Introduction
Colorectal cancer (CRC), which ranks third in terms
of cancer-related morbidity and second in terms of cancer-related
mortality worldwide, is considered a major threat to public health
(1,2). Colorectal carcinogenesis is a
multi-step biological process involving dysregulation of multiple
oncogenes and tumor suppressor genes (3). Despite recent progress in diagnostic
techniques and highly effective medical treatments, the overall
survival rate of patients with CRC remains relatively low (4). At present, in light of the growing
incidence rate and poor prognosis of CRC, there is an urgent demand
to understand the specific molecular mechanisms underlying the
pathogenic process of CRC, as this may facilitate the development
of effective therapeutic strategies to ultimately improve patient
prognosis (5,6).
The high mortality rate associated with CRC is
generally attributed to tumor invasion and metastasis. There are
many factors and systems that are related to tumor invasion and
metastasis, including the epithelial-mesenchymal transition (EMT),
which promotes cancer cell invasion and migration. Specifically,
EMT promotes epithelial cells in a certain region to detach from
epithelial tissue and migrate to other locations, which is the
basis of tumor metastasis in tumorigenesis. Therefore, inhibition
of EMT may effectively control the migration of tumor cells. Recent
studies have revealed that EMT may be regulated by non-coding RNAs
(ncRNAs) as well as coding RNAs; however, data on the function of
ncRNA in EMT remains limited (7,8).
To date, improvements in whole-genome and
transcriptome sequencing technologies have led to the discovery
that the majority of the full mammalian genome can be transcribed;
however, most of the transcripts have limited or no protein-coding
capacity, and are known as ncRNAs (9,10).
Distinct from small ncRNAs (such as siRNAs, miRNAs and piRNAs) that
have been widely studied, long non-coding RNAs (lncRNAs) are
obscure molecules that make up ~80% of ncRNAs and have unclear
functions at present (9). lncRNAs,
which are transcripts of >200 nucleotides in length, are
transcribed by RNA polymerase II (RNAPII); however, they have few
or no open reading frames (ORFs). Extensive evidence suggests that
lncRNAs serve crucial roles in a wide variety of fundamental
biological processes, including embryonic development, epigenetic
silencing, transcriptional and translational control, cell growth,
cell differentiation, cell migration and tumorigenesis (11,12).
Additionally, extensive research has provided strong evidence to
suggest that lncRNAs serve a functional role in a range of human
diseases, including various types of cancer, such as breast
(13), pancreatic (14), lung (15), gastric (10) and cervical (16). Furthermore, it has been ascertained
that numerous lncRNAs participate in both oncogenic and
tumor-suppressing pathways in tumorigenesis (10,17,18).
For instance, lncRNA MVIH may serve as an oncogene, serving to
promote cell proliferation and invasion and indicating poor
prognosis of non-small cell lung cancer (11). Similarly, lncRNA GPR158-AS1 has been
associated with high tumor risk, while lncRNA KCNK15-AS1 has been
shown to be protective against non-small cell lung cancer (19). Recently, evidence has suggested that
various lncRNAs also act as modulators in the carcinogenesis and
progression of human CRC, thus indicating their potential as novel
therapeutic targets. Regarding CRC, research on the molecular and
biological functions of lncRNAs in CRC is still in its infancy, and
has so far had limited success in elucidating the specific
molecular mechanisms. However, previous studies indicated that
lncRNAs may be potential biomarkers for the diagnosis and treatment
of CRC (11,20,21).
In the present study, we first determined that the
expression of XLOC_010588 was higher in cancer tissues compared
with that in adjacent normal tissues in CRC patients, and that
XLOC_010588 was closely associated with metastasis and poor
prognosis. Further functional experiments revealed that XLOC_010588
promoted cell invasion and migration. In addition, we demonstrated
that downregulation of XLOC_010588 may regulate the progression of
CRC invasion and migration via the EMT pathway. In summary, our
findings demonstrated that XLOC_010588 may serve as an oncogene in
CRC invasion and metastasis.
Materials and methods
Ethics statement
In the present study, all tissue microarrays were
purchased from Shanghai Outdo Biotech Co., Ltd. (Shanghai, China),
and were approved by the local Ethics Committee (Zhejiang Taizhou
Hospital Ethics Committee, Zhejiang Taizhou Hospital, Zhejiang,
China). Due to the retrospective nature of the study, the Ethics
Committee waived the requirement for written informed consent from
the patients. All the samples were anonymized.
Tissue preparation
A total of 111 samples of cancer tissue and 70
samples of normal adjacent tissues isolated from CRC patients were
purchased from Shanghai Outdo Biotech Co., Ltd. The tissues
supplied by Shanghai Outdo Biotech Co., Ltd., were collected during
surgery from November 2009 to May 2010, and the longest follow-up
time of the tissue samples was 68 months, and the shortest
follow-up time was only 1 month. Comprehensive clinicopathological
data on the tissue samples, including patient age and sex, tumor
size, differentiation and T stage, and presence of lymph node
metastasis and distant metastasis, were also collected.
In situ hybridization (ISH)
XLOC_010588 expression was detected by ISH in the
cancer tissues and normal adjacent tissues of CRC patients.
Following dewaxing of the microarray sections, we applied an
Enhanced Sensitive ISH Detection kit II (AP) (Boster Biological
Technology, Ltd., Wuhan, China). Following completion of the
prehybridization, digoxigenin-labeled oligonucleotide probes
(DIG-5′ ATTCTAACA TAATATCCCTGCAGT 3′-DIG) were hybridized at 40°C
overnight, and then the kit was used according to the
manufacturer's instructions. Based on both the intensity and
proportion of XLOC_010588-positive cells, the staining scores were
determined using a relatively simple, reproducible scoring method.
On a scale of 0–3, the staining intensity was scored as follows: 0,
none; 1, weak; 2, medium; and 3, strong. The extent of staining was
scored on a scale of 0–100%. The product of the staining intensity
and staining extent scores was used as the final score for
XLOC_010588 staining, with the final staining scores ranging from 0
to 300 (22). The average score of
each sample was used to assess cutoff scores for XLOC_010588
overexpression using receiver operating characteristic (ROC) curves
(23).
Cell lines
The human CRC cell lines Caco-2, HT-29, SW480, SW620
and HCT116, as well as normal human intestinal epithelial cells
(HIEC-6), were obtained from the Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China).
The HIECs were cultured in Dulbecco's modified Eagle's medium
(DMEM; HyClone Laboratories; GE Healthcare Life Sciences, Logan,
UT, USA); the Caco-2, HT29 and HCT116 cell lines were cultured in
RPMI-1640 medium (HyClone Laboratories; GE Healthcare Life
Sciences); and the SW480 and SW620 cell lines were cultured in
Leibovitz's L-15 medium (Gibco Cell Culture; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). All media were supplemented
with 10% fetal bovine serum (FBS) and 100 U/ml
penicillin/streptomycin (both from HyClone Laboratories; GE
Healthcare Life Sciences). All cell lines were maintained in a
humidified chamber containing 5% CO2 at 37°C.
RNA extraction and RT-qPCR
analysis
Total RNA was extracted from human CRC cell lines
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions, and then reverse
transcribed into cDNA with a PrimeScript RT Reagent kit with gDNA
Eraser (Perfect Real-Time) (Takara Bio, Inc., Otsu, Japan). The
primer sequences were as follows: XLOC_010588 forward,
5′-TGTGAAGAGGAGAACATAAAAGG-3′ and reverse,
5′-AAGCAAGATAATACAGTGGCGA-3′; and GAPDH forward,
5′-CTCCTCCTGTTCGACAGTCAGC-3′ and reverse,
5′-CCCAATACGACCAAATCCGTT-3′. The target cDNA was amplified in a
10-µl reaction mixture containing SYBR Premix Ex Taq II (Takara
Bio, Inc.), which was completed on an ABI 7500 Real-Time PCR system
(Thermo Fisher Scientific, Inc.). The amplification profile was
95°C for 5 min, followed by 42 cycles of denaturation at 95°C for
15 sec, and annealing and extension at 60°C for 60 sec. All
experiments were performed in triplicate. Relative gene expression
was determined using the comparative delta-delta Cq method (2-ΔΔCq)
(17). GAPDH was used as an
internal control for normalization.
Transfection
Based on the expression of XLOC_010588 in the CRC
cell lines, SW620 cells were selected for a knockdown study and
HCT116 cells for an overexpression experiment.
To achieve knockdown of XLOC_010588, after being
seeded into 6-well plates and incubated overnight, the SW620 cells
were transiently transfected with small interfering RNA (siRNA)
against XLOC_010588 (si-XLOC_010588) or a nonspecific control siRNA
(Guangzhou RiboBio Co., Ltd., Guangzhou, China) using Lipofectamine
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions. The sequences of the three
designed XLOC_010588 siRNAs were as follows: si-XLOC_010588 #1,
GCAGGGATAGAGCTTGCTT; si-XLOC_010588 #2, GCTGCTGACAGCAATTAAT; and
si-XLOC_010588 #3, GGTCCCTGCTTTGGTTTAA. At 48 h after transfection,
the cells were harvested to detect the knockout efficiency via
RT-qPCR.
For overexpression, the HCT116 cells were
transiently transfected with plasmids (pcDNA3.1 or
pcDNA3.1-XLOC_010588) (Hanbio Biotechnology Co., Ltd., Shanghai,
China), with the subsequent steps being the same as those for the
SW620 cell line.
After confirming that the cell models were
successful, the cells were collected for the following
experiments.
Wound healing assay
The SW620 and HCT116 cells were seeded into 6-well
plates, and subjected to cell transfection. After the cells had
been cultured to ~90% confluence, the cell layer was scratched with
a sterile 10-µl pipette tip and washed three times with PBS to
remove the detached cells. Subsequently, the scratch width change
was observed at 0, 6, 24 and 48 h by light microscopy. Cell
migration was measured using ImageJ 6.0 with the following formula:
Migration area ratio = the proportion of closed wound area/the
whole field of view area.
Migration and invasion assay
Cell migration and invasion assays were performed
using 24-well Transwell chambers (Corning Inc., Corning, NY, USA).
For the migration assay, a total of 8×104 cells in
serum-free media were seeded in the upper chambers after
transfection for 24 h, while medium containing 20% FBS was added to
the lower chambers. The invasion assay was set up in the same way,
except that the upper chambers were coated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). After 48 h of incubation,
the cells on the filter surface were fixed with 4% paraformaldehyde
and stained with crystal violet. Under a light microscope, five
high-power fields of view were randomly selected in which to count
the cells.
Immunofluorescence (IF)
Cells were seeded on 24×24-mm glass coverslips in
6-well plates. After transfection for 24 h, the cells were washed
three times with PBS and fixed with 4% paraformaldehyde at room
temperature for 30 min. After a PBS wash, the cells were
permeabilized with 0.1% Triton X-100. After further PBS washes, the
cells were blocked with 5% bovine serum albumin (BSA) for 30 min at
room temperature, then incubated with primary antibodies (1:200;
E-cadherin cat. no. 3195S; vimentin cat. no. 5741S, Slug cat. no.
9585P; all from Cell Signaling Technology, Inc., Beverly, MA, USA),
(1:200; E-cadherin cat. no. 610181; Slug cat. no. 564614; both from
BD Biosciences), followed by fluorescent secondary antibodies for
30 min at room temperature (1:100; FITC-goat anti-mouse IgG cat.
no. E031210-01; TRITC-goat anti-rabbit IgG cat. no. E031320-01;
ZSGB-BIO, Beijing, China). The cells were subsequently washed with
PBS, after which DAPI was used to stain the cell nuclei. The cells
were observed using an inverted fluorescence microscope (Nikon
Corp., Tokyo, Japan).
Western blotting
Total protein was extracted from the transfected
cells using radioimmunoprecipitation assay (RIPA) buffer
supplemented with the protease inhibitor phenylmethanesulfonyl
fluoride (PMSF), and the concentration of total protein was
measured using a BCA Protein assay kit (Beyotime Institute of
Biotechnology, Haimen, China). Subsequently, the extracted protein
samples were separated by 10% SDS-PAGE and transferred onto
polyvinylidene difluoride (PVDF) membranes. Following blocking of
the PVDF membranes with 5% BSA at room temperature for 1 h, the
membranes were incubated with various antibodies (1:1,000;
E-cadherin cat. no. 3195S; vimentin cat. no. 5741S; Slug cat. no.
9585P; all from Cell Signaling Technology, Inc.), (1:1,000;
E-cadherin cat. no. 610181; Slug cat. no. 564614; both from BD
Biosciences) at 4°C overnight, washed three times for 5 min each
with 1X Tris-buffered saline with Tween-20 (TBST), and then
incubated with secondary antibodies (1:2,500; ZB-2301 cat. no.
109525; ZB-2305 cat. no. 122627; ZSGB-BIO) at room temperature for
2 h. Finally, the proteins were visualized by
electrochemiluminescence (22).
Statistical analysis
All data are presented as the mean ± standard
deviation from three independent experiments. Student's t-tests or
ANOVA analysis were used to estimate the statistical significance
of differences between two groups or multiple groups regarding
clinical data and the results of cell experiments. Survival curves
were analyzed by the Kaplan-Meier method with the log-rank test.
P<0.05 was considered to indicate a statistically significant
difference. Statistical analyses were performed using SPSS software
(version 18.0; SPSS, Inc., Chicago, IL, USA) and presented using
GraphPad Prism software (version 6; GraphPad Software, Inc., La
Jolla, CA, USA).
Results
XLOC_010588 is upregulated in human
CRC tissues and associated with aggressive phenotypes
To explore the association between the expression of
XLOC_010588 in CRC and the clinicopathological data of the
patients, we performed an in situ hybridization analysis on 111
cancer tissue samples and 70 normal adjacent tissues from CRC
patients (Table I). The results
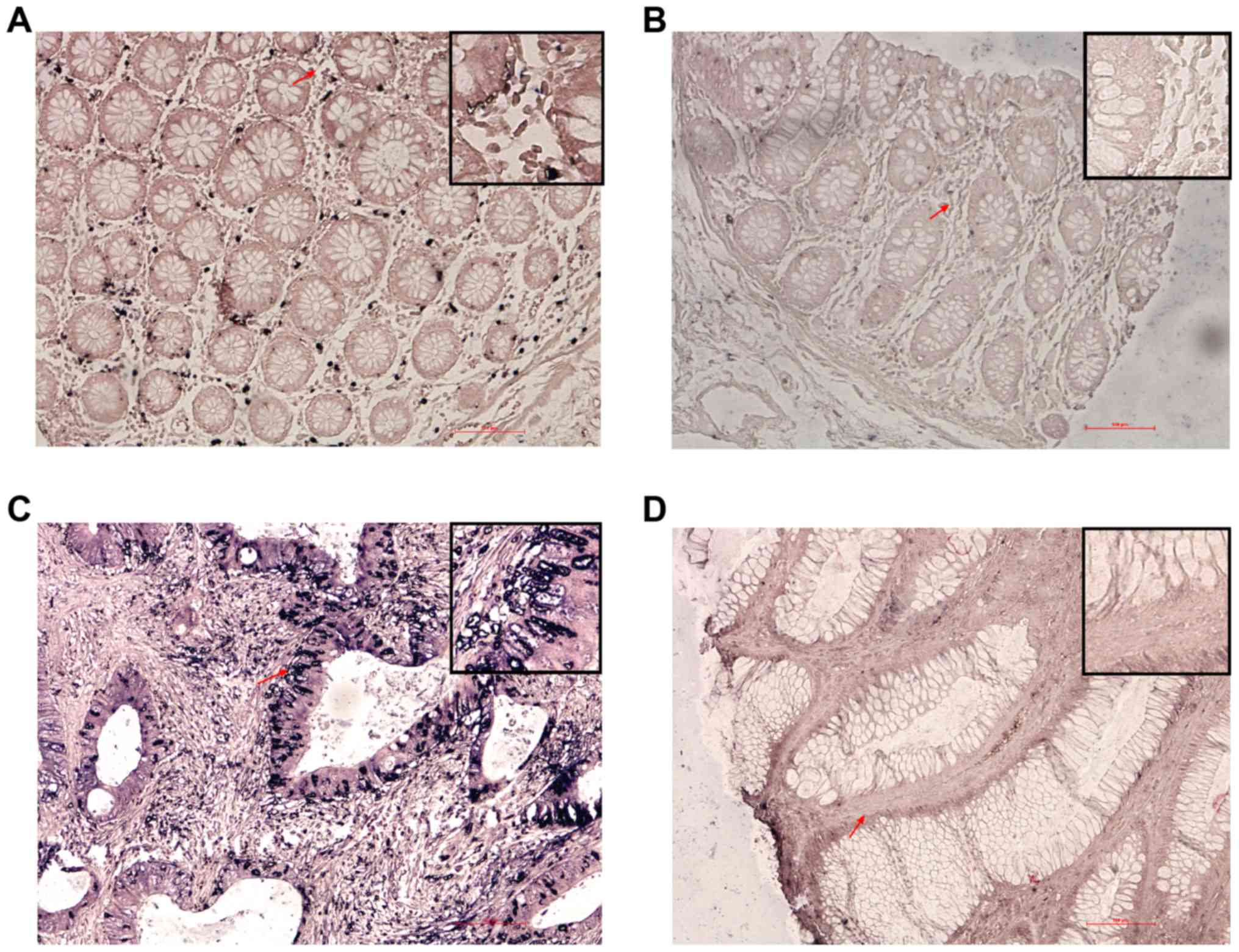
demonstrated that XLOC_010588 was specifically stained in both
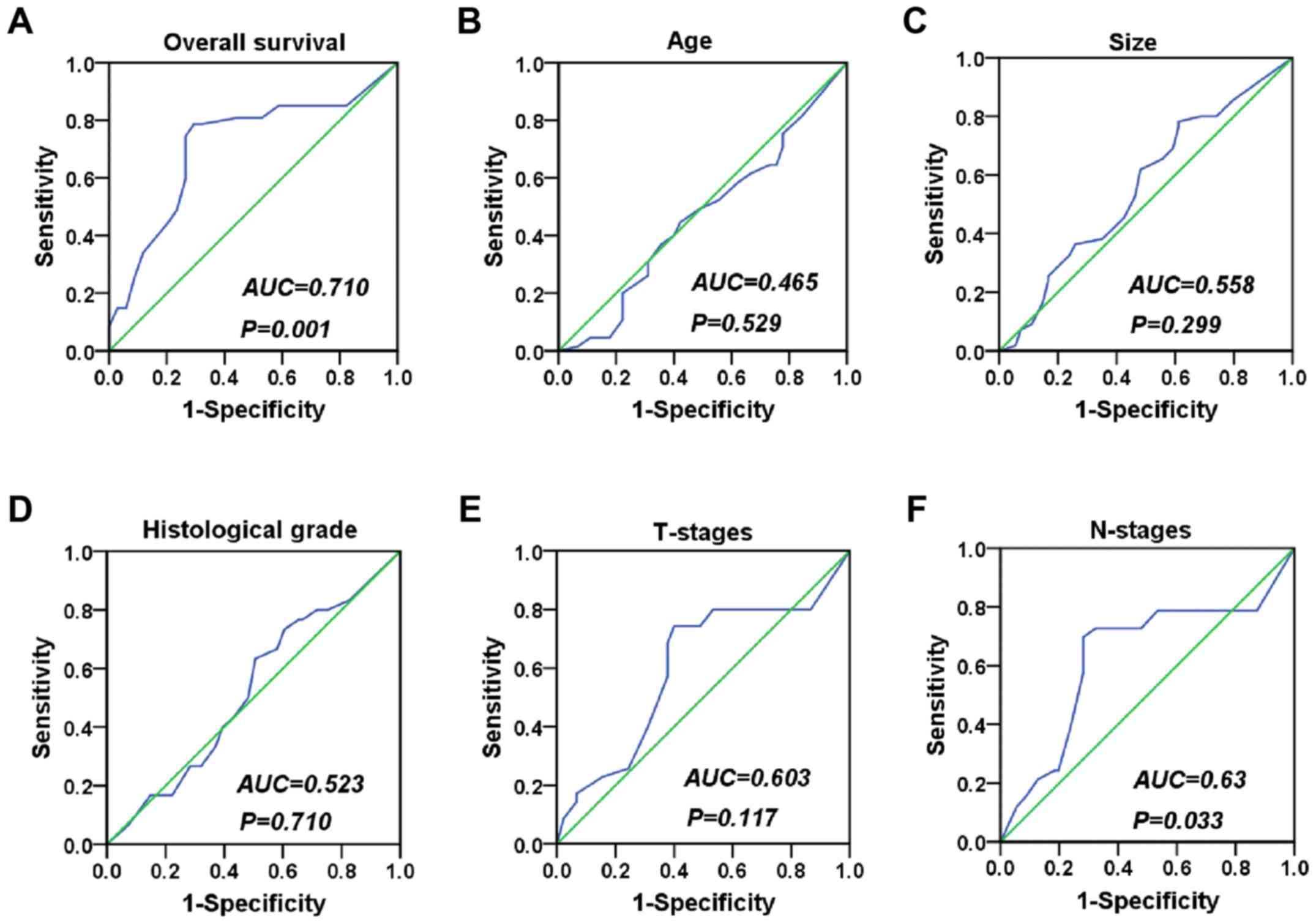
malignant epithelial cells and benign epithelial cells (Fig. 1). We used the ROC curve to obtain
cutoff value (cutoff value = 155) (Fig.
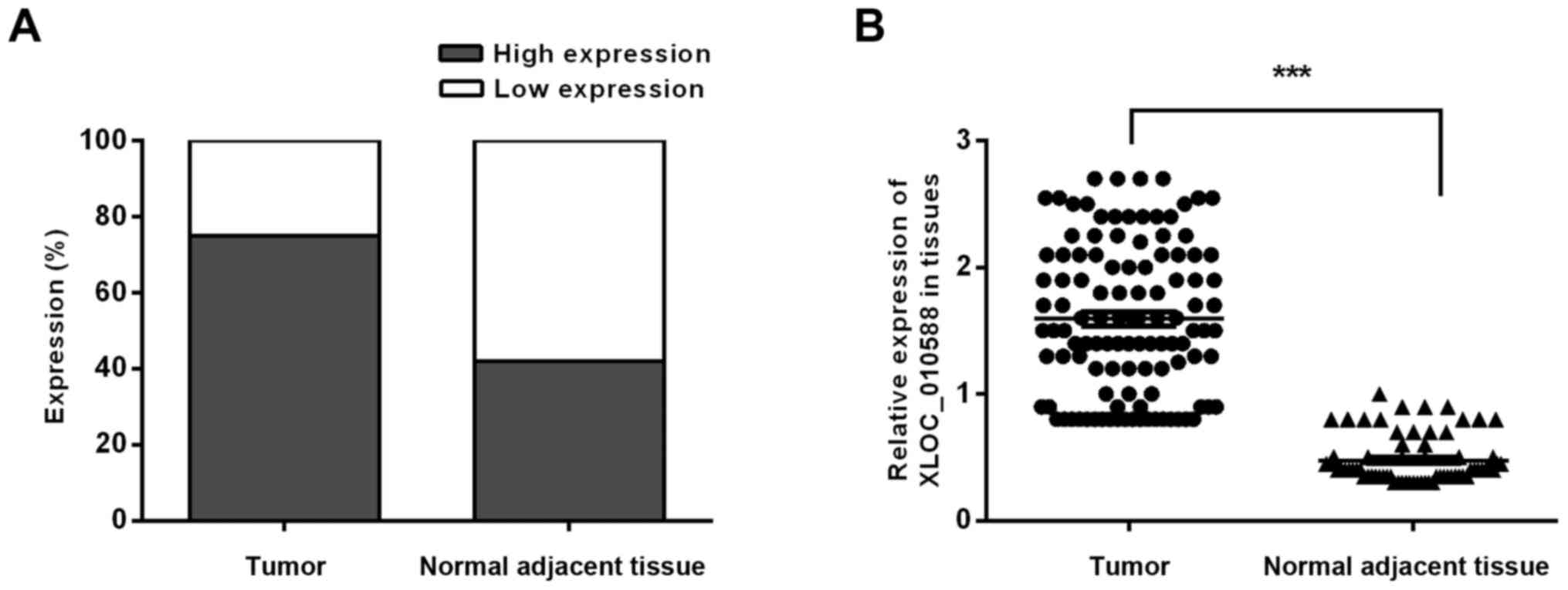
2), and the statistical results indicated that XLOC_010588 was
expressed at a high level in 58.02% of the cancer tissues but only
24.29% of normal adjacent tissues (P<0.001; Fig. 3). We subsequently analyzed the
expression level of XLOC_010588 with regard to clinicopathological
data in patients with CRC, and observed that the expression level
of XLOC_010588 was significantly associated with patient sex
(P=0.03), T-stage (P=0.013) and lymph node metastasis (P=0.002),
whereas it was not associated with patient age, tumor size, tumor
differentiation degree and distant metastasis (all P>0.05;
Table II).
 | Table I.Clinical and pathological
characteristics of patients with CRC in the present study. |
Table I.
Clinical and pathological
characteristics of patients with CRC in the present study.
| Parameters | No. of patients | % |
|---|
| Total number | 111 | 100 |
| Age (years) |
|
|
| ≤60 | 45 | 40.5 |
| >60 | 65 | 58.6 |
| Unknown | 1 | 0.9 |
| Sex |
|
|
| Male | 65 | 58.6 |
| Female | 46 | 41.4 |
| Size (cm) |
|
|
| ≤5 | 54 | 48.7 |
| >5 | 55 | 49.5 |
| Unknown | 2 | 1.8 |
| Histology
grade |
|
|
| I–II | 81 | 73 |
| III | 30 | 27 |
| T-stages |
|
|
| 1–2 | 45 | 40.5 |
| 3–4 | 35 | 31.5 |
| Unknown | 31 | 28 |
| N-stages |
|
|
| Yes | 33 | 29.7 |
| No | 71 | 64 |
| Unknown | 7 | 6.3 |
| M-stages |
|
|
| Yes | 4 | 3.6 |
| No | 107 | 96.4 |
| Table II.Correlation between the
clinicopathological features and the expression of XLOC-010588. |
Table II.
Correlation between the
clinicopathological features and the expression of XLOC-010588.
|
|
| XLOC
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | No. of
patients | High | Low | P-value |
|---|
| Age (years) | 110/111 |
|
| 0.846 |
| ≤60 | 45 | 23 | 22 |
|
| >60 | 65 | 32 | 33 |
|
| Sex | 111/111 |
|
| 0.03 |
| Male | 65 | 26 | 39 |
|
| Female | 46 | 28 | 18 |
|
| Size (cm) | 109/111 |
|
| 0.502 |
| ≤5 | 54 | 25 | 29 |
|
| >5 | 55 | 29 | 26 |
|
| Histology
grade | 111/111 |
|
| 0.862 |
| I–II | 81 | 39 | 42 |
|
| III | 30 | 15 | 15 |
|
| T-stages | 80/113 |
|
| 0.013 |
| 1–2 | 45 | 21 | 24 |
|
| 3–4 | 35 | 26 | 9 |
|
| N-stages | 111/111 |
|
| 0.002 |
| Yes | 33 | 24 | 9 |
|
| No | 71 | 28 | 43 |
|
| Unknown | 7 |
|
|
|
| M-stages | 111/111 |
|
| 0.065 |
| Yes | 4 | 4 | 0 |
|
| No | 107 | 57 | 50 |
|
Upregulation of XLOC_010588 is
associated with poor prognosis in patients with CRC
To ascertain the effect of XLOC_010588 on the
prognosis of CRC patients, the expression of XLOC_010588 was
analyzed with regard to the survival rate of CRC patients using the
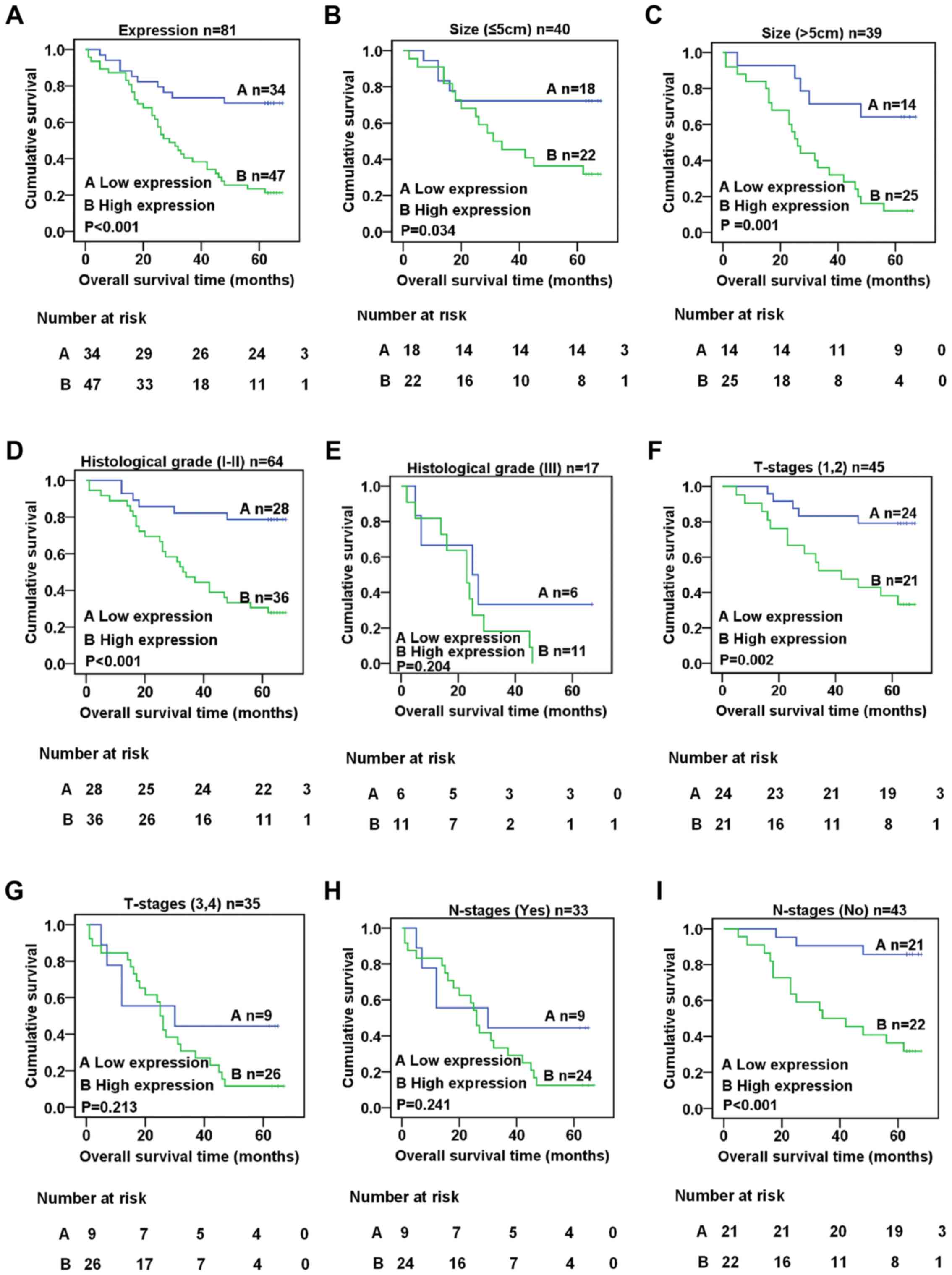
Kaplan-Meier method and the log-rank test. The results indicated
that the expression level of XLOC_010588 was closely associated
with the survival rate of CRC patients, with higher expression
being associated with a lower survival rate (log-rank = 16.353,
P<0.001, Fig. 4A). We further
performed a stratified analysis of XLOC_010588 expression with
regard to tumor size, histological grade, T- and N-stages. The
results revealed that, regardless of tumor size, the expression
level of XLOC_010588 and the survival period of CRC patients were
related (log-rank = 4.490, P=0.034, Fig. 4B; log-rank = 10.079, P=0.001,
Fig. 4C). Additionally, in CRC
patients with histological grade I/II tumors (log-rank = 14.474,
P<0.001; Fig. 4D), T-stage 1/2
tumors (log-rank = 9.455, P=0.002; Fig.
4F) and the absence of lymph node metastasis (log-rank =
13.089, P<0.001; Fig. 4I),
XLOC_010588 expression was significantly associated with the
survival rate, whereas no such association was observed in patients
with histological grade III tumors, T-stage 3/4 tumors and lymph
node metastasis (P>0.05; Fig. 4E, G
and H). To confirm whether the expression of XLOC_010588 is an
independent risk factor that affects the prognosis of CRC, we
performed univariate and multivariate analyses based on the data
referring to the Kaplan-Meier analysis of overall survival with
regard to patient clinicopathological features, and the results
revealed that high expression of XLOC_010588 was an independent
risk factor for poor prognosis in CRC (P=0.005; Table III).
| Table III.Summary of overall survival analyses
by univariate and multivariate COX regression analysis. |
Table III.
Summary of overall survival analyses
by univariate and multivariate COX regression analysis.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Factor | RR | 95% CI | P-value | RR | 95% CI | P-value |
|---|
| Age (years) | 1.504 | 0.826–2.736 | 0.182 |
|
|
|
| Sex | 0.825 | 0.480–1.416 | 0.485 |
|
|
|
| Tumor size | 1.496 | 0.868–2.577 | 0.147 |
|
|
|
| Histological
grade | 2.948 | 1.618–5.369 | <0.001 | 2.478 | 1.238–4.960 | 0.010 |
| T-stages | 3.020 | 1.735–5.257 | <0.001 | 4.484 | 0.962–20.89 | 0.056 |
| N-stages | 2.873 | 1.626–5.076 | <0.001 | 0.475 | 0.107–2.117 | 0.329 |
| M-stages | 2.380 | 0.850–6.667 | 0.099 |
|
|
|
| XLOC
expression | 3.848 | 1.901–7.788 | <0.001 | 3.233 | 1.431–7.302 | 0.005 |
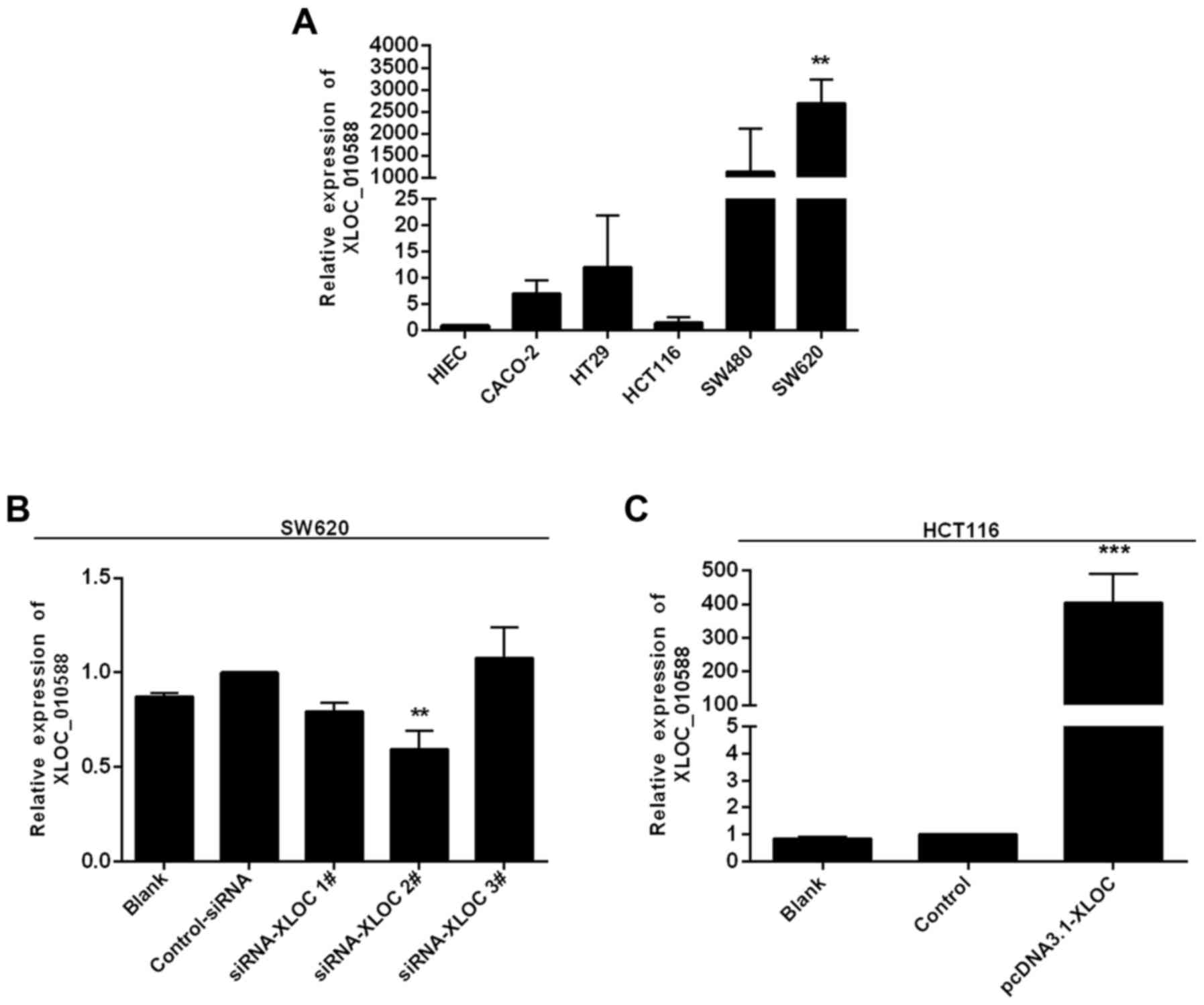
XLOC_010588 expression is generally
increased in human CRC cell lines
The literature reports that low expression of
XLOC_010588 indicates a poor prognosis and promotes cell
proliferation through upregulation of c-Myc in cervical cancer. To
define the role of XLOC_010588 in CRC, we performed an RT-qPCR
analysis to examine the expression of XLOC_010588 in 5 human CRC
cell lines, namely Caco-2, HT-29, SW480, SW620 and HCT116, and in
the normal human intestinal epithelial cell line HIEC-6. The
results demonstrated that XLOC_010588 expression in the majority of
the CRC cell lines was markedly higher than that in the HIEC cells
(Fig. 5A). Among the CRC cells,
XLOC_010588 expression was significantly increased in the SW620
cells (P<0.01), and thus these cells were used as a model for
RNAi knockdown of XLOC_010588. Conversely, HCT116 cells exhibited a
similar expression level of XLOC_010588 to that of the controls,
and thus were used as a model for pcDNA3.1-XLOC_010588
transfection. The SW620 and HCT116 cells were used in subsequent
experiments to assess cell migration and invasion in CRC in
vitro.
XLOC_010588 promotes cell invasion and
migration in CRC cell lines in vitro
Due to the markedly high expression of XLOC_010588
in CRC cell lines, particularly in SW620 cells, we hypothesized
that XLOC_010588 may be associated with the invasion and migration
of CRC, and potentially serve a crucial role in these processes. To
verify this hypothesis, we silenced XLOC_010588 in SW620 cells
using siRNA, while overexpressing XLOC_010588 in HCT116 cells,
after which RT-qPCR was performed at 48 h post-transfection to
confirm the silencing and overexpression efficiencies. We
determined that XLOC_010588 expression was markedly decreased
following transfection with si-XLOC_010588 (P=0.009; Fig. 5B), and increased following
transfection with pcDNA3.1-XLOC_010588 (P<0.001; Fig. 5C).
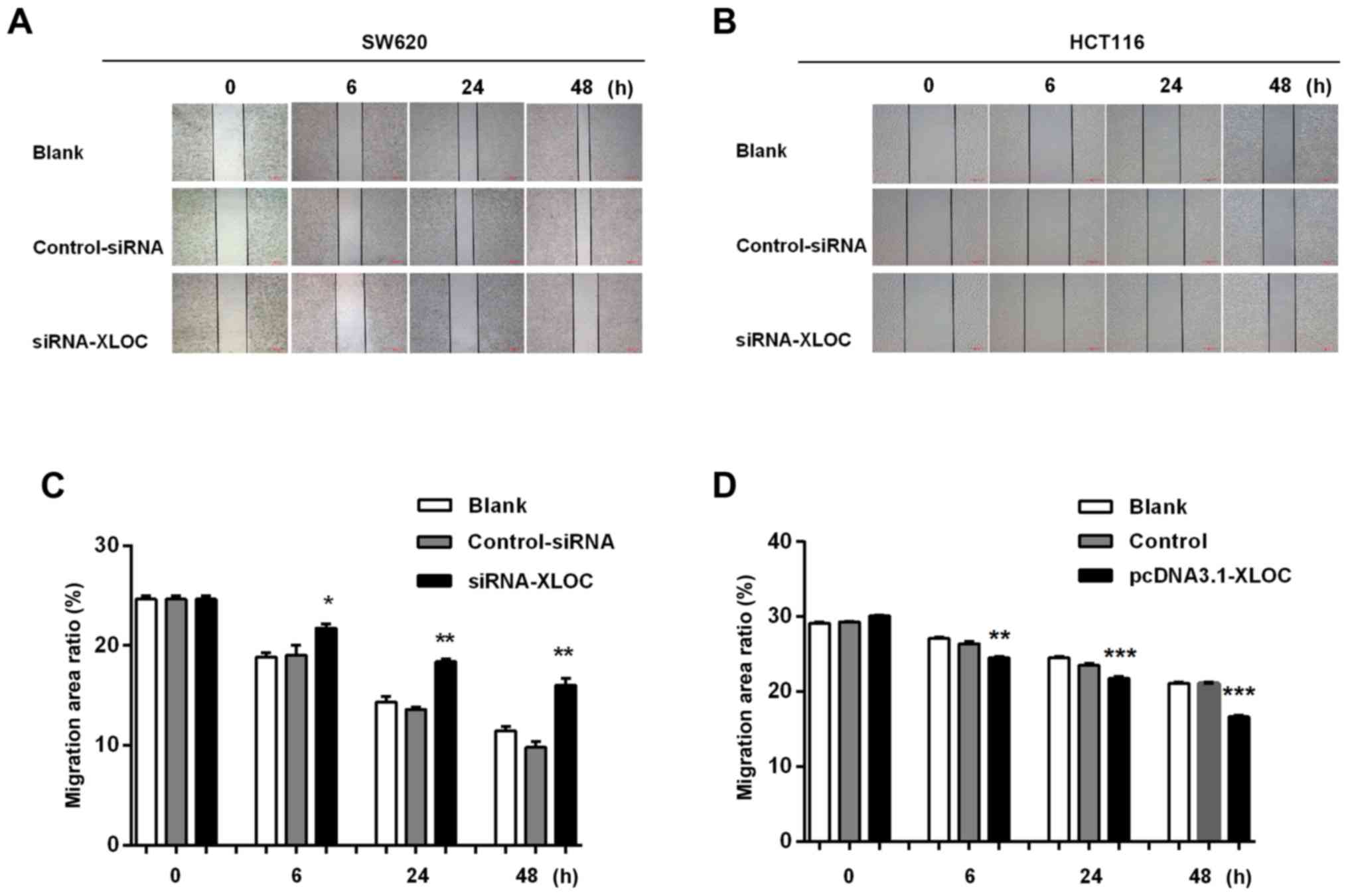
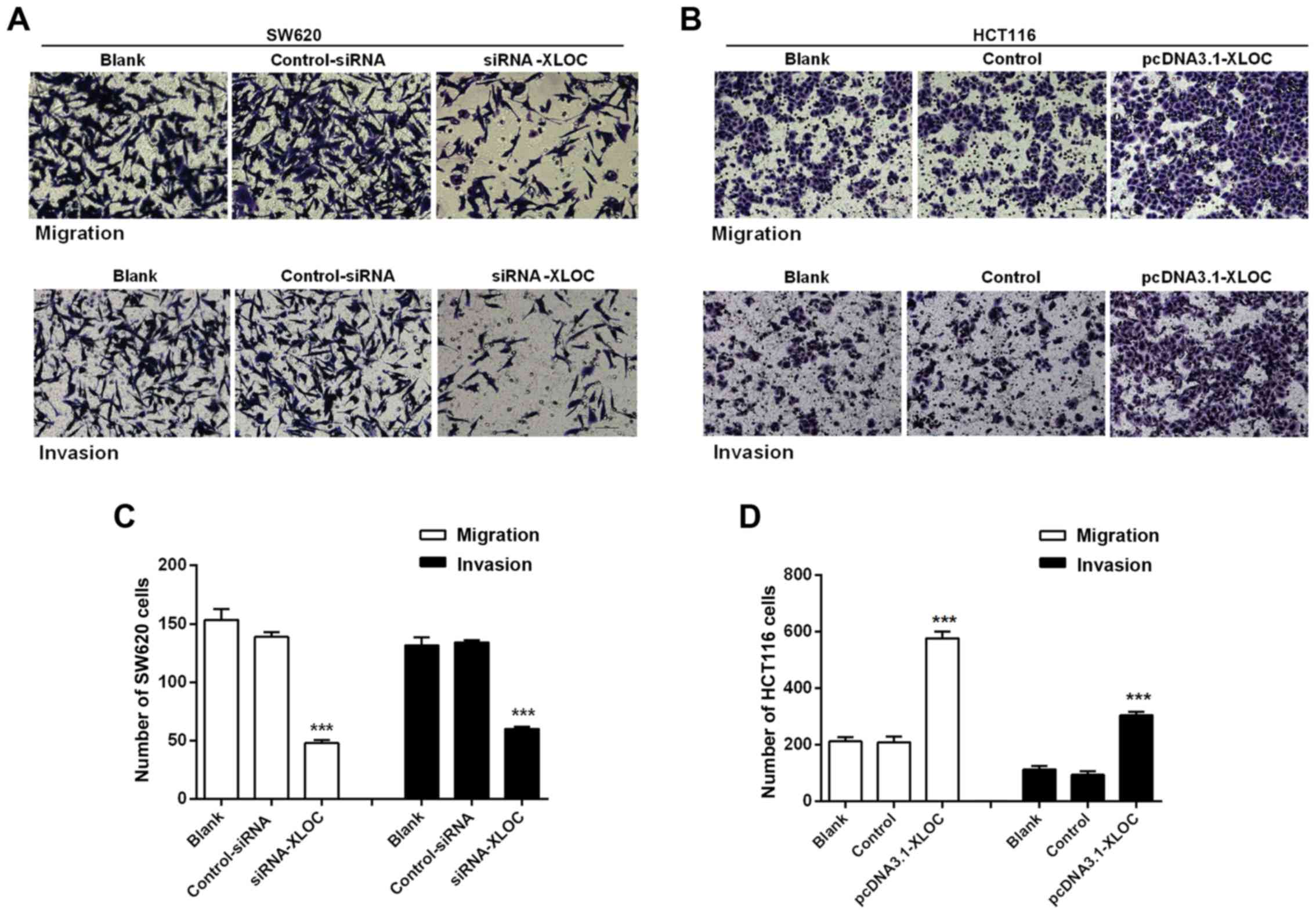
The subsequent wound-healing assay revealed that
XLOC_010588 silencing inhibited cell migration when compared with
si-control transfection (P<0.05; Fig. 6A and C), while the overexpression of
XLOC_010588 promoted cell migration (P<0.05; Fig. 6B and D). Furthermore, through the
Transwell assays, we observed that si-XLOC_010588 interference
significantly reduced cell migration and invasion abilities
compared with the si-control (P<0.001; Fig. 7A and C), while cell invasion and
migration abilities following XLOC_00588 overexpression were
significantly enhanced compared with the control group (P<0.001;
Fig. 7B and D).
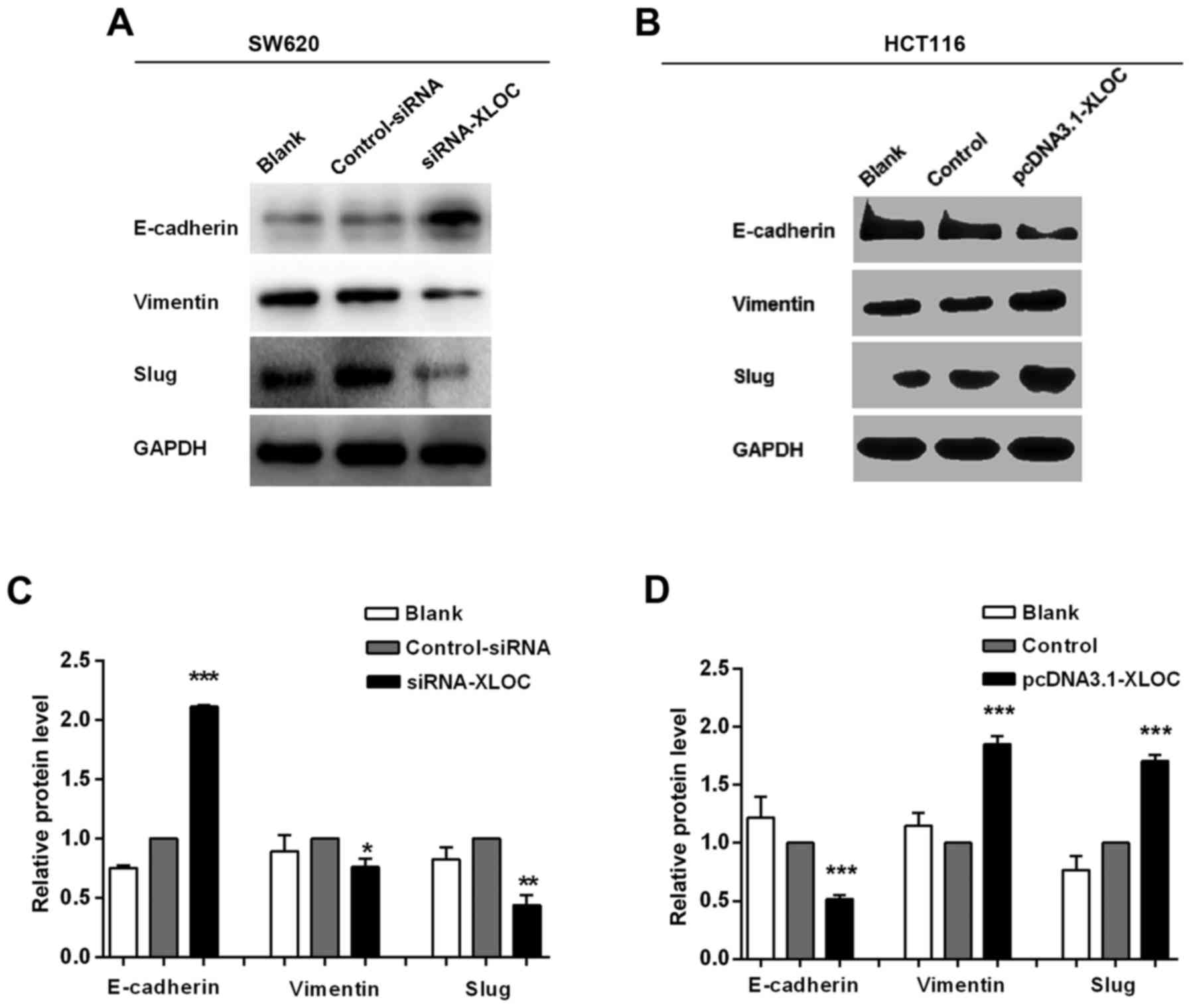
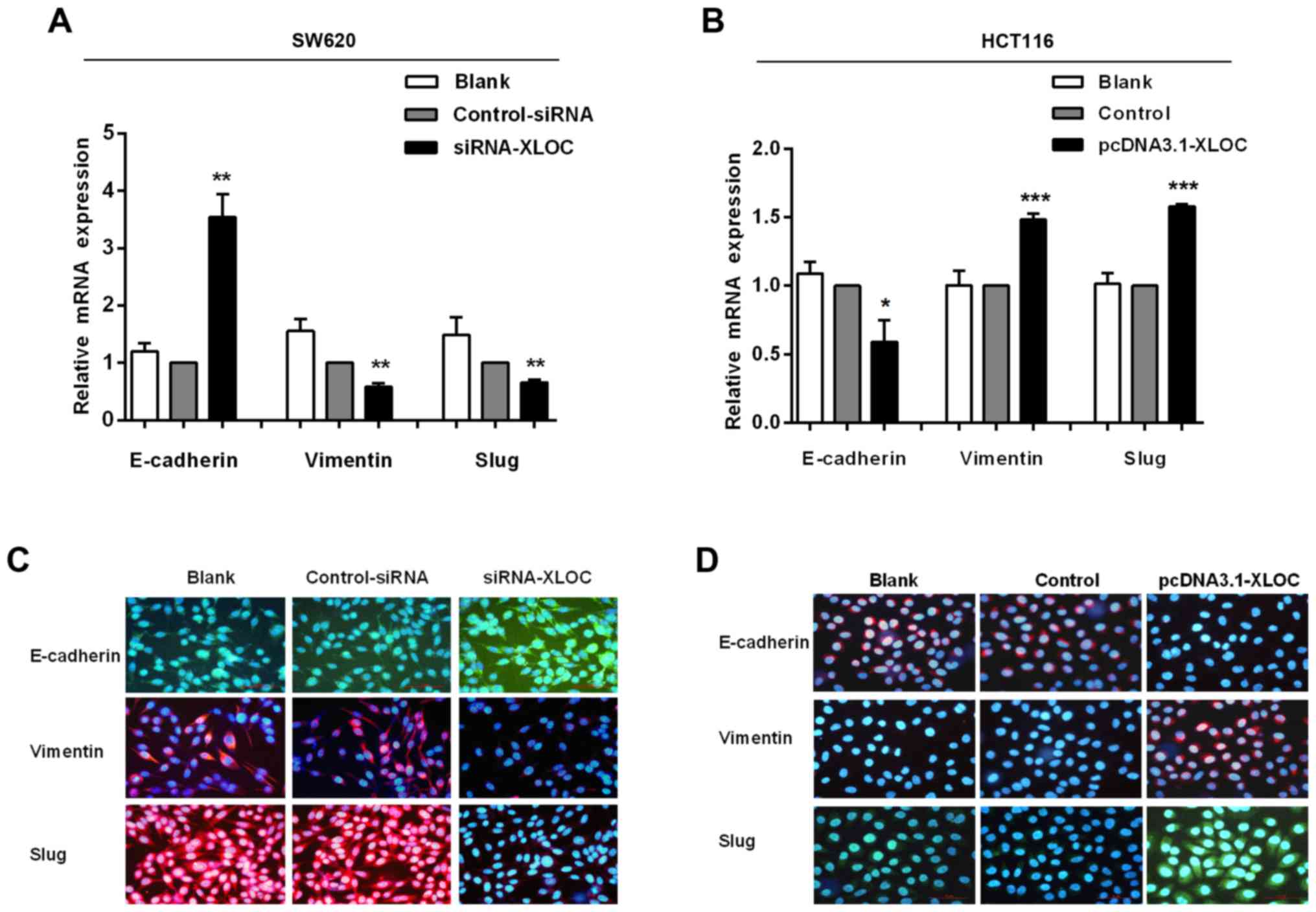
XLOC_010588 in CRC cells promotes the
EMT pathway in CRC cells
The migration and invasion assays demonstrated that
si-XLOC_010588 treatment significantly impaired cell migration and
invasion capacities compared with si-control, while the
overexpression cell model exhibited the opposite results. On
account of EMT being an important factor in cell migration and
invasion, and to further investigate the underlying mechanism of
XLOC_010588 in CRC cell migration and invasion, it was next
analyzed whether XLOC_010588 regulated EMT signaling. The
expression levels of E-cadherin, vimentin and Slug were analyzed by
western blotting (Fig. 8), RT-qPCR
(Fig. 9A and B) and
immunofluorescence (Fig. 9C and D).
We determined that the knockdown of XLOC_010588 markedly promoted
E-cadherin expression at the mRNA (P<0.01) and protein
(P<0.001) levels, while reducing the mRNA and protein levels of
Slug (both P<0.01) and vimentin (P<0.01 and P<0.05,
respectively).
Discussion
In recent years, diagnostic and therapeutic
techniques have undergone rapid development, and the mortality rate
of CRC patients has markedly decreased as a consequence, while the
quality of life of patients has improved substantially. However,
metastasis and recurrence remain key factors that lead to poor
prognosis of CRC patients, and further investigation into the
pathogenesis of CRC, to ultimately identify more effective
treatments for this fatal disease, is therefore required (1,24).
Recent studies have demonstrated that ncRNAs play multiple roles in
biological processes and human diseases; thus, they have become a
focus of research for biologists and clinicians (1,25).
Compared with protein-coding genes, non-coding genes appear to be
expressed at a lower level. However, evidence has ascertained that
these non-coding genes are closely associated with certain human
tumor types, including CRC (18).
Certain ncRNAs have been recognized as contributing
factors in CRC. However, there is still a lack of relevant evidence
to demonstrate their functions and underlying mechanisms in CRC
(22). Previous studies have
revealed a potential key role of lncRNAs in CRC. Notably, an
experiment by Liao et al revealed that XLOC_010588
expression was downregulated in cervical cancer, which was the
first time this phenomenon was reported (16). However, the results of the present
study revealed that XLOC_010588 was expressed at a higher level in
cancer tissues compared with adjacent normal tissues. Furthermore,
it was also overexpressed in a panel of CRC cell lines, and thus we
suspect that XLOC_010588 has a cancer-specific expression pattern,
indicating that XLOC_010588 may act as an oncogene or tumor
suppressor in different cancer types (7,17). A
survey of the available literature indicates that XLOC_010588
promotes cell proliferation through upregulation of c-Myc in
cervical cancer (16). In the
present experiment, we attempted to ascertain, besides its role in
tumor proliferation, whether XLOC_010588 participates in the
processes of invasion and migration in CRC (6). EMT is the process by which tumor cells
differentiate into mesenchymal cells, which have an increased
capacity to obtain movement ability; during this process, cells
gradually reorganize or downregulate their cytoskeleton and basal
epithelial-specific epithelial genes, including (E)-cadherin, while
simultaneously upregulating expression of vimentin and Slug, with
Slug further suppressing the expression of E-cadherin. This EMT
process is typically observed during tumor invasion and migration
(26). Recently, a number of
studies have established lncRNAs to play a dominant role in the
regulation of EMT (8), and a range
of EMT-related lncRNAs have been identified, including
lncRNA-GHET1, lncRNA-SPRY4 and lncRNA-TUG1, among others.
Therefore, we speculated that XLOC_010588 is also a potential
EMT-related lncRNA (7). In the
present study, we demonstrated that the abilities of CRC cells to
invade and migrate were weakened when XLOC_010588 was knocked down,
while these abilities were enhanced when XLOC_010588 was
overexpressed. We also ascertained the expression levels of EMT
markers in SW620 cells and HCT116 cells, revealing that the
overexpression of XLOC_010588 decreased the expression of
E-cadherin and increased the expression of vimentin and Slug, while
the knockdown of XLOC_010588 reversed these effects (17,27).
Overall these results indicated that XLOC_010588 affected the
invasion and migration of CRC by regulating EMT-related genes.
In conclusion, our experiment revealed the
relationship between XLOC_010588 expression and CRC for the first
time. We established that XLOC_010588 was expressed at a higher
level in cancer tissues compared with adjacent normal tissues, and
that this high expression was closely associated with CRC T-stage
and lymph node metastasis. However, due to there being a small
sample size of patients with distant metastasis, we did not obtain
significant data regarding XLOC_010588 expression and distant
metastasis, and thus further research on this is warranted.
Kaplan-Meier analysis also revealed that XLOC_010588 overexpression
in tumor tissues was strongly connected with poorer prognosis of
CRC patients. These results indicated that XLOC_010588 promotes the
progression of CRC, and may serve as a negative prognostic factor
for CRC patients. Furthermore, our study demonstrated that the
expression of XLOC_010588 was markedly higher in CRC cells, and
that XLOC_010588 promoted the invasion and migration of CRC cells.
It was also revealed that XLOC_010588 may regulate the progression
of CRC via the EMT pathway (28).
In our experiments, we revealed a crucial role of XLOC_010588 in
the invasion and migration of CRC. However, the specific mechanism
underlying this effect of XLOC_010588, potentially involving
regulation of the EMT pathway, requires further investigation in
our future studies. In conclusion, the present findings indicated
that XLOC_010588 is a functional oncogene, and revealed that
XLOC_010588 may be a novel therapeutic target in patients with CRC
(29,30).
Acknowledgements
The authors would like to thank Dr LJ and SL
(Department of Pharmacology, School of Pharmaceutical Sciences,
China Medical University) for their help with the statistical
analysis.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL, LZ and MS conceived and designed the study. YL
and TG performed the experiments. YL wrote the study. YL, MS, LZ,
YZ, LG, HZhang, HZhou and PM reviewed and edited the manuscript.
All authors read and approved the manuscript.
Ethics approval and consent to
participate
In the present study, all tissue microarrays were
purchased from Shanghai Outdo Biotech Co., Ltd., and were approved
by the local Ethics Committee (Zhejiang Taizhou Hospital Ethics
Committee, Zhejiang, China). Due to the retrospective nature of the
study, the Ethics Committee waived the requirement for written
informed consent from the patients.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang F, Ni H, Sun F, Li M and Chen L:
Overexpression of lncRNA AFAP1-AS1 correlates with poor prognosis
and promotes tumorigenesis in colorectal cancer. Biomed
Pharmacother. 81:152–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen X, Liu B, Yang R, Guo Y, Li F, Wang L
and Hu H: Integrated analysis of long non-coding RNAs in human
colorectal cancer. Oncotarget. 7:23897–23908. 2016.PubMed/NCBI
|
|
3
|
Sun L, Xue H, Jiang C, Zhou H, Gu L, Liu
Y, Xu C and Xu Q: lncRNA DQ786243 contributes to proliferation and
metastasis of colorectal cancer both in vitro and in vivo. Biosci
Rep. 36:e003282016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen X, Zhu H, Wu X, Xie X, Huang G, Xu X,
Li S and Xing C: Downregulated pseudogene CTNNAP1 promote tumor
growth in human cancer by downregulating its cognate gene CTNNA1
expression. Oncotarget. 7:55518–55528. 2016.PubMed/NCBI
|
|
5
|
Han D, Gao X, Wang M, Qiao Y, Xu Y, Yang
J, Dong N, He J, Sun Q, Lv G, et al: Long noncoding RNA H19
indicates a poor prognosis of colorectal cancer and promotes tumor
growth by recruiting and binding to eIF4A3. Oncotarget.
7:22159–22173. 2016.PubMed/NCBI
|
|
6
|
Ye LC, Chen T, Zhu DX, Lv SX, Qiu JJ, Xu
J, Yuan FL and Wei Y: Downregulated long non-coding RNA CLMAT3
promotes the proliferation of colorectal cancer cells by targeting
regulators of the cell cycle pathway. Oncotarget. 7:58931–58938.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kong J, Sun W, Li C, Wan L, Wang S, Wu Y,
Xu E, Zhang H and Lai M: Long non-coding RNA LINC01133 inhibits
epithelial-mesenchymal transition and metastasis in colorectal
cancer by interacting with SRSF6. Cancer Lett. 380:476–484. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou J, Li X, Wu M, Lin C, Guo Y and Tian
B: Knockdown of long noncoding RNA GHET1 inhibits cell
proliferation and invasion of colorectal cancer. Oncol Res.
23:303–309. 2016. View Article : Google Scholar
|
|
9
|
Song X, Cao G, Jing L, Lin S, Wang X,
Zhang J, Wang M, Liu W and Lv C: Analysing the relationship between
lncRNA and protein-coding gene and the role of lncRNA as ceRNA in
pulmonary fibrosis. J Cell Mol Med. 18:991–1003. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang F, Xue X, Bi J, Zheng L, Zhi K, Gu Y
and Fang G: Long noncoding RNA CCAT1, which could be activated by
c-Myc, promotes the progression of gastric carcinoma. J Cancer Res
Clin Oncol. 139:437–445. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nie FQ, Zhu Q, Xu TP, Zou YF, Xie M, Sun
M, Xia R and Lu KH: Long non-coding RNA MVIH indicates a poor
prognosis for non-small cell lung cancer and promotes cell
proliferation and invasion. Tumour Biol. 35:7587–7594. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wan L, Kong J, Tang J, Wu Y, Xu E, Lai M
and Zhang H: HOTAIRM1 as a potential biomarker for diagnosis of
colorectal cancer functions the role in the tumour suppressor. J
Cell Mol Med. 20:2036–2044. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo W, Wang Q, Zhan Y, Chen X, Yu Q, Zhang
J, Wang Y, Xu XJ and Zhu L: Transcriptome sequencing uncovers a
three-long noncoding RNA signature in predicting breast cancer
survival. Sci Rep. 6:279312016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou M, Diao Z, Yue X, Chen Y, Zhao H,
Cheng L and Sun J: Construction and analysis of dysregulated
lncRNA-associated ceRNA network identified novel lncRNA biomarkers
for early diagnosis of human pancreatic cancer. Oncotarget.
7:56383–56394. 2016.PubMed/NCBI
|
|
15
|
Tu Z, He D, Deng X, Xiong M, Huang X, Li
X, Hao L, Ding Q and Zhang Q: An eight-long non-coding RNA
signature as a candidate prognostic biomarker for lung cancer.
Oncol Rep. 36:215–222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liao LM, Sun XY, Liu AW, Wu JB, Cheng XL,
Lin JX, Zheng M and Huang L: Low expression of long noncoding
XLOC_010588 indicates a poor prognosis and promotes proliferation
through upregulation of c-Myc in cervical cancer. Gynecol Oncol.
133:616–623. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang L, Zhao Z, Feng W, Ye Z, Dai W, Zhang
C, Peng J and Wu K: Long non-coding RNA TUG1 promotes colorectal
cancer metastasis via EMT pathway. Oncotarget. 7:51713–51719.
2016.PubMed/NCBI
|
|
18
|
Huang G, Wu X, Li S, Xu X, Zhu H and Chen
X: The long noncoding RNA CASC2 functions as a competing endogenous
RNA by sponging miR-18a in colorectal cancer. Sci Rep. 6:265242016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou M, Guo M, He D, Wang X, Cui Y, Yang
H, Hao D and Sun J: A potential signature of eight long non-coding
RNAs predicts survival in patients with non-small cell lung cancer.
J Transl Med. 13:2312015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Y, Zheng ZP, Li H, Zhang HQ and Ma FQ:
ANRIL is associated with the survival rate of patients with
colorectal cancer, and affects cell migration and invasion in
vitro. Mol Med Rep. 14:1714–1720. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang P, Xu ZP, Chen T and He ZY: Long
noncoding RNA expression profile analysis of colorectal cancer and
metastatic lymph node based on microarray data. Onco Targets Ther.
9:2465–2478. 2016.PubMed/NCBI
|
|
22
|
Chen N, Guo D, Xu Q, Yang M, Wang D, Peng
M, Ding Y, Wang S and Zhou J: Long non-coding RNA FEZF1-AS1
facilitates cell proliferation and migration in colorectal
carcinoma. Oncotarget. 7:11271–11283. 2016.PubMed/NCBI
|
|
23
|
Wu H, Guan S, Sun M, Yu Z, Zhao L, He M,
Zhao H, Yao W, Wang E, Jin F, et al: Ano1/TMEM16A overexpression is
associated with good prognosis in PR-positive or HER2-negative
breast cancer patients following Tamoxifen treatment. PLoS One.
10:e01261282015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie X, Tang B, Xiao YF, Xie R, Li BS, Dong
H, Zhou JY and Yang SM: Long non-coding RNAs in colorectal cancer.
Oncotarget. 7:5226–5239. 2016.PubMed/NCBI
|
|
25
|
Yan B, Yao J, Liu JY, Li XM, Wang XQ, Li
YJ, Tao ZF, Song YC, Chen Q and Jiang Q: lncRNA-MIAT regulates
microvascular dysfunction by functioning as a competing endogenous
RNA. Circ Res. 116:1143–1156. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen F, Cai WS, Feng Z, Chen JW, Feng JH,
Liu QC, Fang YP, Li KP, Xiao HQ, Cao J, et al: Long non-coding RNA
SPRY4-IT1 pormotes colorectal cancer metastasis by regulate
epithelial-mesenchymal transition. Oncotarget. 8:14479–14486.
2016.
|
|
27
|
Sun J, Ding C, Yang Z, Liu T, Zhang X,
Zhao C and Wang J: The long non-coding RNA TUG1 indicates a poor
prognosis for colorectal cancer and promotes metastasis by
affecting epithelial-mesenchymal transition. J Transl Med.
14:422016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sunamura N, Ohira T, Kataoka M, Inaoka D,
Tanabe H, Nakayama Y, Oshimura M and Kugoh H: Regulation of
functional KCNQ1OT1 lncRNA by β-catenin. Sci Rep. 6:206902016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bian Z, Jin L, Zhang J, Yin Y, Quan C, Hu
Y, Feng Y, Liu H, Fei B, Mao Y, et al: lncRNA-UCA1 enhances cell
proliferation and 5-fluorouracil resistance in colorectal cancer by
inhibiting miR-204-5p. Sci Rep. 6:238922016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang P, Chen T, Xu Z, Zhu H, Wang J and He
Z: Long noncoding RNA GAPLINC promotes invasion in colorectal
cancer by targeting SNAI2 through binding with PSF and NONO.
Oncotarget. 7:42183–42194. 2016.PubMed/NCBI
|