ad-green fluorescent protein (Ad-GFP)
were inoculated into QBI-293A cells for the amplification of
recombinant adenovirus. When 293A cells became rounded and formed
aggregates under microscopy, the cells were considered to be
infected by adenovirus, and these cells were collected and
centrifuged. The supernatants were subjected to the same step at
least three times and then collected. The Ad-hIL-24 or Ad-GFP
adenoviruses were diluted to 10−4, 10−5,
10−6, 10−7 and 10−8, dispensed
into 96-well culture plates, and incubated at 37°C in the presence
of 5% CO2 for 24 h. Following incubation, the
fluorescent cells were counted under a fluorescence microscope, and
the infection rate and adenoviral titer were calculated. A549/DDP
cells were infected with Ad-hIL-24 and Ad-GFP at various
multiplicities of infection (MOIs; 25, 50, 100, 150 and 200). GFP
expression and infection efficiency were determined under
fluorescence microscopy to select the optimal MOI for maximal
transgene expression.
CCK-8 assay
The viability of treated cells was determined by
CCK-8 assay (21). Cells
(1×103) in the logarithmic phase of growth were
inoculated into 96-well plates in 100-µl aliquots and incubated at
37°C. Following incubation, 10 µl of CCK-8 assay solution (Dojindo)
was added to each well. The blank wells contained saline. The
absorbance value (A value) for each well was measured at 450 nm
with a microplate reader. The cell growth trend was observed by
plotting a growth curve, taking the A value as the vertical
coordinate and the culture time as the horizontal coordinate. After
incubation for 24 h, the cells were treated with 100 µl DDP at 5,
10, 25 or 50 µg/ml, followed by the addition of 10 µl CCK-8 assay
solution, and then the value of the cells in each well was
assessed. SPSS 17.0 software was used to calculate the DDP dose
effect on A549/DDP cells, by using the linear regression equation:
Y = a + bX (where Y represents the DDP dose, and X represents the
inhibition rate). The half maximal inhibitory concentration
(IC50), defined as the dose of drugs to induce 50%
inhibition (cellular death) was calculated, and this dose was used
in subsequent experiments. In the same way, A549/DDP cells were
treated with Ad-GFP, DDP, Ad-hIL-24, or Ad-hIL-24 plus DDP
(A549/DDP cells were infected with adenovirus using the
aforementioned method, and the MOI and DDP dose were calculated
according to the IC50 value). According to the formula,
the required amount of virus (ML) = (MOI × tumor cell number)/viral
titer. Saline was used as a negative control. The inhibition rate
of A549/DDP cells was counted using the following formula:
inhibition rate (%) = (A value of the control-A value of the
experimental group)/A value of the control × 100%.
ELISA assay
The treated cells were cultured with FBS-free medium
for 72 h, and the cell media were then harvested for use in the
IL-24 assay with the IL-24 ELISA kit. Briefly, 100 µl of culture
medium was added to each ELISA well and incubated for 2 h at 37°C.
After incubation, the liquid was removed, and 100 µl
biotin-conjugated antibody was added and incubated for 1 h at 37°C.
The supernatant was aspirated and three washes were performed.
After the final wash, the remaining liquid was completely removed
by aspiration or decanting. The plate was then inverted and blotted
against clean paper towels. Into each well, 100 µl horseradish
peroxidase (HRP)-avidin was added, and incubated for 1 h at 37°C,
before the aspiration/wash process was repeated. TMB substrate (90
µl) was next added into each well and incubated for 15–30 min at
37°C, prior to the addition of 50 µl Stop Solution. The optical
density in each well was determined using a microplate reader at
450 nm.
Western blotting
Akt, p-Akt, and P-gp expression in the treated
A549/DDP cells were detected as previously described with some
modifications (22). Briefly,
A549/DDP cells were treated with Ad-GFP, Ad-hIL-24, DDP, or
Ad-hIL-24 plus DDP for 24 h. The treated cells were collected, and
the cellular protein was extracted and assessed by BCA protein
assay. Total protein (~100 µg) was then loaded into each lane of an
acrylamide gel and subjected to SDS-PAGE. Following
electrophoresis, the proteins were transferred onto a PVDF
membrane, and the membrane was blocked by incubation in 5% non-fat
dry milk in 0.1% Tween-20 phosphate-buffered saline (PBS-T) for 1 h
at room temperature. The membranes were incubated with antibodies
against Akt, p-Akt or P-gp at 4°C overnight. After being washed
with PBS-T, the membranes were incubated with HRP-conjugated
secondary antibodies for 1 h. The membranes were washed and then
developed with a Super enhanced chemiluminescence detection kit. A
gel imaging system was used to analyze the gray value of each
protein band. The relative photographic density was quantified.
GAPDH was used as an internal control to verify the basal
expression level and equal protein loading. The abundance ratio
relative to GAPDH was determined.
Hoechst 33342 staining
Hoechst 33342 staining (Sigma-Aldrich, Shanghai,
China) was used to observe the nuclei of A549/DDP cells. Initially,
a moderate density of A549/DDP cells was added to each well of a
6-well plate. After incubation for 24 h, the cell wells were
treated with Ad-GFP, Ad-hIL-24, DDP, or DDP plus Ad-hIL-24 for 48
h, while the cell medium alone was added to serve as a negative
control group. According to the instructions of the Hoechst 33342
kit, the treated cells were washed with PBS and fixed with 4%
paraformaldehyde at room temperature for 15 min. The cells were
then incubated with Hoechst 33342 (0.5 g/ml) for 15 min. After
removing the staining solution from the wells, the cells were
washed. The stained nuclei were observed by fluorescence
microscopy.
Flow cytometry
Cell apoptosis was detected by flow cytometry. The
treated cells were stained using Annexin V-fluorescein
isothiocyanate (FITC)/propidium iodide (PI), according to the
instructions of the Annexin V kit. Briefly, A549/DDP cells
(5×106) were transfected with Ad-GFP, Ad-hIL-24, DDP, or
DDP plus Ad-hIL-24, and incubated for 48 h. After incubation, the
treated cells were collected and washed with cold PBS. Annexin
V-FITC (5 µl) and binding buffer (500 µl) were added to the cells
and incubated for 15 min at room temperature. After incubation, the
cells were analyzed by flow cytometry.
Cell cycle analysis
A549/DDP cells were treated with Ad-GFP, DDP,
Ad-hIL-24, or Ad-hIL-24 plus DDP for 24 h. The treated cells were
collected, and subjected to cell cycle analysis as previously
described (23). Briefly,
1×104 A549/DDP cells were seeded into 6-well plates at
30% confluence. After being treated with Ad-hIL-24, the cells were
collected, and then incubated overnight with pre-cooled 70%
ethanol. The cells were washed once with PBS and then incubated
with PI for 15 min for cell cycle analysis after filtering through
a 400-micron mesh sieve. The cell samples were then subjected to
flow cytometry.
Statistical analysis
Data are presented as the mean ± standard deviation
(SD). More than two groups were compared using a single-factor
analysis of variance method. P<0.05 was considered to indicate
statistical significance.
Results
Ad-hIL-24 amplification and infected
rate in A549/DDP cells
QBI-293A cells were infected with Ad-hIL-24 or
Ad-GFP for 48 h to amplify the vectors. When viewed under an
inverted microscope, the infected cells appeared round or flaky, or
formed grape-like aggregates. After infection with Ad-GFP, many
fluorescent cells (those infected with recombinant adenovirus) were
observed under the fluorescence microscope. QBI-293A cells were
repeatedly infected to determine the viral titer up to
108 pfu/ml. A549/DDP cells were infected with Ad-hIL-24
or Ad-GFP at various MOIs (25, 50, 100, 150 or 200 MOI) for 48 h.
Considering that a high rate of infection and low cytotoxicity
offers the best MOI, in this study we selected 100 as the optimal
MOI for infecting cells. A549/DDP cells were infected with
Ad-hIL-24 and Ad-GFP, and the infected cells were counted under a
fluorescence microscope (Fig. 1A)
to determine the infection rates. The infection rates were
79.3±3.7% at 24 h, and 93.2±5.6% at 48 h (Fig. 1B).
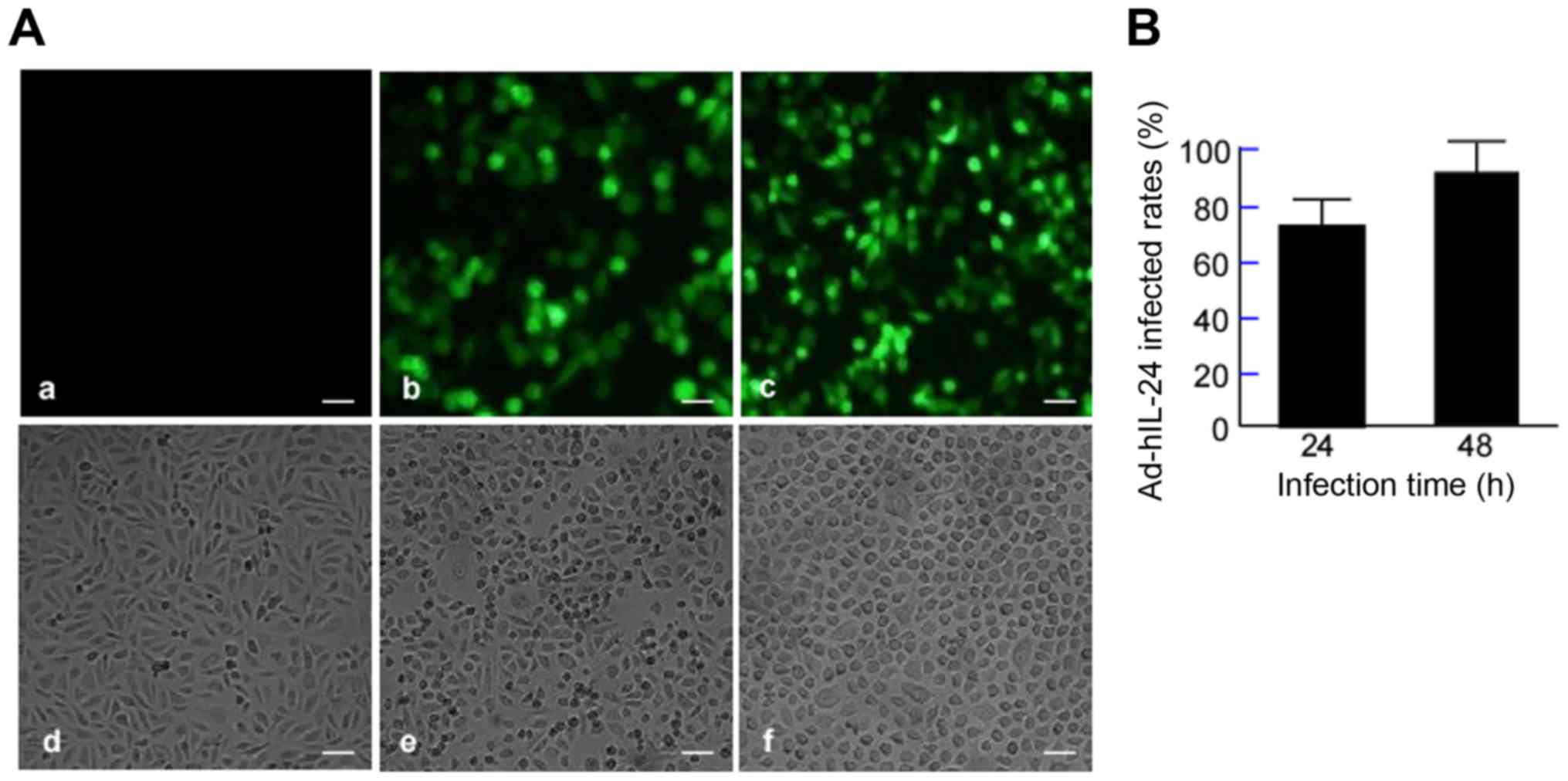 | Figure 1.Adenovirus-mediated human interleukin
24 gene (Ad-hIL-24) infects A549/DDP cells. A549/DDP cells were
infected with Ad-hIL-24 and Ad-GFP, and incubated for 24 or 48 h.
Ad-GFP served as an internal control. The treated cells were fixed
with paraformaldehyde and stained with a fluorescent antibody.
Saline (DDP solvent) served as the blank controls. (A) Green
fluorescent protein (GFP) expression was observed under
fluorescence microscopy. Scale bar, 25 µm. Magnification, ×100.
Upper panel, fluorescence microscopy images: a, saline; b, cells
infected with Ad-hIL-24 for 24 h; c, cells infected with Ad-hIL-24
for 48 h; Lower panel, light-field images: d, saline; e, cells
infected with Ad-hIL-24 for 24 h; f, cells infected with Ad-hIL-24
for 48 h. (B) GFP cells were counted under fluorescence microscopy,
and the infected rates were assessed. |
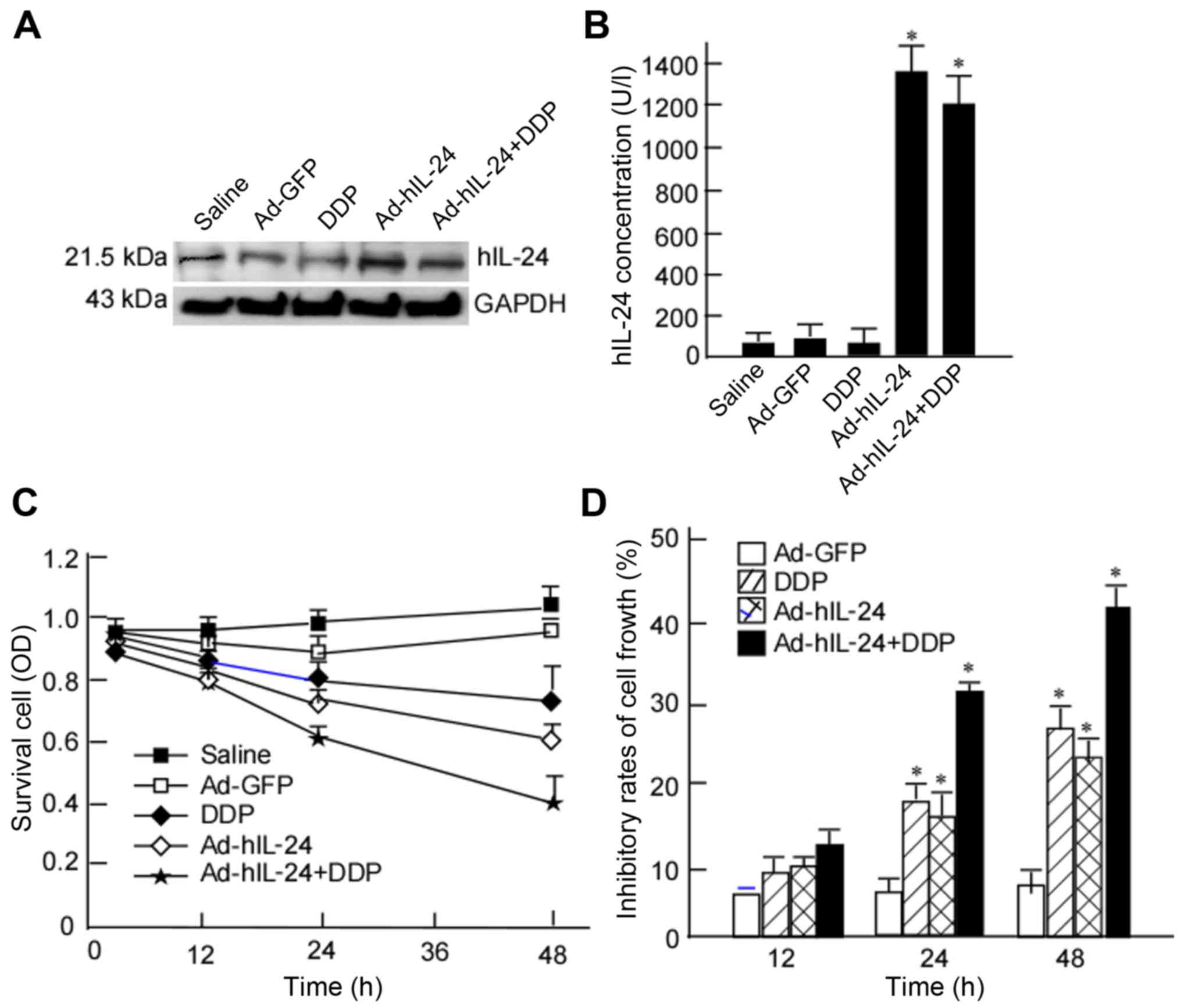
Inhibitory effect of Ad-hIL-24 on
A549/DDP cell growth
A549/DDP cells are tolerant to DDP. A549/DDP cells
were treated with DDP, Ad-hIL-24, or Ad-hIL-24 plus DDP, and hIL-24
expression levels were detected in the treated cells. hIL-24 was
effectively expressed in the cells transfected with Ad-hIL-24
(Fig. 2A), and a high concentration
of hIL-24 was detected in the cell media from the Ad-hIL-24
treatment group (Fig. 2B). A549/DDP
cells were treated with DDP at various concentrations, and then the
viability of the treated cells was detected using the CCK-8 assay.
Using the linear regression equation y = 45.611× - 0.643, the
IC50 value of DDP in A549/DDP cells was calculated. The
results revealed that A549/DDP cells displayed a high resistance to
DDP (IC50, 22.0 µg/ml). This concentration was used as
the optimal dose of DDP in subsequent experiments.
To observe the effect of Ad-hIL-24 on A549/DDP cell
growth, A549/DDP cells were treated with Ad-hIL-24 for 12, 24, and
48 h, and then the viability of the infected cells was detected
using the CCK-8 assay. A549/DDP cell viability was markedly
decreased after infection with Ad-hIL-24 (Fig. 2C), and was found to gradually
decrease with increasing infection time, especially at 48 h after
infection. The viability of these cells was significantly decreased
compared with the blank control (Fig.
2C). Furthermore, cell viability was lower in the Ad-hIL-24
plus DDP group than in the groups treated with Ad-hIL-24 or DDP
alone (Fig. 2C). This indicated
that Ad-hIL-24 could increase the extent of inhibition exerted by
DDP on A549/DDP cell viability.
Inhibition rates of Ad-hIL-24 in
A549/DDP cells
A549/DDP cells were infected with Ad-hIL-24 at 100
MOI and treated with 22.0 µg/ml DDP for 12, 24 or 48 h. hIL-24
expression was detected by western-blotting. hIL-24 expression in
the Ad-hIL-24 plus DDP group was lower than that in the Ad-hIL-24
group (Fig. 2A and B). In the group
of A549/DDP cells treated with DDP alone, hIL-24 expression was not
significantly different compared with the control. However, when
DDP was combined with Ad-hIL-24 treatment, hIL-24 expression was
markedly decreased (Fig. 2A and B).
This revealed that there may be a synergistic reaction between
Ad-hIL-24 and DDP. The cell viability was detected using a CCK-8
assay. Following a 24-h infection, the inhibitory rates in the
Ad-hIL-24, DDP, and Ad-hIL-24 plus DDP groups were 17.63±1.55%,
11.57±1.92%, 30.03±1.01%, respectively, which were high compared
with that in the control group (6.67±1.34%; P<0.05; Fig. 2D). After a 48-h infection, the
inhibitory rates were 27.00±2.00%, 19.37±1.70%, and 42.93±2.59%,
respectively, which were significantly higher than that in the
control group (7.27±1.93%; P<0.05; Fig. 2D). The inhibitory rates were also
higher at 48 h than at 24 h (P<0.05). This indicated that the
inhibition rate was time-dependent, increasing with the increasing
reaction time of Ad-hIL-24. The inhibitory rate of the combined
treatment group (42.93±2.59%) was significantly higher than that of
the groups treated with Ad-hIL-24 alone (27.00±2.00%) or DDP alone
(19.37±1.70%; P<0.05; Fig. 2D).
The inhibitory rate increased from 19.37% in the DDP group to
42.93% in the combined group, amounting to a 2.22-times increase in
growth inhibition (P<0.05; Fig.
2D). These results indicated that Ad-hIL-24 significantly
enhanced the inhibition of A549/DDP cell viability by DDP.
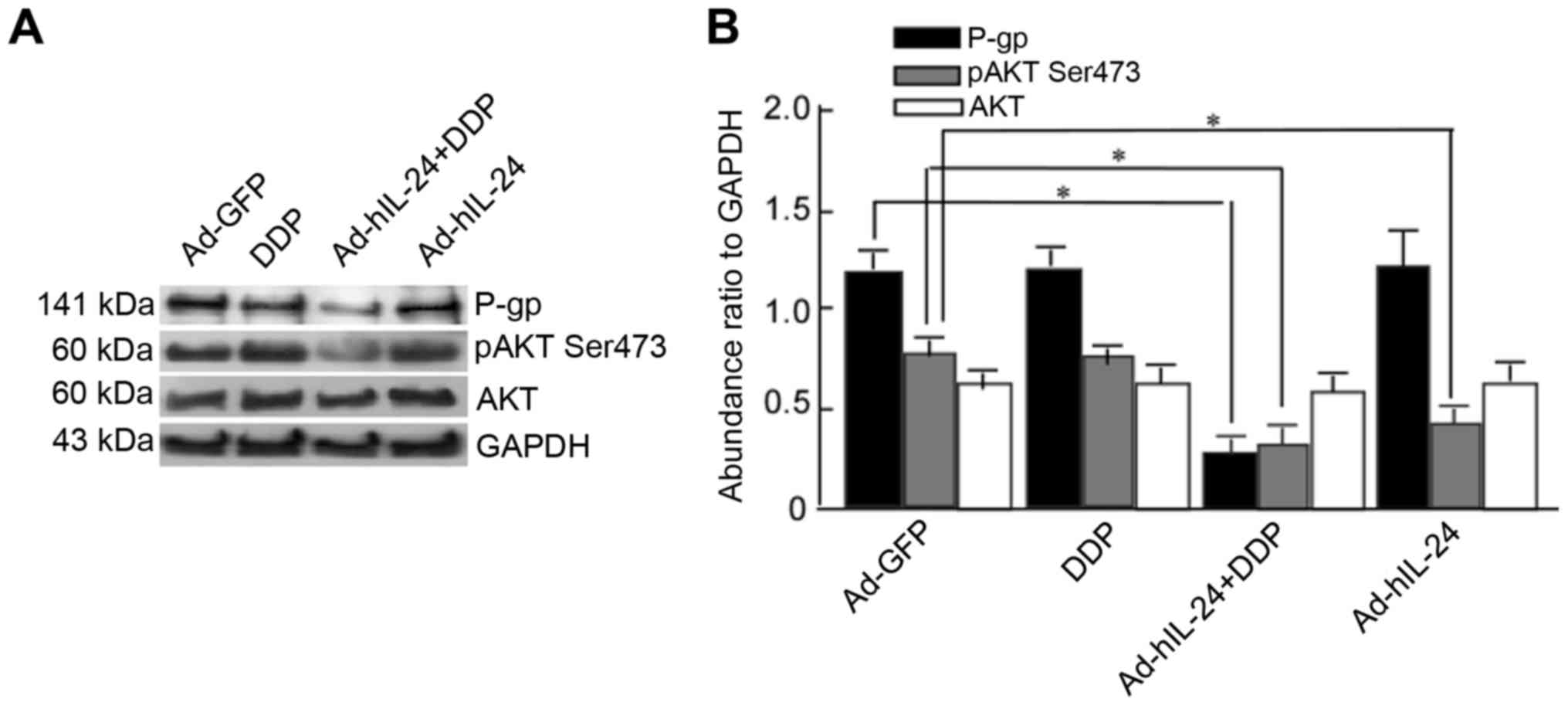
hIL-24 induces decreases in p-Akt and
P-gp expression levels
hIL-24 may exert a reversal effect on drug
resistance in A549/DDP cells. P-gp pumps drugs out of cancer cells,
and decreases the effect of chemotherapeutic drugs, thereby
rendering cells drug-resistant (6,7). To
further investigate whether the effect of hIL-24 was associated
with P-gp, A549/DDP cells were treated with Ad-hIL-24, DDP, or
Ad-hIL-24 plus DDP, and P-gp expression was detected by
western-blotting. P-gp expression was markedly decreased in the
combined group when compared with the groups treated with Ad-hIL-24
or DDP alone (Fig. 3A and B;
P<0.05). This suggested that Ad-hIL-24 combined with DDP could
effectively decrease P-gp expression, and that Ad-hIL-24-mediated
growth inhibition may be associated with decreasing P-gp
expression. Components of the phosphoinoside-3-kinase (PI3K)/AKT
signaling pathway are overexpressed in many tumors; when PI3K is
activated, PIP2 and PIP3 expression levels increase and they bind
to AKT through their pleckstrin homology (PH) domains, leading to
AKT phosphorylation. p-Akt inhibits antiapoptotic signals and
promotes cell apoptosis. Thus, we detected p-AKT expression in the
treated cells, and obtained the expected results. P-gp and p-Akt
expression in the Ad-hIL-24 plus DDP group were significantly
decreased when compared with the DDP or Ad-hIL-24 groups (Fig. 3A and B; P<0.05). However, the
expression of total Akt was not significantly different among the
groups (Fig. 3A and B). This
indicated that Ad-hIL-24 plus DDP decreased p-AKT expression, and
implied that the role of Ad-hIL-24 in DDP-mediated apoptosis was
directly related to decreased p-AKT expression.
Morphological changes of A549/DDP
cells with Ad-hIL-24 treatment
The aforementioned results showed that Ad-hIL-24
decreased p-AKT expression. p-AKT downregulation has been shown to
induce cell apoptosis (24). We
next observed the morphological changes in A549/DDP cells following
Ad-hIL-24 treatment. A549/DDP cells were treated with Ad-hIL-24,
DDP, or Ad-hIL-24 plus DDP for 48 h, then stained with Hoechst
33342. The morphology of the treated cells was observed under a
fluorescence microscope. After treatment, the cell fluorescence
intensity increased and some were evenly stained, indicating that
the cells had begun to undergo apoptosis (Fig. 4A). In the combined treatment group,
a large number of apoptotic cells were present (Fig. 4A-e); the number of apoptotic cells
was greater than that in the Ad-hIL-24 (Fig. 4A-c) and DDP alone treatment groups
(Fig. 4A-d).
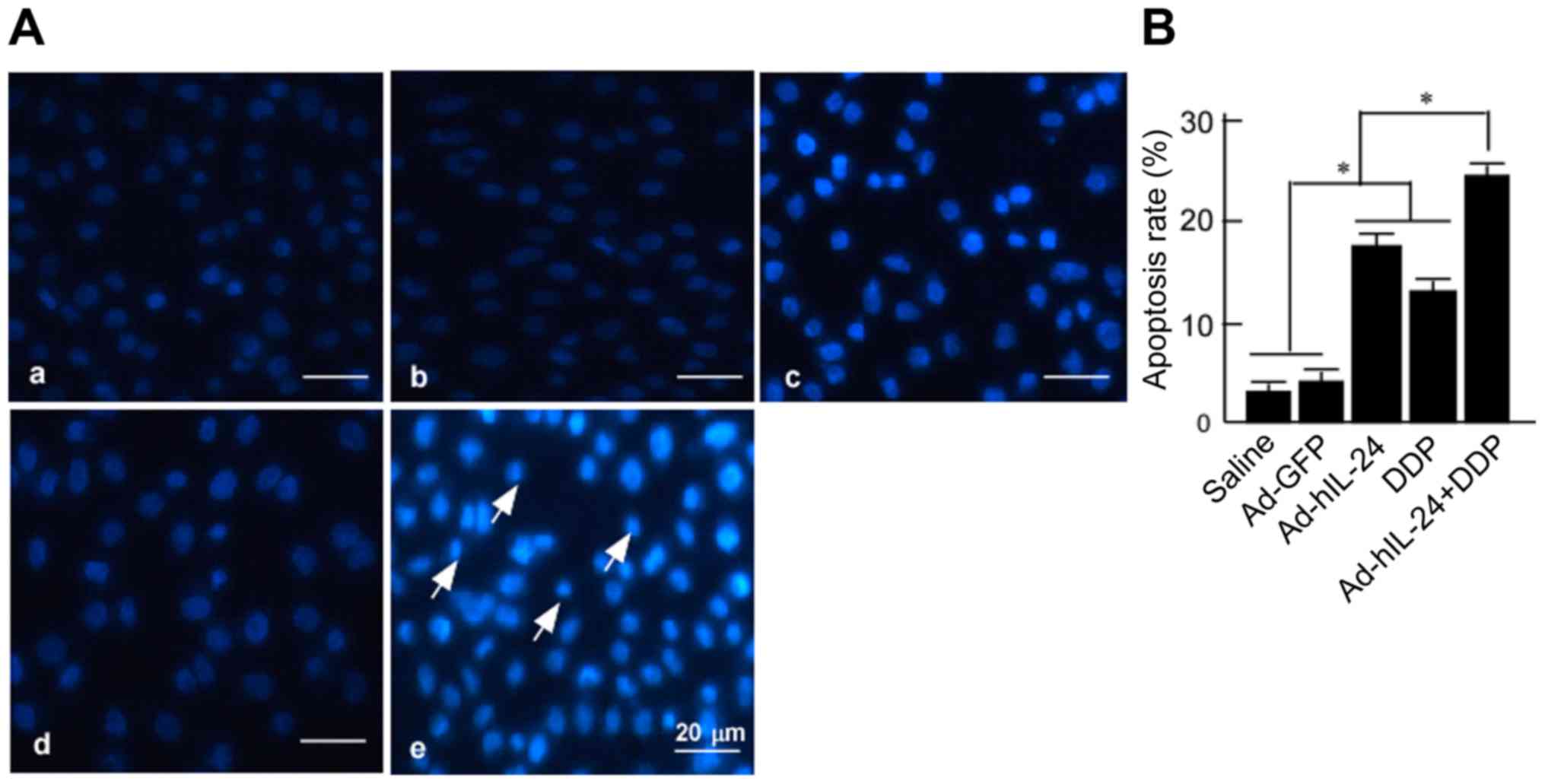 | Figure 4.Morphological analysis of
Ad-hIL-24-mediated A549/DDP cell apoptosis. A549/DDP cells were
treated with DDP, Ad-hIL-24, and Ad-hIL-24 plus DDP, and incubated
for 48 h. The treated cells were fixed with paraformaldehyde and
then stained with Hoechst 33342. Saline served as the blank
control, and Ad-GFP as the vector control. (A) The apoptotic cells
were observed under fluorescence microscopy: a, saline; b,
Ad-vector; c, Ad-hIL-24; d, DDP; e, Ad-hIL-24 plus DDP. Scale bar,
50 µm. Magnification, ×400. (B) The rates of apoptotic cells were
counted. Data are presented as the means ± SD from three
independent experiments statistically using the Students t test
(*P<0.05). |
Apoptotic cells were counted under a fluorescence
microscope, and the apoptosis rate was calculated. The apoptosis
rates of the Ad-hIL-24, DDP, and Ad-hIL-24 plus DDP groups were
respectively 17.50±1.32%, 12.83±1.04% and 24.50±1.00%, all of which
were higher than that of the control (P<0.05) (Fig. 4B). The apoptotic rate of the
combined group (24.50±1.00%) was higher than that of the Ad-hIL-24
(17.50±1.32%) and DDP (12.83±1.04%) groups (P<0.05). This
implied that Ad-hIL-24 increased the apoptosis-inducing effect of
DDP on A549/DDP cells.
Flow cytometric analysis of
hIL-24-mediated apoptosis
To further confirm that hIL-24 enhanced DDP-mediated
cell apoptosis, flow cytometry was used to detect the apoptosis of
cells treated with Ad-hIL-24, DDP, or Ad-hIL-24 plus DDP. Apoptotic
cells were observed in the Ad-hIL-24 plus DDP groups, and were more
frequent in these groups than in the control group (Fig. 5A). The flow cytometry data revealed
that the apoptosis rates in the groups treated with Ad-hIL-24, DDP,
and Ad-hIL-24 plus DDP were 27.90±0.98%, 16.40±0.95% and
39.61±1.38%, respectively, which were higher than that of the blank
and vector control groups (7.62±0.85%, 8.25±1.51%). The rate of
apoptosis in the combined group (39.61±1.38%) was obviously
increased when compared with the groups treated with Ad-hIL-24
(27.90±0.98%) or DDP alone (16.40±0.95%) (P<0.05) (Fig. 5B). Those results revealed that
Ad-hIL-24 enhances the DDP-induced apoptosis of A549/DDP cells.
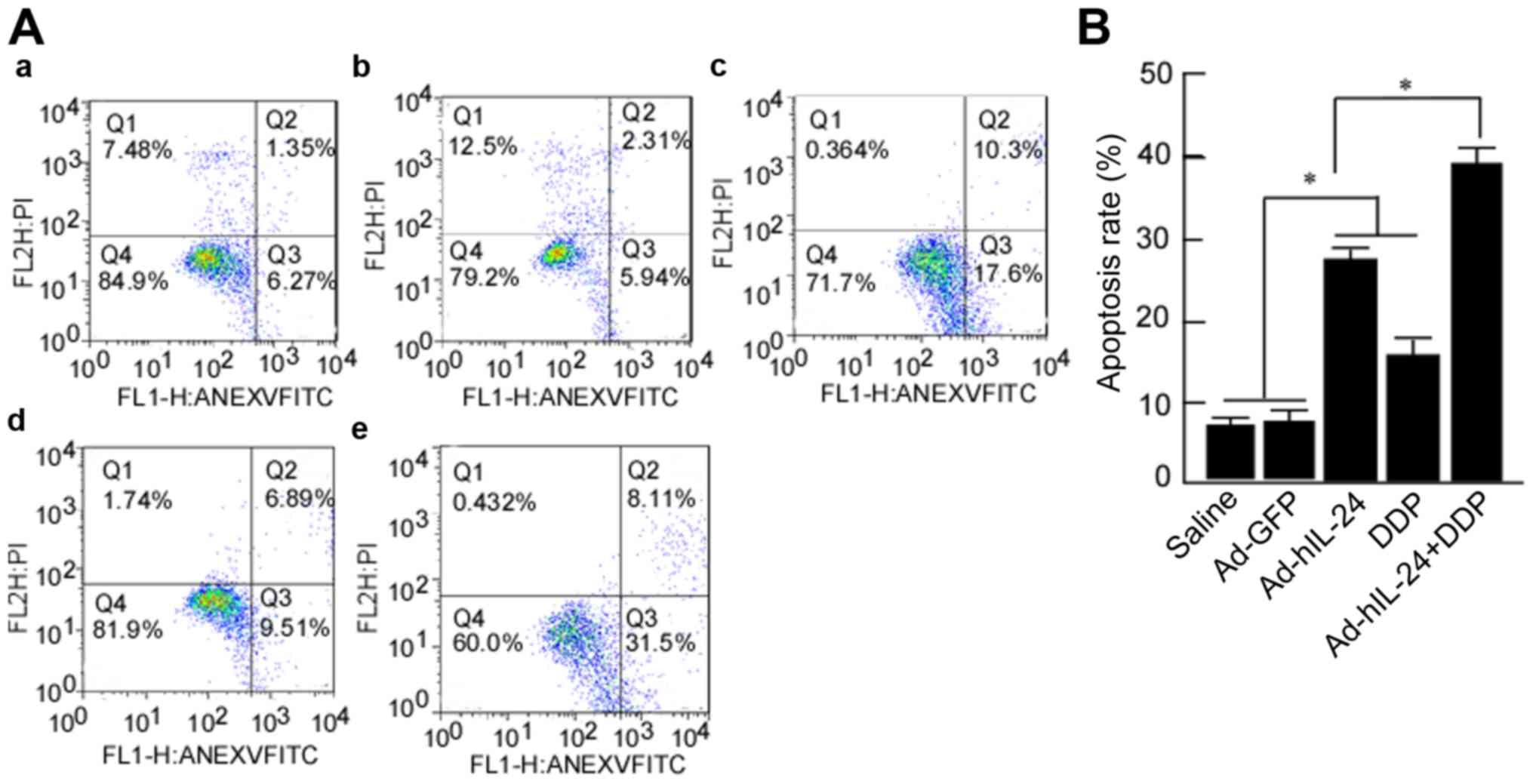 | Figure 5.Flow cytometric analysis of
Ad-hIL-24-mediated A549/DDP cell apoptosis. A549/DDP cells were
treated with DDP, Ad-hIL-24, and Ad-hIL-24 plus DDP, and incubated
for 48 h. Then the treated cells were fixed with ethanol and
stained with Annexin V-FITC. Saline served as the blank control,
and Ad-GFP as the vector control. (A) The apoptotic cells were
analyzed with flow cytometry: a, saline; b, Ad-vector; c,
Ad-hIL-24; d, DDP; e, Ad-hIL-24 plus DDP. Scale bar, 50 µm.
Magnification ×400. (B) The rates of apoptotic cells were counted.
Data are presented as the means ± SD from three independent
experiments statistically using the Students t test
(*P<0.05). |
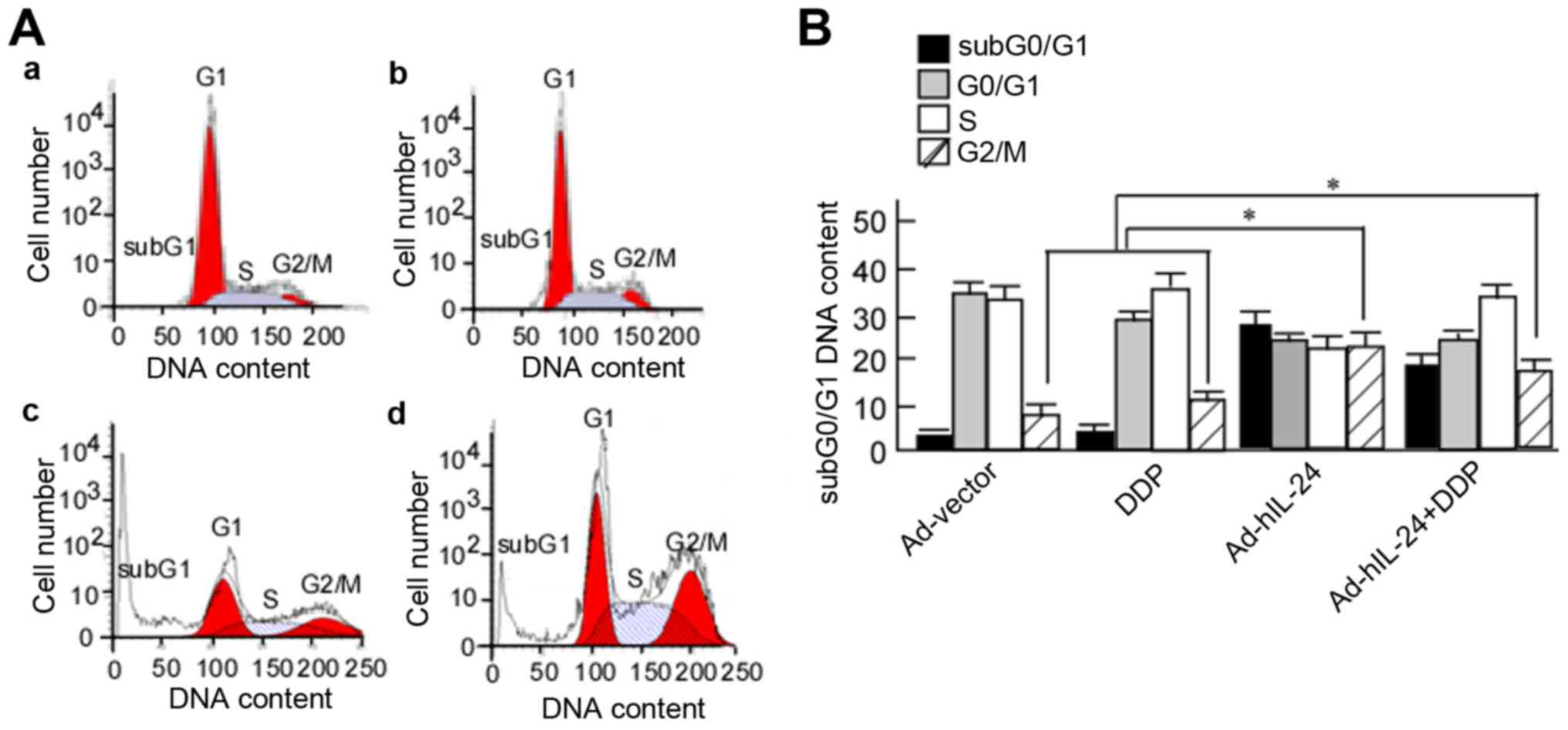
hIL-24 induces A549/DDP cell-cycle
arrest
The aforementioned results revealed that Ad-hIL-24
decreased p-AKT expression, which induced cell-cycle arrest
(25). The next step was to observe
the cell cycle of A549/DDP cells. A549/DDP cells were treated with
Ad-hIL-24, DDP, or Ad-hIL-24 plus DDP for 48 h, and the cell cycle
distribution was analyzed using flow cytometry. The cells were
arrested at the G2/M phase after being treated with hIL-24
(Fig. 6A-c and -d). The proportion
of cells in the subG1, G1, S and G2/M phases was determined.
G2/M-phase cells accounted for 8.5±1.32%, 13.2±2.14%, 24.6±4.33%,
and 19.50±4.12% of cells in the Ad-vector, DDP, Ad-hIL-24, and DDP
plus Ad-hIL-24 groups, respectively. Compared with the Ad-vector
and DDP groups, the number of G2/M-phase cells was markedly
increased in the Ad-hIL-24 and Ad-hIL-24 plus DDP groups (Fig. 6B, P<0.05). This demonstrated that
Ad-hIL-24 induced A549/DDP cell-cycle arrest.
Discussion
The mortality and morbidity associated with lung
cancer represent a serious threat to human health (26). Lung cancer patients often have an
irritating cough, expectoration, hemoptysis, and other symptoms
during the early stages of the disease, and its development is
related to factors such as tumor location and pathological type
(27). Lung cancer is divided into
non-small cell and small cell lung cancer, with non-small cell lung
cancer accounting for ~80% of all cases. Treatments for lung cancer
are primarily based on the pathological type and the extent of
local invasion and metastasis. In the early stages, treatment of
lung cancer is predominantly surgical, while chemotherapy is an
important alternative in the later stages (28). Platinum drugs alone or in
combination with other drugs are considered the standard first-line
chemotherapy agents for lung cancer treatment (29). The main mechanism of platinum drugs
involves DNA recombination in tumor cells, which disrupts the
structure and function of DNA and hinders DNA replication. These
treatments exert anti-tumor effects, but also produce various side
effects, including those affecting the digestive system,
hematopoietic system and immune system in patients. Moreover,
patients develop resistance to repeated chemotherapy, which
decreases the survival rate (30).
Reversing drug resistance and decreasing the toxic and side effects
of chemotherapy drugs are two clinical concerns that must be
addressed urgently.
The mechanism underlying tumor MDR is highly complex
and involves many factors, although the overexpression of P-gp in
tumor cells is one of the most important factors (31). To date, many studies have focused on
the reversal of drug resistance in lung cancer, including the use
of calcium channel blockers, anti-arrhythmia drugs and some
traditional Chinese medicine components. Although certain drugs can
momentarily increase chemotherapeutic drug concentrations to kill
tumor cells, they cannot be widely used in a clinical setting
because of their instability and non-specific nature, as well as
the associated side effects. Therefore, it is essential to
identifying more effective therapeutic strategies or drugs that can
reverse lung cancer MDR and improve the survival rate of lung
cancer patients.
Gene therapy is expected to provide a new
therapeutic approach for decreasing drug resistance in cancer; this
approach has the advantage of high selectivity and targeted
inhibition of tumor cells. Currently, viral vectors are chiefly
used for gene therapy. Adenoviral vectors share homology with human
genes and offer high levels of safety. In addition, adenoviral
vectors are widely used due to their large capacity with respect to
exogenous genes and range of hosts. Ad-hIL-24 was constructed with
a recombinant adenoviral vector, and has been shown to exert an
inhibitory effect on various tumors (32–34).
Recently, Ad-hIL-24 was demonstrated to be capable of reversing
tumor drug resistance. Ad-hIL-24 can not only induce cell
apoptosis, but also reduce MDR gene expression (35,36).
In the present study, we found that Ad-hIL-24 had a high rate of
infection of lung cancer cells, and therefore may act as a suitable
vector for tumor gene-therapy.
To investigate the drug-resistance-reversing effect
of Ad-hIL-24, A549/DDP cells were used in this study. First, the
inhibitory effect of Ad-hIL-24 on A549/DDP cells was detected.
Ad-hIL-24 could significantly inhibit A549/DDP growth, and
combining it with DDP enhanced this effect. This indicated that
Ad-hIL-24 had an inhibitory effect on lung cancer cells that are
resistant to DDP. We also observed that Ad-hIL-24 induced A549/DDP
cell apoptosis. Based on the fluorescence microscopy findings,
Ad-hIL-24 increased DDP-mediated A549/DDP cell apoptosis, and this
was further confirmed by flow cytometric analysis. This suggests
that Ad-hIL-24 has a reversal effect on the drug resistance of lung
cancer and that it improves the sensitivity of lung cancer cells to
DDP. Additionally, P-gp overexpression causes tumor cells to
develop drug resistance. P-gp is a type of cell membrane protein,
encoded by the MDR-1 gene (37),
that can pump chemotherapeutic drugs out of cells, thereby
decreasing the concentration of chemotherapeutic drugs inside cells
and enhancing tumor cell resistance to these agents (38). We investigated whether hIL-24 could
reverse the resistance of A549/DDP cells via altering the levels of
P-gp, and found that Ad-hIL-24 significantly decreased P-gp
expression. Ad-hIL-24 may decrease the activity of P-gp as a drug
pump, enhancing the cytotoxic effect of DDP on A549/DDP cells, and
eventually reverse the resistance of A549/DDP cells to DDP.
Akt is a key functional molecule of the PI3K/AKT
signaling pathway, and p-AKT can resist antiapoptotic signals and
promote cell apoptosis (39). In
the current study, Ad-hIL-24 plus DDP decreased p-AKT expression,
while the total Akt expression did not change significantly.
Ad-hIL-24-mediated apoptosis may occur due to decreased Akt
phosphorylation. A decrease in p-AKT expression induced cell
apoptosis and arrested cells at the G2/M phase (24,25).
Therefore, hIL-24 may reverse the resistance of A549/DDP cells
through the induction of cell apoptosis and arresting cells at the
G2/M phase. Additionally, IL-24 reversed the MDR of tumors through
the PI3K/AKT signaling pathway (40–42),
the double-stranded RNA-dependent protein kinase pathway, the Bcl-2
protein family (43), the
mitochondrial pathway (44), and
the endoplasmic reticulum stress pathway (45). Based on this, we speculated that
Ad-hIL-24 may reverse A549/DDP cell resistance by affecting the
PI3K/AKT signaling pathway. Further discussion is required and
future experiments using the Rh123 method to assess the
accumulation of chemotherapeutic drugs in drug-resistant tumor
cells may help to elaborate the mechanism underlying the
Ad-hIL-24-induced reversal of lung cancer MDR.
In summary, Ad-hIL-24 reversed the MDR of A549/DDP
cells by inhibiting cell growth and inducing apoptosis. When
Ad-hIL-24 is combined with DDP, the reversal effect is enhanced
compared with the single treatments. Regarding the mechanism,
Ad-hIL-24 combined with DDP decreased P-gp expression, and its
reversal effect may be through reducing the pumping of DDP out of
the A549/DDP cells, thereby increasing DDP concentration in the
cells. Ad-hIL-24 may also inhibit Akt phosphorylation and inhibit
antiapoptotic signals to promote cell apoptosis and cell-cycle
arrest at the G2/M phase, thus reversing tumor drug resistance.
Acknowledgements
The authors acknowledge financial support from the
National Natural Science Foundation of China (nos. 81372282,
81402368, 81402265 and 81502346), the Foundation of State Key
Laboratory of Oncology in South China (HN2011-04), the Fundamental
Research Funds for the Guangdong Province (2011B061300053), and the
Zunyi Medical College Master Start Project (F-719).
Glossary
Abbreviations
Abbreviations:
|
Ad-GFP
|
Ad-green fluorescent protein
|
|
Ad-hIL-24
|
adenovirus-mediated human interleukin
24 gene
|
|
CCK-8
|
Cell Counting Kit-8
|
|
DDP
|
cisplatin
|
|
hIL-24
|
human interleukin-24
|
|
HRP
|
horseradish peroxidase
|
|
IC50
|
half maximal inhibitory
concentration
|
|
MDR
|
multidrug resistance
|
|
MOI
|
multiplicity of infection
|
|
PBS
|
phosphate-buffered saline
|
|
p-AKT
|
phosphorylated-AKT
|
|
P-gp
|
P-glycoprotein
|
|
PI3k
|
phosphoinoside-3-kinase
|
|
PMSF
|
phenylmethanesulfonyl fluoride
|
|
TBS
|
Tween-20 phosphate-buffered saline
|
References
|
1
|
She J, Yang P, Hong Q and Bai C: Lung
cancer in China: Challenges and interventions. Chest.
143:1117–1126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Camps C, del Pozo N, Blasco A, Blasco P
and Sirera R: Importance of quality of life in patients with
non-small-cell lung cancer. Clin Lung Cancer. 10:83–90. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Satgé D, Kempf E, Dubois JB, Nishi M and
Trédaniel J: Challenges in diagnosis and treatment of lung cancer
in people with intellectual disabilities: Current state of
knowledge. Lung Cancer Int. 2016:67876482016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takase N, Hattori Y, Kiriu T, Itoh S, Kawa
Y, Yamamoto M, Urata Y, Shimada T, Tsujino K, Soejima T, et al:
Concurrent chemoradiotherapy with cisplatin and S-1 or vinorelbine
for patients with stage III unresectable non-small cell lung
cancer: A retrospective study. Respir Investig. 54:334–340. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lunacsek OE, Ravelo A, Coutinho AD, Hazard
SJ, Green MR, Willey J, Eaddy M and Goertz HP: First-line treatment
with bevacizumab and platinum doublet combination in non-squamous
non-small cell lung cancer: A retrospective cohort study in US
oncology community practices. Drugs Real World Outcomes. 3:333–343.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Teixeira SF, de Azevedo RA, Silva AC,
Braga RC, Jorge SD, Barbuto JA, Andrade CH and Ferreira AK:
Evaluation of cytotoxic effect of the combination of a pyridinyl
carboxamide derivative and oxaliplatin on NCI-H1299 human non-small
cell lung carcinoma cells. Biomed Pharmacother. 84:1019–1028. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu DL, Huang F and Lu HZ: Drug-resistant
proteins in breast cancer: Recent progress in multidrug resistance.
Ai Zheng. 22:441–444. 2003.(In Chinese). PubMed/NCBI
|
|
8
|
Kim I, Xu W and Reed JC: Cell death and
endoplasmic reticulum stress: Disease relevance and therapeutic
opportunities. Nat Rev Drug Discov. 7:1013–1030. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lage H: An overview of cancer multidrug
resistance: A still unsolved problem. Cell Mol Life Sci.
65:3145–3167. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Joshi AA, Vaidya SS, St-Pierre MV, Mikheev
AM, Desino KE, Nyandege AN, Audus KL, Unadkat JD and Gerk PM:
Placental ABC transporters: Biological impact and pharmaceutical
significance. Pharm Res. 33:2847–2878. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kong LL, Shen GL, Wang ZY, Zhuang XM, Xiao
WB, Yuan M, Gong ZH and Li H: Inhibition of P-glycoprotein and
multidrug resistance-associated protein 2 regulates the
hepatobiliary excretion and plasma exposure of thienorphine and its
glucuronide conjugate. Front Pharmacol. 7:2422016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang NN, Zhao LJ, Wu LN, He MF, Qu JW,
Zhao YB, Zhao WZ, Li JS and Wang JH: Mechanistic analysis of
taxol-induced multidrug resistance in an ovarian cancer cell line.
Asian Pac J Cancer Prev. 14:4983–4988. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sourisseau T, Helissey C, Lefebvre C,
Ponsonnailles F, Malka-Mahieu H, Olaussen KA, André F, Vagner S and
Soria JC: Translational regulation of the mRNA encoding the
ubiquitin peptidase USP1 involved in the DNA damage response as a
determinant of Cisplatin resistance. Cell Cycle. 15:295–302. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zuo J, Jiang H, Zhu YH, Wang YQ, Zhang W
and Luan JJ: Regulation of MAPKs signaling contributes to the
growth inhibition of 1,7-dihydroxy-3,4-dimethoxyxanthone on
multidrug resistance A549/taxol cells. Evid Based Complement
Alternat Med. 2016:20187042016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mao Z, Bian G, Sheng W, He S, Yang J and
Dong X: Adenovirus-mediated IL-24 expression enhances the
chemosensitivity of multidrug-resistantgastric cancer cells to
cisplatin. Oncol Rep. 30:2288–2296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dash R, Bhutia SK, Azab B, Su ZZ, Quinn
BA, Kegelmen TP, Das SK, Kim K, Lee SG, Park MA, et al:
mda-7/IL-24: A unique member of the IL-10 gene family promoting
cancer-targeted toxicity. Cytokine Growth Factor Rev. 21:381–391.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu J, Sheng W, Xie Y, Shan Y, Miao J,
Xiang J and Yang J: The in vitro and in vivo antitumor activity of
adenovirus-mediated interleukin-24 expression for laryngocarcinoma.
Cancer Biother Radiopharm. 25:29–38. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu X, Xia W, Li S, Blumenfeld J, Zhang B,
Yang J, Miao J and Gu ZJ: Antitumor effect and underlying mechanism
of RGD-modified adenovirus mediated IL-24 expression on myeloid
leukemia cells. Int Immunopharmacol. 28:560–570. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen F, Zhao Q, Wang S, Wang H and Li X:
Upregulation of Id3 inhibits cell proliferation and induces
apoptosis in A549/DDP human lung cancer cells in vitro. Mol Med
Rep. 14:313–318. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie Y, Lv H, Sheng W, Miao J, Xiang J and
Yang J: Synergistic tumor suppression by adenovirus-mediated
inhibitor of growth 4 and interleukin-24 gene cotransfer in
hepatocarcinoma cells. Cancer Biother Radiopharm. 26:681–695. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zheng D, Chen Z, Chen J, Zhuang X, Feng J
and Li J: Exogenous hydrogen sulfide exerts proliferation,
anti-apoptosis, migration effects and accelerates cell cycle
progression in multiple myeloma cells via activating the Akt
pathway. Oncol Rep. 36:1909–1916. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang F, Zou F, Peng Z, Huang D, Wu Y, Chen
Y, Duan C, Cao Y, Mei W, Tang X, et al:
N,N'-dinitrosopiperazine-mediated ezrin protein phosphorylation via
activation of Rho kinase and protein kinase C is involved in
metastasis of nasopharyngeal carcinoma 6-10B cells. J Biol Chem.
286:36956–36967. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kang T, Wei Y, Honaker Y, Yamaguchi H,
Appella E, Hung MC and Piwnica-Worms H: GSK-3 beta targets Cdc25A
for ubiquitin-mediated proteolysis, and GSK-3 beta inactivation
correlates with Cdc25A overproduction in human cancers. Cancer
Cell. 13:36–47. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiao M, Tang Y, Wang YL, Yang L, Li X,
Kuang J and Song GL: ART1 silencing enhances apoptosis of mouse
CT26 cells via the PI3K/Akt/NF-κB pathway. Cell Physiol Biochem.
32:1587–1599. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Y, Zhang P, Qiu F, Chen L, Miao C, Li
J, Xiao W and Ma E: Inactivation of PI3K/Akt signaling mediates
proliferation inhibition and G2/M phase arrest induced by
andrographolide in human glioblastoma cells. Life Sci. 90:962–967.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen W, Zhang S and Zou X: Evaluation on
the incidence, mortality and tendency of lung cancer in China.
Thorac Cancer. 1:35–40. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee PN and Gosney JR: The effect of time
changes in diagnosing lung cancer type on its recorded
distribution, with particular reference to adenocarcinoma. Regul
Toxicol Pharmacol. 81:322–333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mak KS, van Bommel AC, Stowell C, Abrahm
JL, Baker M, Baldotto CS, Baldwin DR, Borthwick D, Carbone DP, Chen
AB, et al Lung Cancer Working Group of ICHOM, : Defining a standard
set of patient-centred outcomes for lung cancer. Eur Respir J.
48:852–860. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamada K, Ichiki M, Takahashi K, Hisamatsu
Y, Takeoka H, Azuma K, Shukuya T, Nishikawa K, Tokito T, Ishii H,
et al: A multicenter phase II trial of S-1 combined with
bevacizumab after platinum-based chemotherapy in patients with
advanced non-squamous non-small cell lung cancer. Cancer Chemother
Pharmacol. 78:501–507. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Darwish MA, Abo-Youssef AM, Khalaf MM,
Abo-Saif AA, Saleh IG and Abdelghany TM: Vitamin E mitigates
cisplatin-induced nephrotoxicity due to reversal of
oxidative/nitrosative stress, suppression of inflammation and
reduction of total renal platinum accumulation. J Biochem Mol
Toxicol. 31:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Takara K, Sakaeda T, Kakumoto M,
Tanigawara Y, Kobayashi H, Okumura K, Ohnishi N and Yokoyama T:
Effects of alpha-adrenoceptor antagonist doxazosin on MDR1-mediated
multidrug resistance and transcellular transport. Oncol Res.
17:527–533. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pan XT, Zhu QY, Li DC, Yang JC, Zhang ZX,
Zhu XG and Zhao H: Effect of recombinant adenovirus vector mediated
human interleukin-24 gene transfection on pancreatic carcinoma
growth. Chin Med J. 121:2031–2036. 2008.PubMed/NCBI
|
|
33
|
Zheng SY, Ge JF, Zhao J, Jiang D and Li F:
Adenovirus-mediated IL-24 confers radiosensitization to human lung
adenocarcinoma in vitro and in vivo. Mol Biol Rep. 42:1069–1080.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lv C, Su Q, Liang Y, Hu J and Yuan S:
Oncolytic vaccine virus harbouring the IL-24 gene suppresses the
growth of lung cancer by inducing apoptosis. Biochem Biophys Res
Commun. 476:21–28. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu J, Mo Y, Wang X, Liu J, Zhang X, Wang
J, Hu L, Yang C, Chen L and Wang Y: Conditionally replicative
adenovirus-based mda-7/IL-24 expression enhances sensitivity of
colon cancer cells to 5-fluorouracil and doxorubicin. J
Gastroenterol. 48:203–213. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fang P, Zhang X, Gao Y, Ding CR, Cui F and
Jiao SC: Reversal effect of melanoma differentiation associated
gene-7/interleukin-24 on multidrug resistance in human
hepatocellular carcinoma cells. Anat Rec. 295:1639–1646. 2012.
View Article : Google Scholar
|
|
37
|
Bajaj G, Rodriguez-Proteau R, Venkataraman
A, Fan Y, Kioussi C and Ishmael JE: MDR1 function is sensitive to
the phosphorylation state of myosin regulatory light chain. Biochem
Biophys Res Commun. 398:7–12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yun UJ, Lee JH, Koo KH, Ye SK, Kim SY, Lee
CH and Kim YN: Lipid raft modulation by Rp1 reverses multidrug
resistance via inactivating MDR-1 and Src inhibition. Biochem
Pharmacol. 85:1441–1453. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu XS, Du J, Fan YJ, Liu FJ, Cao LL, Liang
N, Xu DG and Zhang JD: Activation of endoplasmic reticulum stress
promotes autophagy and apoptosis and reverses chemoresistance of
human small cell lung cancer cells by inhibiting the PI3K/AKT/mTOR
signaling pathway. Oncotarget. 7:76827–76839. 2016.PubMed/NCBI
|
|
40
|
Burris HA III: Overcoming acquired
resistance to anticancer therapy: Focus on the PI3K/AKT/mTOR
pathway. Cancer Chemother Pharmacol. 71:829–842. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mosca E, Barcella M, Alfieri R, Bevilacqua
A, Canti G and Milanesi L: Systems biology of the metabolic network
regulated by the Akt pathway. Biotechnol Adv. 30:131–141. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Polivka J Jr and Janku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pataer A, Chada S, Roth JA, Hunt KK and
Swisher SG: Development of Ad-mda7/IL-24-resistant lung cancer cell
lines. Cancer Biol Ther. 7:103–108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang BX, Duan YJ, Dong CY, Zhang F, Gao
WF, Cui XY, Lin YM and Ma XT: Novel functions for mda-7/IL-24 and
IL-24 delE5: Regulation of differentiation of acute myeloid
leukemic cells. Mol Cancer Ther. 10:615–625. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shi H, Wei LL, Yuan CF, Yang JX, Yi FP, Ma
YP and Song FZ: Melanoma differentiation-associated
gene-7/interleukin 24 inhibits invasion and migration of human
cervical cancer cells in vitro. Saudi Med J. 28:1671–1675.
2007.PubMed/NCBI
|