Introduction
Gastric cancer is a malignant disease which has a
high mortality rate and is the second leading cause of
cancer-related deaths worldwide (1). Although recent advances in surgery,
chemotherapy, and radiation therapy have improved the outcome of
gastric cancer, patients who undergo curative surgical resection
often succumb to recurrent disease due to tumor invasion and
metastasis. Hence, to treat gastric cancer, it is necessary to
elucidate the molecular mechanisms involved in the development and
invasiveness of gastric cancer.
Epithelial-mesenchymal transition (EMT) is a
physiological and pathological process where epithelial cells lose
their polarity and cell-cell adhesion signature, and acquire the
characteristics of mesenchymal cells (2). EMT plays a critical role in many
aspects of cancer behavior, including phenotypic conversion that is
implicated in the initiation of metastasis in gastric cancer
progression (3). During EMT, the
expression of some epithelial cell markers, such as E-cadherin
decreases, while the expression of mesenchymal cell markers, such
as vimentin increases. In addition, the EMT-related transcription
factor, SOX10 (4), is upregulated,
during this process. The Wnt/β-catenin signaling pathway is also
associated with EMT; in fact, aberrant β-catenin subcellular
localization expression is considered a surrogate marker of
aberrant Wnt signaling pathway activation.
REC8 is a key meiosis-specific component of the
cohesion complex that binds sister chromatids in preparation for
the two divisions of meiosis. REC8 has also been implicated in DNA
damage repair and maintenance of chromosome stability (5). Previous studies revealed that REC8 was
hypermethylated in melanomas and malignant gastrointestinal stromal
tumors. Moreover, REC8 was associated with poor tumor prognosis in
patients (6–8). The novel role of REC8 as a
tumor-suppressor gene was recently identified through the robust
epigenetic inactivation of REC8 by PI3K. REC8 was also revealed to
play an important role in many human cancers (9). These results indicated that REC8 has
the potential to suppress tumors in gastric cancer.
We thus hypothesized that REC8 and EMT are involved
in the progression of gastric cancer. We determined that REC8
inhibited the proliferation, migration and invasion of gastric
cancer cells in vitro. Furthermore, we used genome-wide
expression microarray to analyze the association of REC8 expression
with differential gene expression profiles and determined that the
expression of early growth response-1 (EGR1), a transcription
factor mediated by the AGE-RAGE signaling pathway (10) was regulated by REC8. However, the
role of EGR1 on EMT remains unclear due to opposite functions that
have been reported in studies (11,12).
In the present study, we determined the function of
REC8 by assessing gain- and loss-of-function of REC8 and its effect
on EMT in gastric cancer cells. Moreover, we revealed that REC8
expression regulated the expression of EGR1 and mediated EMT.
Finally, we assessed the manner in which REC8 regulated the
expression of EGR1, specifically through its interaction.
Therefore, we demonstrated the effects of REC8 on gastric cancer
cell proliferation and migration, and revealed that REC8 inhibited
cell EMT by downregulating EGR1 in gastric cancer via REC8 and ERG1
interactions.
Materials and methods
Cell culture
The BGC823 and SGC-7901 cell lines were obtained
from the Cell Bank of the Chinese Academy of Sciences (Chinese
Academy of Sciences, Shanghai, China). Cells were routinely grown
in RPMI-1640 medium supplemented with 10% (v/v) fetal bovine serum
(FBS) at 37°C in a humidified atmosphere of 5% CO2.
REC8 shRNA, REC8 and EGR1
overexpression constructs and infection
REC8 shRNA was purchased from Origene Technologies,
Inc. (cat. no. TG309883; Rockville, MD, USA). For the
overexpression of REC8 and EGR1, full-length homo sapiens REC8 and
EGR1 sequences (based on NCBI) were cloned into the pcDNA3.1
vector. The shRNA plasmid pcDNA3.1-REC8 or pcDNA3.1-EGR1 were
transiently transfected into cells using Lipofectamine 2000 reagent
(cat. no. 11668019; Invitrogen/Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) according to the manufacturers instructions.
Cell proliferation analysis
Cells (1×104) transfected with vector or
pcDNA3.1-REC8 were seeded into 96-well culture plates and cultured
for 72 h. At 0, 24, 48 and 72 h, CCK-8 reagent was added to each
well, according to the manufacturers instructions (cat. no. 96992;
Sigma-Aldrich, St. Louis, MO, USA). The optical density values (OD
values) were assessed at 450 nm using a plate reader (Thermo
Scientific™Multiskan™ GO; Thermo Fisher Scientific).
Wound healing assay
Cells (1×105) were seeded and cultured in
a 24-well plate. After the cells were confluent, the cell layer was
carefully wounded using sterile tips (200 µl). The falling cells
were washed off with phosphate-buffered saline (PBS). After
incubation for 0 and 48 h in serum-free medium, the images of the
cells were captured by light microscope at a low magnification (4×,
objective) and wound healing was assessed.
Transwell invasion assays
The precoated Transwell filters (8-µm pore size)
were set in the 24-well plate. After being serum-starved for 12 h,
the BGC823 cells (1×105) were loaded into the upper
chamber. Conditioned medium containing 10% FBS was added to each of
the wells, and the cells were allowed to invade by penetrating
through the membrane during a 24-h incubation period at 37°C. The
cells on the lower surface of the Transwell filters were stained
with 5% crystal violet. The results were calculated from five
random fields taken by light microscope.
Microarrays and gene expression
analysis
Total RNA was extracted using TRIzol reagent,
according to the manufacturers instructions. Whole Human Genome
Oligo Microarray (Agilent Technologies, Inc., Santa Clara, CA, USA)
was performed by KangChen Bio-tech (Shanghai, China) to observe the
difference in transcriptional profiles between the pcDNA3.1-vector
and -REC8 cells. The microarray datasets revealed the differential
expression of genes and these were quantile normalized and analyzed
by the GeneSpring GX software package (Version 11.0; Agilent
Technologies). The Gene Ontology (GO) biological process and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analysis were performed. The Database for Annotation, Visualization
and Integrated Discovery (DAVID) 6.7 was used and the results were
ranked by P-values.
cDNA preparation and real-time
PCR
Total RNA was extracted using TRIzol reagent,
according to the manufacturers instructions. For quantitative
reverse-transcription polymerase chain reaction (qRT-PCR), total
RNA was reverse-transcribed to cDNA by using Bestar qPCR RT kit
(cat. no. DBI-2220; DBI Bioscience, Ludwigshafen, Germany). Next,
cDNA was quantified by real-time PCR on an Mx3000P real-time PCR
system (Agilent Technologies) with DBI Bestar® SYBR
Green qPCR Master Mix (cat. no. DBI-2043; DBI Bioscience). Results
were normalized to the expression of β-actin. The 2−∆∆Ct
method was used for calculating expression levels. The PCR primers
are shown in Table I.
 | Table I.Primers used in qRT-PCR. |
Table I.
Primers used in qRT-PCR.
| ID | Sequences (5–3) | Product length
(bp) |
|---|
| Β-actin Forward |
ATCGTGCGTGACATTAAGGAGAAG | 179 |
| Β-actin Reverse |
AGGAAGGAAGGCTGGAAGAGTG |
|
| REC8 Forward |
CATCCCACCAGAAGAACGG | 110 |
| REC8 Reverse |
GCACCAAAGGCATCTCCAT |
|
| EGR1 Forward |
TTCGACCTGCTCATCTTCGG | 229 |
| EGR1 Reverse |
CGATGCGTGAGTCCATGTGT |
|
| TGFβ1 Forward |
GCAACAATTCCTGGCGATAC | 134 |
| TGFβ1 Reverse |
AAGGCGAAAGCCCTCAAT |
|
| ATG12 Forward |
TAGAGCGAACACGAACCATCC | 153 |
| ATG12 Reverse |
CACTGCCAAAACACTCATAGAGA |
|
| SOX10 Forward |
CCTCACAGATCGCCTACACC | 161 |
| SOX10 Reverse |
CATATAGGAGAAGGCCGAGTAGA |
|
| E-cadherin
Forward |
CCCTGTTGGTGTCTTTATTATTG | 185 |
| E-cadherin
Reverse |
ATTCGGGCTTGTTGTCATTC |
|
| Vimentin
Forward |
GACGCCATCAACACCGAGTT | 238 |
| Vimentin
Reverse |
CTTTGTCGTTGGTTAGCTGGT |
|
Protein extraction and western blot
analysis
Cells were collected and lysed on ice in RIPA buffer
containing a protease inhibitor cocktail. Protein concentration was
assessed and 100 µg of protein from each sample was loaded and
separated by 10% polyacrylamide gel electrophoresis and transferred
onto PVDF membranes. After incubation with the specific primary
antibodies: REC8 antibody (1:1,000; rabbit monoclonal, cat. no.
ab192241; Abcam, San Francisco, CA, USA), EGR1 antibody (1:1,000;
rabbit polyclonal, cat. no. ab6054; Abcam), TGFβ1 antibody
(1:1,000; rabbit polyclonal, cat. no. ab92486; Abcam), SOX10
antibody (1:1,000; rabbit polyclonal, cat. no. ab108408; Abcam),
vimentin antibody (1:1,000; rabbit monoclonal, cat. no. ab92547;
Abcam), E-cadherin antibody (1:1,000; rabbit polyclonal, cat. no.
3195; Cell Signaling Technology, Danvers, MA, USA), GAPDH antibody
(1:4,000; rabbit polyclonal, cat. no. 2118; Cell Signaling
Technology), ATG12 antibody (1:1,000; mouse monoclonal, cat. no.
sc-271688; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C
overnight, the membranes were incubated with HRP-conjugated
secondary antibodies (1:10,000; cat no. A4416 and A6154;
Sigma-Aldrich). ECL luminescent reagent (cat. no. 32109;
Invitrogen/Thermo Fisher Scientific) was added and the signals were
recorded on X-ray film in a darkroom. GAPDH was used as the loading
control.
Immunofluorescence assay
Cells were fixed with 4% paraformaldehyde and
incubated with anti-β-catenin antibody (1:100; rabbit monoclonal
β-catenin antibody, cat. no. 8480; Cell Signaling Technology)
overnight at 4°C. After blocking and washing, the coverslips were
incubated with anti-rabbit IgG (H+L) F(ab)2 Fragment
(Alexa Fluro 488 conjugate) (1:500; cat. no. 4412; Cell Signaling
Technology). The nuclei were stained with DAPI
(4′,6-diamidino-2-phenylindole) for 5 min. After 3 washes, the
slides were immediately examined with a confocal microscope (Zeiss
710, 63× oil; Carl Zeis, Heidenheim, Germany) and the images were
captured.
Immunoprecipitation assays
Cell lysates were incubated with protein-A/G linked
agarose (1 mg of protein/40 µl of beads) for 1 h at 4°C to exclude
any non-specific binding. The supernatant collected after
centrifugation was incubated with 1 µg of the specific antibody
REC8 (1:1,000; rabbit monoclonal, cat. no. ab192241; Abcam)
overnight at 4°C. The solution was then incubated with protein-A/G
agarose beads for specific binding. IgG was used as the negative
control. Beads were washed with RIPA buffer after incubation and
dissolved in an SDS-PAGE loading buffer. The subsequent steps were
performed as in the western blot analysis and have been previously
described.
Statistical analysis
All data were obtained from at least three
independent experiments. The significance of the differences
between groups was determined using a two-tailed Students-t test or
analysis of variance (ANOVA) followed by Tukeys test. P<0.05 was
considered to indicate a statistically significant result.
Results
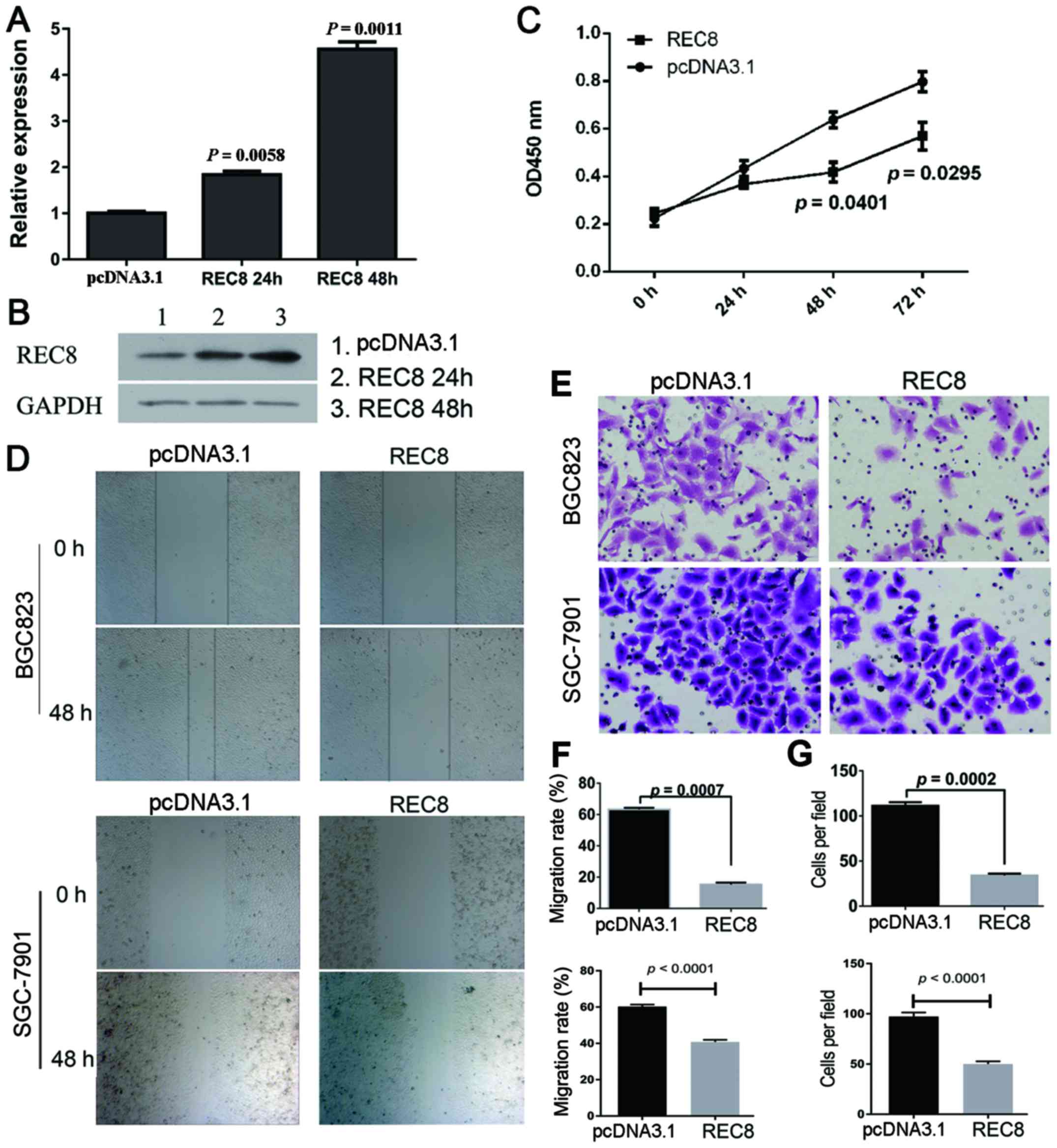
Overexpression of REC8 inhibits the
proliferation, migration and invasion of gastric cancer cells
To assess the effect of REC8 on gastric cancer cell
proliferation, migration and invasion, we overexpressed REC8 by
transient transfection in BGC823 cells. Cells overexpressing REC8
protein were successfully obtained, as observed by the RT-PCR
(Fig. 1A) and western blotting data
(Fig. 1B).
We assessed tumor cell proliferation in response to
REC8 overexpression, using the CCK-8 assay in BCG823 cell lines. As
shown in Fig. 1C, the BGC823 cells
overexpressing REC8 exhibited significantly reduced proliferation
compared with vector controls at 48 and 72 h (P=0.0401 and
P=0.0295). We further examined the effect of REC8 overexpression on
the motility of BGC823 cells using the scratch wound healing assay.
As shown in Fig. 1D and F, the
migration of BGC823 and SGC-7901 cells overexpressing REC8 was
significantly reduced (P<0.001). In addition, we examined the
invasion of BGC823 and SGC-7901 cells overexpressing REC8 using a
chamber invasion assay, and found that the invasion rates of these
cells were also decreased (P=0.0002; Fig. 1E and G).
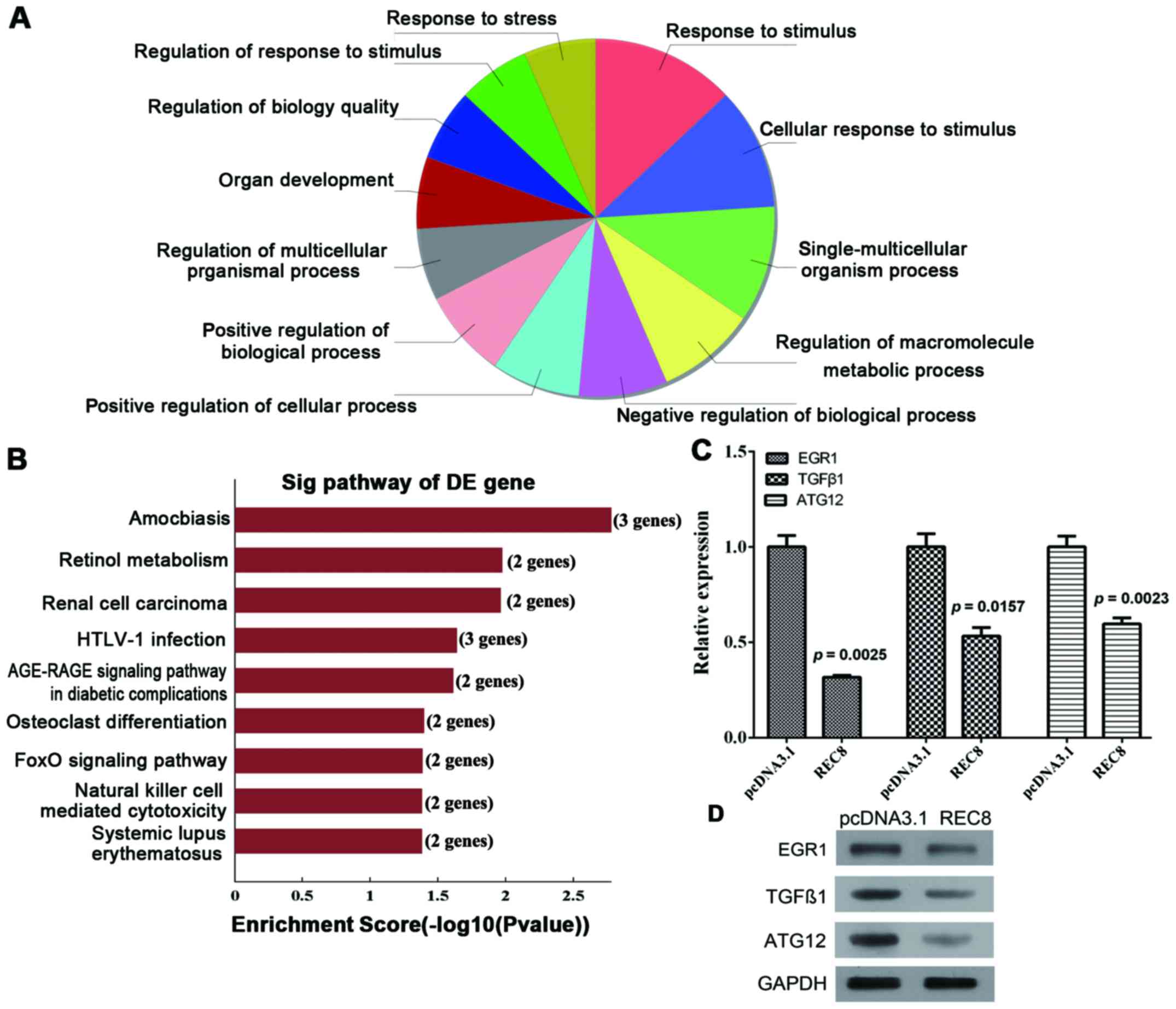
Overexpression of REC8 induces the
AGE-RAGE pathway
We used genomic microarray to analyze the
transcriptome profiles induced by REC8 expression in BGC823 cells.
A total of 1,657 genes were differentially expressed by at least
2-fold between BGC823-REC8 and -pcDNA3.1 cells. We analyzed the
biological processes of the downregulated genes using GO and the
KEGG pathway databases with bioinformatic methods (Fig. 2A and B). Among the 9 significantly
enriched KEGG terms and/or pathways (Table II), the AGE-RAGE signaling pathway
is associated with EMT, while the FoxO signaling pathway is
associated with apoptosis. The expression of EGR1 and TGFβ1 in the
AGE-RAGE pathway and the expression of ATG12 and TGFβ1 in the FoxO
pathway were significantly downregulated by overexpression of REC8.
The qRT-PCR and western blot assays further validated the results
of the microarray (Fig. 2C and D;
P=0.0025, P=0.0157 and P=0.0023, respectively). Since the
expression of EGR1 was downregulated significantly, we hypothesized
that the AGE-RAGE signaling pathway should be further investigated
in future research.
| Table II.KEGG pathway enrichment analysis of
the expression of downregulated genes between BGC823-REC8 and
-pcDNA3.1 cells. |
Table II.
KEGG pathway enrichment analysis of
the expression of downregulated genes between BGC823-REC8 and
-pcDNA3.1 cells.
| Pathway ID | Definition | P-value
(Fisher) | Genes |
|---|
| hsa05146 | Amoebiasis -
Homo sapiens (human) | 0.001670065 |
SERPINB3/SERPINB4/TGFB1 |
| hsa00830 | Retinol metabolism
- Homo sapiens (human) | 0.01058397 | CYP3A5/DHRS3 |
| hsa05211 | Renal cell
carcinoma - Homo sapiens (human) | 0.01089918 | ARNT2/TGFB1 |
| hsa05166 | HTLV-I infection -
Homo sapiens (human) | 0.0230317 |
EGR1/PPP3R1/TGFB1 |
| hsa04933 | AGE-RAGE signaling
pathway in diabetic complications Homo sapiens (human) | 0.02441325 | EGR1/TGFB1 |
| hsa04380 | Osteoclast
differentiation - Homo sapiens (human) | 0.03999841 | PPP3R1/TGFB1 |
| hsa04068 | FoxO signaling
pathway - Homo sapiens (human) | 0.04110769 | ATG12/TGFB1 |
| hsa04650 | Natural killer cell
mediated cytotoxicity - Homo sapiens (human) | 0.04166677 | PPP3R1/SH3BP2 |
| hsa05322 | Systemic Lupus
erythematosus - Homo sapiens (human) | 0.04166677 |
HIST1H2AD/HIST2H2AA4 |
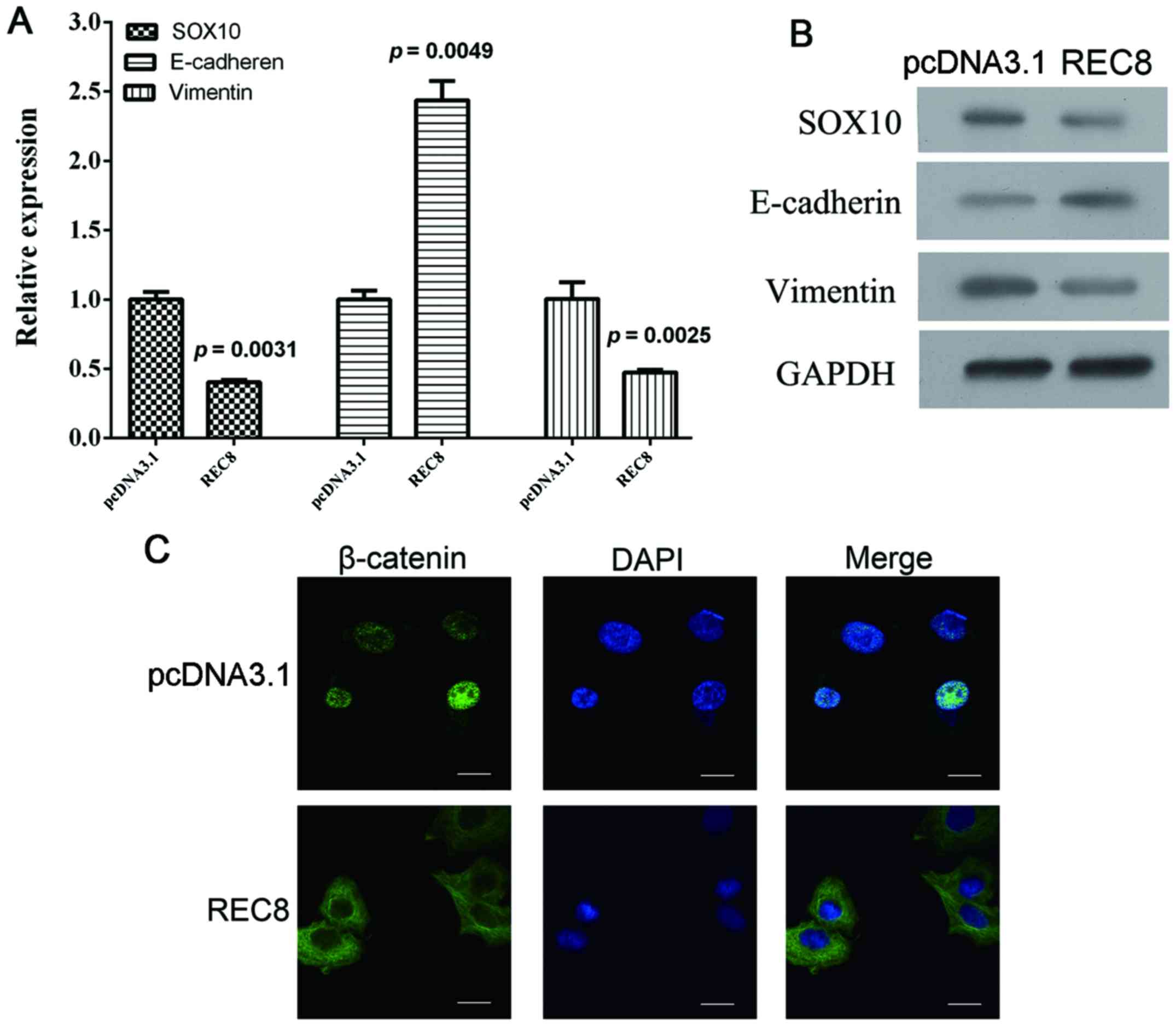
REC8 inhibits EMT in gastric cancer
cells through downregulation of EGR1 expression
When the REC8 overexpression plasmid was transfected
into BGC823 cells, we detected an associated increase of
E-cadherin, a decrease of SOX10 and vimentin at both the mRNA and
protein levels (Fig. 3A and B;
P=0.0049, P=0.0031 and P=0.0025, respectively). We also observed a
nuclear accumulation of β-catenin using immunofluorescence assays
(Fig. 3C). Collectively, these
results demonstrated that REC8 appeared to inhibit EMT in gastric
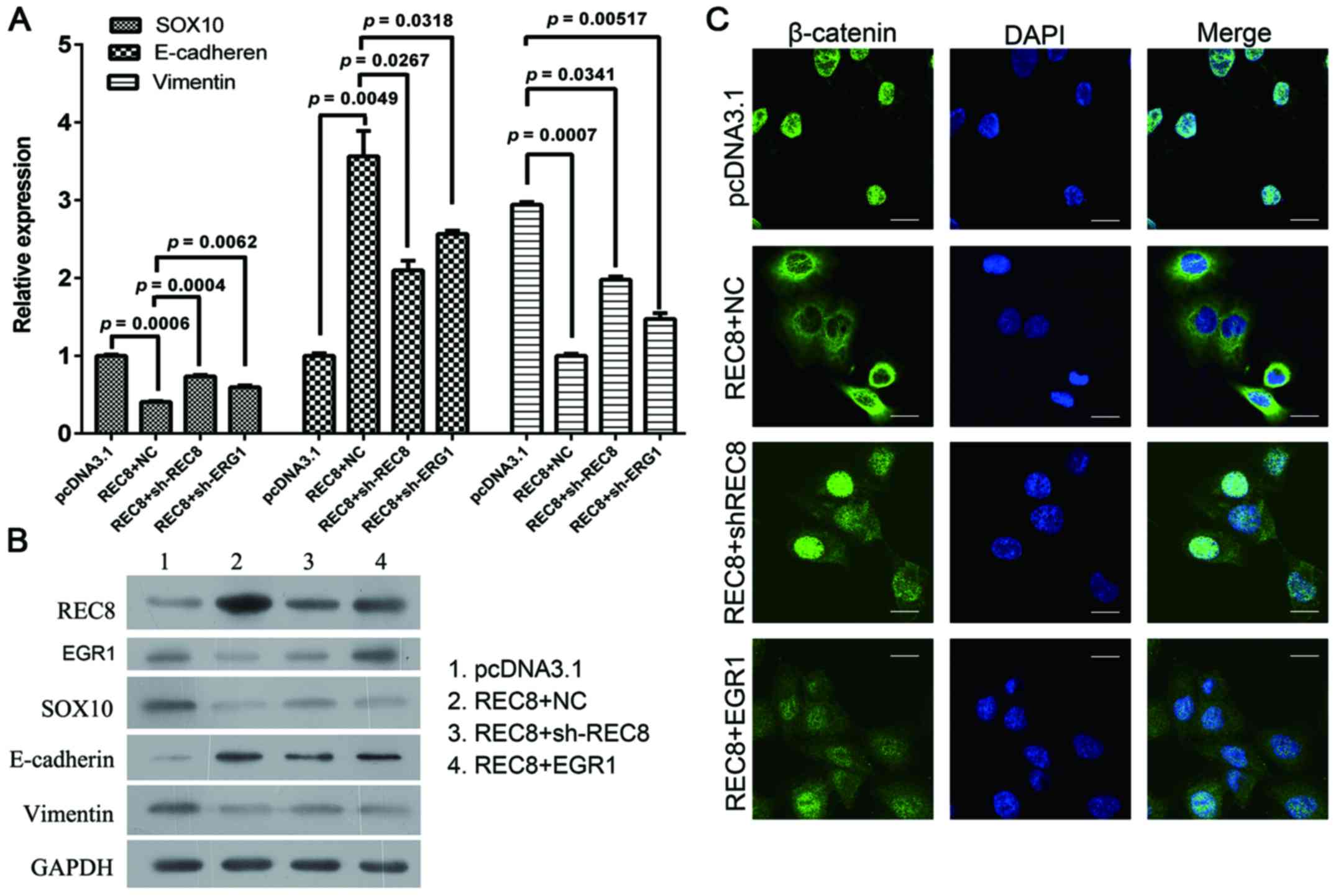
cancer cells. In our previous microarray analysis, we found that
REC8 regulated EGR1 expression. We, therefore transfected
REC8-overexpressed BGC823 cells with REC8 shRNA or EGR1
overexpression plasmids and assessed the expression levels of REC8
and EGR1, and the expression characteristics of EMT markers using
qRT-PCR, western blot and immunofluorescence assays. Consistent
with the previous results obtained, the artificial overexpression
of REC8 decreased the expression of EGR1. When we ablated REC8 in
REC8-transfected cells using REC8 shRNA or overexpressed EGR1 by
using the pcDNA3.1-EGR1 plasmid, we found that the expression of
REC8 or overexpressed EGR1 reduced the anti-EMT effect observed in
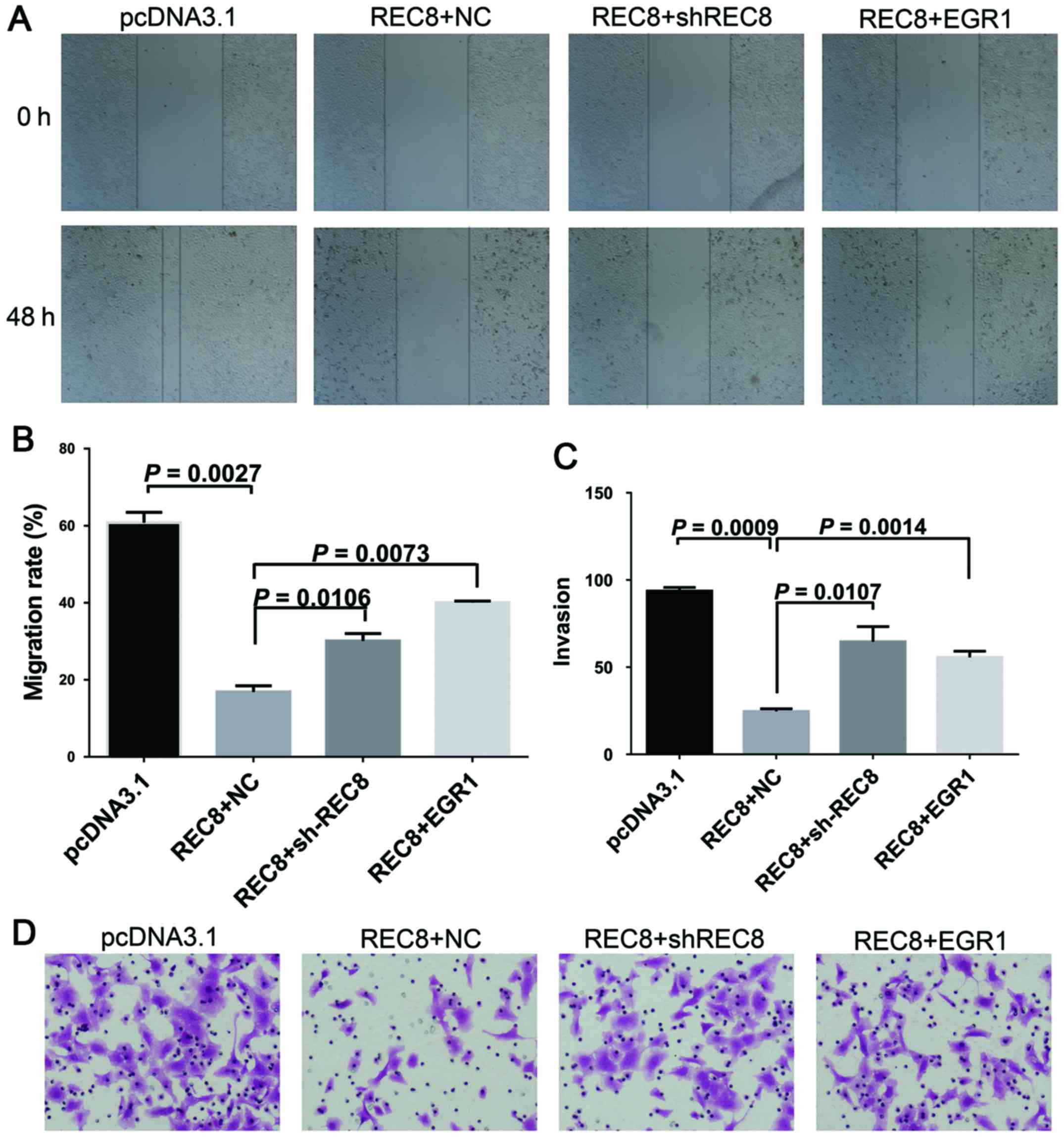
REC8 overexpressed gastric cancer cells (Fig. 4). In addition, we examined whether
REC8 regulated cell motility and invasion through EGR1 expression
using the wound healing and the chamber invasion assays. When we
ablated REC8 expression or overexpressed EGR1, we found that the
anti-motility effect of REC8 in REC8 overexpressed gastric cancer
cells was reduced, like we predicted (Fig. 5). These results revealed that REC8
contributed to the anti-EMT effect through EGR1 expression.
REC8 and EGR1 physically interact and
therefore REC8 regulates EGR1 expression
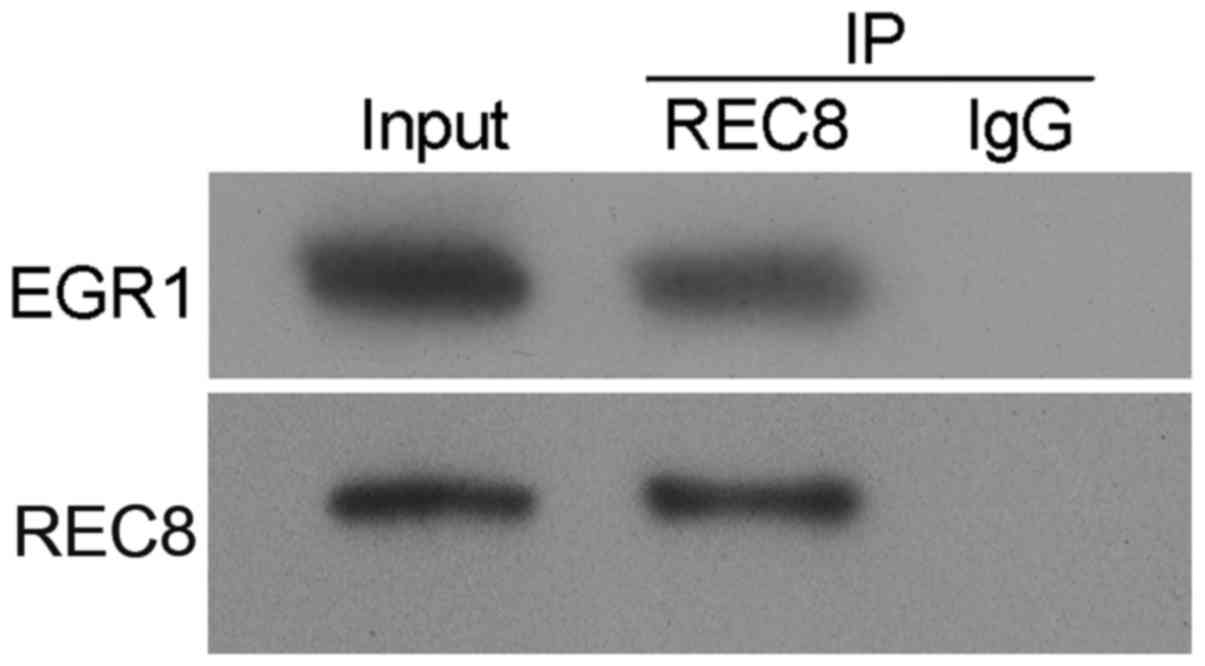
To further investigate the manner in which REC8
decreased EGR1 expression in gastric cancer cells, we performed an
immunoprecipitation assay, and observed a strong interaction
between REC8 and EGR1 (Fig. 6).
This result revealed that REC8 decreased EGR1 expression by
interacting with it.
Discussion
REC8 is an important component of the meiotic
prophase chromosome axis that mediates sister chromatid cohesion,
homologous chromosome pairing, crossover recombination and
chromosome synapsis (13,14). In the present study, we hypothesized
that REC8 acts as a tumor-suppressor gene and we demonstrated for
the first time that overexpressed REC8 inhibited gastric cancer
cell proliferation and migration.
However, the regulatory mechanisms of REC8 in
gastric cancer are not known. To understand how the carcinogenic
process is influenced by REC8 in gastric cancer, we searched for
differential expression profiles and signaling pathways induced by
REC8 overexpression using microarrays. We ascertained that EGR1,
TGFβ1 and ATG12 were downregulated by REC8 overexpression and were
involved in the AGE-RAGE and FoxO signaling pathways. The FoxO
pathway plays an important role in cell growth and cell survival
(15), while the AGE-RAGE pathway
is involved in EMT (16–18).
Gastric cancer is the second leading cause of
cancer-related deaths in the world (19). Even though patients undergo curative
therapy, they still succumb to recurrent disease due to metastasis.
Therefore, to treat gastric cancer, it is important to understand
the molecular mechanisms underlying metastasis, thus finding the
relevant therapeutic targets. EMT is a process where epithelial
cells lose their characteristics including their spindle-cell
shape, polarity, intercellular separation and pseudopodia formation
(20–22). EMT is critical in many cancer
biological behaviors, such as migration and invasion. The
multifaceted REC8-mediated anticancer effects play a causal but
unclear role in mammalian oncogenesis, thus in the present study,
we focused on EMT and the AGE-RAGE pathway. Furthermore, EGR1, a
protein in the AGE-RAGE pathway is an important factor in promoting
EMT (11,23). To explore the manner in which EGR1
expression promotes EMT in gastric cancer, we searched for EGR1
binding proteins using a protein interaction prediction program. We
found that EGR1 interacted with SOX10, a marker of EMT. In support
of our hypothesis, we observed marked changes in morphology,
biochemistry, histochemistry and motility in BGC823-REC8 cells.
Specifically, we observed an inhibition of EMT and a less motile
and invasive phenotype than observed in vector cells. In previous
studies, REC8 was considered to be a tumor suppressor via other
factors and signaling pathways involved in the regulation of cell
proliferation, apoptosis, the cell cycle and migration (1,24–33).
Furthermore, based on data from epigenetic inactivation
experiments, the antitumor effect of REC8 was attributed to
methylation (34). As we previously
described, EGR1 was demonstrated to be downregulated by REC8.
Overexpression of EGR1 in REC8-transfected cells induced motility
and EMT in gastric cancer cells which was the reverse of what was
observed in REC8 overexpressed cells. These results revealed that
EGR1 is a novel downstream protein of REC8 that regulates EMT in
gastric cancer cells.
To understand the manner in which EGR1 expression
was downregulated by REC8 in gastric cancer, we demonstrated that
REC8 interacted with EGR1 and therefore downregulated EGR1. These
results revealed that EGR1 promotes EMT depending on the amount of
uncombined EGR1. The uncombined EGR1 can either bind with SOX10
directly or act as a transcription factor, and activate oncogenes
to regulate EMT. The mechanism of REC8 in EMT in gastric cancer
cells requires further study. In the present study we revealed that
the EGR1 protein contributed to EMT regulation. In future research,
the underlying mechanism of EGR1 in the regulation of EMT needs to
be illuminated more clearly.
In conclusion, our results revealed that REC8
decreased EMT in gastric cancer cells by regulating the expression
of EGR1 and interacting with it. REC8 or EGR1 are potential targets
in the treatment of gastric cancer.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 81301758), the Natural
Science Foundation of Guangdong (no. 2016A030310254) and the
Postdoctoral Science Foundation of China (no. 2016M600648).
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
REC8
|
meiotic recombination protein REC8
homolog
|
|
EGR1
|
early growth response-1
|
|
EMT
|
epithelial-mesenchymal transition
|
References
|
1
|
Arseneault R, Chien A, Newington JT,
Rappon T, Harris R and Cumming RC: Attenuation of LDHA expression
in cancer cells leads to redox-dependent alterations in
cytoskeletal structure and cell migration. Cancer Lett.
338:255–266. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang D, Duan H, Huang H, Tong X, Han Y,
Ru G, Qu L, Shou C and Zhao Z: Cisplatin resistance in gastric
cancer cells is associated with HER2 upregulation-induced
epithelial-mesenchymal transition. Sci Rep. 6:205022016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang J, Xiao D, Li G, Ma J, Chen P, Yuan
W, Hou F, Ge J, Zhong M, Tang Y, et al: EphA2 promotes
epithelial-mesenchymal transition through the Wnt/β-catenin pathway
in gastric cancer cells. Oncogene. 33:2737–2747. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dravis C, Spike BT, Harrell JC, Johns C,
Trejo CL, Southard-Smith EM, Perou CM and Wahl GM: Sox10 regulates
stem/progenitor and mesenchymal cell states in mammary epithelial
cells. Cell Reports. 12:2035–2048. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nasmyth K: Disseminating the genome:
Joining, resolving, and separating sister chromatids during mitosis
and meiosis. Annu Rev Genet. 35:673–745. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Laird PW: The power and the promise of DNA
methylation markers. Nat Rev Cancer. 3:253–266. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Furuta J, Nobeyama Y, Umebayashi Y, Otsuka
F, Kikuchi K and Ushijima T: Silencing of Peroxiredoxin 2 and
aberrant methylation of 33 CpG islands in putative promoter regions
in human malignant melanomas. Cancer Res. 66:6080–6086. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okamoto Y, Sawaki A, Ito S, Nishida T,
Takahashi T, Toyota M, Suzuki H, Shinomura Y, Takeuchi I, Shinjo K,
et al: Aberrant DNA methylation associated with aggressiveness of
gastrointestinal stromal tumour. Gut. 61:392–401. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu D, Shen X, Zhu G and Xing M: REC8 is a
novel tumor suppressor gene epigenetically robustly targeted by the
PI3K pathway in thyroid cancer. Oncotarget. 6:39211–39224. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu Y, Toure F, Qu W, Lin L, Song F, Shen
X, Rosario R, Garcia J, Schmidt AM and Yan SF: Advanced glycation
end product (AGE)-receptor for AGE (RAGE) signaling and
up-regulation of Egr-1 in hypoxic macrophages. J Biol Chem.
285:23233–23240. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun S, Ning X, Zhai Y, Du R, Lu Y, He L,
Li R, Wu W, Sun W and Wang H: Egr-1 mediates chronic
hypoxia-induced renal interstitial fibrosis via the PKC/ERK
pathway. Am J Nephrol. 39:436–448. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Chen X, Wang J, Guang W, Han W,
Zhang H, Tan X and Gu Y: EGR1 decreases the malignancy of human
non-small cell lung carcinoma by regulating KRT18 expression. Sci
Rep. 4:54162014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yoon SW, Lee MS, Xaver M, Zhang L, Hong
SG, Kong YJ, Cho HR, Kleckner N and Kim KP: Meiotic prophase roles
of Rec8 in crossover recombination and chromosome structure.
Nucleic Acids Res. 44:9296–9314. 2016.PubMed/NCBI
|
|
14
|
Watanabe Y and Nurse P: Cohesin Rec8 is
required for reductional chromosome segregation at meiosis. Nature.
400:461–464. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cohen S, Lee D, Zhai B, Gygi SP and
Goldberg AL: Trim32 reduces PI3K-Akt-FoxO signaling in muscle
atrophy by promoting plakoglobin-PI3K dissociation. J Cell Biol.
204:747–758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Raghavan CT and Nagaraj RH: AGE-RAGE
interaction in the TGFβ2-mediated epithelial to mesenchymal
transition of human lens epithelial cells. Glycoconj J. 33:631–643.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song JS, Kang CM, Park CK, Yoon HK, Lee
SY, Ahn JH and Moon HS: Inhibitory effect of receptor for advanced
glycation end products (RAGE) on the TGF-β-induced alveolar
epithelial to mesenchymal transition. Exp Mol Med. 43:517–524.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li JH, Wang W, Huang XR, Oldfield M,
Schmidt AM, Cooper ME and Lan HY: Advanced glycation end products
induce tubular epithelial-myofibroblast transition through the
RAGE-ERK1/2 MAP kinase signaling pathway. Am J Pathol.
164:1389–1397. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hay ED: An overview of
epithelio-mesenchymal transformation. Acta Anat (Basel). 154:8–20.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ozen E, Gozukizil A, Erdal E, Uren A,
Bottaro DP and Atabey N: Heparin inhibits hepatocyte growth factor
induced motility and invasion of hepatocellular carcinoma cells
through early growth response protein 1. PLoS One. 7:e427172012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Z, Zhang B, Li W, Fu L, Fu L, Zhu Z
and Dong JT: Epigenetic silencing of miR-203 upregulates SNAI2 and
contributes to the invasiveness of malignant breast cancer cells.
Genes cancer. 2:782–791. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ying J, Srivastava G, Hsieh WS, Gao Z,
Murray P, Liao SK, Ambinder R and Tao Q: The stress-responsive gene
GADD45G is a functional tumor suppressor, with its response to
environmental stresses frequently disrupted epigenetically in
multiple tumors. Clin Cancer Res. 11:6442–6449. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Regel I, Eichenmüller M, Joppien S, Liebl
J, Häberle B, Müller-Höcker J, Vollmar A, von Schweinitz D and
Kappler R: IGFBP3 impedes aggressive growth of pediatric liver
cancer and is epigenetically silenced in vascular invasive and
metastatic tumors. Mol Cancer. 11:92012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pruitt SC, Bailey KJ and Freeland A:
Reduced Mcm2 expression results in severe stem/progenitor cell
deficiency and cancer. Stem Cells. 25:3121–3132. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Medina-Ramirez CM, Goswami S, Smirnova T,
Bamira D, Benson B, Ferrick N, Segall J, Pollard JW and Kitsis RN:
Apoptosis inhibitor ARC promotes breast tumorigenesis, metastasis,
and chemoresistance. Cancer Res. 71:7705–7715. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu JY, Qian D, He LR, Li YH, Liao YJ, Mai
SJ, Tian XP, Liu YH, Zhang JX, Kung HF, et al: PinX1 suppresses
bladder urothelial carcinoma cell proliferation via the inhibition
of telomerase activity and p16/cyclin D1 pathway. Mol Cancer.
12:1482013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim MO, Lee YJ, Park JH, Ryu JM, Yun SP
and Han HJ: PKA and cAMP stimulate proliferation of mouse embryonic
stem cells by elevating GLUT1 expression mediated by the NF-κB and
CREB/CBP signaling pathways. Biochim Biophys Acta. 1820:1636–1646.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kabbout M, Garcia MM, Fujimoto J, Liu DD,
Woods D, Chow CW, Mendoza G, Momin AA, James BP, Solis L, et al:
ETS2 mediated tumor suppressive function and MET oncogene
inhibition in human non-small cell lung cancer. Clin Cancer Res.
19:1–3395. 2013. View Article : Google Scholar
|
|
32
|
Du W, Jiang P, Mancuso A, Stonestrom A,
Brewer MD, Minn AJ, Mak TW, Wu M and Yang X: TAp73 enhances the
pentose phosphate pathway and supports cell proliferation. Nat Cell
Biol. 15:991–1000. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Deep G, Jain AK, Ramteke A, Ting H,
Vijendra KC, Gangar SC, Agarwal C and Agarwal R: SNAI1 is critical
for the aggressiveness of prostate cancer cells with low
E-cadherin. Mol Cancer. 13:372014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu J, Liang Q, Wang J, Wang K, Gao J,
Zhang J, Zeng Y, Chiu PW, Ng EK and Sung JJ: REC8 functions as a
tumor suppressor and is epigenetically downregulated in gastric
cancer, especially in EBV-positive subtype. Oncogene. 36:182–193.
2017. View Article : Google Scholar : PubMed/NCBI
|