Introduction
Gastric cancer is the fourth most common cancer and
the second leading cause of cancer-related deaths worldwide
(1). An estimated 951,600 new
gastric cancer cases and 723,100 gastric cancer-related deaths
occurred worldwide in 2012 (2). The
patients are usually diagnosed at an advanced stage with metastases
and more than half of radically resected gastric cancer patients
relapse, either locally or with distant metastases. Therefore, the
5-year survival is less than 10% and the prognosis of patients
remains poor (1).
Diallyl disulfide (DADS), one of the sulfur
compounds derived from garlic, exhibits anticancer activity by
modulating signaling molecules in various pathways, indicating that
DADS could be used as a potential therapeutic agent for the
treatment or prevention of cancer (3).
Transforming growth factor-β (TGF-β) plays a pivotal
role in cancer progression and metastasis by inducing
epithelial-mesenchymal transition (EMT), in which cancer cells
acquire the capability of motility and invasion (4). TGF-β induces EMT not only through the
Smad-mediated gene expression regulation, but also by activating
non-Smad signaling, such as PI3K/Akt, ERK, JNK, p38, Src tyrosine
kinase and Rho GTPases pathways (4,5).
TGF-β1, one of the members of the transforming
growth factor family, induces EMT via the downregulation of the
expression of E-cadherin and the upregulation of the expression of
vimentin in gastric cancer cells (6). Crosstalk between the TGF-β1/Smad and
other pathways is critical during the development of TGF-β1-induced
EMT. Wnt/β-catenin pathway has been demonstrated to mediate EMT.
Apart from Wnt-dependent β-catenin transactivation, TGF-β1
regulates β-catenin nuclear translocation through a Smad-dependent
manner (7). In addition,
TGF-β1-induced EMT is mediated by ERK-dependent β-catenin
upregulation and nuclear translocation in renal tubular epithelial
cells (7). The β-catenin inhibitor
can reverse TGF-β1-induced EMT in human airway epithelial cells
(8). In addition, DADS inhibits the
activation of the β-catenin pathway and EMT in breast cancer cells
(9).
Activation of Rac1 GTPase/Pak1 pathway is involved
in TGF-β1-induced EMT in prostate cancer cells (10). We have demonstrated that DADS
restrained EMT, migration and invasion through the downregulation
of Rac1-Pak1/Rock1-LIMK1 (11) and
uPAR-ERK-Fra-1 (12) pathways in
gastric cancer cells and inhibited the Wnt-1/β-catenin pathway
through the upregulation of miR-200b and miR-22 in gastric cancer
cells (13).
We proposed that DADS has inhibitory effects on
TGF-β1-induced EMT and invasion, which may be associated with the
downregulation of Rac1 and β-catenin in gastric cancer cells.
In the present study, we verified that TGF-β1
upregulated Rac1 and β-catenin in gastric cancer cells and that
Rac1 regulated the expression of β-catenin. DADS treatment
inhibited the expression of TGF-β1, resulting in the downregulation
of Rac1 and β-catenin. The underlying mechanisms of DADS
suppressive effects on TGF-β1-induced EMT, invasion and growth of
gastric cancer were investigated.
Materials and methods
Cell culture and cell line
establishment
Human gastric cancer MGC803 cell line was obtained
from the Cancer Research Institute, Xiangya Medical College,
Central South University in China. Cells were cultured in RPMI-1640
medium (Gibco; Life Technologies, Vienna, Austria) containing 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.,
Vienna, Austria) with the addition of 100 U/ml penicillin, 100 U/ml
streptomycin and maintained at 37°C in a humidified atmosphere of
containing 5% CO2. To establish TGF-β1-overexpressing
cell lines, the plasmid pCMV6 containing the full-length cDNA of
human TGF-β1 (pCMV6-TGF-β1) was constructed by OriGene Technologies
(Rockville, MD, USA). MGC803 cells were transfected with
TGF-β1-expressing plasmid and the control vector (pCMV6-Neo) using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.,
Carlsbad, CA, USA) according to the manufacturer's protocol. The
stable transfectants were established after G418 (Invitrogen;
Thermo Fisher Scientific, Inc.) selection. The expression levels of
TGF-β1 in stable cell lines were evaluated by western blot
analysis.
Reagents and antibodies
Diallyl disulfide (DADS), purchased from Fluka Co.
(Milwaukee, WI, USA), was dissolved in Tween-80 and stored at −20°C
after a 100-fold dilution with saline. Human recombinant TGF-β1
protein was purchased from R&D Systems (Minneapolis, MN, USA).
The primary antibodies against TGF-β1 (cat. no. ab92486), Rac1
(cat. no. ab33186), β-catenin (cat. no. ab16051), Ki-67 (cat. no.
ab66155), CD34 (cat. no. ab81289) and FITC-conjugated anti-mouse
(cat. no. ab6785) or anti-rabbit (cat. no. ab6717) secondary
antibodies were provided by Abcam (Cambridge, MA, UK). The primary
antibodies against E-cadherin (cat. no. 24E10) and vimentin (cat.
no. D21H3) were obtained from Cell Signaling Technology (Danvers,
MA, USA). The mouse monoclonal against β-actin antibody (cat. no.
sc-8432) and horseradish peroxidase (HRP)-conjugated secondary
antibodies (cat. no. sc-2004 and cat. no. sc-2005) were purchased
from Santa Cruz Biotechnology (Santa Cruz, CA, USA). TGF-β1
receptor inhibitor SB431542 and Rac1 inhibitor NSC23766 were
obtained from Cayman Chemical (Ann Arbor, MI, USA). A total of 5
ng/ml TGF-β1, 30 mg/l DADS, 10 µmol/l SB431542 and 50 µmol/l
NSC23766 were used for experiments in vitro.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from the cells using TRIzol
reagent (Gibco-BRL; Thermo Fisher Scientific, Inc., Grand Island,
USA). Reverse transcription was carried out using the RT-PCR system
(Promega, Madison, WI, USA). PCR analysis was performed using the
GeneAmp PCR kit (Promega). Primer sequences were as follows: TGF-β1
forward, TCTCCAGGCATTTCCACTATTC and reverse,
CTCAGGCATTCGTCAACATCTA; Rac1 forward, TGCCTGCTGTTGTAAATGTCTC and
reverse, AAAGTTCAGTGCTCGGTGTTCT; β-catenin forward,
GGAAGGGACAGTATCGTTTGTT and reverse, GCCTCAGCATCTACCAGCATAG;
E-cadherin forward, CTCCCAATACATCTCCCTTCAC and reverse,
CGCCTCCTTCTTCATCATAGTAA; vimentin forward, GCGAGGAGAGCAGGATTTCT and
reverse, TCTTGTAGGAGTGTCGGTTGTT; β-actin forward,
CTGGGACGACATGGAGAAAA and reverse, AAGGAAGGATGGAAGAGTGC. The PCR
products were analyzed on 2% agarose gel containing ethidium
bromide. Densitometric quantitation of products was determined
using the Labwork analysis software (Labworks LLC, Lehi, UT, USA).
The relative abundance was expressed as the ratio of the object
gene to β-actin.
Western blot analysis
For total protein extraction, cells were lysed
directly on ice for 30 min in lysis buffer [10 mmol/l Tris-HCl (pH
7.6), 100 mmol/l NaCl, 1 mmol/l EDTA (pH 8.0), 100 µg/ml PMSF and 1
µg/ml aprotinin]. The cell lysates were centrifugated at 12,000 rpm
for 10 min and the supernatants were collected. Then protein
contents were determined using a BCA protein assay kit (Pierce,
Rockford, IL, USA).
Protein extracts were loaded on a 10%
SDS-polyacrylamide gel for electrophoresis and transferred onto
polyvinylidene fluoride (PVDF) membrane. The blots were blocked in
5% skim milk in Tris-buffered saline (TBS) containing 0.1% Tween-20
for 2 h at room temperature, and then incubated with primary
antibodies (1:200-500) at 4°C overnight. The membranes were washed
in TBS-T and then incubated with HRP-conjugated secondary
antibodies (1:1,000-1:2,000). After washed with TBS-T, the
membranes were developed by an enhanced chemiluminescence plus (ECL
Plus) kit (Amersham Biosciences, Buckinghamshire, UK) and bands
were visualized on X-ray film (Kodak, Rochester, NY, USA).
Membranes were re-incubated with anti-β-actin antibody to verify
equal protein-sample loading. The target protein amounts were
normalized towards β-actin quantity using densitometry, then
relative fold changes in protein levels were calculated as ratios
between treated vs. control group values.
Cell migration and invasion
assays
Invasion assays were performed using
Transwell® plates (Corning, Inc., Corning, NY, USA) as
previously described (12).
Briefly, MGC803 cells were seeded onto Matrigel-coated filters
(8-µm pore size), then were treated with TGF-β1 (5 ng/ml) or DADS
(30 mg/l) alone, or incubated with TGF-β1 + SB431542 (10 µmol/l) or
TGF-β1 + NSC23766 (50 µmol/l) for 24 h or left untreated. The cells
that had invaded the lower surface of the filter were fixed and
stained with hematoxylin. Invasiveness was determined by counting
cells in four microscopic fields per well, and the extent of
invasion was expressed as an average number of cells per
microscopic field. Invasion rates were expressed as the ratio of
the treated group value to the control group value. Transwell
migration assays were conducted using the same procedure as for the
invasion assay, except using the Matrigel-uncoated filters.
Gastric tumor growth in nude mice
Transfected and untransfected MGC803 cells were
injected into the subcutis of the right axillary of male athymic
BALB/c nude mice (4 weeks old). The mice were purchased from the
Experimental Center of the Chinese Academy of Science in Shanghai.
The mice were housed in an environment controlled for temperature
(22±2°C), light (12 h light/dark cycle) and humidity (60±10%). The
animals were maintained under specific pathogen-free conditions in
accordance with the NIH Guide for the Care and Use of Laboratory
Animals. The animals were randomly divided into six groups, and
each group consisted of five mice. The mice were treated with
normal saline, DADS (100 mg/kg) (11), SB431542 (10 mg/kg) (14) and NSC23766 (5 mg/kg) (15) via intraperitoneal injection every 2
days until the termination of the experiment. Tumor volume
(cm3) was examined every 6 days and calculated using a
standard formula (width2 × length × 0.5). Average tumor
volumes are presented (n=5 for each group) starting from the
twelfth day and continuing until mice were sacrificed at 48 days by
cervical dislocation under anesthesia. The xenografts were removed
and the tumor size and weight were assessed at 48 days. Tumor
tissues were then fixed in formalin and embedded in paraffin.
Tissue sections (5 µm-thick) were prepared for subsequent
immunohistochemistry analysis. All experiments were performed
according to the guidelines for animal use of the Ethics Committee
of the University of South China.
Immunohistochemistry
Briefly, after slides were dewaxed in xylene and
hydrated in graded alcohol solutions, antigen retrieval was
performed by heat treatment in 10 mM sodium citrate buffer (pH
8.0). Slides were incubated in 3% H2O2
solution to quench endogenous peroxidase activity and then
incubated with normal goat serum for 20 min. Slides were incubated
with primary antibodies (dilution 1:100) at 4°C overnight. Positive
signals were developed with peroxidase-conjugated secondary
antibodies and 0.5% diaminobenzidine/H2O2
followed by counterstaining with Mayer's hematoxylin, dehydration,
clearing and mounting. The slides that were treated with normal
goat serum were evaluated as negative controls.
Statistical analysis
All results are presented as the mean ± SD for three
independent experiments. Student's t-tests and one-way ANOVA were
used to analyze the differences in expression among groups.
P<0.05 were considered to indicate a statistically significant
result. Statistical analyses were conducted using the SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA).
Results
DADS downregulates TGF-β1, Rac1,
β-catenin and vimentin and upregulates E-cadherin in MGC803
cells
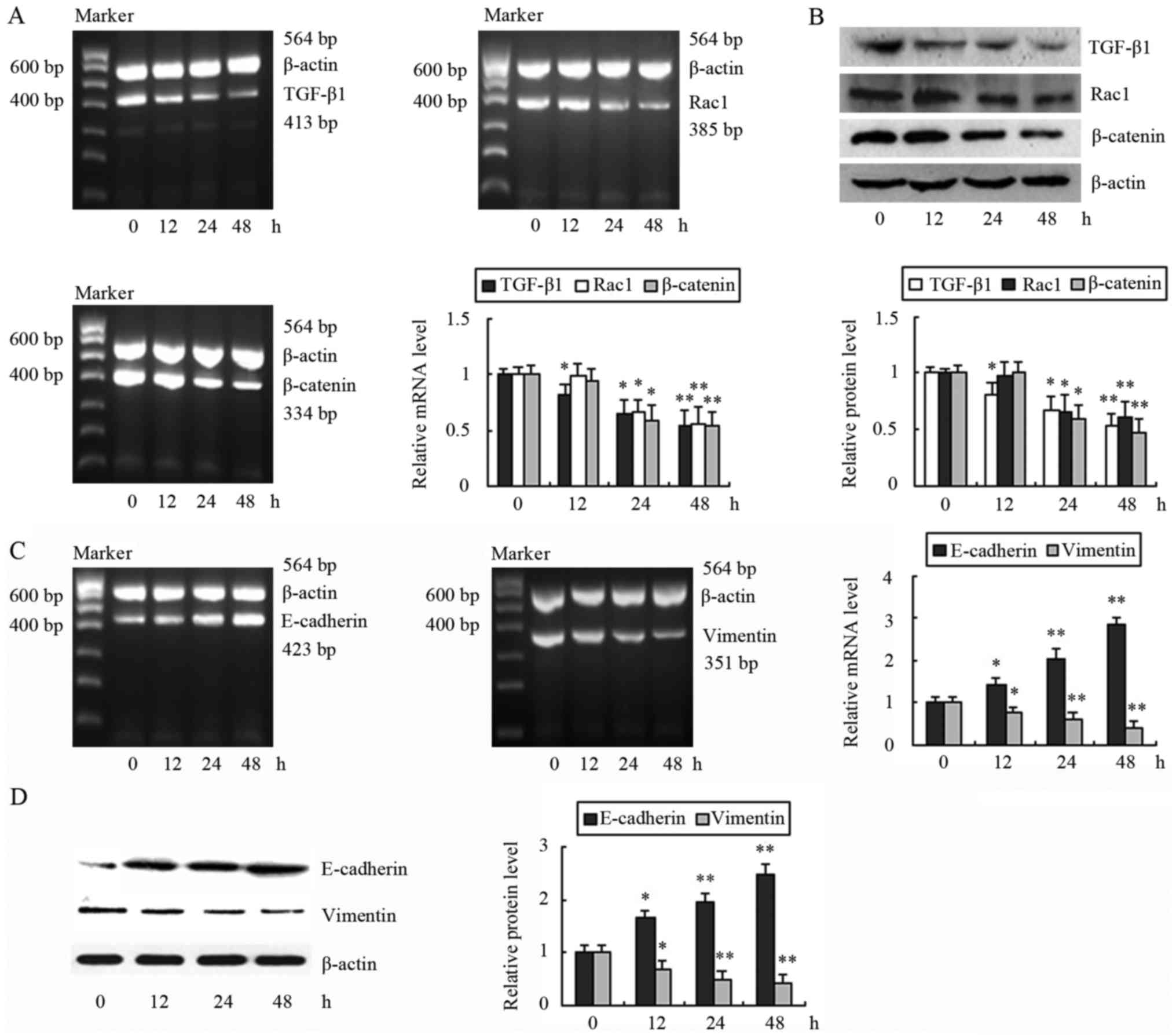
We first treated cells with 30 mg/l DADS for
different time-points and we examined the effects of DADS on the
expression of TGF-β1, Rac1 and β-catenin. The mRNA and protein
levels of TGF-β1 were reduced after cells were treated with DADS
for 12, 24 and 48 h in a time-dependent manner (Fig. 1A and B). In contrast, Rac1 and
β-catenin were decreased in the mRNA and protein levels after
incubation for 24 h (Fig. 1A and
B). These data indicated that DADS can reduce the expression of
TGF-β1, Rac1 and β-catenin. In addition, we observed that a
decrease in TGF-β1 occurred after 12 h of incubation, which was
earlier than the decreases in Rac1 and β-catenin (24 h). We
proposed that downregulation of TGF-β1 by DADS may result in the
expression changes of its downstream effectors, Rac1 and β-catenin.
The expression of E-cadherin in the mRNA and protein level was
increased by DADS after 12 h of treatment. In contrast, the mRNA
and protein levels of vimentin were reduced (Fig. 1C and D).
DADS antagonizes TGF-β1-induced
upregulation of TGF-β1, Rac1 and β-catenin
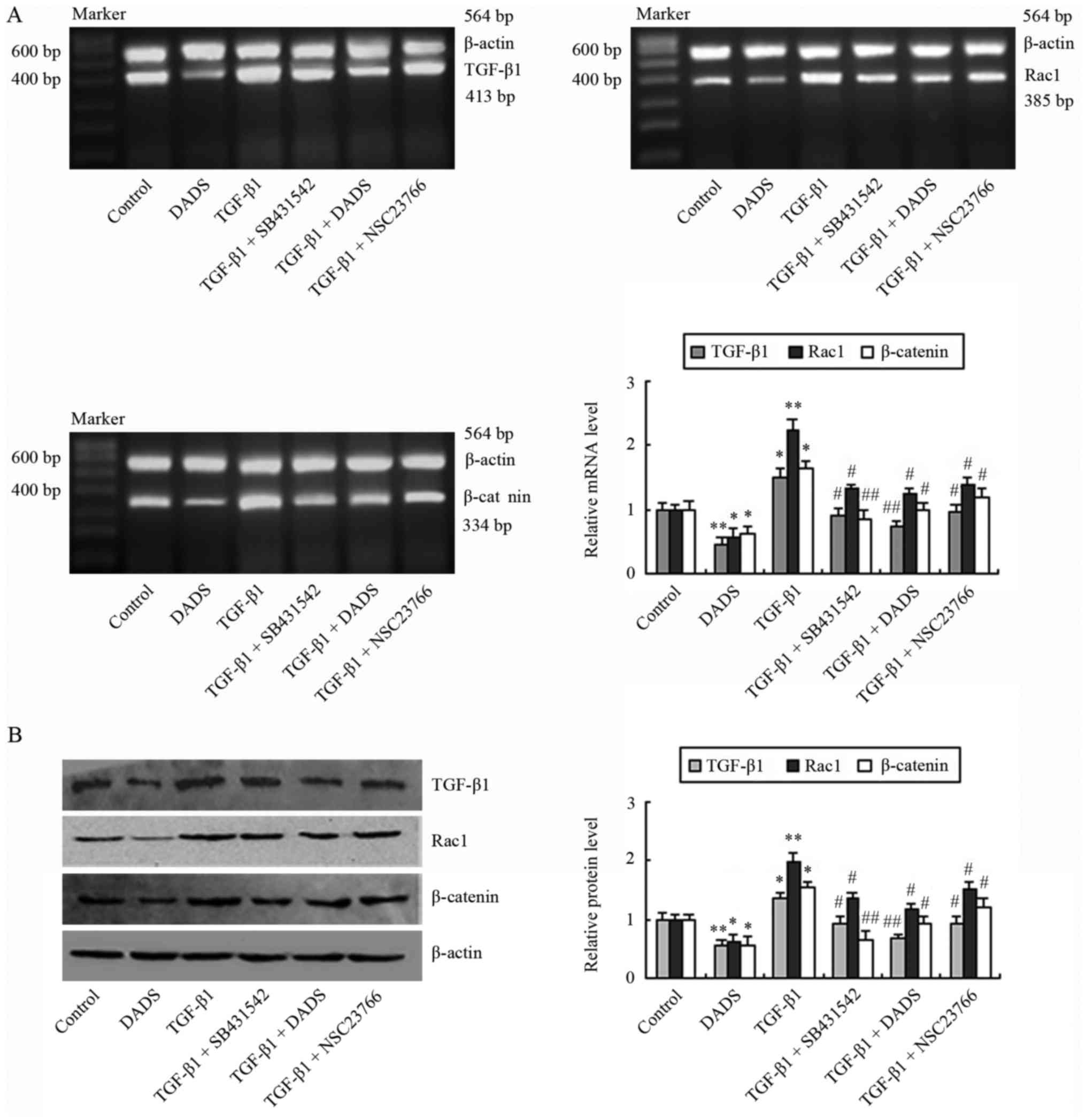
Subsequently, we explored the effects of TGF-β1 on
the expression of TGF-β1, Rac1 and β-catenin in MGC803 cells. The
expression levels were determined after the cells were incubated
with TGF-β1 (5 ng/ml) for 24 h. The mRNA and protein expression
levels of TGF-β1, Rac1 and β-catenin were elevated in cells exposed
to TGF-β1 (Fig. 2). The results
indicated that TGF-β1 can induce upregulation of TGF-β1, Rac1 and
β-catenin. Compared with the TGF-β1-treated group, TGF-β1, Rac1 and
β-catenin protein levels were decreased in the TGF-β1 + SB431542
group (Fig. 2B). Similarly, these
protein levels were decreased after cells were treated with TGF-β1
in the presence of a Rac1 inhibitor, NSC23766. DADS (30 mg/l)
treatment produced similar effects to those of SB431542 and
NSC23766 (Fig. 2).
DADS suppresses TGF-β1-induced EMT and
invasion by blocking TGF-β1 and Rac1
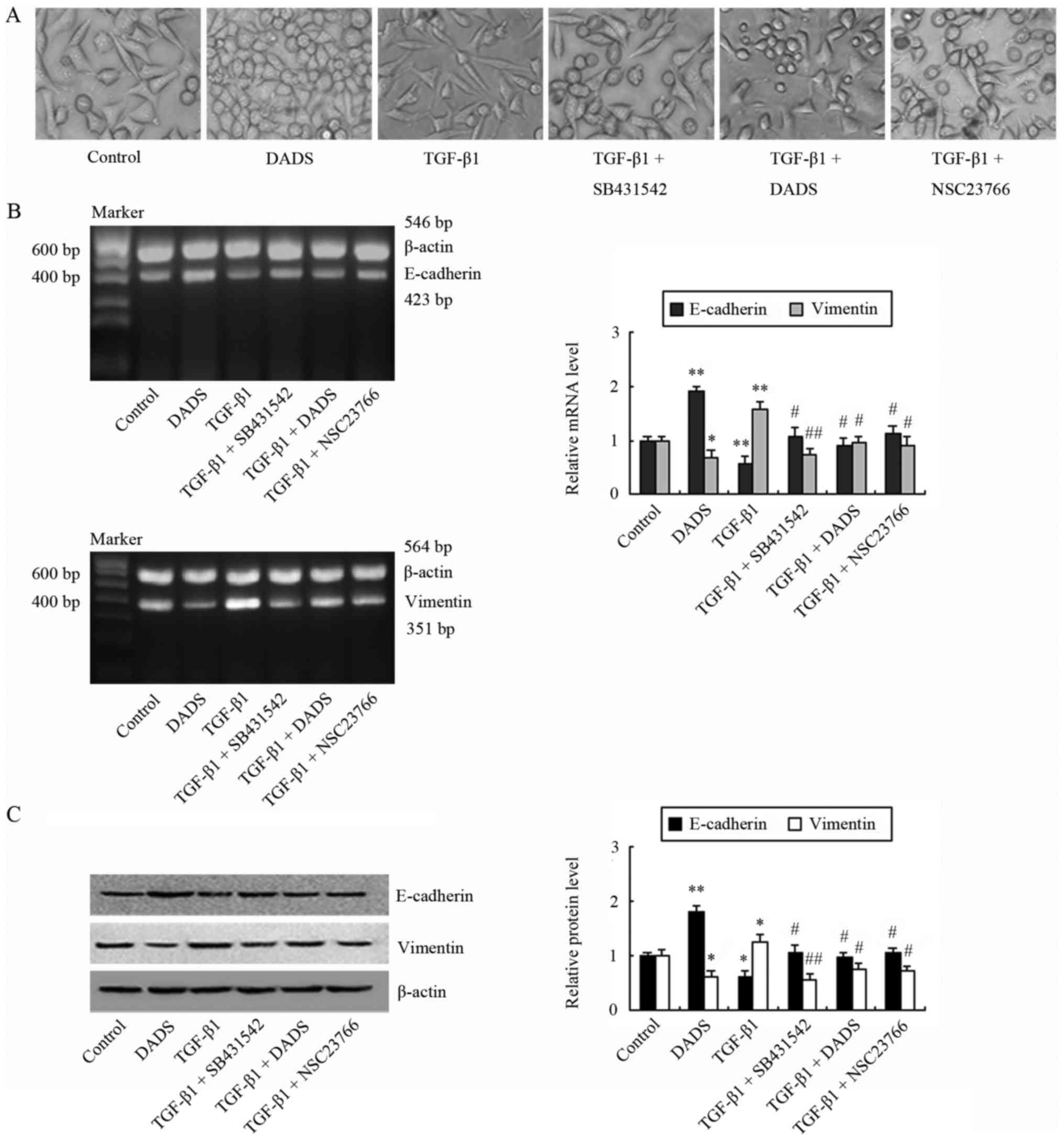
We observed that TGF-β1 treatment induced a
morphological change (spindle-like morphology) and a decrease in
cell-cell junctions, compared with the control group (Fig. 3A). In line with these morphological
changes, an increase of vimentin and a decrease of E-cadherin in
the mRNA and protein levels were demonstrated in the TGF-β1-treated
group (Fig. 3B and C). Conversely,
SB431542 and NSC23766 reversed these changes of morphology and EMT
markers, which occurred in TGF-β1-treated cells (Fig. 3B and C). DADS exerted similar
effects as SB431542 and NSC23766, decreasing vimentin and
increasing E-cadherin, concomitantly with significant inhibition of
morphological changes similar to mesenchymal cells (Fig. 3).
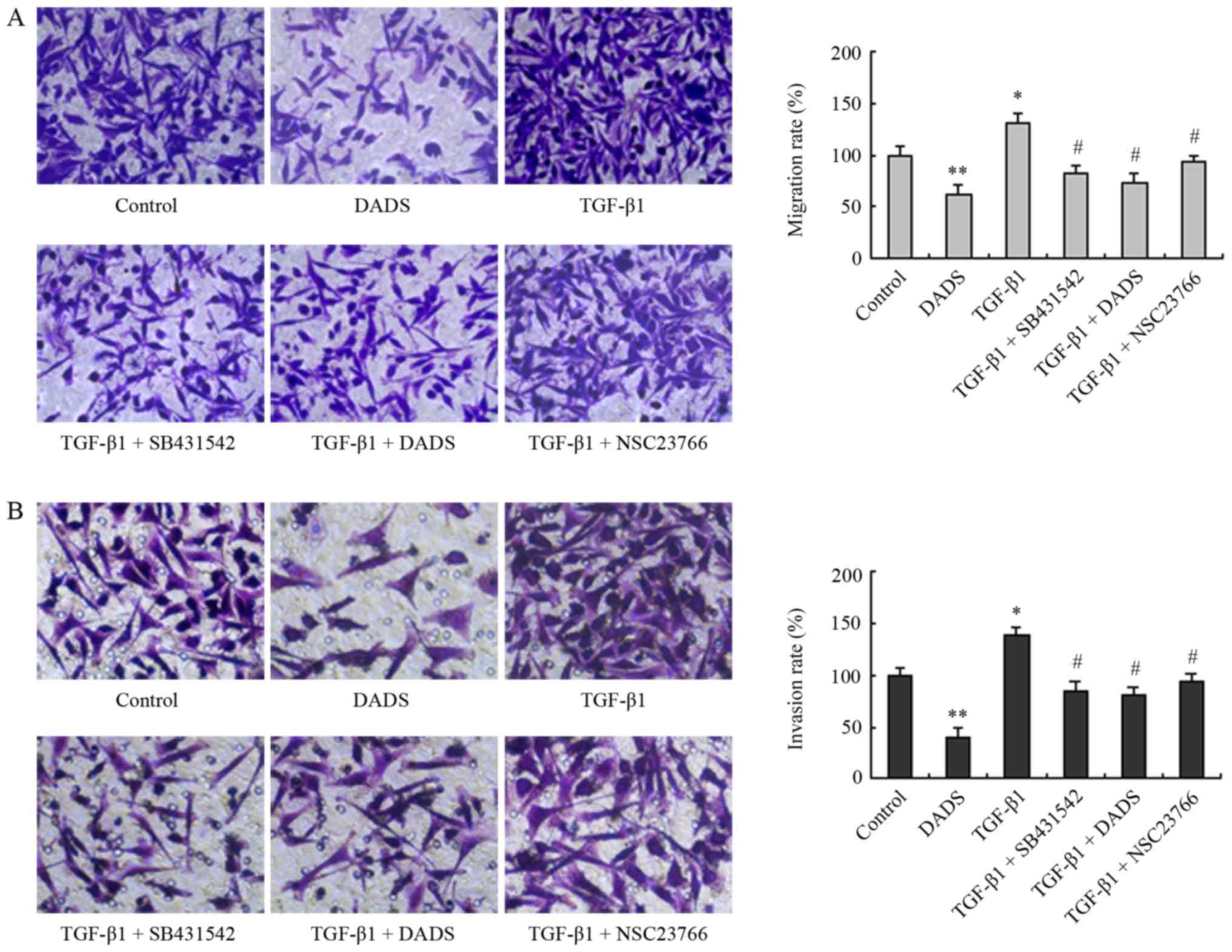
Subsequently, we further demonstrated that TGF-β1
treatment increased the rates of cell migration and invasion, while
DADS neutralized these effects of TGF-β1, as did SB431542 and
NSC23766 (Fig. 4). These data
indicated that Rac1 mediated EMT induced by TGF-β1, whereas
downregulation of TGF-β1/Rac1 signaling by DADS resulted in
inhibition of EMT, migration and invasion.
DADS, TGF-β1 receptor inhibitor
SB431542 and Rac1 inhibitor NSC23766 suppress TGF-β1-induced tumor
growth in vivo
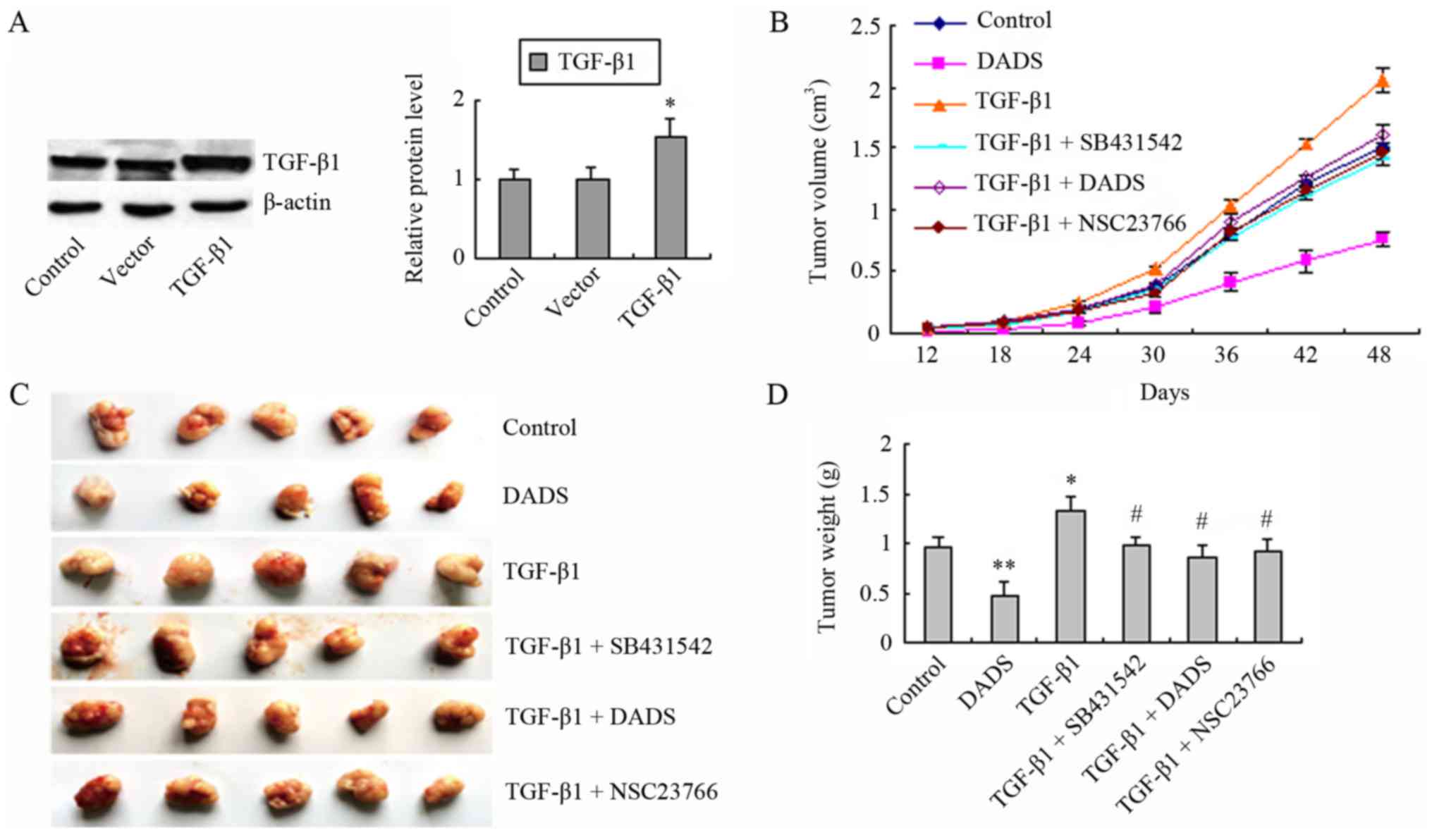
We have previously reported that DADS inhibits tumor
growth by downregulating LIMK1, a downstream effector of Rac1
(11). We constructed a
TGF-β1-overexpressing MGC803 cell line that exhibited increased
TGF-β1 expression compared to the empty vector group and the
control group (Fig. 5A). The
transfected and untransfected cells were subcutaneously injected
into nude mice. We examined the effect of TGF-β1 on tumor growth in
nude mice, and determined whether the suppression of TGF-β1/Rac1 by
DADS led to inhibition of gastric cancer MGC803 cell proliferation
in vivo. The mice were subjected to different treatments and
the tumor volume was examined every 6 days. Compared to the control
group, the TGF-β1 group demonstrated an increase in tumor volume,
whereas a decreased tumor volume was observed in the DADS group
(Fig. 5B). The TGF-β1 + DADS,
TGF-β1 + SB431542 and TGF-β1 + NSC23766 groups exhibited reduced
tumor volumes, compared to the TGF-β1 group (Fig. 5B). After 48 days, the xenografts
were removed from the mice. Similar changes were observed in tumor
volume and weight (Fig. 5C and D).
These data indicated that DADS antagonized TGF-β1-induced tumor
growth via the downregulation of TGF-β1/Rac1 signaling.
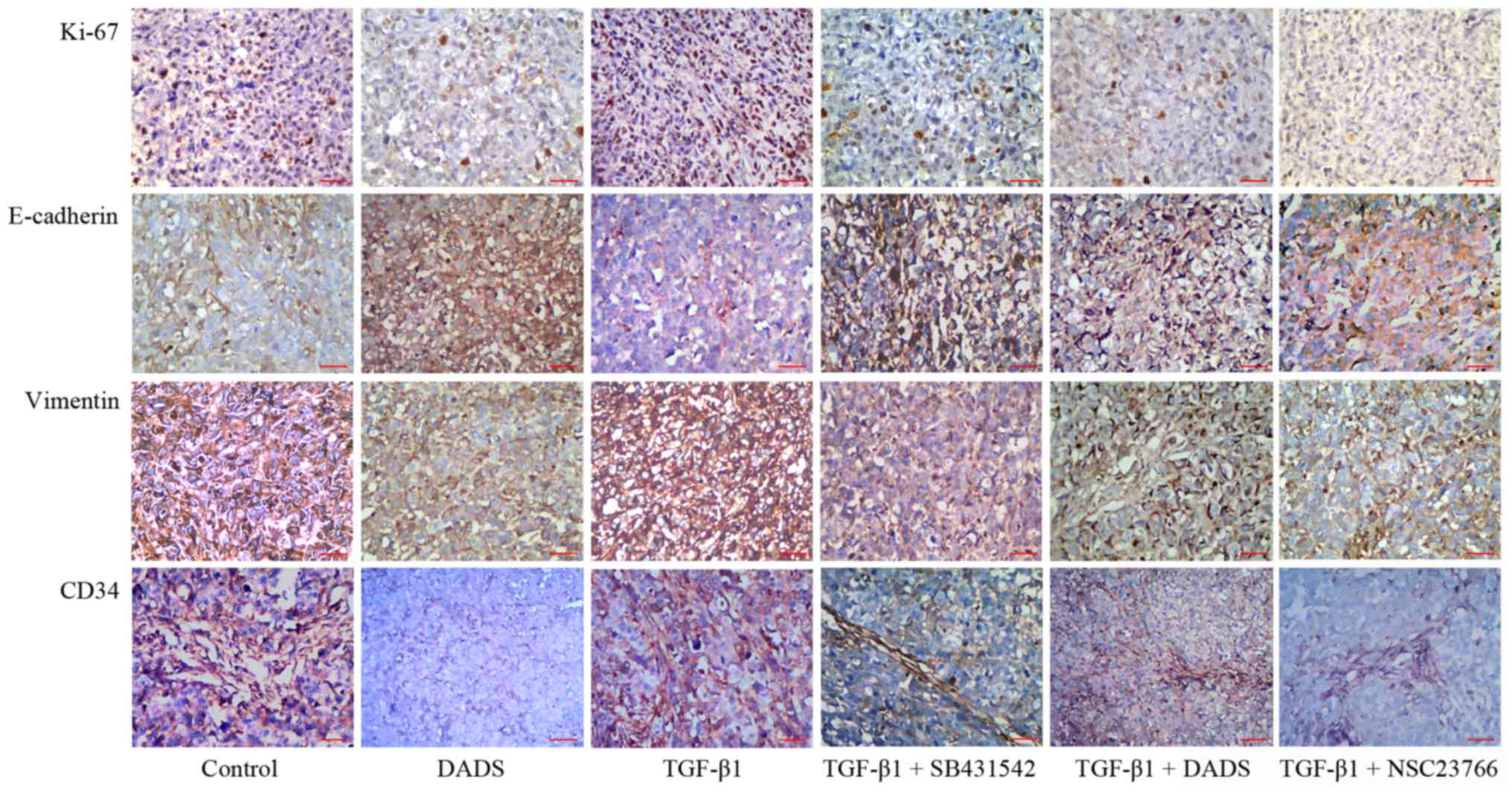
Effects of DADS on TGF-β1-induced
E-cadherin, vimentin, Ki-67 and CD34 expression in vivo
We detected the protein expression levels of
E-cadherin, vimentin, Ki-67 and CD34 in transplanted tumor tissues
using immunohistochemistry. DADS reduced vimentin, Ki-67 and CD34
protein levels, and increased the expression of E-cadherin
(Fig. 6). These results were
consistent with our previous data (11). The opposite effects were observed in
the TGF-β1 group. SB431542 and NSC23766 attenuated the inhibitory
effect of TGF-β1 on E-cadherin expression and weakened the enhanced
effects of TGF-β1 on vimentin, Ki-67 and CD34 expression.
Furthermore, DADS exerted the same effects on the expression of
these proteins as these inhibitors (Fig. 6).
Discussion
TGF-β1 downregulates the expression of E-cadherin
and upregulates the expression of vimentin, inducing EMT in gastric
cancer cells (6). SB431542 (a
TGF-β1 receptor inhibitor) blocks the TGF-β1 signaling pathway
(16,17) and reverses TGF-β1-induced EMT in
esophageal cancer cells, by downregulating the expression of
N-cadherin and vimentin and upregulating the expression of
E-cadherin (17). In the present
study, we demonstrated that DADS decreased the expression of TGF-β1
and exerted same effects as SB431542, which abolished the enhanced
effects of TGF-β1 on EMT and invasion in gastric cancer cells.
Therefore, DADS inhibited EMT through the downregulation of TGF-β1
in gastric cancer cells.
We observed that Rac1 and β-catenin expression
levels were increased in TGF-β1 treated gastric cancer cells which
indicated that TGF-β1 may positively regulate the expression of
Rac1 and β-catenin. DADS and SB431542 abrogated the TGF-β1 induced
upregulation of the expression of Rac1 and β-catenin which
indicated that downregulation of TGF-β1 by DADS resulted in reduced
Rac1 and β-catenin expression. Rac1 (10) and β-catenin (7) pathways are involved in TGF-β1-induced
EMT. DADS inactivates the β-catenin pathway and inhibits EMT in
breast cancer cells (9). TGF-β1
induces EMT in prostate cancer cells via the activation of the
Rac1/Pak1 pathway (10). DADS
downregulates the Rac1/LIMK1 pathway, inhibiting EMT in gastric
cancer cells (11). The decreased
expression of Rac1 and β-catenin may contribute to the suppression
of TGF-β1-induced EMT in gastric cancer cells.
We revealed that TGF-β1 promoted the expression of
TGF-β1, while DADS and the TGF-β1 receptor inhibitor antagonized
this effect. Furthermore, NSC23766, a Rac1 specific inhibitor,
decreased the expression of TGF-β1 in TGF-β1-treated cells,
indicating that Rac1 also positively regulated the expression of
TGF-β1. NSC23766 inactivates Rac1 and results in abolishing colon
cancer cell migration and invasion (18). Cigarette smoke extract (CSE)-induced
EMT in pulmonary epithelial cells is associated with elevated Rac1
expression and increased TGF-β1 release and Rac1 inhibition by
NSC23766 or knockdown decreases TGF-β1 release and abolishes
CSE-induced EMT (19). TGF-β1 can
induce the upregulated expression of Rac1 (20). Thus, a reciprocal positive interplay
in expression regulation may exist between Rac1 and TGF-β1, and
DADS may inhibit this positive feedback regulation mechanism.
There exists crosstalk between Rac1 and
Wnt/β-catenin pathways. Rac1 acts as an upstream regulator of
β-catenin. Overexpression of Rac1 augments Wnt3a-stimulated
transcription of β-catenin target genes (21). Rac1-mediated JNK2 activation by
Wnt3a promotes β-catenin phosphorylation and nuclear localization
in ST2 cells (22). Rac1/PAK1 is
required for the superactivation of β-catenin in colon cancer cells
(23), and Rac1 promotes the
formation of nuclear β-catenin-lymphoid enhancer factor 1 (LEF-1)
complexes (24). Even without Wnt
stimulation, Rac1 can still recruit β-catenin to its target genes
and act as a co-activator in β-catenin/TCF (T cell factor)-mediated
transcription in colon cancer cells (25). Furthermore, the interaction of
active or inactive Rac1 with β-catenin is required for the nuclear
translocation of β-catenin and Rac1 can promote β-catenin target
gene transcription in breast cancer cells (26). TGF-β1 induces Rac1 activation in
prostate cancer cells (10). Rac1
inhibitors attenuate Wnt/β-catenin pathway in breast cancer cells,
reducing cell migration and invasion (27). We hypothesized that the increased
Rac1 expression by TGF-β1 may facilitate the expression and
activation of β-catenin in gastric cancer cells, while the reduced
expression of Rac1 by DADS may reverse this effect of TGF-β1.
Rac1-mediated activation of β-catenin regulates the
expression of Snail and MMP9, whereas knockdown of Rac1 decreases
the expression and activation of β-catenin, resulting in impairing
trophoblast invasion (28), which
indicates that Rac1 may regulate the expression of β-catenin in
addition to activating β-catenin. TGF-β1-induced EMT is mediated by
ERK-dependent β-catenin upregulation and nuclear translocation in
renal tubular epithelial cells (7).
Rac1-mediated ERK activation is involved in TGF-β1-induced EMT in
keratinocytes (29). We previously
verified that DADS can downregulate Rac1/LIMK1 (11) and ERK (12) pathways in gastric cancer cells. We
revealed that DADS, SB431542 and NSC23766 prohibited TGF-β1-induced
upregulation of the expression of β-catenin. These data indicated
that DADS reduced the expression of β-catenin through
downregulation of the TGF-β1/Rac1 pathway in gastric cancer cells,
which may, in part, account for the inhibitory effects of DADS on
TGF-β1-induced EMT and invasion.
TGF-β1-mediated activation of Rac1/Pak1 pathway is
associated with prostate tumor xenograft growth (10). Downregulation of Rac1/Pak1/LIMK1
(11) and Wmt/β-catenin (13) pathways is associated with the growth
inhibition of gastric cancer cells in vitro and in
vivo. In in vivo experiments, we further verified that
DADS can reverse TGF-β1-induced EMT by blocking the TGF-β1/Rac1
pathway, which was supported by the upregulation of vimentin and
the downregulation of E-cadehrin. Ki-67 is widely used as a marker
to assess cell proliferation (30)
and the overexpression of Ki-67 is related to poor prognosis of
patients with gastric cancer (31).
CD34 is a specific angiogenic marker and its expression is
modulated by Pak1 (32). We have
demonstrated that the decreased expression of Ki-67 and CD34 due to
DADS-induced downregulation of LIMK1 is in accordance with the
tumor growth inhibition (11). We
illustrated that TGF-β1-induced tumor growth was attenuated by
TGF-β1 receptor inhibitor SB431542 and Rac1 inhibitor NSC23766,
concomitantly with the reduced expression of Ki-67 and CD34 in the
transplanted gastric tumor.
In conclusion, DADS inhibited EMT, invasion and
growth of gastric cancer cells by decreasing TGF-β1 expression,
concomitantly with reduced Rac1 and β-catenin expression. These
data indicated that the downregulation of TGF-β1/Rac1 pathway may,
in part, account for the molecular mechanisms through which DADS
exerts anti-EMT and antitumor growth effects in gastric cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Scientific Foundation of China (nos. 31000629, 31100935,
81102854, 81374013 and 81641112), The Key Project of Scientific
Research Foundation of Hunan Province Education Department of China
(no. 09A077), The Patency Foundation of Innovation Platform of
Hunan Provincial University of China (no. 09K074), The Key Project
of Scientific Research Foundation of Health and Family Planning
Committee of Hunan Province (no. A2015-2), The Scientific Research
Foundation of Health and Family Planning Committee of Hunan
Province (no. B2015-182) and The Construct Program of the Key
Discipline in Hunan Province of China (no. 2011-76).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
BS, JS and YZ conceived and designed the study. BS
and JS were involved in drafting and revising the manuscript. BS
and YZ performed the RT-PCR. JS performed the immunohistochemistry.
ED, FL and TT performed the cell culture and western blot analysis.
HX and YHW performed the cell migration and invasion assays. XZ and
HL were involved in the acquisition and analysis of the data. HJ,
XHA and QS reviewed and edited the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All experiments were performed according to the
guidelines for animal use of the Ethics Committee of the University
of South China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TGF-β1
|
transforming growth factor-β1
|
|
DADS
|
diallyl disulfide
|
|
EMT
|
epithelial-mesenchymal transition
|
|
LIMK1
|
LIM kinase-1
|
|
PAK1
|
P21 activated kinase-1
|
|
ROCK1
|
Rho-associated, coiled-coil containing
protein kinase 1
|
|
uPAR
|
urokinase-type plasminogen activator
receptor
|
|
ERK
|
extracellular regulated protein
kinases
|
|
MMP9
|
matrix metalloproteinase-9
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
AKT, PKB
|
protein kinase B
|
|
JNK
|
c-Jun N-terminal kinase
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
CSE
|
cigarette smoke extract
|
|
HRP
|
horseradish peroxidase
|
References
|
1
|
Orditura M, Galizia G, Sforza V,
Gambardella V, Fabozzi A, Laterza MM, Andreozzi F, Ventriglia J,
Savastano B, Mabilia A, et al: Treatment of gastric cancer. World J
Gastroenterol. 20:1635–1649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yi L and Su Q: Molecular mechanisms for
the anti-cancer effects of diallyl disulfide. Food Chem Toxicol.
57:362–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Katsuno Y, Lamouille S and Derynck R:
TGF-β signaling and epithelial-mesenchymal transition in cancer
progression. Curr Opin Oncol. 25:76–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heldin CH and Moustakas A: Signaling
receptors for TGF-β family members. Cold Spring Harb Perspect Biol.
8:a0220532016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang H, Liu L, Wang Y, Zhao G, Xie R, Liu
C, Xiao X, Wu K, Nie Y, Zhang H and Fan D: KLF8 involves in
TGF-beta-induced EMT and promotes invasion and migration in gastric
cancer cells. J Cancer Res Clin Oncol. 139:1033–1042. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo L, Peng W, Tao J, Lan Z, Hei H, Tian
L, Pan W, Wang L and Zhang X: Hydrogen sulfide inhibits
transforming growth factor-β1-induced EMT via Wnt/catenin pathway.
PLoS One. 11:e01470182016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moheimani F, Roth HM, Cross J, Reid AT,
Shaheen F, Warner SM, Hirota JA, Kicic A, Hallstrand TS, Kahn M, et
al: Disruption of β-catenin/CBP signaling inhibits human airway
epithelial-mesenchymal transition and repair. Int J Biochem Cell
Biol. 68:59–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang J, Yang B, Xiang T, Peng W, Qiu Z,
Wan J, Zhang L, Li H, Li H and Ren G: Diallyl disulfide inhibits
growth and metastatic potential of human triple-negative breast
cancer cells through inactivation of the β-catenin signaling
pathway. Mol Nutr Food Res. 59:1063–1075. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Al-Azayzih A, Gao F and Somanath PR: P21
activated kinase-1 mediates transforming growth factor β1-induced
prostate cancer cell epithelial to mesenchymal transition. Biochim
Biophys Acta. 1853:1229–1239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Su B, Su J, Zeng Y, Liu F, Xia H, Ma YH,
Zhou ZG, Zhang S, Yang BM, Wu YH, et al: Diallyl disulfide
suppresses epithelial-mesenchymal transition, invasion and
proliferation by downregulation of LIMK1 in gastric cancer.
Oncotarget. 7:10498–10512. 2016.PubMed/NCBI
|
|
12
|
Su B, Su J, He H, Wu Y, Xia H, Zeng X, Dai
W, Ai X, Ling H, Jiang H, et al: Identification of potential
targets for diallyl disulfide in human gastric cancer MGC-803 cells
using proteomics approaches. Oncol Rep. 33:2484–2494. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang H, Kong Y, Guo J, Tang Y and Xie X,
Yang L, Su Q and Xie X: Diallyl disulfide suppresses proliferation
and induces apoptosis in human gastric cancer through Wnt-1
signaling pathway by up-regulation of miR-200b and miR-22. Cancer
Lett. 340:72–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nyati S, Schinske K, Ray D, Nyati M, Ross
BD and Rehemtulla A: Molecular imaging of TGFβ-induced Smad2/3
phosphorylation reveals a role for receptor tyrosine kinases in
modulating TGFβ signaling. Clin Cancer Res. 17:7424–7439. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hwaiz R, Rahman M, Syk I, Zhang E and
Thorlacius H: Rac1-dependent secretion of platelet-derived CCL5
regulates neutrophil recruitment via activation of alveolar
macrophages in septic lung injury. J Leukoc Biol. 97:975–984. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pan R, Zhang Y, Zang B, Tan L and Jin M:
Hydroxysafflor yellow A inhibits TGF-β1-induced activation of human
fetal lung fibroblasts in vitro. J Pharm Pharmacol. 68:1320–1330.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pang L, Li Q, Wei C, Zou H, Li S, Cao W,
He J, Zhou Y, Ju X, Lan J, et al: TGF-β1/Smad signaling pathway
regulates epithelial-to-mesenchymal transition in esophageal
squamous cell carcinoma: In vitro and clinical analyses of cell
lines and nomadic Kazakh patients from northwest Xinjiang, China.
PLoS One. 9:e1123002014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Makrodouli E, Oikonomou E, Koc M, Andera
L, Sasazuki T, Shirasawa S and Pintzas A: BRAF and RAS oncogenes
regulate Rho GTPase pathways to mediate migration and invasion
properties in human colon cancer cells: A comparative study. Mol
Cancer. 10:1182011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shen HJ, Sun YH, Zhang SJ, Jiang JX, Dong
XW, Jia YL, Shen J, Guan Y, Zhang LH, Li FF, et al: Cigarette
smoke-induced alveolar epithelial-mesenchymal transition is
mediated by Rac1 activation. Biochim Biophys Acta. 1840:1838–1849.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang F, Zhuge YZ, Li YJ and Gu JX:
S-adenosylmethionine inhibits the activated phenotype of human
hepatic stellate cells via Rac1 and matrix metalloproteinases. Int
Immunopharmacol. 19:193–200. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Buongiorno P, Pethe VV, Charames GS,
Esufali S and Bapat B: Rac1 GTPase and the Rac1 exchange factor
Tiam1 associate with Wnt-responsive promoters to enhance
beta-catenin/TCF-dependent transcription in colorectal cancer
cells. Mol Cancer. 7:732008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu X, Tu X, Joeng KS, Hilton MJ, Williams
DA and Long F: Rac1 activation controls nuclear localization of
beta-catenin during canonical Wnt signaling. Cell. 133:340–353.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu G, Wang Y, Huang B, Liang J, Ding Y,
Xu A and Wu W: A Rac1/PAK1 cascade controls β-catenin activation in
colon cancer cells. Oncogene. 31:1001–1012. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jamieson C, Lui C, Brocardo MG,
Martino-Echarri E and Henderson BR: Rac1 augments Wnt signaling by
stimulating β-catenin-lymphoid enhancer factor-1 complex assembly
independent of β-catenin nuclear import. J Cell Sci. 128:3933–3946.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pethe VV, Charames GS and Bapat B: Rac1b
recruits Dishevelled and β-catenin to Wnt target gene promoters
independent of Wnt3A stimulation. Int J Oncol. 39:805–810.
2011.PubMed/NCBI
|
|
26
|
Saha SK, Choi HY, Kim BW, Dayem AA, Yang
GM, Kim KS, Yin YF and Cho SG: KRT19 directly interacts with
β-catenin/RAC1 complex to regulate NUMB-dependent NOTCH signaling
pathway and breast cancer properties. Oncogene. 36:332–349. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
De P, Carlson JH, Jepperson T, Willis S,
Leyland-Jones B and Dey N: RAC1 GTP-ase signals Wnt-beta-catenin
pathway mediated integrin-directed metastasis-associated tumor cell
phenotypes in triple negative breast cancers. Oncotarget.
8:3072–3103. 2017.PubMed/NCBI
|
|
28
|
Fan M, Xu Y, Hong F, Gao X, Xin G, Hong H,
Dong L and Zhao X: Rac1/β-catenin signalling pathway contributes to
trophoblast cell invasion by targeting Snail and MMP9. Cell Physiol
Biochem. 38:1319–1332. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Santibáñez JF, Kocić J, Fabra A, Cano A
and Quintanilla M: Rac1 modulates TGF-beta1-mediated epithelial
cell plasticity and MMP9 production in transformed keratinocytes.
FEBS Lett. 584:2305–2310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Booth DG, Takagi M, Sanchez-Pulido L,
Petfalski E, Vargiu G, Samejima K, Imamoto N, Ponting CP, Tollervey
D, Earnshaw WC, et al: Ki-67 is a PP1-interacting protein that
organises the mitotic chromosome periphery. Elife. 3:e016412014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang M, Wang X, Zhao Q, Liu T, Yao G, Chen
W, Li Z, Huang X and Zhang Y: Combined evaluation of the expression
of NUCKS and Ki-67 proteins as independent prognostic factors for
patients with gastric adenocarcinoma. Tumour Biol. 35:7505–7512.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bagheri-Yarmand R, Vadlamudi RK, Wang RA,
Mendelsohn J and Kumar R: Vascular endothelial growth factor
up-regulation via p21-activated kinase-1 signaling regulates
heregulin-beta1-mediated angiogenesis. J Biol Chem.
275:39451–39457. 2000. View Article : Google Scholar : PubMed/NCBI
|