Introduction
With an estimated 230,000 new cases and 40,000
deaths in 2013, breast cancer has the highest incidence and is the
second leading cause of cancer-related death among women in the
United States (1). Four subtypes of
breast cancers, namely luminal A, luminal B, HER2+ and
basal-like (significantly but not completely overlaps with the
triple-negative breast cancer, TNBC), are classified according to
the intrinsic gene expression profile (2,3). While
the luminal subtypes respond well to hormone therapies, over 50% of
patients with HER2+ breast cancer develop
trastuzumab-resistance within 1 to 2 years of treatment (4,5). More
than 70% of TNBC patients have residual invasive disease after
neoadjuvant chemotherapy and are at high risk of disease relapse
(6). Recent evidence supports that
a small fraction of cancer cells, termed cancer stem cells (CSCs),
are capable of self-renewing and differentiating into non-stem
cancer cells and are responsible for tumor initiation, drug
resistance and metastasis (7–10).
Therefore, combining CSC-targeting agents with conventional
chemotherapies seems to be a promising strategy for eradicating
both CSCs and bulk tumor cells (11,12).
Reprogramming of energy metabolism is one of the
hallmarks of cancer (13). Over 80
years ago, Otto Warburg observed that cancer cells favored aerobic
glycolytic metabolism in the presence of oxygen (14). Warburg hypothesized that cancer
resulted from impaired cellular mitochondrial metabolism. It is
clear now that the Warburg effect is not due to the impairment of
mitochondrial function in tumors. Indeed, depletion of
mitochondrial DNA has been shown to decrease colony formation in
soft agar and tumor initiation in mice (15–19),
which are the key indicators of CSCs. Recent studies have also
demonstrated that mitochondrial features of CSCs differ from those
of non-stem cancer cells (20–22),
and attenuating mitochondrial metabolism could suspend tumor
metastasis and prolong tumor latency in xenograft models (19,23).
This phenomenon indicates that mitochondria are functionally
indispensable to sustaining CSCs. Therefore, targeting mitochondria
is emerging as a new strategy for eradicating CSCs.
Doxycycline is a commonly used tetracycline analogue
of antibiotics. With ideal pharmacokinetics and minor side effects,
doxycycline has been used in clinics for five decades. The
mechanism of doxycycline's action is binding to mitochondrial
ribosome, which then disrupts the biogenesis of bacterial
mitochondria. In addition to bacterial mitochondria, doxycycline
has also been reported to affect mitochondria in eukaryotes
(24). In cancer, doxycycline was
found to inhibit the self-renewal ability of CSCs in many types of
cancer, including breast cancer (25–28),
indicating the potentiality of using the ‘old’ antibiotic for a new
treatment-targeting CSCs.
Although doxycycline-mediated CSC inhibition has
been linked to mitochondria (20,24,25),
it remains unknown what type of CSCs doxycycline could inhibit.
Breast cancer stem cells (BCSCs) can transition between two
phenotypic states. One is a more proliferative epithelial-like
state characterized by the expression of the CSC marker aldehyde
dehydrogenase (ALDH) and the other is a more quiescent
mesenchymal-like state characterized by the expression
CD44+/CD24− (29). In this study, we selected BT474,
SK-BR-3, SUM149, and SUM159 breast cancer cell lines to examine
doxycycline's effects on BCSCs. BT474 and SK-BR-3 are both
HER2+ breast cancer cell lines and according to the
literature and our previous publication, this subtype has a higher
number of ALDH+ epithelial-like BCSCs compared to other
subtypes but does not have the CD44+/CD24−
mesenchymal-like BCSC population (30,31).
In contrast, SUM159 is a Claudin-low TNBC cell line, which has a
high percentage of mesenchymal-like bulk tumor cells that are also
CD44+/CD24− (30). As a result, the
CD44+/CD24− markers cannot be used to define
the mesenchymal-like BCSCs in the Claudin-low subtype. SUM149, on
the other hand, is a basal-like TNBC and has both ALDH+
and CD44+/CD24− BCSCs. Hence, SUM149 is a
suitable cell line for testing the effects of doxycycline on
ALDH+ as well as CD44+/CD24− BCSC
populations.
In the present study, we report that doxycycline can
reduce the ALDH+ BCSC population. Mechanistically, our
results suggest that doxycycline inhibits ALDH+ BCSCs
via inhibiting reactive oxygen species (ROS) production and their
downstream p38 MAPK signaling pathway.
Materials and methods
Cell lines and chemicals
BT474 (ATCC, Manassas, VA, USA) and SK-BR-3 (ATCC)
cells were grown in RPMI-1640 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% FBS and 1X
antibiotic-antimycotic (Invitrogen; Thermo Fisher Scientific,
Inc.). SUM149 and SUM159 cell lines were kindly provided by Dr Max
Wicha (University of Michigan, Ann Arbor, MI, USA). SUM149 and
SUM159 cells were grown in F12 (Invitrogen; Thermo Fisher
Scientific, Inc.) containing 5% FBS, 1X antibiotic-antimycotic, 5
µg/ml of insulin (Invitrogen; Thermo Fisher Scientific, Inc.) and 1
µg/ml of hydrocortison (Sigma-Aldrich, St. Louis, MO, USA). Cells
were cultured in a 5% CO2 incubator at 37°C. p38 MAPK
inhibitor SB203580 was purchased from Cayman Chemical Co. (Ann
Arbor, MI, USA). Doxycycline hyclate was purchased from
Sigma-Aldrich.
Mammosphere formation assay
Mammosphere formation was performed as previously
described (32). Single cells were
seeded in low-attachment 6-well plates (Corning Inc., Corning, NY,
USA) at a density of 5,000 cells/well. Cells were cultured in 2 ml
of MammoCult™ (Stemcell Technologies, Inc., Vancouver, BC, Canada)
with doxycycline from 0–10 µM. Doxycycline was replenished every 2
days, and mammospheres were counted on day 6. To test the
self-renewal ability of CSCs, secondary mammosphere formation was
performed in the absence of doxycycline. Briefly, primary spheres
were dissociated to single cells enzymatically (trypsin) and
mechanically (23G needle). Secondary mammosphere formation was
performed by plating 5,000 cells/well of the dissociated single
cells from the primary mammospheres.
Aldefluor assay
Cells were treated with 0, 0.1, 1 or 10 µg/ml of
doxycycline for 7 days. The aldehyde dehydrogenase (ALDH) activity
was then determined by Aldefluor assay (StemCell Technologies Inc.,
Cambridge, MA, USA) according to the manufacturer's instructions.
Diethylaminobenzaldehyde (DEAB) was used as a negative control for
gating. To test the importance of the p38 pathway in maintaining
ALDH+ BCSCs, BT474 cells were treated with SB203580, a
p38-specific inhibitor, for 2 days and then Aldefluor assay was
conducted. To inhibit chemotherapy-induced Aldefluor-positive CSCs,
cells were pretreated with 10 µM of doxycycline for 3 days, and
then were treated with a combination of doxycycline and 10 nM of
paclitaxel for another 4 days.
CD44 and CD24 analysis
Cells were treated with 0, 0.1, 1 or 10 µg/ml of
doxycycline for 7 days, and then were harvested for CD44 (BD
Biosciences, San Jose, CA, USA) and CD24 (BioLegend, Inc., San
Diego, CA, USA) antibody staining. The cells were then analyzed by
flow cytometry.
Analysis of reactive oxygen species
(ROS)
Cells were treated with 10 µg/ml of doxycycline for
7 days and then ROS were determined by a 2′,7′-dichlorofluorescin
diacetate (DCFDA)-based kit (Abcam, Cambridge, MA, USA) according
to the manufacturer's instructions. Briefly, the cells were
incubated with 20 µM of DCFDA at 37°C for 30 min. Samples were then
spiked with 300 µl of ice-cold 1X buffer containing DAPI and kept
on ice before the ROS level was measured by flow cytometry.
Immunoblotting
Breast cancer cells were treated with 0, 0.1, 1 or
10 µg/ml of doxycycline for 7 days. Cells were then lysed using
RIPA buffer containing proteinase inhibitor cocktail (Thermo Fisher
Scientific, Inc.) and Calbiochem® phosphatase inhibitors
(MilliporeSigma, Burlington, MA, USA). Proteins were separated by
SDS-PAGE and probed with antibodies. Phosphorylated p38 MAPK
(1:1,000; cat. no. 4511; rabbit mAb), p38 MAPK (1:1,000; cat. no.
8690; rabbit mAb) and vinculin (1:2,000; cat. no. 13901; rabbit
mAb) antibodies were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Statistical analysis
Two-tailed Student's t-test was used to compare the
statistical difference between two groups. One-way ANOVA was used
if the comparison involved more than two groups. A P-value <0.05
was considered to indicate statistical significance.
Results
Doxycycline inhibits ALDH+
BCSCs
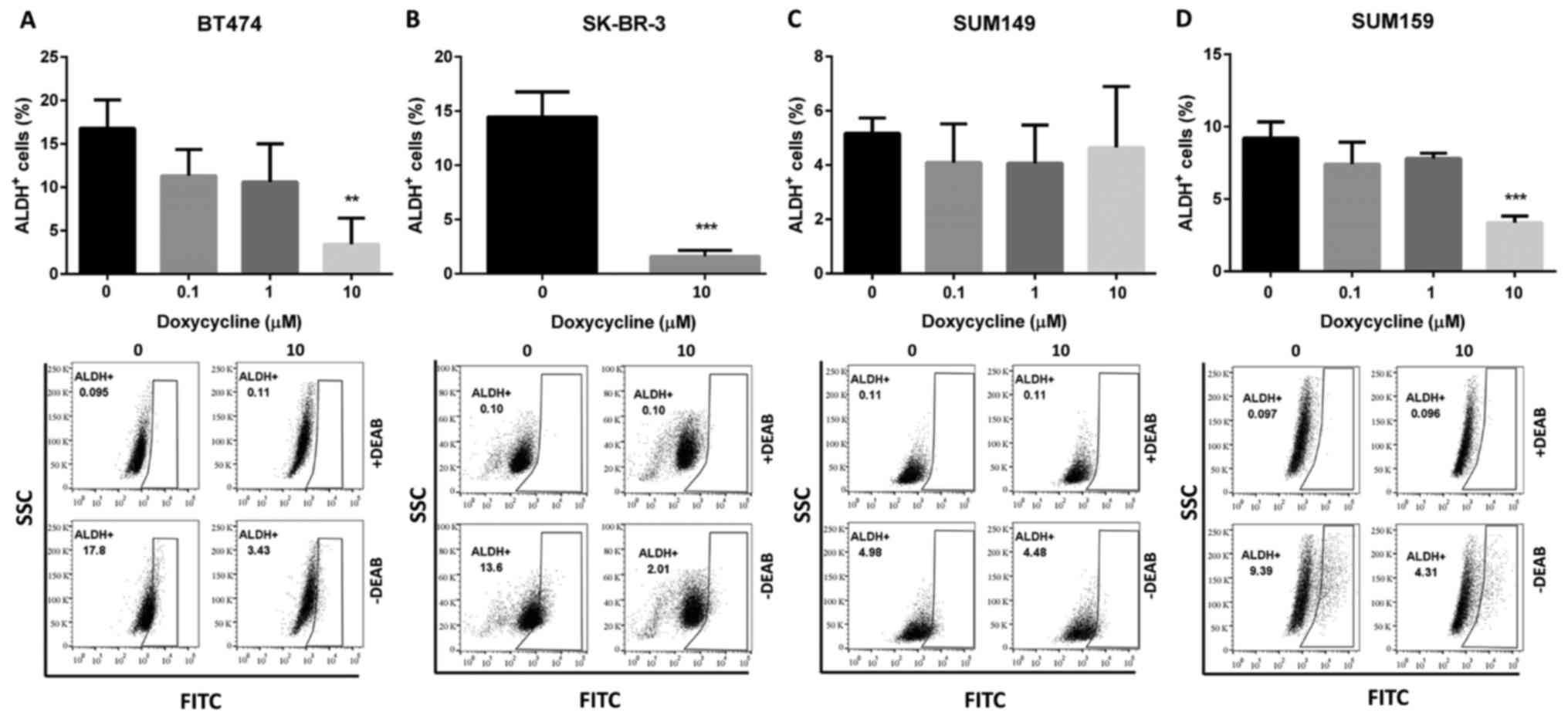
To test doxycycline's ability to inhibit
ALDH+ BCSCs, we treated breast cancer cell lines with
doxycycline and then measured ALDH activity in these cells. ALDH is
an important biomarker of CSCs in many types of cancer (33). In breast cancer, cells with high
ALDH activity have self-renewal ability to regenerate tumors that
recapitulate the heterogeneity of the parental tumors (8). Our results demonstrated that
doxycycline at 10 µM significantly decreased the percentage of
cells with high ALDH activity in the BT474, SK-BR-3 and SUM159
cells (Fig. 1). These results
suggested that doxycycline could be used to target ALDH+
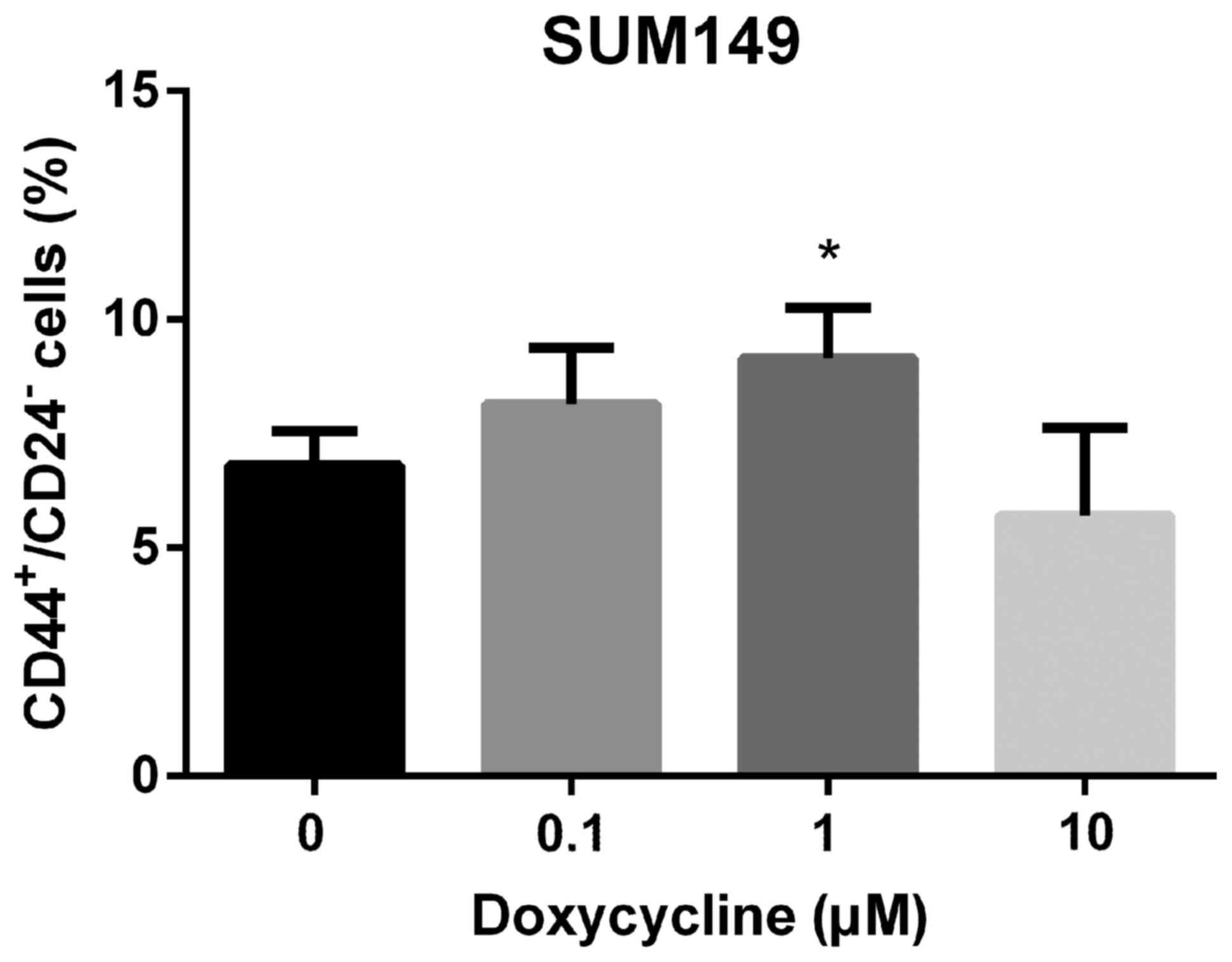
BCSCs. However, in SUM149 cells, doxycycline did not decrease
ALDH+ (Fig. 1) or
CD44+/CD24− (Fig.
2) BCSCs. This result may be due to the characteristics of the
SUM149 cell line (see Discussion).
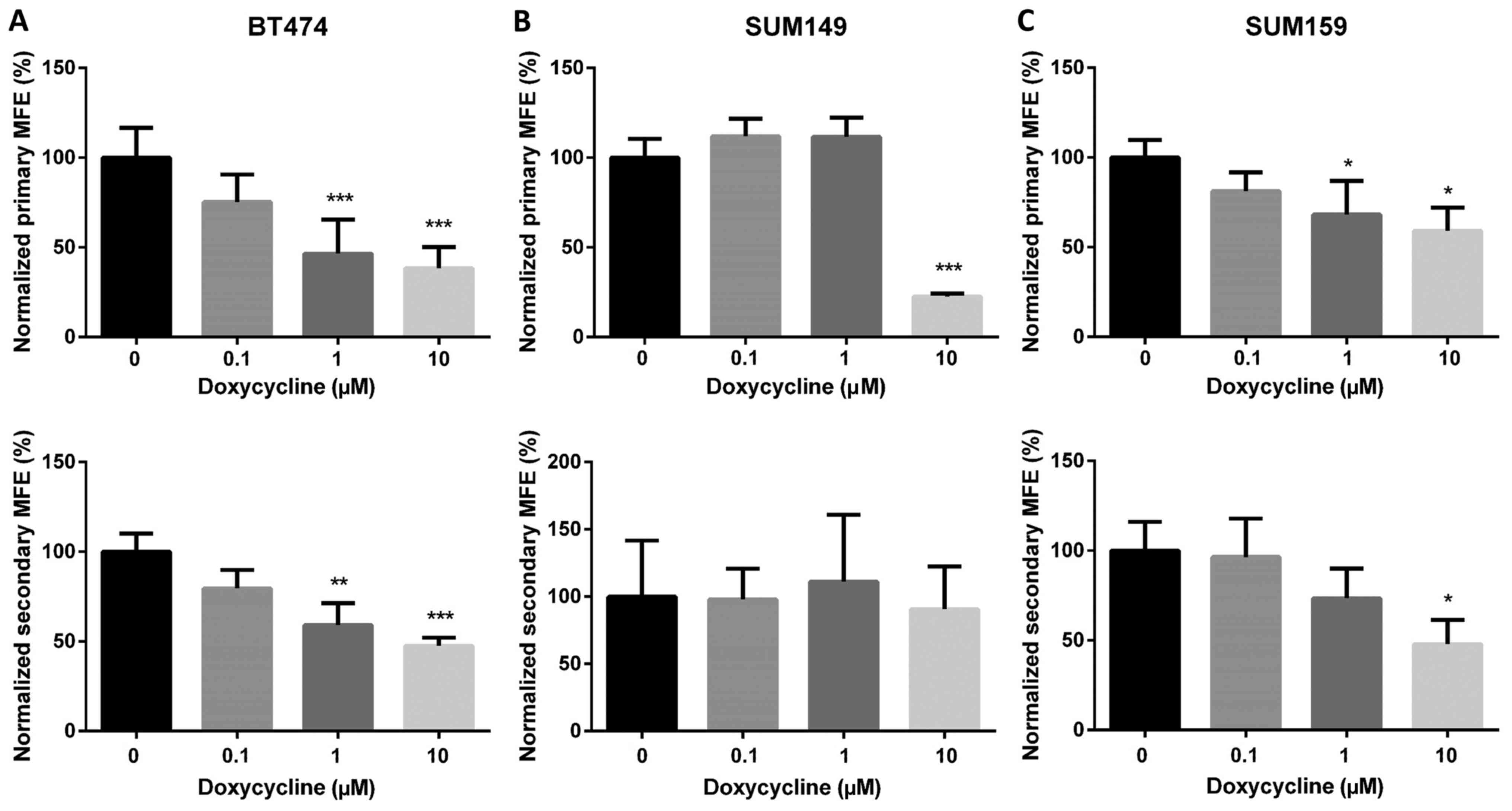
To further confirm whether doxycycline could
functionally inhibit BCSCs, we treated BT474, SUM149 and SUM159
cells with various concentrations of doxycycline in the primary
mammmosphere culture. Mammosphere formation is an in vitro
surrogate assay to evaluate self-renewal ability of BCSCs. In BT474
and SUM159, primary mammosphere formation was significantly
inhibited by doxycycline in a concentration-dependent manner,
whereas in SUM149, it was inhibited only at 10 µM (Fig. 3). The primary mammospheres were then
dissociated and reseeded to form secondary mammospheres in the
absence of doxycycline. In the secondary mammosphere culture, a 50%
decrease in mammospheres was observed in the 10 µM
doxycycline-pretreated BT474 and SUM159 cells (Fig. 3), indicating that doxycycline could
inhibit the self-renewal ability of BCSCs in these cell lines.
Doxycycline inhibits reactive oxygen
species and their downstream p38 signaling
In mammalian cells, doxycycline inhibits
mitochondrial biogenesis by binding to 28S small mitochondrial
ribosome (24,34). Mitochondria is the main organelle of
ROS generation. High mitochondrial mass (20) and elevated ROS levels (35) have been reported to sustain
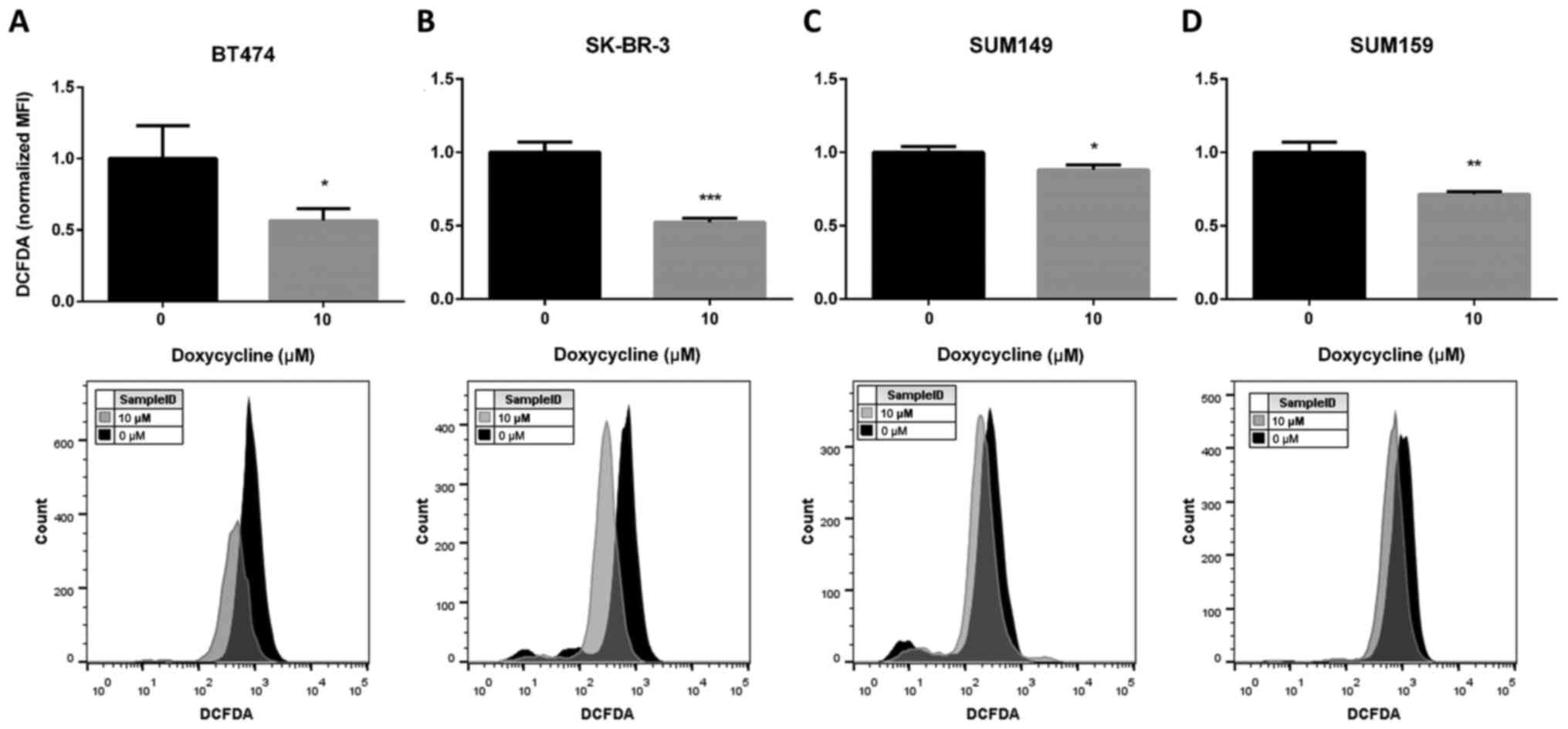
ALDH+ CSCs. We demonstrated that doxycycline treatment
significantly inhibited ALDH+ BCSCs (Fig. 1). Therefore, we hypothesized that
doxycycline inhibited ALDH+ BCSCs via ROS attenuation.
To test if doxycycline could decrease cellular ROS levels, we
performed DCFDA assays after doxycycline treatment and analyzed
samples by flow cytometry. As expected, a significant decrease in
ROS levels was observed in the doxycycline-treated cells (Fig. 4).
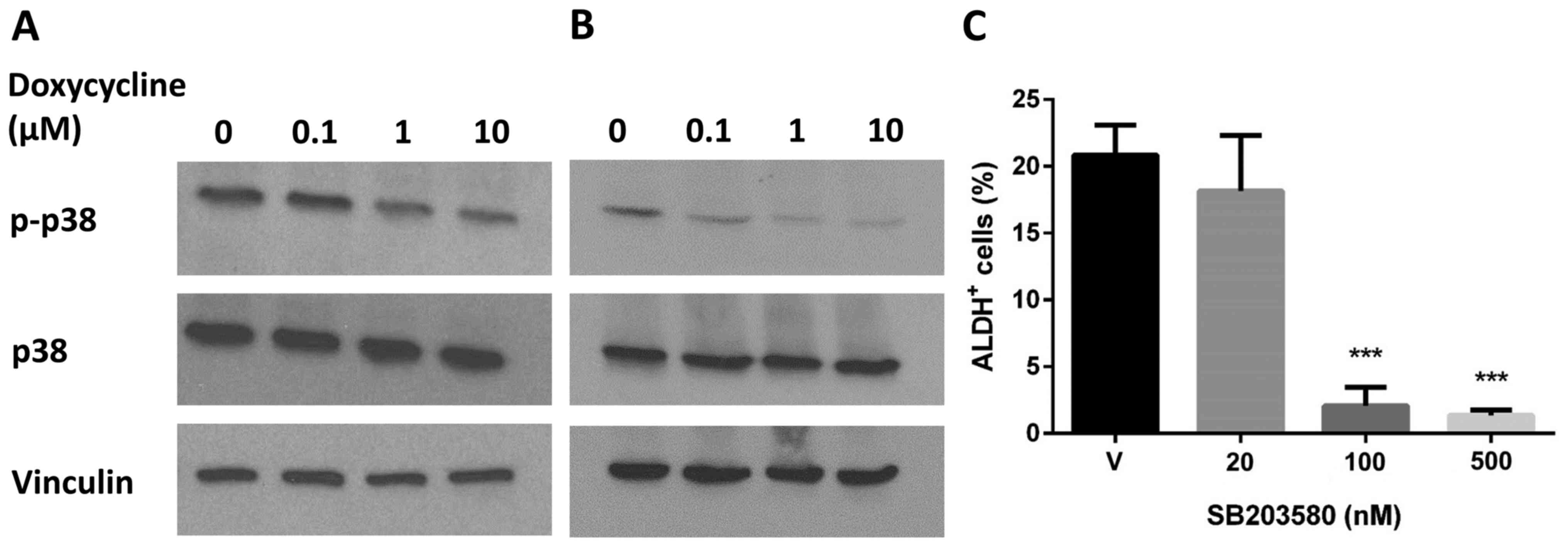
Next, we examined whether the p38 MAPK signaling
downstream of ROS was affected by doxycycline. We found that
doxycycline treatment resulted in a decrease in p38 phosphorylation
in a dose-dependent manner in the BT474 and SUM159 cell lines
(Fig. 5A and B). To further test
the correlation between p38 MAPK signaling and ALDH+
BCSCs, we treated BT474 cells with a p38 MAPK-specific inhibitor
SB203580 and then performed the Aldefluor assay. The result showed
that SB203580 abolished ALDH activity (Fig. 5C), indicating that p38 MAPK plays a
key role in ALDH+ BCSC maintenance, which is targeted by
doxycycline treatment.
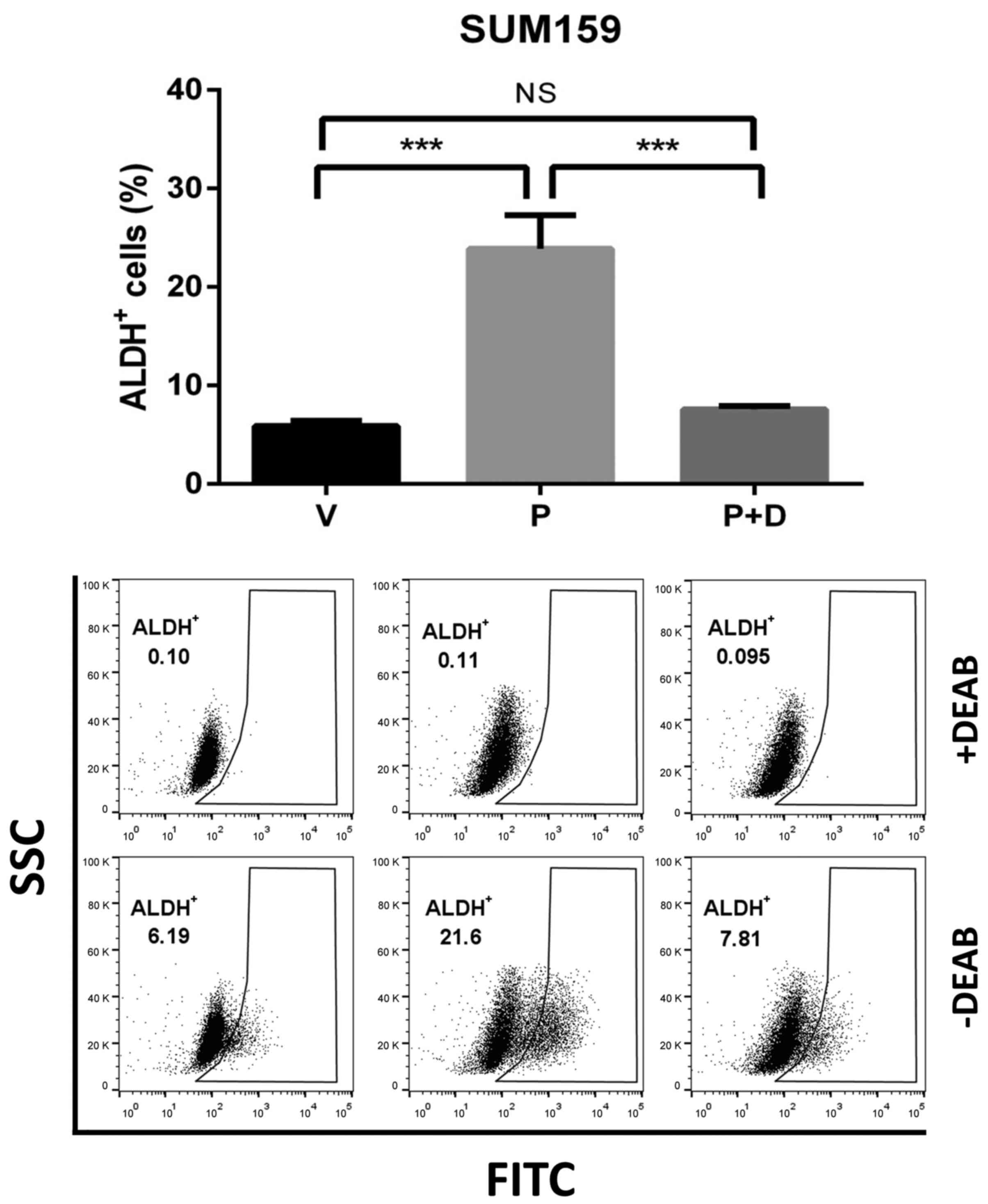
Doxycycline attenuates
paclitaxel-induced enrichment of ALDH+ BCSCs
Paclitaxel has been reported to kill the bulk of
tumor cells, yet enriching ALDH+ CSCs via elevating the
ROS level (35). To ascertain
whether doxycycline could ameliorate paclitaxel-induced enrichment
of ALDH+ BCSCs, SUM159 cells were pre-treated with
doxycycline and then in combination with paclitaxel. In agreement
with the previous report, paclitaxel treatment resulted in
approximately 4 times more ALDH+ BCSCs as compared to
the vehicle control. However, this enrichment of ALDH+
BCSCs induced by paclitaxel was significantly inhibited when cells
were pre-treated and later co-treated with doxycycline (Fig. 6).
Discussion
Recent studies have demonstrated that metastasis and
drug resistance of cancer are driven by small subpopulations of
cells termed cancer stem cells (CSCs). CSCs are therefore emerging
as important therapeutic targets for cancer treatment. In contrast
to conventional cytotoxic chemotherapy which aims to kill the bulk
of the tumor, CSC targeting therapy focuses on blocking specific
signaling pathways which CSCs rely on. Thus, combining chemotherapy
and CSC targeting therapy could help reach the goal of eradicating
the entire tumor. In the present study, we found that doxycycline
significantly decreased ALDH+ BCSCs by inhibiting MAPK
signaling, the downstream pathway of ROS. While applied in
combination with paclitaxel, doxycycline also attenuated
paclitaxel-induced enrichment of ALDH+ BCSCs, implying
the potentiality of combining the two drugs for removing both the
bulk of cancer cells and CSCs.
High mitochondrial mass is associated with the
ALDH+ CSC population (20). Since doxycycline has been shown to
interrupt mitochondrial biogenesis in eukaryotic systems (24), we hypothesized that doxycycline can
be used as an inhibitor for ALDH+ CSCs. The hypothesis
is supported by our results of aldefluor and mammosphere formation
assays. However, we also found that doxycycline failed to decrease
the CD44+/CD24− BCSC population (Fig. 2). CD44+/CD24−
are cell-surface markers acquired by epithelial cancer cells when
they undergo epithelial-to-mesenchymal transition (EMT), a
developmental program that enriches CSCs (36). CD44+/CD24− EMT
CSCs have characteristics that are distinct from those of
ALDH+ CSCs. Unlike proliferative and epithelial-like
ALDH+ CSCs, CD44+/CD24− EMT CSCs
are quiescent and mesenchymal-like (29,37).
Recent studies have reported that doxycycline can inhibit the
propagation of mitochondrial-related hypoxic CSCs (27), whereas doxycycline-resistance may
occur when cancer cells switch to a purely glycolytic phenotype
(28). The relationship between
CD44+/CD24− EMT CSCs and the glycolytic
phenotype is yet to be determined. Nonetheless, it is likely that
only mitochondrial-driven ALDH+ CSCs but not
CD44+/CD24− EMT CSCs are sensitive to
doxycycline.
Mitochondria are an important source of ROS
generation in most mammalian cells (38). ROS play an important role in
stabilizing hypoxia-induced factor 1α (HIF-1α), which is known to
induce ALDH+ CSCs (35,39,40).
Studies have shown that the p38 MAPK pathway, a downstream pathway
of ROS, is required for HIF-1α signaling (41,42).
Knockdown of p38 MAPK in the HER2-overexpressing MCF-7 cell line
can inhibit ALDH+ CSCs, cancer cell migration and
invasion (43,44). In the present study, we demonstrated
that doxycycline significantly decreased intracellular ROS levels,
p38 MAPK phosphorylation and ALDH+ CSCs. Cancer cells
treated with a p38 MAPK-specific inhibitor also exhibited a
significant reduction in ALDH+ CSCs, indicating that
doxycycline inhibited ALDH+ CSCs potentially via
blocking the p38 MAPK signaling pathway. However, more evidence is
needed to further support this hypothesis. Future studies will
focus on directly investigating the involvement of p38 MAPK in
doxycycline-mediated inhibition of ALDH+ CSCs. First,
knockdown of p38 MAPK could be carried out in HER2+ and
TNBC cell lines to ascertain whether ALDH+ CSC
population is affected. Second, a constitutively active p38 MAPK
could be overexpressed to examine its ability to prevent or
decrease doxycycline's effect on ALDH+ CSCs.
It is worth noting that doxycycline failed to
inhibit ALDH+ CSC population and secondary mammosphere
formation in SUM149 cells (Figs. 1C
and 3B). The number of mammospheres
formed is mainly determined by the number of stem cells seeded in
the culture. The results, however, can be affected if the treatment
changes the proliferation of cells. Therefore, to evaluate whether
doxycycline can really affect CSCs, we performed the secondary
mammosphere formation assays in the absence of doxycycline. Hence,
the effect of doxycycline on proliferation was avoided, and the
mammospheres should be decreased if the number of CSCs has been
reduced by doxycycline in the primary assays. In SUM149 cells, we
found that doxycycline significantly decreased primary but not
secondary mammosphere formation. The reason may be that doxycycline
inhibits cell proliferation (data not shown) instead of decreasing
CSCs in SUM149 cells. In addition, SUM149 has been reported as an
inflammatory breast cancer cell line that constitutively adapts to
hypoxia (45,46). Therefore, SUM149 can behave as if it
is continuously hypoxic even under normoxia (45). This may explain why doxycycline
decreases the ROS level but fails to inhibit ALDH+ BCSCs
in SUM149 cells.
Recent studies and our results indicate the
potentiality of repurposing doxycycline, an old drug as a new
treatment to target CSCs. Doxycycline is an FDA-approved antibiotic
since 1960s. With limited toxicity to cells, doxycycline is
relatively safe to be used concomitantly with chemotherapy drugs in
patients (25). A recent clinical
trial demonstrated that pathogenic bacteria-negative patients with
lymphoma still benefit from doxycycline (47). More phase II clinical trials are
ongoing to test the use of doxycycline as a CSC-targeting agent. In
addition to targeting CSCs, doxycycline was also found to
ameliorate tumor metastasis via inhibition of matrix
metallopeptidases (48,49). As such, we propose that doxycycline
is an ideal drug that can be used in combination with cytotoxic
chemotherapy drugs to eradicate both CSCs and bulk tumor cells.
Acknowledgements
The authors thank the Flow Cytometry Core of
University of Michigan, supported by the NCI Grant P30CA046592 from
the National Institutes of Health, for the technical support on
flow cytometry work in this study.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
CCL conceived and designed the study and wrote the
paper. CCL, RRM, NOS and SLT performed the experiments. MCL
reviewed and edited the manuscript. MCL and DS supervised the
research and manuscript preparation. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guiu S, Michiels S, André F, Cortes J,
Denkert C, Di Leo A, Hennessy BT, Sorlie T, Sotiriou C, Turner N,
et al: Molecular subclasses of breast cancer: How do we define
them? The IMPAKT 2012 working group statement. Ann Oncol.
23:2997–3006. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parker JS, Mullins M, Cheang MC, Leung S,
Voduc D, Vickery T, Davies S, Fauron C, He X, Hu Z, et al:
Supervised risk predictor of breast cancer based on intrinsic
subtypes. J Clin Oncol. 27:1160–1167. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lan KH, Lu CH and Yu D: Mechanisms of
trastuzumab resistance and their clinical implications. Ann N Y
Acad Sci. 1059:70–75. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nahta R, Yu D, Hung MC, Hortobagyi GN and
Esteva FJ: Mechanisms of disease: Understanding resistance to
HER2-targeted therapy in human breast cancer. Nat Clin Pract Oncol.
3:269–280. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu KD, Zhu R, Zhan M, Rodriguez AA, Yang
W, Wong S, Makris A, Lehmann BD, Chen X, Mayer I, et al:
Identification of prognosis-relevant subgroups in patients with
chemoresistant triple-negative breast cancer. Clin Cancer Res.
19:2723–2733. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lapidot T, Sirard C, Vormoor J, Murdoch B,
Hoang T, Caceres-Cortes J, Minden M, Paterson B, Caligiuri MA and
Dick JE: A cell initiating human acute myeloid leukaemia after
transplantation into SCID mice. Nature. 367:645–648. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Korkaya H, Kim GI, Davis A, Malik F, Henry
NL, Ithimakin S, Quraishi AA, Tawakkol N, D'Angelo R, Paulson AK,
et al: Activation of an IL6 inflammatory loop mediates trastuzumab
resistance in HER2+ breast cancer by expanding the cancer stem cell
population. Mol Cell. 47:570–584. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou BB, Zhang H, Damelin M, Geles KG,
Grindley JC and Dirks PB: Tumour-initiating cells: Challenges and
opportunities for anticancer drug discovery. Nat Rev Drug Dis.
8:806–823. 2009. View
Article : Google Scholar
|
|
12
|
Wicha MS: Targeting self-renewal, an
Achilles' heel of cancer stem cells. Nat Med. 20:14–15. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Warburg O: The Metabolism of Tumor.
Richard R. Smith; New York: 1931
|
|
15
|
Magda D, Lecane P, Prescott J, Thiemann P,
Ma X, Dranchak PK, Toleno DM, Ramaswamy K, Siegmund KD and Hacia
JG: mtDNA depletion confers specific gene expression profiles in
human cells grown in culture and in xenograft. BMC Genomics.
9:5212008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morais R, Zinkewich-Péotti K, Parent M,
Wang H, Babai F and Zollinger M: Tumor-forming ability in athymic
nude mice of human cell lines devoid of mitochondrial DNA. Cancer
Res. 54:3889–3896. 1994.PubMed/NCBI
|
|
17
|
Cavalli LR, Varella-Garcia M and Liang BC:
Diminished tumorigenic phenotype after depletion of mitochondrial
DNA. Cell Growth Differ. 8:1189–1198. 1997.PubMed/NCBI
|
|
18
|
Weinberg F, Hamanaka R, Wheaton WW,
Weinberg S, Joseph J, Lopez M, Kalyanaraman B, Mutlu GM, Budinger
GR and Chandel NS: Mitochondrial metabolism and ROS generation are
essential for Kras-mediated tumorigenicity. Proc Natl Acad Sci USA.
107:8788–8793. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tan AS, Baty JW, Dong LF, Bezawork-Geleta
A, Endaya B, Goodwin J, Bajzikova M, Kovarova J, Peterka M, Yan B,
et al: Mitochondrial genome acquisition restores respiratory
function and tumorigenic potential of cancer cells without
mitochondrial DNA. Cell Metab. 21:81–94. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Farnie G, Sotgia F and Lisanti MP: High
mitochondrial mass identifies a sub-population of stem-like cancer
cells that are chemo-resistant. Oncotarget. 6:30472–30486. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yan B, Stantic M, Zobalova R,
Bezawork-Geleta A, Stapelberg M, Stursa J, Prokopova K, Dong L and
Neuzil J: Mitochondrially targeted vitamin E succinate efficiently
kills breast tumour-initiating cells in a complex II-dependent
manner. BMC Cancer. 15:4012015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pasdar EA, Smits M, Stapelberg M,
Bajzikova M, Stantic M, Goodwin J, Yan B, Stursa J, Kovarova J,
Sachaphibulkij K, et al: Characterisation of
mesothelioma-initiating cells and their susceptibility to
anti-cancer agents. PLoS One. 10:e01195492015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
LeBleu VS, O'Connell JT, Herrera Gonzalez
KN, Wikman H, Pantel K, Haigis MC, de Carvalho FM, Damascena A,
Chinen Domingos LT, Rocha RM, et al: PGC-1α mediates mitochondrial
biogenesis and oxidative phosphorylation in cancer cells to promote
metastasis. Nat Cell Biol. 16(992–1003): 1–15. 2014.
|
|
24
|
Moullan N, Mouchiroud L, Wang X, Ryu D,
Williams EG, Mottis A, Jovaisaite V, Frochaux MV, Quiros PM,
Deplancke B, et al: Tetracyclines disturb mitochondrial function
across eukaryotic models: A call for caution in biomedical
research. Cell Rep: S2211-1247(15)00180-1. 2015. View Article : Google Scholar
|
|
25
|
Lamb R, Ozsvari B, Lisanti CL, Tanowitz
HB, Howell A, Martinez-Outschoorn UE, Sotgia F and Lisanti MP:
Antibiotics that target mitochondria effectively eradicate cancer
stem cells, across multiple tumor types: Treating cancer like an
infectious disease. Oncotarget. 6:4569–4584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lamb R, Fiorillo M, Chadwick A, Ozsvari B,
Reeves KJ, Smith DL, Clarke RB, Howell SJ, Cappello AR,
Martinez-Outschoorn UE, et al: Doxycycline down-regulates DNA-PK
and radiosensitizes tumor initiating cells: Implications for more
effective radiation therapy. Oncotarget. 6:14005–14025. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
De Francesco EM, Maggiolini M, Tanowitz
HB, Sotgia F and Lisanti MP: Targeting hypoxic cancer stem cells
(CSCs) with Doxycycline: Implications for optimizing
anti-angiogenic therapy. Oncotarget. 8:56126–56142. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
De Francesco EM, Bonuccelli G, Maggiolini
M, Sotgia F and Lisanti MP: Vitamin C and Doxycycline: A synthetic
lethal combination therapy targeting metabolic flexibility in
cancer stem cells (CSCs). Oncotarget. 8:67269–67286. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu S, Cong Y, Wang D, Sun Y, Deng L, Liu
Y, Martin-Trevino R, Shang L, McDermott SP, Landis MD, et al:
Breast cancer stem cells transition between epithelial and
mesenchymal states reflective of their normal counterparts. Stem
Cell Reports. 2:78–91. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brooks MD, Burness ML and Wicha MS:
Therapeutic implications of cellular heterogeneity and plasticity
in breast cancer. Cell Stem Cell. 17:260–271. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Burnett JP, Korkaya H, Ouzounova MD, Jiang
H, Conley SJ, Newman BW, Sun L, Connarn JN, Chen CS, Zhang N, et
al: Trastuzumab resistance induces EMT to transform
HER2+ PTEN− to a triple negative breast
cancer that requires unique treatment options. Sci Rep.
5:158212015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dontu G, Abdallah WM, Foley JM, Jackson
KW, Clarke MF, Kawamura MJ and Wicha MS: In vitro propagation and
transcriptional profiling of human mammary stem/progenitor cells.
Genes Dev. 17:1253–1270. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu X, Chai S, Wang P, Zhang C, Yang Y,
Yang Y and Wang K: Aldehyde dehydrogenases and cancer stem cells.
Cancer Lett. 369:50–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ahler E, Sullivan WJ, Cass A, Braas D,
York AG, Bensinger SJ, Graeber TG and Christofk HR: Doxycycline
alters metabolism and proliferation of human cell lines. PLoS One.
8:e645612013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Samanta D, Gilkes DM, Chaturvedi P, Xiang
L and Semenza GL: Hypoxia-inducible factors are required for
chemotherapy resistance of breast cancer stem cells. Proc Natl Acad
Sci USA. 111:E5429–E5438. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu S, Clouthier SG and Wicha MS: Role of
microRNAs in the regulation of breast cancer stem cells. J Mammary
Gland Biol Neoplasia. 17:15–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Murphy MP: How mitochondria produce
reactive oxygen species. Biochem J. 417:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Iriondo O, Rábano M, Domenici G,
Carlevaris O, López-Ruiz JA, Zabalza I, Berra E and Vivanco M:
Distinct breast cancer stem/progenitor cell populations require
either HIF1α or loss of PHD3 to expand under hypoxic conditions.
Oncotarget. 6:31721–31739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Conley SJ, Gheordunescu E, Kakarala P,
Newman B, Korkaya H, Heath AN, Clouthier SG and Wicha MS:
Antiangiogenic agents increase breast cancer stem cells via the
generation of tumor hypoxia. Proc Natl Acad Sci USA. 109:2784–2789.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gao N, Jiang BH, Leonard SS, Corum L,
Zhang Z, Roberts JR, Antonini J, Zheng JZ, Flynn DC, Castranova V
and Shi X: p38 signaling-mediated hypoxia-inducible factor 1alpha
and vascular endothelial growth factor induction by Cr(VI) in DU145
human prostate carcinoma cells. J Biol Chem. 277:45041–45048. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kwon SJ, Song JJ and Lee YJ: Signal
pathway of hypoxia-inducible factor-1alpha phosphorylation and its
interaction with von Hippel-Lindau tumor suppressor protein during
ischemia in MiaPaCa-2 pancreatic cancer cells. Clin Cancer Res.
11:7607–7613. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu M, Ren Z, Wang X, Comer A, Frank JA, Ke
ZJ, Huang Y, Zhang Z, Shi X, Wang S and Luo J: ErbB2 and p38gamma
MAPK mediate alcohol-induced increase in breast cancer stem cells
and metastasis. Mol Cancer. 15:522016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu M, Wang S, Ren Z, Frank JA, Yang XH,
Zhang Z, Ke ZJ, Shi X and Luo J: Chronic ethanol exposure enhances
the aggressiveness of breast cancer: The role of p38γ. Oncotarget.
7:3489–3505. 2016.PubMed/NCBI
|
|
45
|
Silvera D and Schneider RJ: Inflammatory
breast cancer cells are constitutively adapted to hypoxia. Cell
Cycle. 8:3091–3096. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wynn ML, Yates JA, Evans CR, Van
Wassenhove LD, Wu ZF, Bridges S, Bao L, Fournier C, Ashrafzadeh S,
Merrins MJ, et al: RhoC GTPase is a potent regulator of glutamine
metabolism and N-acetylaspartate production in inflammatory breast
cancer cells. J Biol Chem. 291:13715–13729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ferreri AJ, Ponzoni M, Guidoboni M, Resti
AG, Politi LS, Cortelazzo S, Demeter J, Zallio F, Palmas A, Muti G,
et al: Bacteria-eradicating therapy with doxycycline in ocular
adnexal MALT lymphoma: A multicenter prospective trial. J Natl
Cancer Inst. 98:1375–1382. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Shen LC, Chen YK, Lin LM and Shaw SY:
Anti-invasion and anti-tumor growth effect of doxycycline treatment
for human oral squamous-cell carcinoma-in vitro and in vivo
studies. Oral Oncol. 46:178–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Duivenvoorden WC, Popović SV, Lhoták S,
Seidlitz E, Hirte HW, Tozer RG and Singh G: Doxycycline decreases
tumor burden in a bone metastasis model of human breast cancer.
Cancer Res. 62:1588–1591. 2002.PubMed/NCBI
|