Introduction
Regulatory T-cell receptors serve as
immunotherapeutic targets for enhancing activation of antitumor
immune responses or reversing immunosuppressive mechanisms of tumor
resistance to immune surveillance and destruction. Cytotoxic
T-lymphocyte antigen-4 (CTLA-4), an essential inhibitory regulator,
is responsible for the early stages of T-cell expansion that
opposes the action of CD28-mediated costimulation (1). Following T-cell activation, CTLA-4 is
rapidly upregulated, then binds to B7 molecules with a higher
affinity than CD28 (2,3). CTLA-4 may abolish the initiation of
the responses of T cells by raising the threshold of signals
required for full activation of T cells, and it also may terminate
the ongoing T-cell responses. Based on the significant regulatory
effect of CTLA-4 on immune responses, antibodies against either
mouse or human CTLA-4 have been developed for boosting
immunological responses against cancer (4). Anti-CTLA-4 antibodies have been
confirmed to confer a blockage effect on CTLA-4/B7 interactions
in vivo, and they can enhance T-cell responses to peptides,
superantigens, and parasites (5).
It has been shown that anti-CTLA-4 antibodies are able to induce
the rejection of newly implanted murine tumors (6,7). In
addition, promising results have been observed in clinical trials
of anti-human CTLA-4 monoclonal antibodies (mAbs) for the treatment
of late-stage metastatic melanoma (8,9).
Based on their high affinity and specificity, mAbs
have become ideal therapeutic strategies for research, diagnosis
and clinical applications (10,11).
The traditional immunoglobulin G (IgG) molecules consist of two
identical heavy chains and light chains, forming the antigen
binding site together. However, the complex structure, costly
production and unstable behavior of IgG greatly limit their
practical applications (12–14).
Recently, single domain antibodies (sdAbs; also called nanobodies;
Nbs) have emerged as small (~15 kDa) antigen-binding fragments
which are derived from camelid heavy-chain antibodies (15). They present several advantages
including good solubility, thermal stability, and high expression
yield (16–18). Furthermore, Nbs have a natural
tendency for binding epitopes that are inaccessible to conventional
antibodies (19). Nbs have been
evaluated in vitro and in vivo and have been proven
to be a valuable tool for optical molecular imaging of
HER2-positive breast cancer (20).
They are capable of selectively targeting HGF-producing tumors.
Furthermore, treatment of U87 MG-bearing mice with these Nbs
resulted in inhibition of tumor growth and ultimately caused cures
(21). Consequently, these unique
advantages make Nbs an attractive and valuable approach for the
diagnosis and treatment of tumors.
In the present study, we successfully constructed an
immune phage display library against CTLA-4 with the size of
1.85×108 colonies and generated characteristic
anti-CTLA-4 Nbs. We further demonstrated their valuable properties
of high binding rates and anti-melanoma activity.
Materials and methods
Reagents and materials
The human CTLA-4 protein was purchased from Abcam
(Cambridge, UK). Freund's incomplete adjuvant was purchased from
Sigma-Aldrich (St. Louis, MO, USA).
Density gradient centrifugation with Ficoll-Paque™
Plus (GE Healthcare, Beijing, China). Fast Track 2.0 kit and
ThermoScript RT-PCR kit were provided by Invitrogen (Carsbad, CA,
USA). OligodT primers were obtained from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Restriction enzymes
PstI and NotI were provided by New England BioLabs
(NEB) (Ipswich, MA, USA). Anti-mouse IgG-alkaline phosphatase,
NI-NTA Superflow sepharose column and bisphosphate and
phytohemagglutinin (PHA) were acquired from Sigma-Aldrich. The
VCSM13 helper phages, TG1 and WK6 cells were kindly provided by
Professor Serge Muyldermans (Laboratory of Cellular and Molecular
Immunology, Vrije Universiteit Brussel, Brussels, Belgium). Anti-HA
tag antibody and mouse anti-human CTLA-4 mAb were purchased from
Abcam (clone, 16B12) and BD Pharmingen (San Diego, CA, USA; clone,
BNI3), respectively.
Cells and animals
B16/BL6 cells were obtained from the National Center
for International Research of Biological Targeting Diagnosis and
Therapy of Guangxi Medical University. B16/BL6 cells were cultured
in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine
serum (FBS) (both from Gibco, Grand Island, NY, USA) and 1% double
antibiotics (penicillin/streptomycin) at 37°C in a 5%
CO2 incubator.
C57BL/6 mice were obtained from Vital River Company
(Beijing, China) and were raised in specific pathogen-free (SPF)
conditions. All animal experiments were carried out according to
the guidelines of the Federation of European Laboratory Animal
Science Associations. All protocols were approved by the Animal
Ethics Committee of Guangxi Medical University.
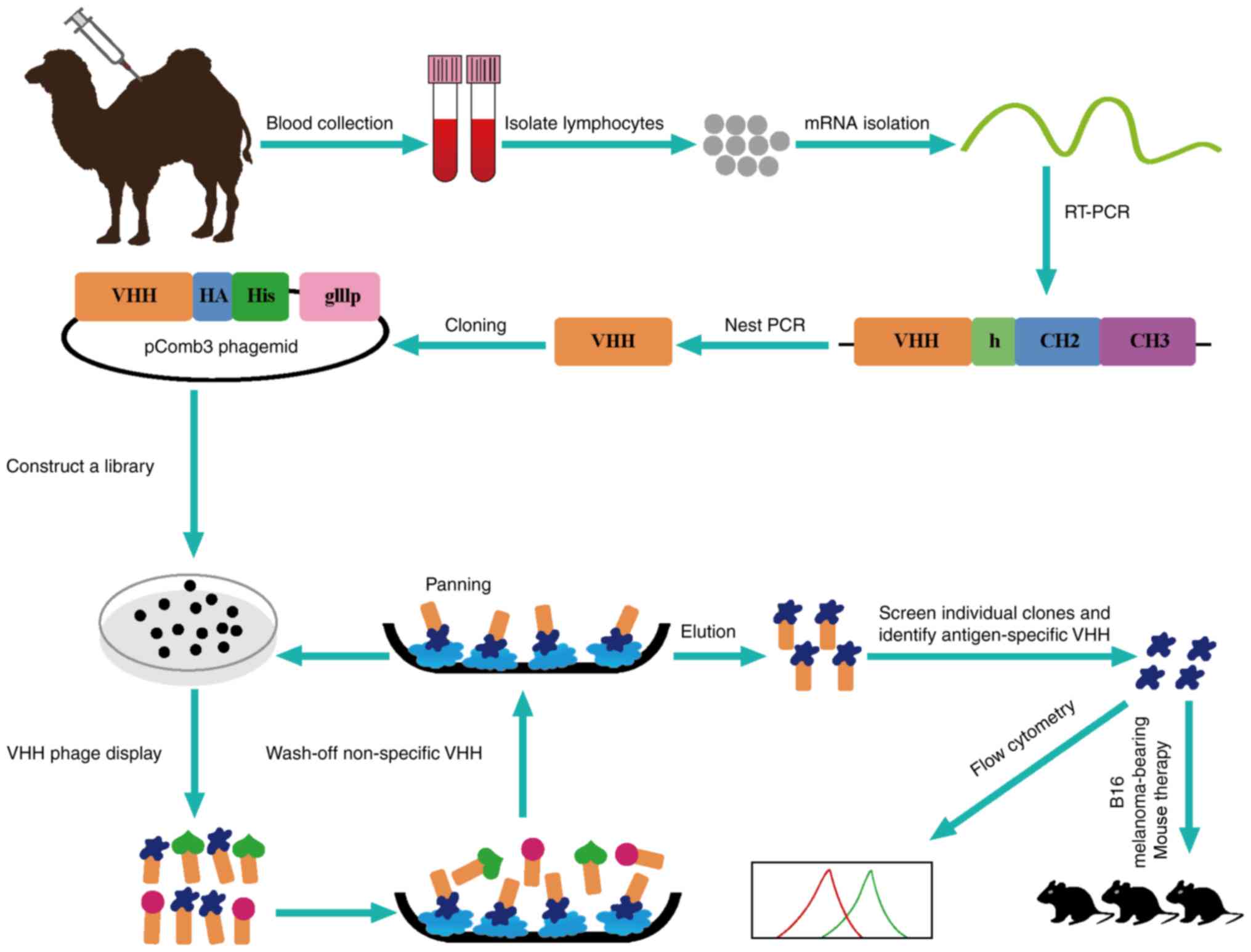
Dromedary camel immunization
A healthy dromedary camel was immunized
subcutaneously 7 times at 7-day intervals with human CTLA-4 protein
(1 mg, 1 ml) mixed with an equal volume of Freund's incomplete
adjuvant for stimulating antigen-specific B cells expressing Nbs
(22). The peripheral blood
lymphocytes were extracted from 100 ml blood sample to construct
the library after the last injection.
Library construction
Lymphocytes were isolated from peripheral blood by
density gradient centrifugation. The total RNA was extracted from
~107 lymphocytes by using Fast Track 2.0 kit and mRNA
(40 µg) was used to synthesize cDNA strands using a ThermoScript
RT-PCR kit with oligodT primers. To avoid contamination
of VH genes, the variable regions of heavy-chain
immunoglobulins (VHH) were amplified by 2-steps nested PCR. The
first step PCR was performed with a template of the first-strand
cDNA using the primers CALL001 and CALL002 (23). This protocol consisted of an initial
denaturation step at 94°C for 7 min, followed by 94°C for 1 min,
55°C for 1 min, and 72°C for 1 min for 30 cycles, and a final
extension step at 72°C for 10 min. The first PCR products consist
of ~700 bp fragments and were used as the template for the second
step PCR. Then, the VHH encoding gene fragments were amplified by
second PCR using degenerated primers including PstI and
NotI restriction sites. The amplified products were ligated
into phagemid pComb3 after digesting by restriction enzymes
PstI and NotI and then electro-transformed into
competent E. coli TG1 cells (24). The transformants were plated onto 2X
YT medium which contained 2% glucose and 100 µg/ml ampicillin. The
transformants were subsequently cultured at 37°C overnight. After
gradient dilution, the size of the library was measured by the
number of colonies. Twenty-four colonies were chosen to detect the
insertion rate of the library by PCR.
Selection of Nbs by phage display
The Nbs against CTLA-4 were selected by phage
display. VHH library was amplified and infected with VCSM13 helper
phages with 3 consecutive rounds of bio-panning (24). The CTLA-4 protein (20 µg) in buffer
(100 mM NaHCO3, pH 8.2) was used as an antigen to coat
microtiter plates of 96-wells at 4°C overnight. After blocking with
0.1% casein in phosphate-buffered saline (PBS) for 2 h, the wells
were incubated with displayed phages by PBS for 1 h at room
temperature. The specific phages were then eluted with 100 mM
triethylamine for 10 min and neutralized with 1.0 M Tris-HCl (pH
7.4) immediately. The exponentially growing culture of TG1 cells
(OD600=0.4–0.6) was infected with the eluted phages.
Then, they were incubated in constant temperature incubator at 37°C
for 30 min. The helper phages VCSM13 were added to rescue the
phages. The process represented one round of bio-panning and these
rescued phage particles were used in the next round of panning.
After 3 rounds, the CTLA-4-specific phages were enriched
gradually.
To obtain positive colonies, 96 individual colonies
were selected randomly for PE-ELISA and cultured in 1 ml Terrific
broth (TB) containing 100 µg/ml ampicillin for 3 h. Then, isopropyl
β-D-1-thiogalactopyranoside (IPTG) was added to induce the
expression of Nbs overnight at 28°C. The supernatant of cells was
collected after an osmotic shock and added into the plate wells
which were coated with human CTLA-4 protein in advance for 1 h,
followed by incubation with mouse anti-HA tag antibody for another
1 h, and subsequently incubated with anti-mouse IgG-alkaline
phosphatase. The chromogenic solution containing bisphosphate
(pNPP) was added after washing with PBS with 0.05% Tween-20 (PBST).
The absorbance was read using an ELISA reader at 405 nm. Finally,
the positive colonies were sequenced and classified according to
the amino acid sequence in the CDR3 region.
Expression and purification
For expressing Nbs, the recombinant phagemids were
transformed into E. coli WK6 electrocompetent cells from the
TG1 strain. These cells were cultured at 37°C in TB medium
containing 0.1% glucose, 2 M MgCl2 and ampicillin (100
µg/ml). The cultures were induced with 1 mM IPTG and incubated
overnight at 28°C when the optical density (OD) reached 0.6–1. The
periplasmic proteins were extracted by osmotic shock and then
purified by immobilized metal affinity chromatography (IMAC) by
NI-NTA superflow sepharose columns in a gradient of increasing
imidazole concentration (pH 7.0) (25). The purity of eluted proteins was
checked by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE).
Flow cytometric experiments
Human peripheral blood mononuclear cells (PBMCs)
were isolated with Ficoll-Hypaque density-gradient centrifugation
of whole blood from healthy donors included in the present study
(informed consent has been provided). The PBMCs were suspended in
RPMI-1640 medium supplemented with 10% FBS for 1 h. The nonadherent
cells were removed, and then T cells were isolated by nylon-wool
separation.
All flow cytometric experiments were performed at
4°C. PHA (1×106) (10 µg/ml) stimulated human T cells
were saturated with PBS/2% BSA solution during 30 min with shaking
to avoid nonspecific binding. Nbs (1 µg) were added to cells in
PBS/2% BSA and incubated for 30 min. After 3 washes in PBS/2% BSA,
cells were incubated for 30 min with 1 µg of PE anti-HA tag
antibody (26,27). After 3 last washes in PBS, binding
was detected by flow cytometry. Mouse anti-human CTLA-4 mAb was
used as a positive control. An irrelevant Nb (anti-CD105) served as
the negative control. Data were analyzed using FlowJo software
10.0.7 FlowJo LLC, Ashland, OR, USA).
B16 melanoma tumor challenge and
treatments
C57BL/6 mice were subcutaneously injected with
1×105 B16/BL6 melanoma cells in the right flank. On days
7, 10, 13 and 16, the mice were treated intraperitoneally with Nb16
(100 µg in 100 µl). Control groups received a corresponding dose of
anti-CTLA-4 mAb, irrelevant Nb and PBS intraperitoneally.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 6.02 (GraphPad Software, Inc., San Diego, CA, USA). Data were
analyzed by two-way analysis of variance (ANOVA) or log-rank
(Mantel-Cox) test. P<0.05 was considered statistically
significant.
Results
Construction of the VHH library
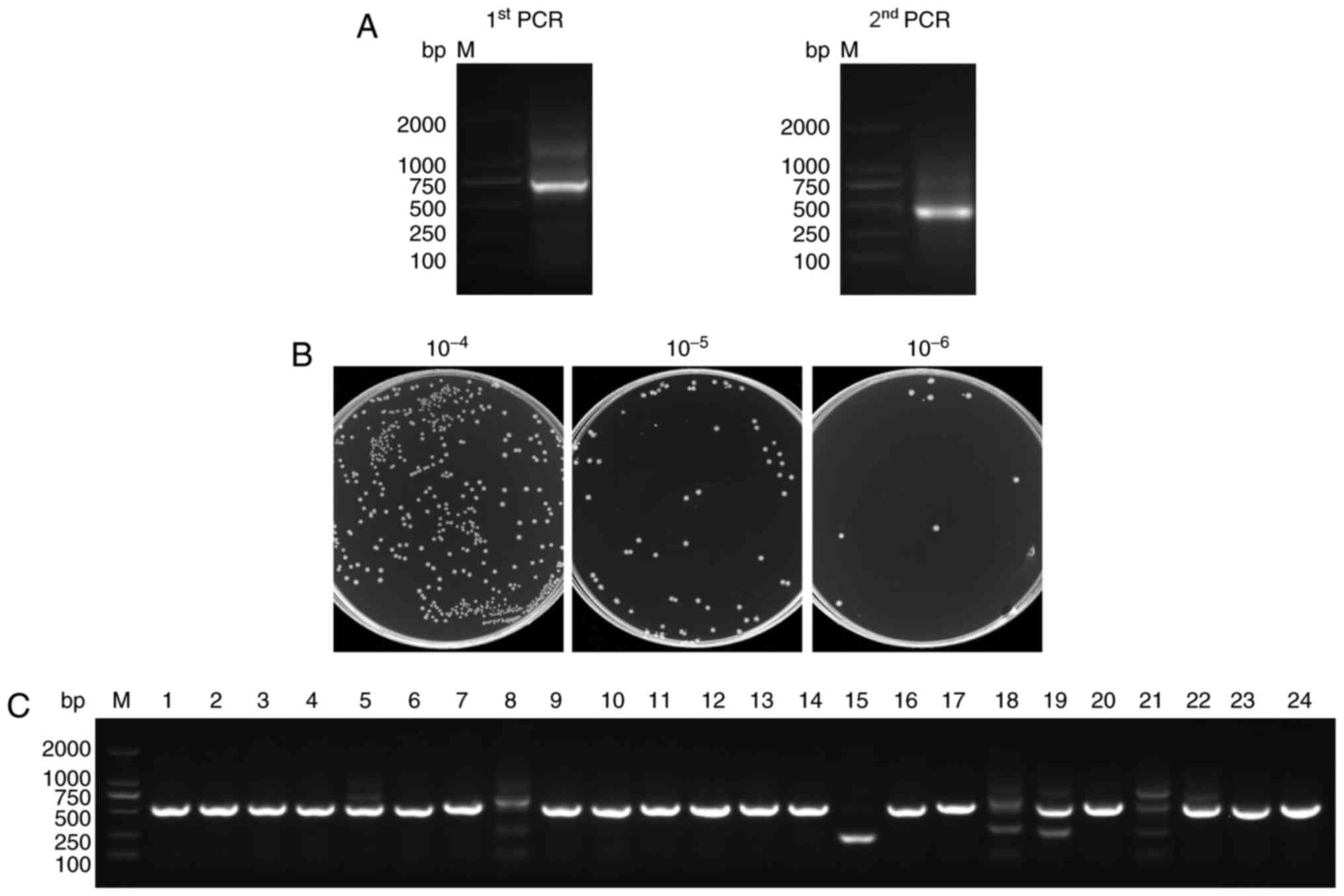
After a healthy camel was immunized with the human
CTLA-4 protein 7 times, peripheral blood lymphocytes were isolated
and the VHH genes were amplified from the lymphocyte cDNA
(Fig. 1). The first step PCR
products contained the 700 bp fragments of the VH-CH2 exons and
served as templates for the second step PCR that generated 400 bp
fragments of the VHH exons (Fig.
2A).
To construct the library, PstI and
NotI sites were introduced at the 5′ and 3′ ends of the VHH
fragments, respectively. In total, 10 µg of VHH fragments and 20 µg
of linearized pComb3 vector were used for the ligation. Then, the
recombinant plasmids were transformed into TG1 cells by 30
electroporation transformations. The size of the library was
calculated by counting the number of colonies after gradient
dilution. Its size was found to reach 1.85×108 colonies
(Fig. 2B) which enabled the
acquisition of Nbs with high specificity and sequence diversity.
Twenty-four individual colonies were selected randomly for PCR
analysis and the PCR results showed a library insertion rate of 95%
(Fig. 2C). All of these suggested
that a high-quality immunized phage display library was
successfully constructed for the subsequent selection of the
CTLA-4-specific Nbs.
Library screening and selection of
CTLA-4-specific Nbs
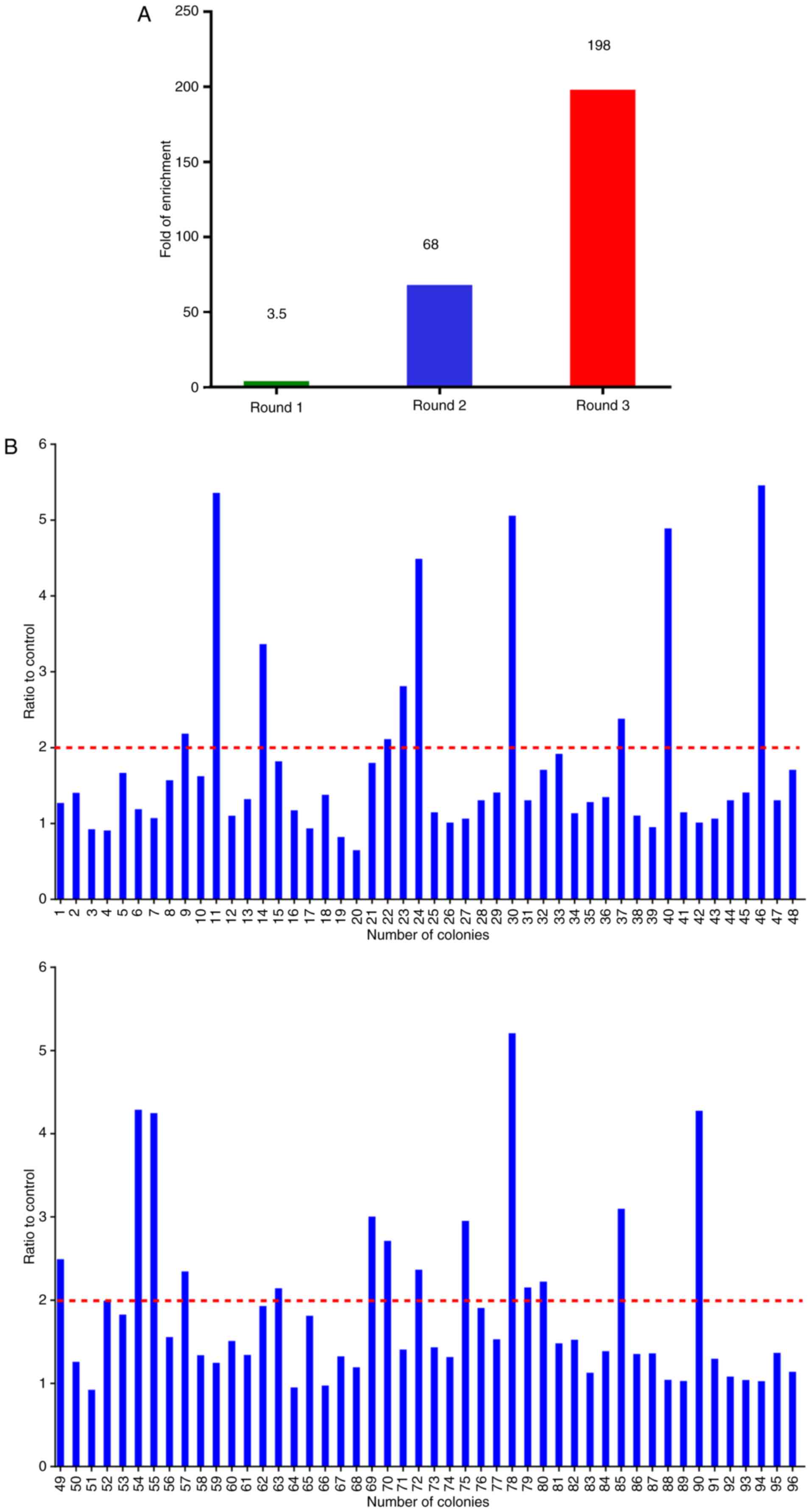
The CTLA-4-specific Nbs were identified by
bio-panning from the phage display library using ~5×1011
phages. Consecutive rounds of bio-panning were performed for
enriching the phages expressing CTLA-4-specific VHHs. After 3
rounds of panning, specific VHHs were enriched 198-fold compared
with the negative control (Fig.
3A). Subsequently, 96 colonies were randomly chosen for
PE-ELISA. Nbs existed in the supernatant of the cultured cells were
disrupted by osmotic shock (25).
Twenty-four colonies were selected as positive colonies whose
binding ratios were >2 (Fig.
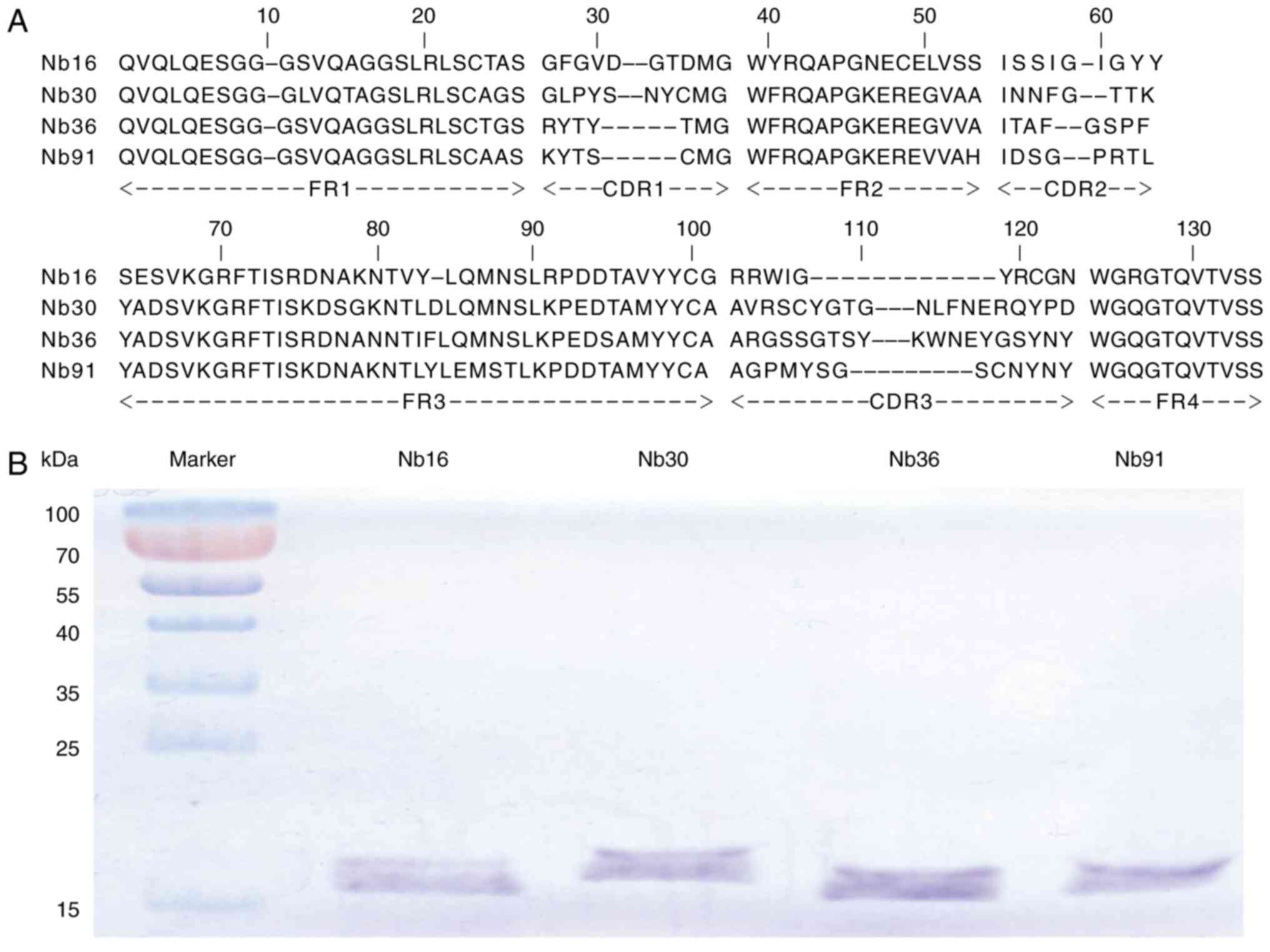
3B). After PE-ELISA, the sequences of positive colonies were
analyzed and then divided into 4 families (Nb16, Nb30, Nb36 and
Nb91) according to the variety of amino acid sequences in CDR3
(Fig. 4A) (28).
Expression and purification of the
Nbs
VHH fragments in the phage display vector pComb3
were transformed into E. coli WK6 strains. These cells
cannot suppress the amber stop codon between VHH and gene
III on pComb3. Upon IPTG induction, soluble Nbs were
expressed in the periplasmic region of WK6 cells. The induced Nbs
were further purified by NI-NTA superflow sepharose columns.
SDS-PAGE analysis showed that Nbs had single bands with high purity
(Fig. 4B). The molecular weights of
4 Nbs were 15.30, 16.41, 16.12 and 15.48 kDa, respectively.
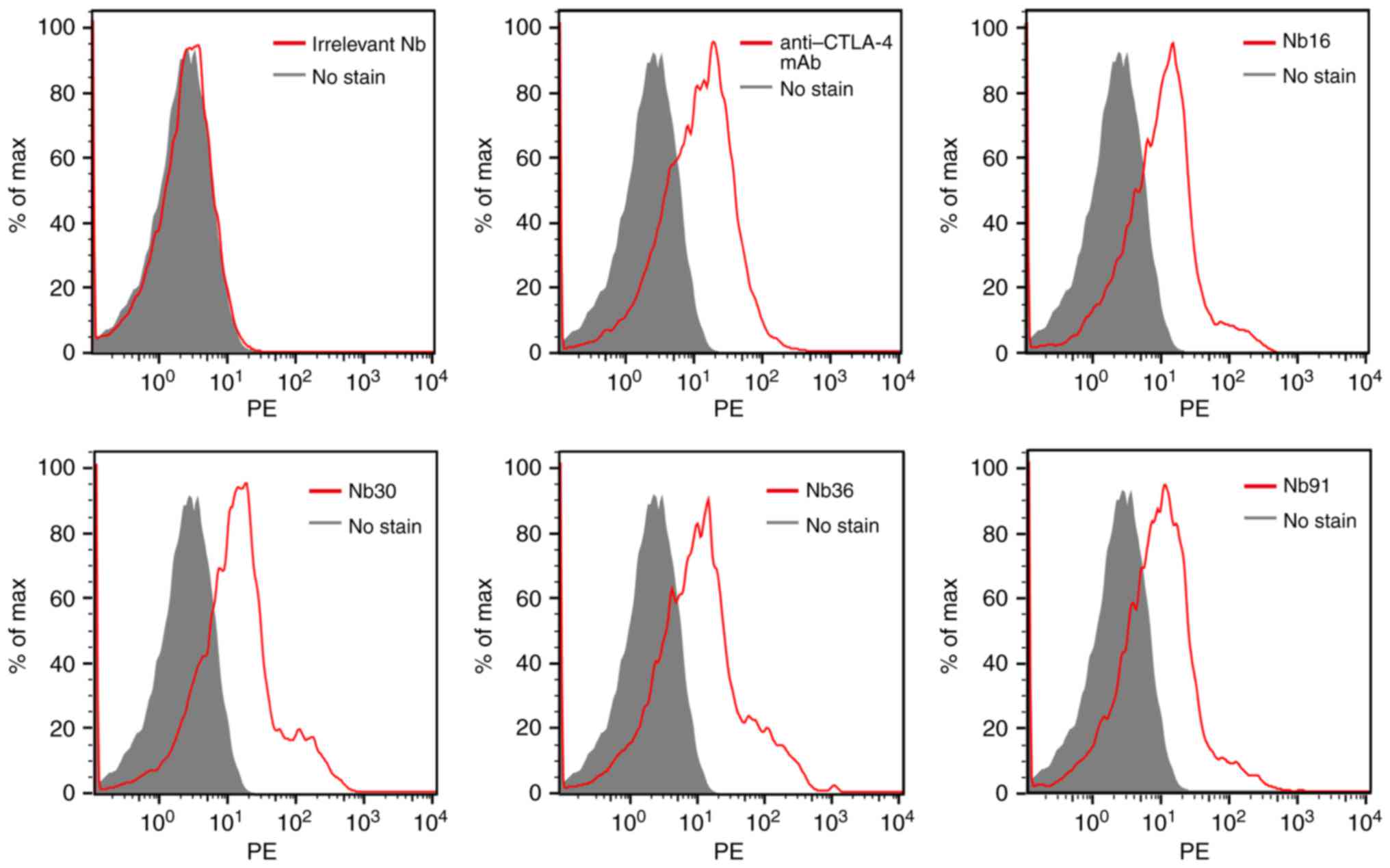
Flow cytometric analysis
To analyze the ability of the selected nanobody to
bind PHA-stimulated human T cells, flow cytometry was performed.
Our results demonstrated that all of the 4 selected Nbs were able
to recognize CTLA-4-expressing T cells, while the irrelevant Nb
could not bind to activated T cells (Fig. 5).
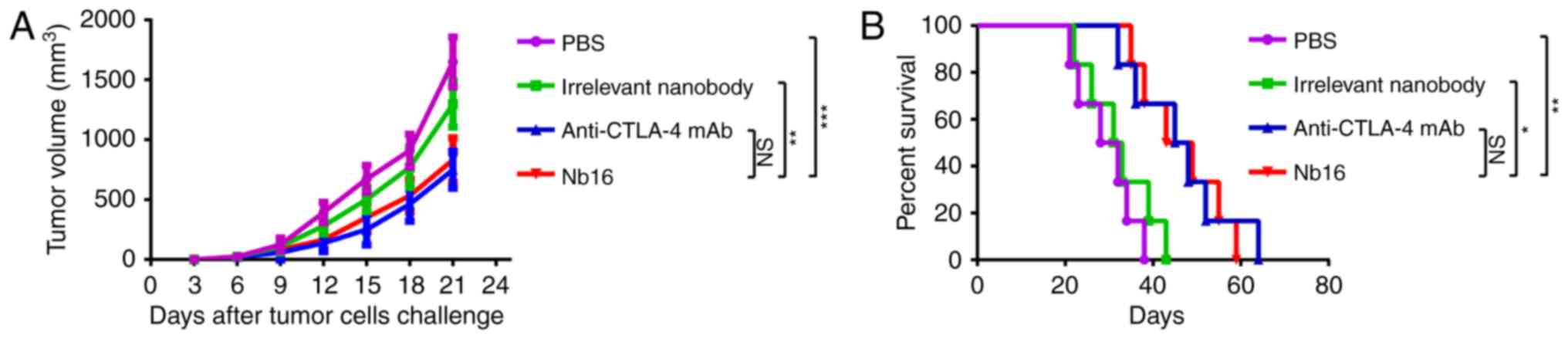
Anti-CTLA-4 Nb16 therapy is effective
against B16 melanoma tumors
To test the contribution of Nb16, C57BL/6 mice were
treated on days 7, 10, 13 and 16 with a B16/BL6 challenge
(1×105 cells). Treatment of mice with Nb16 clearly
delayed melanoma tumor growth (Fig.
6A) and prolonged the survival time of melanoma-bearing mice
(Fig. 6B).
Discussion
In the present study, CTLA-4-specific nanobodies
(Nbs) were selected from the repertoire of an immunized dromedary
camel with phage display technology. The VHH fragments encoded 4
types of CTLA-4-specific Nbs in phage plasmid pComb3 were directly
transformed into WK6 from TG1 strains. SDS-PAGE analysis showed a
band ~15 kDa after purification of soluble Nbs by NI-NTA SuperFlow
sepharose columns. We expressed and isolated Nbs of high quality.
We used anti-CTLA-4 Nbs in vitro by flow cytometry and
subsequently demonstrated that they had a good combination with PHA
stimulated and CTLA-4-positive human T cells. These Nbs showed a
strong binding ability to CTLA-4. After treating B16/BL6 melanomas,
we found that Nb16 delayed melanoma tumor growth and prolonged the
survival time of melanoma-bearing mice.
CTLA-4 is a key surface receptor on T-lymphocytes
that modulates immune responses. Antibody blockade of CTLA-4 has
been shown to enhance antitumor immune responses in murine tumor
models (29). Nbs present other
advantages including their chemical and thermal stability,
specificity, and high yield (30).
Furthermore, their substantial sequence identity (>80%) with
human VH sequences and the small size of 15 kDa make Nbs less
likely to elicit immune responses in humans. Nbs have been reported
to have low immunogenicity; humanization of Nbs has also been
previously described (31). In
clinical trials, anti-CTLA-4 antibodies have shown efficacy in
treating prostate cancer, malignant melanoma and lung cancer
(small-cell and non-small-cell) (32,33).
In summary, immunotherapy has emerged as a prominent
modality in the treatment of tumors (34). For the first time, the present study
shows that anti-CTLA-4 Nb16 has an intrinsic value for delaying
melanoma tumor. This suggests the possibility that anti-CTLA-4 Nbs
may also be advantageous for the treatment of other types of cancer
or even in the clinic (35). Next,
in subsequent research by us, it will be constructed into a
polymer, to further improve its efficacy.
Acknowledgements
The present study was supported, in part, by grants
from the Programs for Changjiang Scholars and Innovative Research
Team in the University (no. IRT_15R13), the National Natural
Scientific Foundation of China (nos. 81372452, 81430055, 81560494
and 81773254), the International Cooperation Project of the
Ministry of Science and Technology of China (no. 2015DFA31320), the
Project for Innovative Research Team in Guangxi Natural Science
Foundation (2015GXNSFFA139001), the Project of Science and
Technology of Guangxi (nos. 14125008-2-12 and 1599005-2-10), and
the Project for International Nanobody Research Center of Guangxi
(no. GuiKe-AD17195001).
References
|
1
|
Peggs KS, Quezada SA, Chambers CA, Korman
AJ and Allison JP: Blockade of CTLA-4 on both effector and
regulatory T cell compartments contributes to the antitumor
activity of anti-CTLA-4 antibodies. J Exp Med. 206:1717–1725. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zamarin D, Holmgaard RB, Subudhi SK, Park
JS, Mansour M, Palese P, Merghoub T, Wolchok JD and Allison JP:
Localized oncolytic virotherapy overcomes systemic tumor resistance
to immune checkpoint blockade immunotherapy. Sci Transl Med.
6:226ra322014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Butte MJ, Keir ME, Phamduy TB, Sharpe AH
and Freeman GJ: Programmed death-1 ligand 1 interacts specifically
with the B7-1 costimulatory molecule to inhibit T cell responses.
Immunity. 27:111–122. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Motoshima T, Komohara Y, Horlad H,
Takeuchi A, Maeda Y, Tanoue K, Kawano Y, Harada M, Takeya M and Eto
M: Sorafenib enhances the antitumor effects of anti-CTLA-4 antibody
in a murine cancer model by inhibiting myeloid-derived suppressor
cells. Oncol Rep. 33:2947–2953. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hurwitz AA, Foster BA, Kwon ED, Truong T,
Choi EM, Greenberg NM, Brug MB and Allison JP: Combination
immunotherapy of primary prostate cancer in a transgenic mouse
model using CTLA-4 blockade. Cancer Res. 60:2444–2448.
2000.PubMed/NCBI
|
|
6
|
Van Elsas A, Hurwitz AA and Allison JP:
Combination immunotherapy of B16 melanoma using anti-cytotoxic T
lymphocyte-associated antigen 4 (CTLA-4) and granulocyte/macrophage
colony-stimulating factor (GM-CSF)-producing vaccines induces
rejection of subcutaneous and metastatic tumors accompanied by
autoimmune depigmentation. J Exp Med. 190:355–366. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leach DR, Krummel MF and Allison JP:
Enhancement of antitumor immunity by CTLA-4 blockade. Science.
271:1734–1736. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hodi FS, O'Day SJ, McDermott DF, Weber RW,
Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel
JC, et al: Improved survival with ipilimumab in patients with
metastatic melanoma. N Engl J Med. 363:711–723. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ribas A, Camacho LH, Lopez-Berestein G,
Pavlov D, Bulanhagui CA, Millham R, Comin-Anduix B, Reuben JM, Seja
E, Parker CA, et al: Antitumor activity in melanoma and anti-self
responses in a phase I trial with the anti-cytotoxic T
lymphocyte-associated antigen 4 monoclonal antibody CP-675,206. J
Clin Oncol. 23:8968–8977. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fernández LÁ and Muyldermans S: Recent
developments in engineering and delivery of protein and antibody
therapeutics. Curr Opin Biotech. 22:839–842. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saerens D, Frederix F, Reekmans G, Conrath
K, Jans K, Brys L, Huang L, Bosmans E, Maes G, Borghs G and
Muyldermans S: Engineering camel single-domain antibodies and
immobilization chemistry for human prostate-specific antigen
sensing. Anal Chem. 77:7547–7555. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hassanzadeh-Ghassabeh G, Devoogdt N, De
Pauw P, Vincke C and Muyldermans S: Nanobodies and their potential
applications. Nanomedicine. 8:1013–1026. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Market E and Papavasiliou FN: V(D)J
recombination and the evolution of the adaptive immune system. PLoS
Biol. 1:E162003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Holt LJ, Herring C, Jespers LS, Woolven BP
and Tomlinson IM: Domain antibodies: proteins for therapy. Trends
Biotechnol. 21:484–490. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Muyldermans S: Nanobodies: Natural
single-domain antibodies. Annu Rev Biochem. 82:775–797. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gueorguieva D, Li S, Walsh N, Mukerji A,
Tanha J and Pandey S: Identification of single-domain, Bax-specific
intra-bodies that confer resistance to mammalian cells against
oxidative-stress-induced apoptosis. FASEB J. 20:2636–2638. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pérez JM, Renisio JG, Prompers JJ, van
Platerink CJ, Cambillau C, Darbon H and Frenken LG: Thermal
unfolding of a llama antibody fragment: A two-state reversible
process. Biochemistry. 40:74–83. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Muyldermans S: Single domain camel
antibodies: Current status. J Biotechnol. 74:277–302.
2001.PubMed/NCBI
|
|
19
|
De Genst E, Silence K, Decanniere K,
Conrath K, Loris R, Kinne J, Muyldermans S and Wyns L: Molecular
basis for the preferential cleft recognition by dromedary
heavy-chain antibodies. Proc Natl Acad Sci USA. 103:pp. 4586–4591.
2006; View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kijanka M, Warnders FJ, El Khattabi M,
Lub-de Hooge M, van Dam GM, Ntziachristos V, de Vries L, Oliveira S
and van Bergen En Henegouwen PM: Rapid optical imaging of human
breast tumour xenografts using anti-HER2 VHHs site-directly
conjugated to IRDye 800CW for image-guided surgery. Eur J Nucl Med
Mol Imaging. 40:1718–1729. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vosjan MJ, Vercammen J, Kolkman JA,
Stigter-van Walsum M, Revets H and van Dongen GA: Nanobodies
targeting the hepatocyte growth factor: Potential new drugs for
molecular cancer therapy. Mol Cancer Ther. 11:1017–1025. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
De Meyer T, Eeckhout D, De Rycke R, De
Buck S, Muyldermans S and Depicker A: Generation of VHH antibodies
against the Arabidopsis thaliana seed storage proteins. Plant Mol
Biol. 84:83–93. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Conrath KE, Lauwereys M, Galleni M,
Matagne A, Frère JM, Kinne J, Wyns L and Muyldermans S:
Beta-lactamase inhibitors derived from single-domain antibody
fragments elicited in the camelidae. Antimicrob Agents Chemother.
45:2807–2812. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vincke C, Gutiérrez C, Wernery U, Devoogdt
N, Hassanzadeh-Ghassabeh G and Muyldermans S: Generation of single
domain antibody fragments derived from camelids and generation of
manifold constructs. Methods Mol Biol. 907:145–176. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu M, Gong X, Hu Y, Ou W and Wan Y:
Streptavidin-biotin-based directional double Nanobody sandwich
ELISA for clinical rapid and sensitive detection of influenza H5N1.
J Transl Med. 12:3522014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nevoltris D, Lombard B, Dupuis E, Mathis
G, Chames P and Baty D: Conformational nanobodies reveal tethered
epidermal growth factor receptor involved in EGFR/ErbB2 predimers.
ACS Nano. 9:1388–1399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hernot S, Unnikrishnan S, Du Z, Shevchenko
T, Cosyns B, Broisat A, Toczek J, Caveliers V, Muyldermans S,
Lahoutte T, et al: Nanobody-coupled microbubbles as novel molecular
tracer. J Control Release. 158:346–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Muyldermans S, Baral T, Retamozzo VC, De
Baetselier P, De Genst E, Kinne J, Leonhardt H, Magez S, Nguyen V,
Revets H, et al: Camelid immunoglobulins and nanobody technology.
Vet Immunol Immunopathol. 128:178–183. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Curran MA, Montalvo W, Yagita H and
Allison JP: PD-1 and CTLA-4 combination blockade expands
infiltrating T cells and reduces regulatory T and myeloid cells
within B16 melanoma tumors. Proc Natl Acad Sci USA. 107:pp.
4275–4280. 2010; View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dumoulin M, Conrath K, Van Meirhaeghe A,
Meersman F, Heremans K, Frenken LG, Muyldermans S, Wyns L and
Matagne A: Single-domain antibody fragments with high
conformational stability. Protein Sci. 11:500–515. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vu KB, Ghahroudi MA, Wyns L and
Muyldermans S: Comparison of llama VH sequences from conventional
and heavy chain antibodies. Mol Immunol. 34:1121–1131. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vaneycken I, Govaert J, Vincke C,
Caveliers V, Lahoutte T, De Baetselier P, Raes G, Bossuyt A,
Muyldermans S and Devoogdt N: In vitro analysis and in vivo tumor
targeting of a humanized, grafted nanobody in mice using pinhole
SPECT/micro-CT. J Nucl Med. 51:1099–1106. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vincke C, Loris R, Saerens D,
Martinez-Rodriguez S, Muyldermans S and Conrath K: General strategy
to humanize a camelid single-domain antibody and identification of
a universal humanized nanobody scaffold. J Biol Chem.
284:3273–3284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gao Y, Gao W, Chen X, Cha N, Wang X, Jia
X, Wang B, Ren M and Ren J: Enhancing the treatment effect on
melanoma by heat shock protein 70-peptide complexes purified from
human melanoma cell lines. Oncol Rep. 36:1243–1250. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vosjan MJ, Vercammen J, Kolkman JA,
Stigter-van Walsum M, Revets H and van Dongen GA: Nanobodies
targeting the hepatocyte growth factor: Potential new drugs for
molecular cancer therapy. Mol Cancer Ther. 11:1017–1025. 2012.
View Article : Google Scholar : PubMed/NCBI
|