Introduction
Lung cancer is the most common cancer and the
leading cause of cancer-related mortality in males worldwide with
roughly 1.8 million new cases diagnosed in 2012 (13% of all
cancers) (1). Risk factors
including exposure to environmental and occupational carcinogens
have been associated with an increased incidence of lung cancer
(2). It is well established that
lung cancer is a clinically and pathologically heterogeneous
disease and has been classified into two main histological types
non-small cell lung cancer (NSCLC) including squamous cell
carcinoma, adenocarcinoma and large cell carcinoma and small cell
lung cancer based on the origin of epithelial cell precursors
(3). NSCLC accounts for ~83% of all
newly diagnosed lung cancers, and most patients (70%) are diagnosed
with advanced disease (4). Despite
treatment advancements, many patients with recurrent disease fail
to respond effectively to chemotherapy due to the development of
resistance with treatment. Following treatment overall survival
remains poor with 5-year survival estimates globally ranging from
10 to 20% (5–7). Therefore, it is critical to find new
treatment options against this deadly disease.
Natural compounds, isolated and exploited from
plants, have been reported to be involved in the modulation of
several biological processes, thus showing a great potential to be
translated into clinical use (8–10).
Moreover, various members of these families of compounds have the
ability to modulate signaling pathways as well as to regulate the
expression of genes involved in cell cycle regulation,
differentiation, and apoptosis (11). Besides being useful in prevention,
some of these molecules could also be helpful for the treatment of
cancer, especially in combination with other drugs (12,13).
Natural compounds often hold unusual structural features which
cannot be easily mimicked (14).
Therefore, such compounds may serve as an invaluable source of drug
discovery for diverse diseases. However, despite the extensive
attention focused on natural products, their exact regulatory
mechanisms remain to be explored.
Puerarin (daidzein 8-C-glucoside,
C21H20C9) is a major isoflavonoid
compound, isolated from the herb Radix Puerariae (15,16).
Previous studies have shown that puerarin has beneficial effects on
cardiovascular and cerebrovascular diseases, including coronary
artery disease, heart failure, hypertension and myocardial
infarction (17–19). In addition, recently studies have
found that puerarin has an anticancer effect on tumor cells
(20). Puerarin was found to
inhibit HT29 cell growth by enhancing the anti-proliferative
effects of other anti-neoplastic agents (21). Co-treatment of puerarin and
5-fluorouracil had a synergistic antitumor effect in gastric
carcinoma (12). Moreover, puerarin
inhibited cell growth and induced apoptosis in breast cancer and
hepatocellular carcinoma cells (22,23).
Although puerarin inhibits the cell growth and induces apoptosis in
many cancer types, there exists sparse research on the
effectiveness of puerarin on NSCLC, and the underlying mechanism of
its protective effects remains elusive. The aim of present study
was to explore the inhibitory effect of puerarin on NSCLC cells,
and elucidate the potential mechanisms of puerarin in lung
cancer.
Materials and methods
Cell culture
NCI-H441 and NCI-H460 human lung adenocarcinoma cell
lines were purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA). Cells were authenticated on the basis of
viability, recovery, growth and morphology. All cells were cultured
in RPMI-1640 medium (Sigma-Aldrich, St. Louis, MO, USA), containing
10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) at 37°C with 5% CO2 in a tissue
culture incubator. These cells are regularly tested to ensure that
they are mycoplasma-free.
Chemicals and reagents
Puerarin was obtained from Sigma-Aldrich with a
purity of 98%, as assessed by reverse-phase high-performance liquid
chromatography. Puerarin was stored at a 100 mM concentration stock
solution in dimethyl sulfoxide at −20°C and diluted with serum-free
culture medium for use in the experiments. PI3K/mTOR inhibitor
rapamycin was purchased from Sigma-Aldrich, and BEZ235 (dactolisib)
was obtained from Selleck Chemicals (Houston, TX, USA).
Cell growth inhibition studies
The inhibitory effect of puerarin on NCI-H441 or
NCI-H460 cell growth in vitro was measured by Cell Counting
Kit-8 (CCK-8; Sigma-Aldrich) according to the protocol of the
manufacturer. Briefly, NCI-H441 and NCI-H460 cells were seeded in a
96-well plate at a density of 3,000 cells/well. Then the cells were
treated with puerarin (5, 10 and 20 µM) for different time-points.
A volume of 1/10 of the medium Cell Counting Kit solution was added
to each well, and the plates were incubated for an additional 2 h
(at 37°C with 5% CO2). The optical density was then
determined at a wavelength of 450 nm using a microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Each experiment
was performed in triplicate and the results are presented as the
inhibition rate (IR), which was calculated using the following
formula: IR (%) = [(A - B)/A] × 100, where A and B are the
absorbance of the control and sample group following different
time-points of incubation, respectively.
Annexin V/PI staining
Dead Cell Apoptosis kit with Annexin V-FITC and
propidium iodide (PI) (Invitrogen, Carlsbad, CA, USA) was used
according to the manufacturer's instructions to quantify the
percentage of cells undergoing apoptosis. Briefly, NCI-H441 cells
were exposured to puerarin (5, 10 and 20 µM) for 72 h. Next, the
cells were washed twice with cold PBS and resuspended in binding
buffer at a concentration of 1×106 cells/ml. Then, 5 µl
of Annexin V-FITC and 10 µl PI were added, and the cells were
incubated for 5 min at room temperature in dark. After incubation,
200 µl of binding buffer was added and the cells were analyzed
immediately by flow cytometry (BD Biosciences, Franklin Lakes, NJ,
USA). The flow cytometric analysis was performed using CellQuest
software (BD Biosciences). Annexin V+/PI−
cells were identified as apoptotic cells, and Annexin
V−/PI+ cells were identified as necrotic
cells. The entire procedure was repeated three times for each
sample.
Western blot analysis
After treatment with puerarin for 72 h, NCI-H441
cells were lysed, and total proteins were extracted. Protein
concentration was determined using the BCA protein assay kit
(Thermo Fisher Scientific, Inc.). After quantification, 30 µg of
protein was loaded for each group, and electrophoresis was
performed on 10% polyacrylamide SDS gel. Following electrophoresis,
the gel was blotted onto polyvinylidene fluoride membranes
(Millipore, Billerica, MA, USA). After blocking in a 5% non-fat dry
milk solution in washing buffer containing 10 mmol/l Tris (pH 7.5),
150 mM NaCl, and 0.05% Tween-20, the membranes were incubated
overnight at 4°C with different antibodies: anti-Bax (#5023),
anti-Bcl2 (#3498), anti-cleaved caspase-3 (#9664), anti-Atg5
(#2630), anti-LC3 (#12741), anti-phospho-Akt (#4060), anti-Akt
(#4691), anti-phospho-ERK (#4370), anti-ERK (#9102) and
anti-β-actin (#4970) (all dilutions, 1:1,000; all from Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). After washing three
times with washing buffer containing 0.05% Tween-20, the membranes
were incubated for 2 h with horseradish peroxidase-coupled
secondary antibodies at room temperature. Signals were detected
using the ECL kit (Amersham Pharmacia Biotech; GE Healthcare,
Chicago, IL, USA).
Monodansylcadaverine (MDC)
staining
The autofluorescent agent monodansylcadaverine (MDC)
(Sigma-Aldrich) was used as a specific autophagolysosome marker to
analyze the autophagic process. After treatment with puerarin for
72 h, NCI-H441 cells were induced to autophagy by 100 nM rapamycin,
and then were incubated with 0.05 mM MDC in PBS at 37°C for 10 min.
After incubation, the cells were washed three times with PBS and
the quantitative analysis of MDC staining was assessed by flow
cytometry (BD Biosciences).
Xenografts in nude mice
Six-week-old male BALB/c nu/nu mice were purchased
from Beijing Vital River Laboratory Animal Technology Co. (Beijing,
China) and maintained under specific pathogen-free conditions. All
mice weighed 16–18 g and were bred in autoclaved, filter-top,
microisolator cages, which were kept in an isolator unit with
filtered air. The mice had access to water and food ad
libitum. The mice were inoculated subcutaneously with
1×107 NCI-H441 cells per mouse and the tumor sizes were
measured using micrometer calipers. Puerarin was solubilized in
normal saline buffer. When tumors grew to a mean volume of 100
mm3, the mice were randomly divided into the following
four groups (n=6/group): normal saline (vehicle), puerarin (25
mg/kg/day), puerarin (50 mg/kg/day), and puerarin (100 mg/kg/day),
which was injected by oral gavage daily for 5 weeks. The tumor
volume (TV) was calculated using the following formula: TV
(mm3) = d2 × (D/2), where d and D are the
shortest and longest diameters, respectively. The tumor sizes and
tumor weights of the mice were measured weekly. The mice were
sacrificed by using CO2 asphyxiation at the end of 5
weeks and the tumors were removed, weighed and measured for
additional analyses. All animal procedures, which complied with the
National Institutes of Health Guide for the Care and Use of
Laboratory Animals (24), were
approved by the Committee on Animal Experimentation of Guangxi
Normal University.
TUNEL assay
Sections of each tumor xenograft were fixed in 4%
formaldehyde, dehydrated with an ethanol gradient, embedded in
paraffin, dewaxed and rehydrated with a decreasing ethanol gradient
(100, 95, 90, 80 and 70%), according to standard instructions. An
in situ apoptosis detection kit (Roche Molecular Systems
Inc., Branchburg, NJ, USA) was used to detect apoptosis. All
procedures were performed according to the manufacturer's
instructions. The specimens were incubated with proteinase K [15
µg/ml in 10 mM Tris/HCl (pH 7.5)] for 20 min at room temperature
after being dewaxed and rehydrated. Next, the specimens were rinsed
with 3% H2O2, and incubated with
equilibration buffer and terminal deoxynucleotidyl transferase
(Beyotime Institute of Biotechnology). The specimens were then
incubated with an anti-digoxigenin-peroxidase conjugate. Finally,
the 3,3′-diaminobenzidine substrate was added to react with the
peroxidase and the specimens were counterstained with hematoxylin,
mounted and observed under a microscope.
Statistical analysis
All values are presented as the mean ± standard
deviation. Differences were determined by two-tailed Student's
t-test and one-way ANOVA using the SPSS 20.0 statistical software
(SPSS, Inc., Chicago, IL, USA). P-values of <0.05 were
indicative of a statistically significant result.
Results
Puerarin inhibits lung carcinoma cell
growth
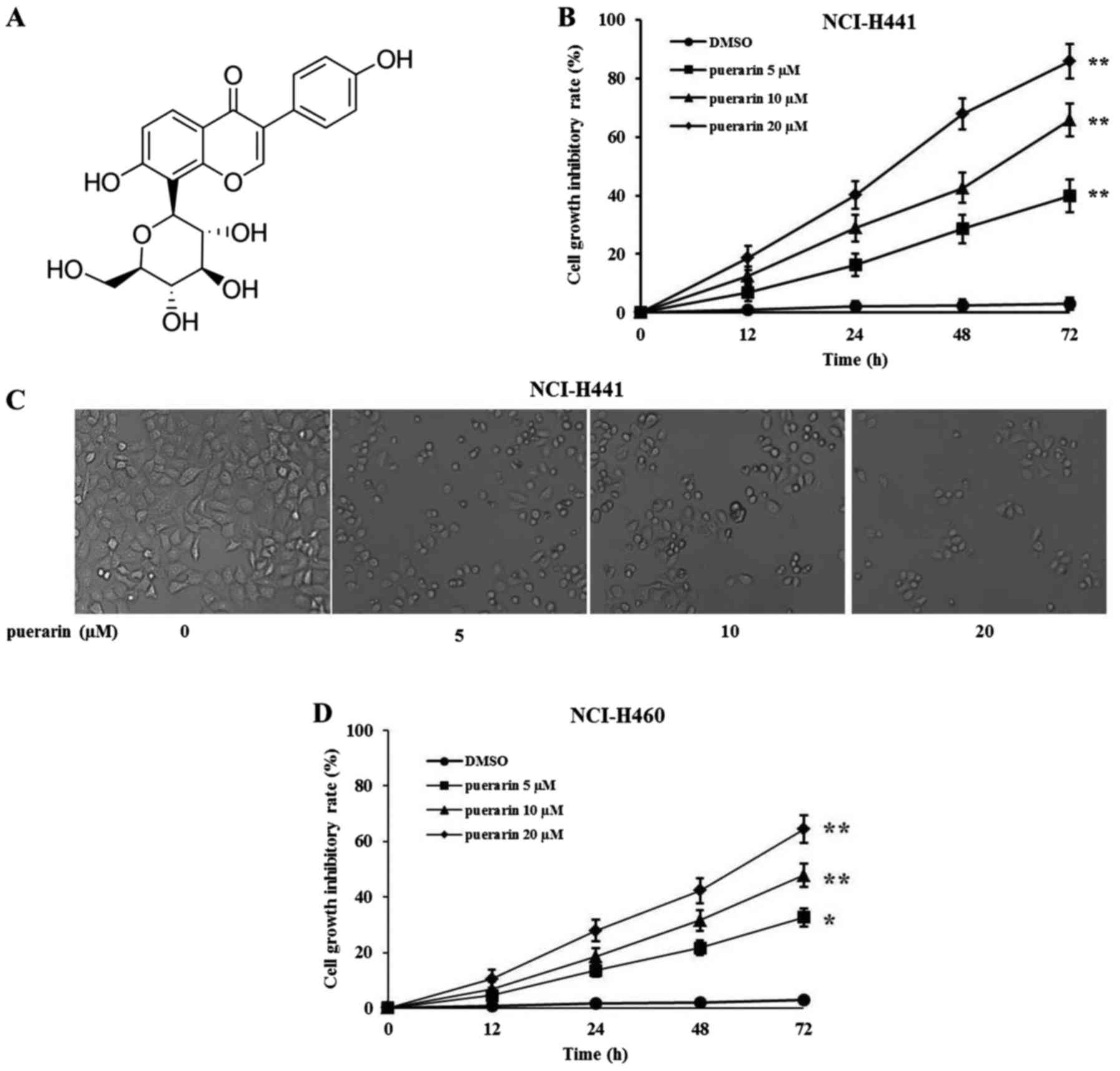
The structure of puerarin is shown in Fig. 1A. To evaluate the effects of
puerarin on cell growth, human lung adenocarcinoma NCI-H441 cells
were treated with different concentrations of puerarin, and cell
viability was determined by Cell Counting Kit-8 (CCK-8) at
different times after treatment. As shown in Fig. 1B, puerarin inhibited the growth of
NCI-H441 cells in a time- and dose-dependent manner (with
increasing concentrations from 5 to 20 µM), and showed significant
inhibition at concentrations of 10 and 20 µM after puerarin
treatment for 24, 48 and 72 h. Cell morphology and density were
also observed under a microscope after a 72-h treatment at the
concentration of 20 µM. Consistent with the cell growth results,
there was a significant reduced cell density in the
puerarin-treated cells compared with the vehicle-treated cells
(Fig. 1C). Moreover, cell
morphology turned from a round shape into a spreading shape with
good morphology, indicating that puerarin may not only inhibit cell
growth, but also induce cell apoptosis. Similarly, puerarin also
inhibited the growth of NCI-H460 cells in a time- and
dose-dependent manner (Fig. 1D).
Compared with the NCI-H460 cell line, NCI-H441 was more sensitive
to puerarin. Therefore, the NCI-H441 cell line was selected for the
following experiments.
Puerarin induces lung cancer cell
apoptosis
Apoptosis is one of the predominant types of
programmed cell death which involves a series of biochemical events
leading to specific cell morphologic characteristics, including
cell shrinkage, nuclear fragmentation, chromatin condensation and
chromosomal DNA fragmentation. As observed in Fig. 1A and B, NCI-H441 cells appeared to
exhibit apoptosis characteristics after puerarin treatment, thus,
flow cytometric analysis by Annexin V and PI staining was performed
to examine the proportion of apoptotic cells. Data indicated that
puerarin treatment significantly increased the number of early
apoptotic cells (Annexin V+/PI−) and late
apoptotic cells (Annexin V+/PI−) in a
dose-dependent manner (Fig. 2A and
B). In order to verify the apoptotic mechanism, the expression
of the mitochondrial pathway proteins Bax and Bcl-2 were detected.
As shown in Fig. 2C, the expression
of Bax was upregulated along with increasing concentrations of
puerarin. In contrast, the levels of Bcl-2 were decreased, leading
to an increase in cleaved caspase-3. The relative protein levels
normalized by β-actin are shown in Fig.
2D. Similarly, puerarin induced apoptosis in the NCI-H460 cells
in a dose-dependent manner (Fig. 2E and
F).
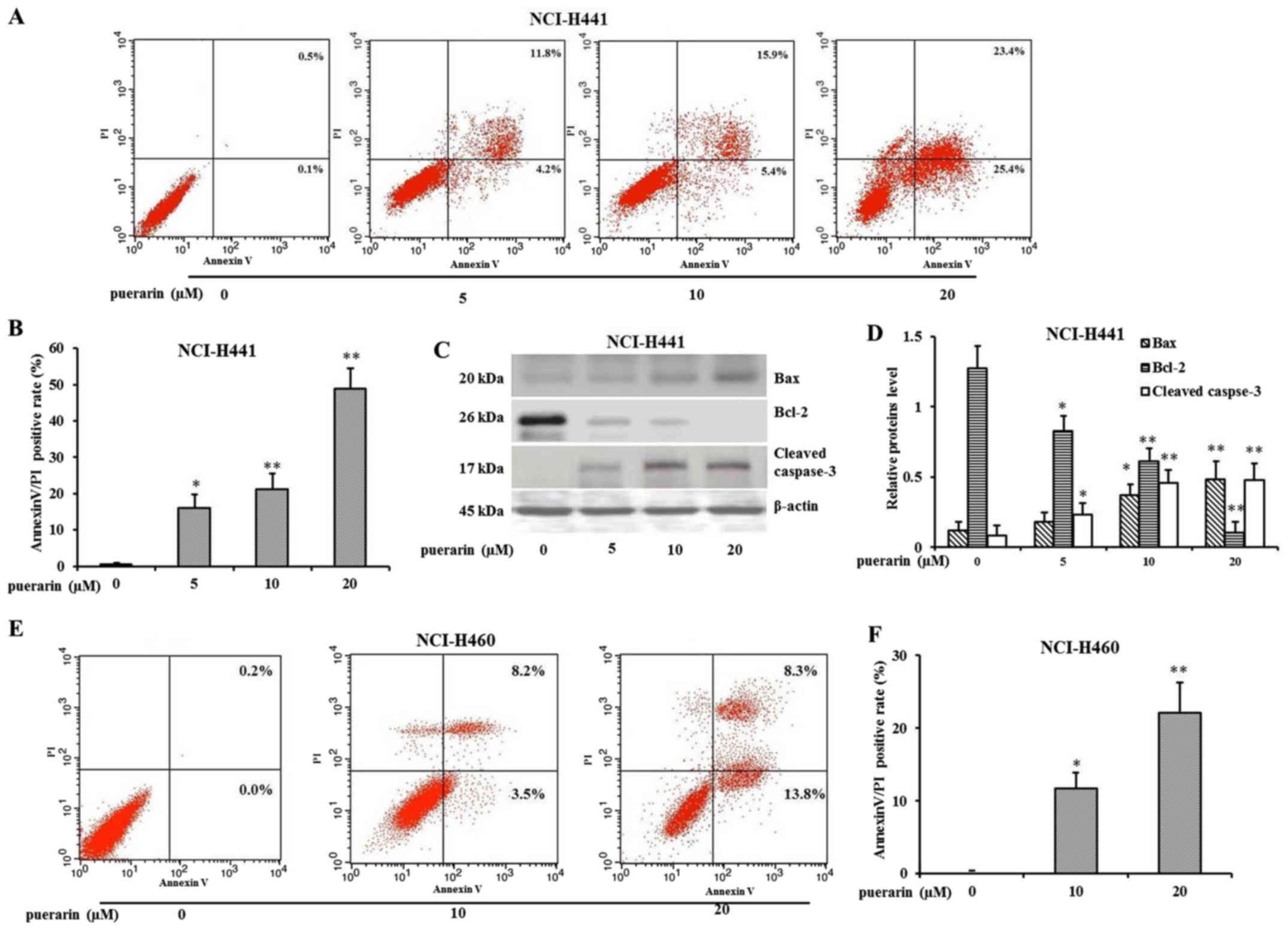 | Figure 2.Puerarin promotes apoptosis in
NCI-H441 or NCI-H460 cells. (A) NCI-H441 cells were treated with
different concentrations of puerarin (5, 10 and 20 µM) for 72 h.
The apoptosis was determined by Annexin V/PI staining, and then was
analyzed by flow cytometry (x-axis, Annexin V; y-axis, PI). (B) The
quantification of the results of Annexin V/PI staining for NCI-H441
cells (*P<0.05, **P<0.01, compared with DMSO control, n=3).
(C) Western blot analysis of Bax, Bcl-2 and cleaved caspase-3
protein following exposure to puerarin. (D) Relative protein levels
were quantified by using β-actin as control (*P<0.05,
**P<0.01, compared with DMSO control, n=3). (E) NCI-H460 cells
were treated with different concentrations of puerarin (0, 10 and
20 µM) for 72 h. The apoptosis was determined by Annexin V/PI
staining, and then was analyzed by flow cytometry (x-axis, Annexin
V; y-axis, PI). (F) The quantification of the results of Annexin
V/PI staining for NCI-H460 cells (*P<0.05, **P<0.01, compared
with DMSO control, n=3). |
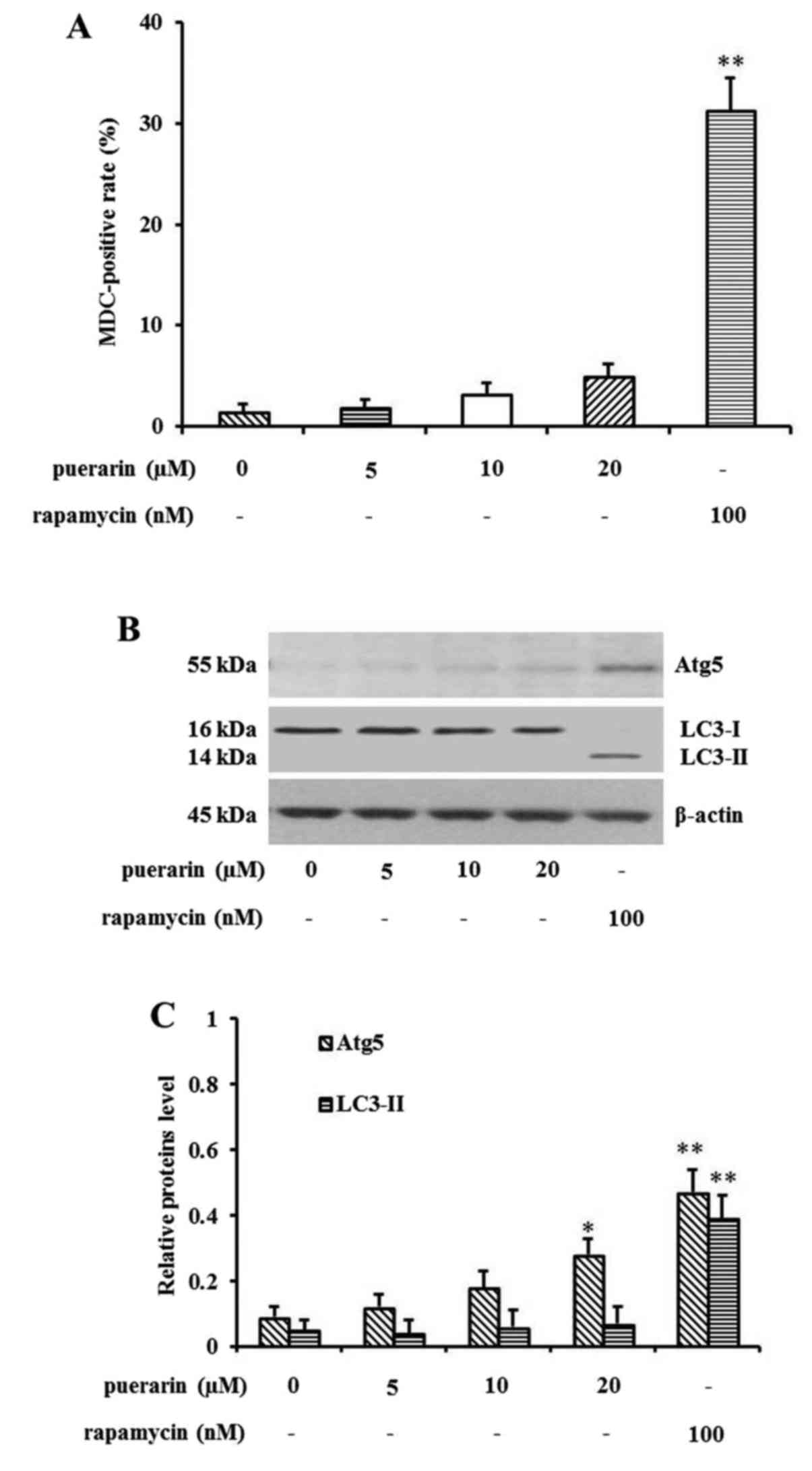
Puerarin slightly induces NCI-H441
cell autophagy
Since it has been reported that puerarin attenuates
anoxia/reoxygenation injury (25)
or prevents cardiac hypertrophy (26) through activation of autophagy, we
investigated the relationship between puerarin and autophagy in
tumor cells. Therefore, we detected the effect of puerarin
treatment on NCI-H441 cell autophagy by using MDC staining assay.
The positive staining of autophagosomes was measured via flow
cytometry and the MDC-positive rate was calculated (Fig. 3A). There was a slight increase in
the MDC-positive rate following puerarin treatment (20 µM) compared
with the control group. Rapamycin, a specific inhibitor of mTOR,
was used as a positive control of autophagy. To further confirm
puerarin-induced NCI-H441 cell autophagy, western blot analysis was
performed to detect the alteration of essential molecules involved
in autophagy. The expression of Atg5, autophagy protein 5, was
gradually upregulated in a concentration-dependent manner. In
contrast, the level of LC3-I, microtubule-associated protein 1
light chain 3 type I, was decreased. However, the expression of
LC3-II, which correlates with the extent of autophagosomes, was not
detectable in this cell line. Cells treated with rapamycin showed a
strong signal of Atg5 and LC3-II, used as a positive control of
autophagy (Fig. 3B and C).
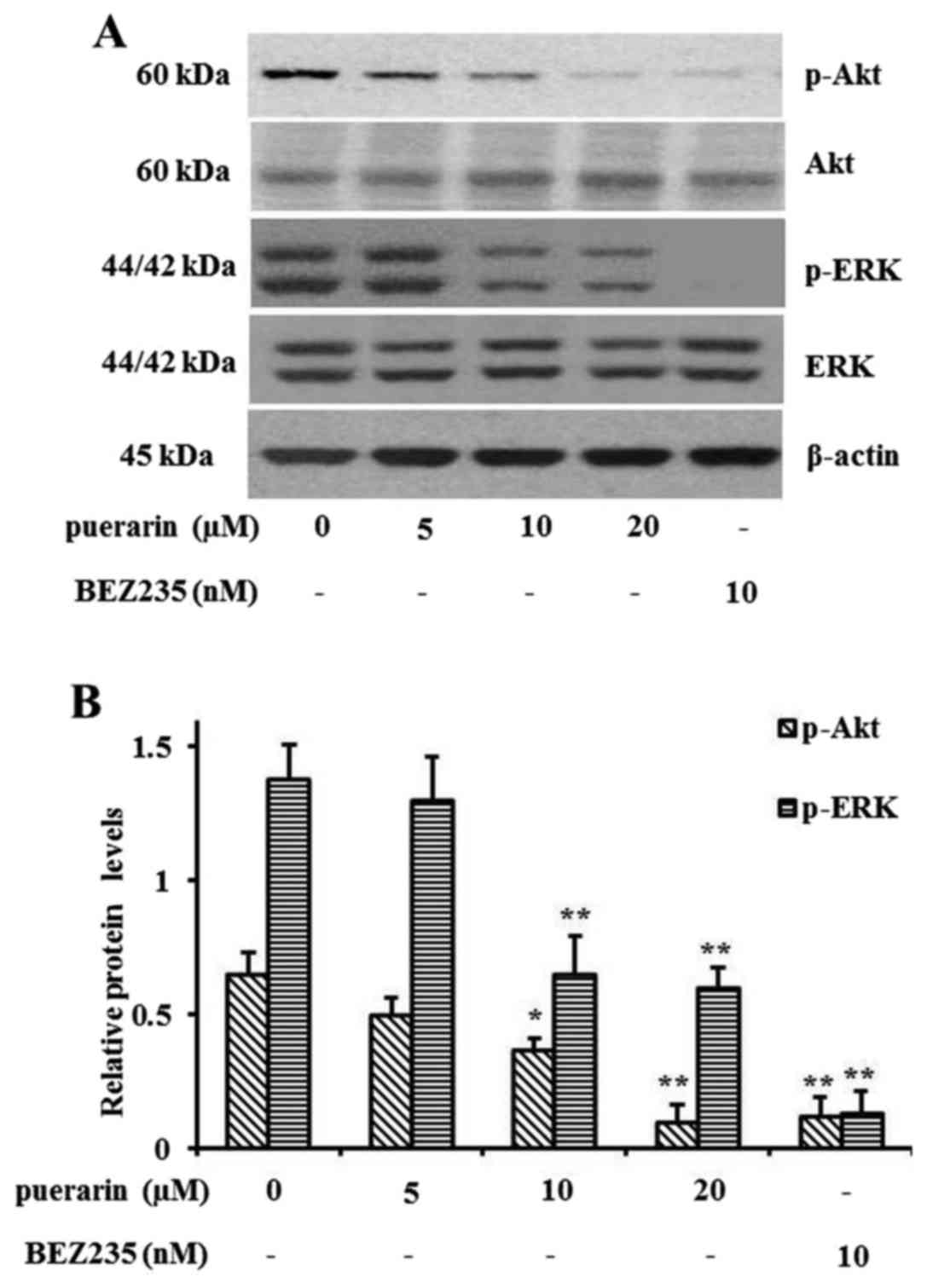
Puerarin induces cell autophagy via
the PI3K/Akt and ERK pathway
Since puerarin activated NCI-H441 cell autophagy,
PI3K/Akt and ERK pathway activation, which are related to cell
autophagy, was also determined by western blot analysis. As shown
in Fig. 4A, the phosphorylation
levels of Akt and ERK were extremely reduced after puerarin
treatment. However, the total protein levels were not affected.
BEZ235, an inhibitor of PI3K/mTOR, was used as a control for the
inhibition of phosphorylation. The relative protein levels
normalized by total Akt or ERK expression are shown in Fig. 4B.
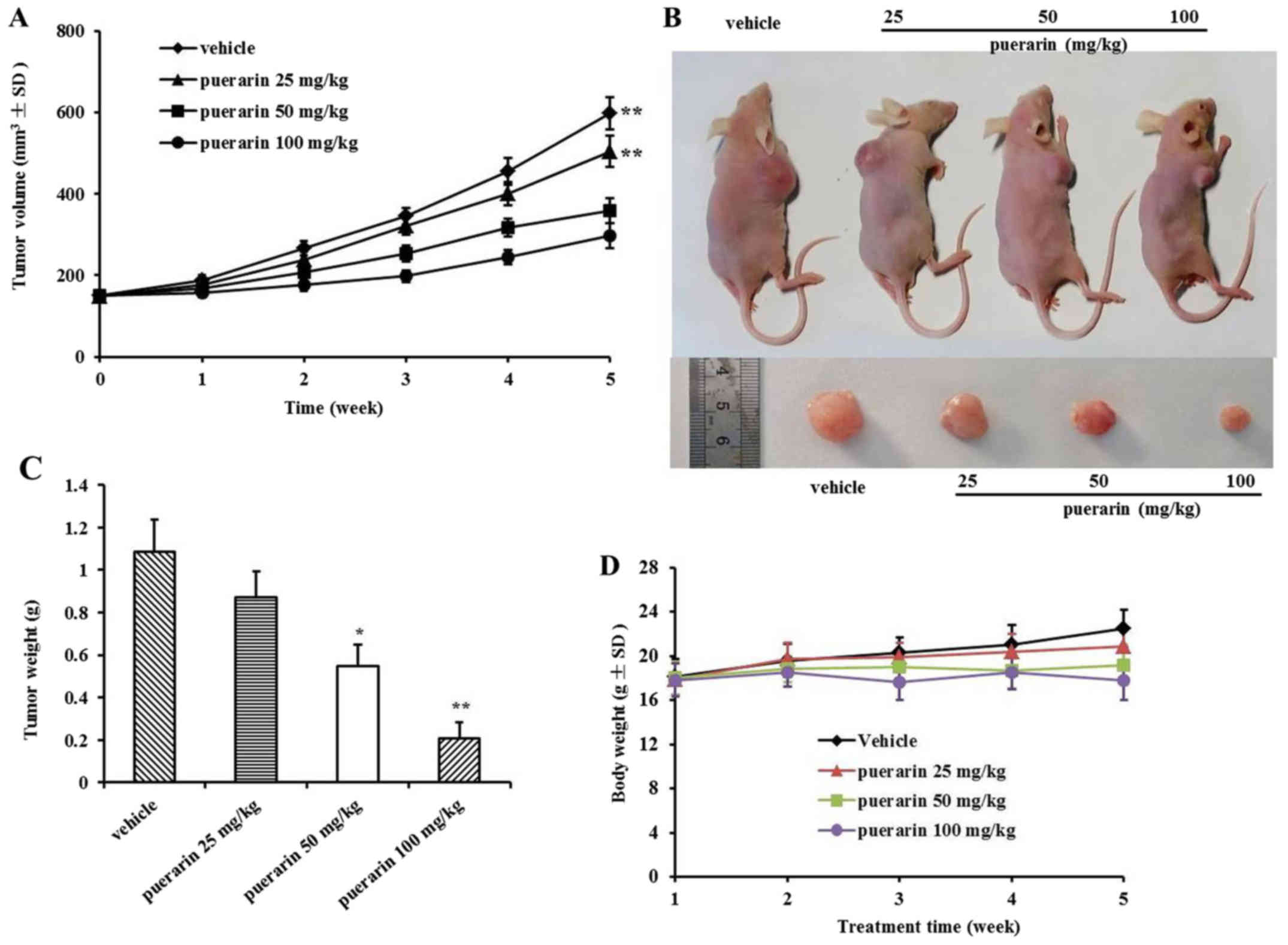
Puerarin prevents tumor development in
vivo
The effect of puerarin on the growth of primary
tumor xenografts in nude mice was examined. Tumor volume was
recorded every week. The volumes of the tumors in the treatment
groups were significantly reduced in a dose-dependent manner
compared with those in the control group, while the inhibition rate
in the 100 mg/kg of puerarin treatment group was more highly
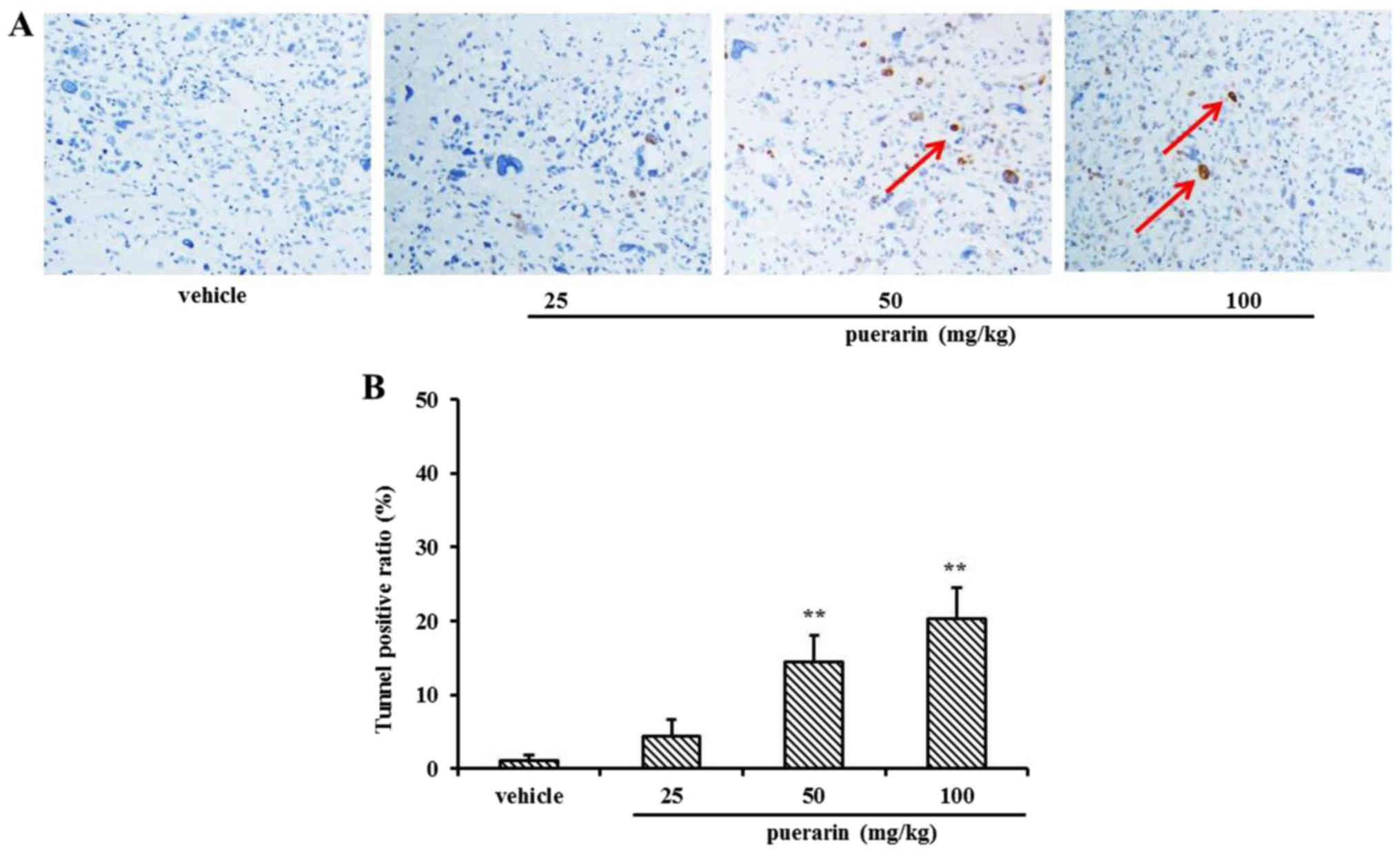
significant than that in the other three groups (Fig. 5A). Representative figures of tumor
dimensions in nude mice at the end of the experiment are shown in
Fig. 5B. At the end of the
experiment, the tumors were harvested and weighed in each group
(Fig. 5C). In addition, there was
no significant body weight loss in each group, which indicated that
puerarin treatments were tolerable.
Detection of apoptotic cells in the
xenograft tumor tissues
At the end of the experiment, 24 mice were
necropsied, and the subcutaneous tumor tissue sections were used
for detecting apoptotic cells by TUNEL assay. As shown in Fig. 6A, significant cell death had
occurred in the tumor masses in the puerarin treatment groups, when
compared with that of the vehicle group. The TUNEL-positive ratio
was calculated by counting the positive-stained cells, which was
then divided by the total cell number (Fig. 6B).
Discussion
Puerarin is the most important phytoestrogen
extracted from the dried root of Pueraria lobata, which is a
commonly used traditional Chinese herb. Researchers have
concentrated on the pharmacological activities of puerarin, which
displays a series of beneficial activities on hangover,
cardiovascular disease, osteoporosis, neurological dysfunction,
fever, and liver injury in clinical treatment and experimental
research (27). In addition, more
and more studies focus on the role of puerarin in cancer: puerarin
inhibited cell growth and induced apoptosis in breast cancer cells
(22), prevented human oophoroma
cell invasion and metastasis (28),
and induced neuroblastoma SH-SY5Y cell death by activating the
PI3K/Akt pathway (29).
In the present study, we conducted an investigation
of the effects of puerarin on NSCLC NCI-H441 cells. Puerarin
inhibited cell growth in a dose and time-dependent manner, and
promoted apoptosis as determined by Annexin V/PI staining. In
addition, puerarin treatment increased the active form of caspase-3
(cleaved caspase-3), with an increase in Bax expression and a
decrease in Bcl-2 levels. The Bcl-2 family of proteins plays an
important role in the regulation of apoptosis in many cellular
systems, by either inhibiting (Bcl-2, Bcl-XL, Bcl-W, Bfl-1, and
Mcl-1) or promoting apoptosis (Bax, Bak, Bad, Bcl-Xs, Bid, and Hrk)
(30). Heterodimerization between
pro- and anti-apoptotic members of this family and relative levels
of both types of proteins may determine the susceptibility to a
given apoptotic stimulus and cell fate (31,32).
Moreover, a high concentration of puerarin induced
NCI-H441 cell autophagy as assessed by MDC staining, and caused a
slight increase in Atg5 expression and reduction in the LC3-I
protein level. Notably, there was no detectable alteration of
LC3-II expression compared with the control. During autophagy, the
nascent proMAP1LC3/LC3 (microtubule-associated protein 1 light
chain 3) is processed post-translationally into the cytoplasmatic
soluble form LC3-I and by conjugation to phosphatidylethanolamine
(PE), generating the membrane-associated LC3-II puncta, which
correlates with the extent of autophagosomes (33). For this reason, LC3 is a
commonly-used autophagic marker (34). We failed to detect a change in
LC3-II expression, perhaps because the samples used for this
experiment were in the early stage of autophagy. To confirm our
hypothesis, we analyzed the phosphorylation levels of Akt and ERK,
since it is known that PI3K/Akt and ERK pathway inactivation is
required during cell autophagy. Indeed, the phosphorylation levels
of Akt and ERK were reduced in a dose-dependent manner, further
indicating that puerarin treatment induced NCI-H441 cell autophagy.
In addition, puerarin prevented tumor growth by inducing tumor cell
apoptosis in the xenograft nude mouse model. This result was
consistent with in vitro data.
Taken together, puerarin, inhibited cell growth,
induced apoptosis and autophagy and could be considered as a
possible chemotherapeutic drug for the treatment of lung
cancer.
Acknowledgements
This study was supported by the Key Laboratory for
Chemistry and Molecular Engineering of Medicinal Resources (Guangxi
Normal University) (CMEMR2016-B01).
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Świątkowska B, Szubert Z, Sobala W and
Szeszenia-Dąbrowska N: Predictors of lung cancer among former
asbestos-exposed workers. Lung Cancer. 89:243–248. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Devesa SS, Bray F, Vizcaino AP and Parkin
DM: International lung cancer trends by histologic type:
male:female differences diminishing and adenocarcinoma rates
rising. Int J Cancer. 117:294–299. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Howlader N, Noone AM, Krapcho M, Garshell
J, Miller D, Altekruse SF, Kosary CL, Yu M, Ruhl J, Tatalovich Z,
Mariotto A, Lewis DR, Chen HS, Feuer EJ and Cronin KA: SEER Cancer
Statistics Review, 1975–2012. National Cancer Institute; Bethesda,
MD: http://seer.cancer.gov/csr/1975_2012/based on
November 2014 SEER data submission, posted to the SEER web site.
April. 2015
|
|
5
|
Allemani C, Weir HK, Carreira H, Harewood
R, Spika D, Wang XS, Bannon F, Ahn JV, Johnson CJ, Bonaventure A,
et al CONCORD Working Group, : Global surveillance of cancer
survival 1995–2009: Analysis of individual data for 25,676,887
patients from 279 population-based registries in 67 countries
(CONCORD-2). Lancet. 385:977–1010. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gupta SK: Role of Crizotinib in previously
treated non-small-cell lung cancer. South Asian J Cancer.
3:138–140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schiller JH, Harrington D, Belani CP,
Langer C, Sandler A, Krook J, Zhu J and Johnson DH; Eastern
Cooperative Oncology Group, : Comparison of four chemotherapy
regimens for advanced non-small-cell lung cancer. N Engl J Med.
346:92–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Angulo P, Kaushik G, Subramaniam D,
Dandawate P, Neville K, Chastain K and Anant S: Natural compounds
targeting major cell signaling pathways: A novel paradigm for
osteosarcoma therapy. J Hematol Oncol. 10:102017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chanvorachote P, Chamni S, Ninsontia C and
Phiboonchaiyanan PP: Potential anti-metastasis natural compounds
for lung lancer. Anticancer Res. 36:5707–5717. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shanmugam MK, Lee JH, Chai EZ, Kanchi MM,
Kar S, Arfuso F, Dharmarajan A, Kumar AP, Ramar PS, Looi CY, et al:
Cancer prevention and therapy through the modulation of
transcription factors by bioactive natural compounds. Semin Cancer
Biol 40–41. 1–47. 2016.
|
|
11
|
Pan MH, Lai CS, Wu JC and Ho CT: Molecular
mechanisms for chemoprevention of colorectal cancer by natural
dietary compounds. Mol Nutr Food Res. 55:32–45. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo XF, Yang ZR, Wang J, Lei XF, Lv XG and
Dong WG: Synergistic antitumor effect of puerarin combined with
5-fluorouracil on gastric carcinoma. Mol Med Rep. 11:2562–2568.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zeng YP, Yang ZR, Guo XF, Jun W and Dong
WG: Synergistic effect of puerarin and 5-fluorouracil on
hepatocellular carcinoma. Oncol Lett. 8:2436–2442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang SF, Wang XL, Yang XQ and Chen N:
Autophagy-associated targeting pathways of natural products during
cancer treatment. Asian Pac J Cancer Prev. 15:10557–10563. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yanagihara K, Ito A, Toge T and Numoto M:
Antiproliferative effects of isoflavones on human cancer cell lines
established from the gastrointestinal tract. Cancer Res.
53:5815–5821. 1993.PubMed/NCBI
|
|
16
|
Tian F, Xu LH, Zhao W, Tian LJ and Ji XL:
The optimal therapeutic timing and mechanism of puerarin treatment
of spinal cord ischemia-reperfusion injury in rats. J
Ethnopharmacol. 134:892–896. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wong KH, Li GQ, Li KM, Razmovski-Naumovski
V and Chan K: Kudzu root: Traditional uses and potential medicinal
benefits in diabetes and cardiovascular diseases. J Ethnopharmacol.
134:584–607. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan Y, Zong J, Zhou H, Bian ZY, Deng W,
Dai J, Gan HW, Yang Z, Li H and Tang QZ: Puerarin attenuates
pressure overload-induced cardiac hypertrophy. J Cardiol. 63:73–81.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yeung DK, Leung SW, Xu YC, Vanhoutte PM
and Man RY: Puerarin, an isoflavonoid derived from Radix puerariae,
potentiates endothelium-independent relaxation via the cyclic AMP
pathway in porcine coronary artery. Eur J Pharmacol. 552:105–111.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu Z and Li W: Induction of apoptosis by
puerarin in colon cancer HT-29 cells. Cancer Lett. 238:53–60. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Ma Y, Zheng Y, Song J, Yang X, Bi
C, Zhang D and Zhang Q: In vitro and in vivo anticancer activity of
a novel puerarin nanosuspension against colon cancer, with high
efficacy and low toxicity. Int J Pharm. 441:728–735. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang WG, Liu XF, Meng KW and Hu SY:
Puerarin inhibits growth and induces apoptosis in SMMC-7721
hepatocellular carcinoma cells. Mol Med Rep. 10:2752–2758. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin YJ, Hou YC, Lin CH, Hsu YA, Sheu JJ,
Lai CH, Chen BH, Lee Chao PD, Wan L and Tsai FJ: Puerariae radix
isoflavones and their metabolites inhibit growth and induce
apoptosis in breast cancer cells. Biochem Biophys Res Commun.
378:683–688. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
National Research Council of the National
Academies, . Guide for the Care and Use of Laboratory Animals:
2011. The National Academies Press; Washington, DC: 2011
|
|
25
|
Ma Y, Gai Y, Yan J, Li J and Zhang Y:
Puerarin attenuates anoxia/reoxygenation injury through enhancing
Bcl-2 associated athanogene 3 Expression, a modulator of apoptosis
and autophagy. Med Sci Monit. 22:977–983. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu B, Wu Z, Li Y, Ou C, Huang Z, Zhang J,
Liu P, Luo C and Chen M: Puerarin prevents cardiac hypertrophy
induced by pressure overload through activation of autophagy.
Biochem Biophys Res Commun. 464:908–915. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei SY, Chen Y and Xu XY: Progress on the
pharmacological research of puerarin: A review. Chin J Nat Med.
12:407–414. 2014.PubMed/NCBI
|
|
28
|
Han J, Yu CQ and Shen W: Inhibitory
effects of puerarin on invasion and metastasis of oophoroma cells
HO-8910. Zhongguo Zhong Xi Yi Jie He Za Zhi. 29:632–635. 2009.(In
Chinese). PubMed/NCBI
|
|
29
|
Zhu G, Wang X, Wu S and Li Q: Involvement
of activation of PI3K/Akt pathway in the protective effects of
puerarin against MPP+-induced human neuroblastoma
SH-SY5Y cell death. Neurochem Int. 60:400–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Choi YH, Kong KR, Kim YA, Jung KO, Kil JH,
Rhee SH and Park KY: Induction of Bax and activation of caspases
during beta-sitosterol-mediated apoptosis in human colon cancer
cells. Int J Oncol. 23:1657–1662. 2003.PubMed/NCBI
|
|
33
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|