Introduction
Cancer remains a major human health threat
worldwide. The incidence of cancer has increased to 14 million new
cases and 8.2 million cancer-related deaths in 2012 (1). Cancer is characterized by the
uncontrolled and invasive growth of cells (2). Chemotherapy is a traditional
treatment, but chemotherapeutic drugs have many side effects
(3,4). Therefore, there is an urgent
requirement for new anticancer drugs with advanced efficiency and
fewer side-effects (5).
Mangrove ecosystems are intertidal wetlands in
subtropical or tropical temperate coastal zones. These ecosystems
have a great commercial and ecological value for humans (6). Endophytes are microbial entities that
colonize within living tissues of living plants without causing
apparent disease symptoms in its hosts and are a relatively
unexplored potential source of novel and bioactive natural
compounds for exploitation in medicine, agriculture and industry
(7). Recently, more attention has
been paid to bioactive natural products of endophytic fungi,
isolated from mangrove plants. The secondary metabolites from
endophytic fungi exhibit a range of biological activities,
including antitumor, anti-inflammatory, antibacterial, antiviral,
antioxidative and anti-angiogenic (8–13). As
a result, many fungal endophytes have been isolated and subjected
to extensive chemical investigations during the past two decades
(14). In our previous study,
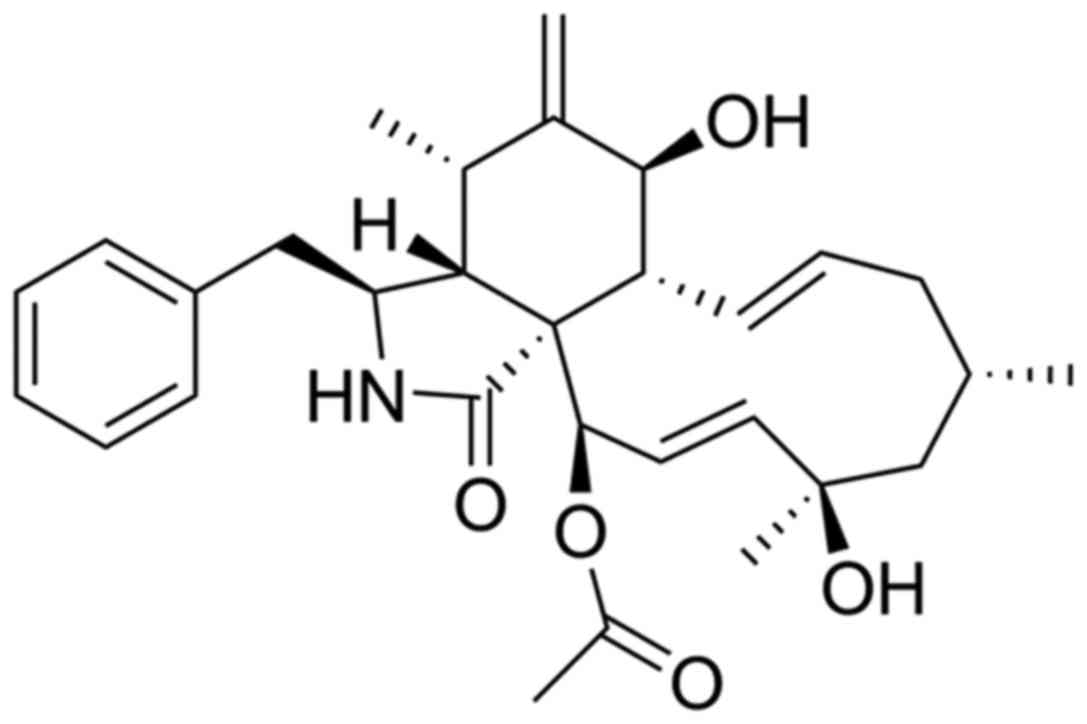
cytochalasin H (CyH) was isolated from the metabolic products of
endophytic fungus Phomopsis liquidambari, derived from a
mangrove in Zhanjiang, China.
Cytochalasins are a class of structurally related
fungal metabolites that are microfilament inhibitors with
substantially the same biological activity as that observed in the
inhibition of cell division, motility, secretion and phagocytosis
(15). Different cytochalasins have
their own unique functions. However, there has been a recent focus
on the anti-inflammatory, anti-fungal, and antitumor
pharmacological effects of CyH, each through different mechanisms
such as the induction of apoptosis and inhibition of angiogenesis
(16,17).
Apoptosis is the process of programmed cell death
regulated by a complex network of proliferation and survival genes.
Its disequilibrium, i.e., either through the acquisition of
anti-apoptotic signals or lack of pro-apoptotic signals, can result
in the failure of treatment or a variety of pathological conditions
such as cancer or autoimmune and degenerative diseases (18). Therefore, chemical agents that act
on molecular targets in apoptotic pathways are likely to be a
promising approach for cancer therapy.
In the present study, we observed the effect of CyH
on apoptosis and migration in A549 cells. To the best of our
knowledge, we found for the first time that CyH can induce
apoptosis and inhibit migration in A549 cells. Our findings provide
useful evidence for the anticancer activity of CyH in the treatment
of lung cancer.
Materials and methods
Drug and reagents
CyH, isolated previously by our laboratory from
mangrove endophytic fungus Phomopsis liquidambari in
Zhanjiang, China, was identified by nuclear magnetic resonance
(NMR) (Fig. 1). CyH was dissolved
in 0.1% dimethyl sulfoxide (DMSO) at a concentration of 1 mM, and
was then diluted in cell culture medium according to experimental
requirements. Cell Counting kit-8 (CCK-8), RIPA lysis buffer,
mitochondrial membrane potential (MMP) assay kit with JC-1, Hoechst
staining kit, cell cycle and apoptosis analysis kit, mouse
anti-human Bax antibody (1:1,000 dilution; cat. no. AB026-1),
rabbit anti-human Bcl-2 antibody (1:1,000 dilution; cat. no.
AB112-1), mouse anti-human β-actin antibody (1:2,000 dilution; cat.
no. AA128-1), and rabbit anti-human Bcl-xL antibody (1:1,000
dilution; cat. no. AB126-1) were purchased from Beyotime Institute
of Biotechnology (Shanghai, China). Phenylmethanesulfonyl fluoride
(PMSF) was purchased from Biosharp Biotechnology (Hefei, China).
Gibco RPMI-1640 medium was purchased from Thermo Fisher Scientific,
Inc. (Waltham, MA, USA).
Cell culture
The human lung adenocarcinoma cell line A549 was
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). A549 cells were cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS), 100 U/ml of
penicillin and 100 µg/ml streptomycin at 37°C and 5% CO2
in an incubator. Cells in the logarithmic phase of growth were used
in the experiment.
CCK-8 assay
The CCK-8 assay was performed to determine the
effect of CyH on the growth of A549 cells. Cells in the logarithmic
growth phase were plated onto 96-well plates at 5×103
cells/well. After adhering, the cells were treated with various
concentrations of CyH for 24, 48 and 72 h, respectively. CCK-8
solution (0 µl) from the CCK-8 was added to each well and the plate
was incubated at 37°C for an additional 1 h. Cell viability was
calculated as the absorbance value (A) at a wavelength of 450 nm
according to the manufacturer's instructions.
Analysis of cell cycle and cell
apoptosis
A549 cells in the logarithmic growth phase were
treated with different concentrations of CyH (0, 6.25, 12.5, 25 and
50 µM) for 48 h. For cell cycle analysis, the cells were harvested
by trypsinization and fixed in 70% ice-cold ethanol overnight.
Afterwards, the cells were centrifuged at 1,000 rpm for 5 min,
washed with PBS, and suspended with 0.5 ml of propidium iodide (PI)
for 30 min at 37°C. Cellular DNA was stained with PI for 10 min at
4°C in the dark before analysis by flow cytometry with a MultiCycle
AV DNA analysis software (Beckman Coulter, Inc., Brea, CA,
USA).
Analysis of MMP
Cell apoptosis was determined using the MMP assay.
MMP was detected using JC-1. Aggregate red fluorescence indicates
high MMP and cells in a normal state, while green fluorescence
indicates reduced MMP such as during the early events of apoptosis.
The change in MMP was detected using fluorescence microscopy (Nikon
Corp., Tokyo, Japan).
Migration assay
The effect of CyH on cell migration was evaluated by
scratch wound healing and chamber migration assays. For the scratch
wound healing assay, 100 µl of A549 cell suspension was seeded onto
6-well plates at a density of 5×104/well, and then
cultured into monolayers. Wounds were generated in the cell
monolayer by scratching with a sterile 10 µl pipette tip. The
medium was then removed and the cells were rinsed 3 times with PBS
to wash away cell debris. Cells were then treated with different
concentrations of CyH (0, 0.05, 0.1, 0.2, 0.4 and 0.8 µM) and the
wound migration distance was imaged at 0, 24, 48 and 72 h,
respectively, under a light microscope (Nikon Corp.) at a
magnification of ×100. For the Transwell chamber migration assay,
A549 cells (5×104/ml) were pre-treated with CyH (0,
0.05, 0.10, 0.20, 0.4 and 0.8 µM) for 48 h and then plated in the
top chambers in FBS-free medium. The medium at the bottom chamber
was supplemented with 10% FBS to allow migration towards the
chemoattractant for 48 h in a 37°C chamber. Cells at the bottom of
the membrane were fixed in 4% paraformaldehyde for 30 min and
stained with 0.1% crystal violet for 1 h. The number of cells was
counted in at least five randomized fields under a light microscope
(Nikon Corp.) at a magnification of ×200.
Analysis of apoptosis-related protein
expression
Proteins were extracted from A549 cells by RIPA
lysis buffer containing protease inhibitors, cultured in 6-well
plates and treated with CyH (0, 6.25, 12.5, 25, 50 and 100 µM) for
48 h. Protein concentrations were detected by the BCA protein
assay. Afterwards, 100 µg of protein was separated on 10% SDS-PAGE
and was then transferred to a PVDF membrane (EMD Millipore,
Billerica, MA, USA). The membrane was blocked with 5% non-fat milk
in TBS containing 0.1% Tween-20 (TBST) for 2 h at room temperature.
The membranes were incubated overnight at 4°C with Bax, Bcl-2,
Bcl-xL, caspase-3 and P53 primary antibodies, respectively and
β-actin was used as an internal control. Target proteins were
detected by ECL reagents and then were exposed to X-ray film
(Carestream Health, Inc., Xiamen, China). The band density was
analyzed with ImageJ software (National Institutes of Health,
Bethesda, MD, USA).
Statistical analysis
All data are showed as mean ± standard deviation.
One-way ANOVA assay was performed for data analysis using SPSS 16.0
statistical software (SPSS, Inc., Chicago, IL, USA). P<0.05 was
indicative of a statistically significant difference.
Results
Effect of CyH on cell viability in
A549 cells
The results from the CCK-8 assay (data not shown)
confirmed that CyH inhibited the proliferation of A549 cells.
IC50 value for CyH in the A549 cells was 159.50±1.048
µM. Subsequently, the optimal experimental concentrations of CyH
(0, 6.25, 12.5, 25, 50 and 100 µM) were chosen.
Analysis of cell cycle distribution
and cell apoptosis
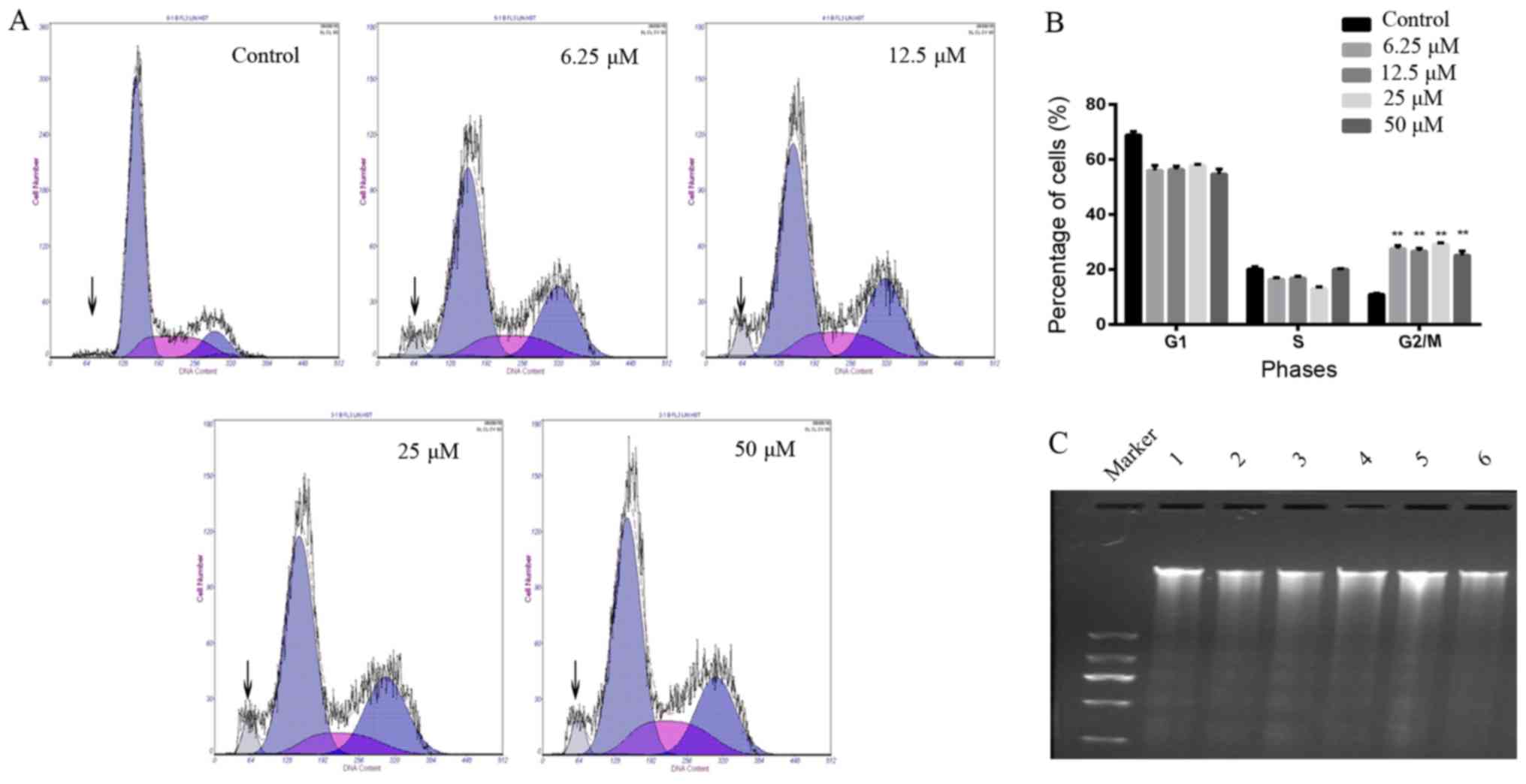
The results from the cell cycle assay confirmed that
the cell cycle was arrested at the G2/M phase and
sub-G1 peaks were found after A549 cells were treated
with different concentrations of CyH (Fig. 2A). The proportion of DNA in cells in
the G2/M phase was increased following treatment of CyH
with different concentrations. Different drug concentration groups
had statistical differences when compared to the control group
(P<0.01, Fig. 2B). Furthermore,
fragmented DNA ladder is an important indicator of apoptosis. As
expected, fragmented DNA ladders were found in the CyH-treated
cells. As shown in Fig. 2C, we
observed fragmented DNA ladders by agarose gel electrophoresis.
Cell apoptosis was further analyzed
using MMP assay
MMP levels were examined using a JC-1 sensitive
fluorescent probe by fluorescence microscopy. Moreover, the red
fluorescence of JC-1 was significantly decreased and the green
fluorescence was markedly increased (Fig. 3), indicating that MMP in the
CyH-treated cells was decreased. Taken together, our results
demonstrated that CyH induced apoptosis in the A549 cells.
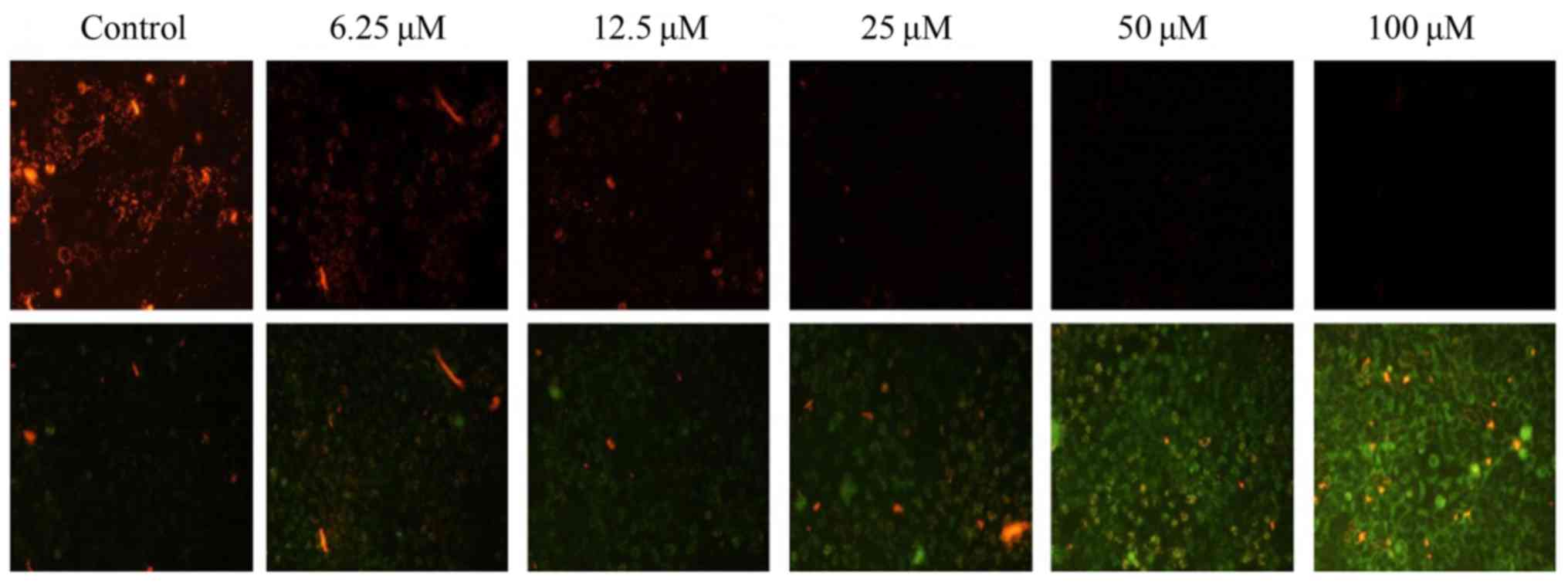 | Figure 3.MMP was measured using JC-1 dye
staining under fluorescence microscopy. A549 cells were treated
with different concentrations (0, 6.25, 12.5, 25, 50 and 100 µM) of
CyH for 48 h. In upper row, red fluorescence represents
mitochondria with intact membrane potential. After treated, the red
fluorescence was decreased. In lower row, green fluorescence
represents de-energized mitochondria. In treated cells, green
fluorescence was noted in A549 cells indicating the dissipation of
the MMP. MMP, mitochondrial membrane potential; CyH, cytochalasin
H. |
Analysis of apoptosis-related protein
expression
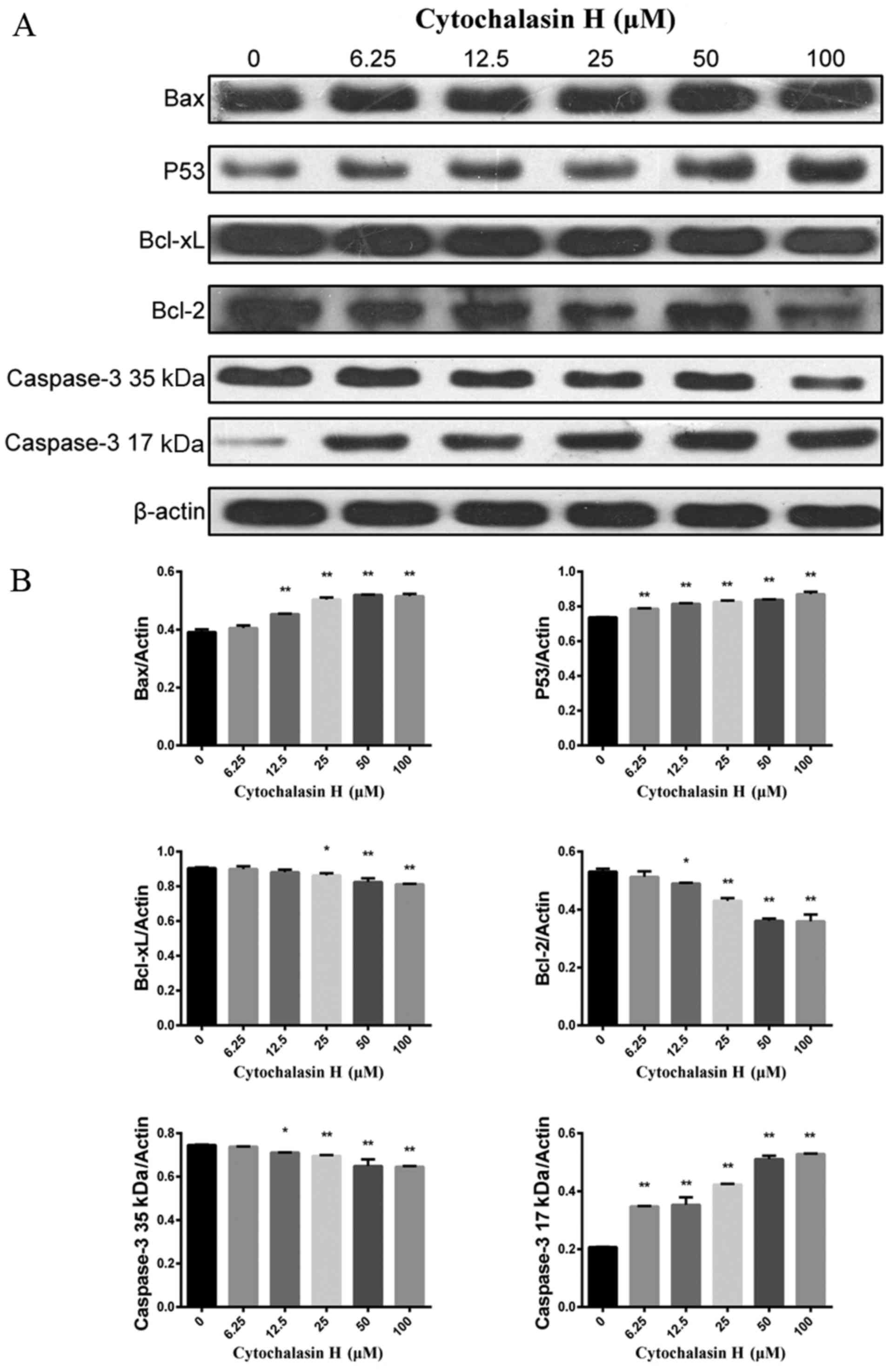
To investigate the mechanism by which CyH induces
the apoptosis of A549 cells, the protein expression levels of
apoptosis-related proteins were determined by western blotting.
Bcl-2 and Bcl-xL, two anti-apoptotic proteins, inhibit programmed
cell death; Bax and P53, two pro-apoptotic proteins, drive the cell
towards apoptosis. As shown in Fig.
4A, as the concentration of CyH increased, the protein
expression levels of Bcl-xL, Bcl-2 and full-length caspase-3 (35
kDa) were decreased while the protein expression levels of Bax, P53
and cleaved caspase-3 (17 kDa) were increased. The differences
between the groups were statistically significant (P<0.01,
Fig. 4B).
Migration assay
To analyze the effect of CyH on the metastasis of
A549 cells, scratch wound healing and Transwell migration assays
were used to analyze cell migration. The results from the scratch
wound healing assay showed that the migration ability in
CyH-treated A549 cells was significantly inhibited compared with
that of the control group (Fig.
5A). Furthermore, it revealed that the migration distances of
CyH-treated A549 cells were significantly decreased compared with
that of the control cells (P<0.01, Fig. 5B). A Transwell migration assay was
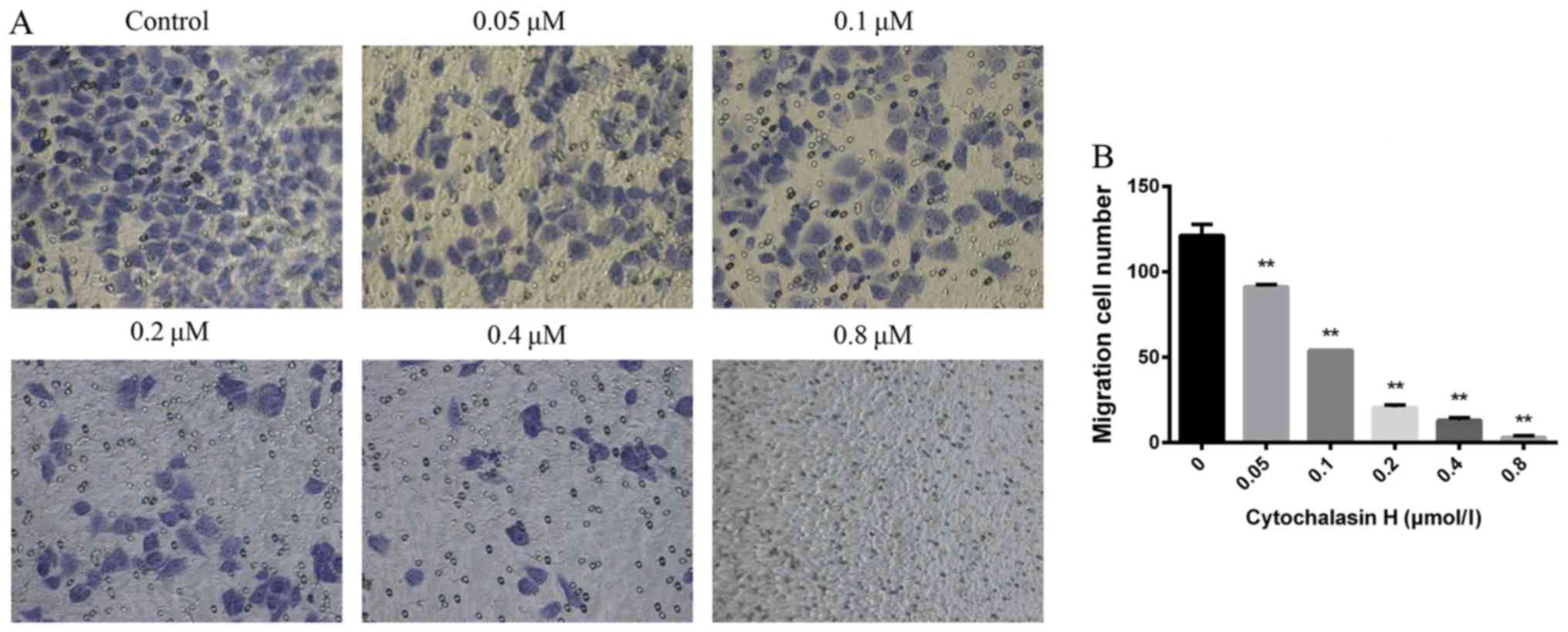
carried out to further confirm the effect of CyH on the migration
of A549 cells. Our results showed that the number of A549 cells
moving into the chamber was reduced gradually as the concentration
of CyH increased compared with the control group (Fig. 6A). These results demonstrated that
the difference between the CyH-treated cells and the control cells
was statistically significant (P<0.01, Fig. 6B), indicating that CyH suppressed
the migration ability of A549 cells.
Discussion
High rates of morbidity and mortality have made
cancers a major public health concern (19). The treatment of cancers mainly
involves radiotherapy combined with chemotherapy and other
comprehensive treatments (20).
However, conventional chemotherapy drugs have serious adverse
reactions and are prone to induce drug resistance, and the
long-term effects are not ideal (21). The secondary metabolites from
mangrove endophytic fungi have the advantages of high biological
activity and low toxicity.
In our previous study, we isolated CyH from the
endophytic fungus Phomopsis liquidambari. In the present
study, we further analyzed the effects of CyH on proliferation in
A549 cells. We found that CyH inhibited the proliferation of A549
cells. The cell cycle is important for the proliferation of cancer
cells (22). The goal of targeting
cell proliferation is to arrest the cell cycle or induce cancer
cell death using cytotoxic compounds. Cells treated with Taxol
extracted from yews are unable to proceed normally through the cell
cycle and are arrested in the G1 and G2/M
phases (23–25). As shown in Fig. 2, our findings suggest that CyH
inhibits the proliferation of A549 cells attributed to
G2/M phase arrest. However, G2/M arrest may
stop cell cycle progression, which will also affect DNA synthesis,
thus further investigation is needed.
Apoptosis, the process of programmed cell death, has
been recognized as one of the major processes that mediate the
inhibition of cell proliferation, which may be targeted by
anticancer agents (26). Cells
undergoing apoptosis exhibit morphological and biochemical
modifications including chromatin segregation, nuclear
condensation, DNA fragmentation, partition of the membrane, and
vesicle formation (27,28). The late-stage of apoptosis can be
visualized by standard agarose gel electrophoresis as a ladder
pattern because of DNA cleavage (29). In the present study, we found
fragmented DNA ladders in the CyH-treated A549 cells (Fig. 2C). To further confirm the effects of
CyH on apoptosis, MMP assay, PI staining, and flow cytometry were
performed. Our results showed that CyH induced apoptosis in the
A549 cells. p53, a tumor-suppressor transcription factor, plays a
vital role in cell cycle arrest and apoptosis in response to
cellular stress (30). The
functional p53 encodes a nuclear phosphoprotein that regulates the
synthesis of gene products involved in growth arrest, DNA repair,
apoptosis and the inhibition of angiogenesis (31). According to the present results, the
expression level of p53 was significantly upregulated following CyH
treatment (Fig. 4). It could be
concluded that CyH induces apoptosis in a p53-dependent pathway.
The potent anticancer activity of p53 has been linked to its
ability to induce apoptosis through the intrinsic
mitochondrial-mediated apoptotic pathway (32). The mitochondrial apoptotic pathway
is mainly regulated by Bcl-2 family proteins. Any imbalances in the
expression level of pro-apoptotic Bax and anti-apoptotic Bcl-2
members lead to the disruption of the outer mitochondrial membrane
(33,34). Upon apoptotic stimulation, the
expression of Bax is increased, leading to a lower level of Bcl-2
(35). Bax forms oligomers and is
transported from the cytoplasm to the mitochondrial membrane,
resulting in mitochondrial membrane depolarization (36). Subsequently, cytochrome c is
released from mitochondria to the cytosol, triggering caspase
pathway activation (37). Caspases,
playing a key role in apoptotic events, adjust cell death, and the
appearance and function of caspase apoptotic features are closely
associated (38). Caspase-3, a
crucial downstream effector of the caspase family, is thought to be
involved in both the mitochondrial apoptotic pathway and the death
receptor pathway (39). Full-length
caspase-3 (35 kDa), an inactive protein, is cleaved between Asp28
and Ser29 as well as between Asp175 and Ser176 to produce cleaved
caspase-3 (17 kDa), an active peptide. Apoptosis will occur in the
cells when full-length caspase-3 (35 kDa) protein levels are
decreased and cleaved caspase-3 (17 kDa) protein levels are
increased. In this study, we demonstrated that Bcl-2, full-length
caspase-3 (35 kDa) and Bcl-xL protein levels were significantly
decreased. Meanwhile, Bax and cleaved caspase-3 (17 kDa) protein
levels were significantly increased (Fig. 4). As a result, the ratios of
anti-apoptotic proteins and pro-apoptotic proteins were
significantly reduced during apoptosis. This imbalance led to the
loss of MMP after CyH treatment (Fig.
3).
Migration is a critical step in the initial
progression of cancer that facilitates metastasis. The scratch
wound healing assay is a classic and common method used for the
discovery and validation of molecules that affect cell migration
and metastasis (40–42). The results from this study indicated
that CyH showed strong anti-migratory activities (Figs. 5 and 6). The underlying mechanisms of how CyH
inhibits migration ability in A549 cells require further
investigated.
In summary, we demonstrated for the first time and
to the best of our knowledge that CyH significantly induced cell
apoptosis and inhibited migration in A549 cells. Furthermore, we
found that CyH induced apoptosis in A549 cells by the
downregulation of Bcl-xL, Bcl-2 and full-length caspase-3 (35 kDa)
protein levels and the upregulation of Bax, P53 and cleaved
caspase-3 (17 kDa) protein levels. These findings suggest that CyH
may be developed into a potential chemotherapeutic drug for the
treatment of lung cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by grants
from the National Natural Science Foundation of China (no. 81372511
to XT); the Special Fund for Scientific and Technological
Development (Basic and Applied Basic Research) of Guangdong
Province in 2017 (Natural Science Foundation of Guangdong Province)
(no. 2017A030313539 to XT); the Guangdong Provincial Department of
Science and Technology (Research and Development of Industrial
Technology in Guangdong Province) (no. 2013B031100002 to XT); and
the ‘Sail Plan’ in Guangdong Province to Cultivate High-Level
Talents (no. 201635011 to XT).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
XT, YM and XW conceived and designed the study. YM,
ZX, XL, BH, LH, JL and ZZ performed the experiments. YM and XT
wrote the report. XT and XW reviewed and edited the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bhavana J, Kalaivani MK and Sumathy A:
Cytotoxic and pro-apoptotic activities of leaf extract of Croton
bonplandianus Baill. against lung cancer cell line A549. Indian
J Exp Biol. 54:379–385. 2016.PubMed/NCBI
|
|
2
|
Ramos-Silva A, Tavares-Carreón F, Figueroa
M, De la Torre-Zavala S, Gastelum-Arellanez A, Rodríguez-García A,
Galán-Wong LJ and Avilés-Arnaut H: Anticancer potential of
Thevetia peruviana fruit methanolic extract. BMC Complement
Altern Med. 17:2412017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Velmurugan BK, Yang HH, Sung PJ and Weng
CF: Excavatolide B inhibits nonsmall cell lung cancer proliferation
by altering peroxisome proliferator activated receptor gamma
expression and PTEN/AKT/NF-Kβ expression. Environ Toxicol.
32:290–301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhu L, Chen Y, Wei C, Yang X, Cheng J,
Yang Z, Chen C and Ji Z: Anti-proliferative and pro-apoptotic
effects of cinobufagin on human breast cancer MCF-7 cells and its
molecular mechanism. Nat Prod Res. 32:493–497. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kumar Nishanth S, Aravind SR, Jacob J,
Gopinath G, Lankalapalli RS, Sreelekha TT and Kumar Dileep BS:
Pseudopyronine B: A potent antimicrobial and anticancer molecule
isolated from a pseudomonas mosselii. Front Microbiol.
7:13072016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gong B, Liu G, Liao R, Song J and Zhang H:
Endophytic fungus Purpureocillium sp. A5 protect mangrove
plant Kandelia candel under copper stress. Braz J Microbiol.
48:530–536. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng MJ, Wu MD, Chan HY, Hsieh SY, Chen
YL, Chen IS, Chan FL, Chen JJ and Yuan GF: Secondary metabolites
produced by Phomopsis sp. 11F0023, an endophytic fungus in
Eragrostis amabilis. Chem Nat Compd. 51:431–434. 2015.
View Article : Google Scholar
|
|
8
|
Bhatia DR, Dhar P, Mutalik V, Deshmukh SK,
Verekar SA, Desai DC, Kshirsagar R, Thiagarajan P and Agarwal V:
Anticancer activity of Ophiobolin A, isolated from the endophytic
fungus Bipolaris setariae. Nat Prod Res. 30:1455–1458. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ju ZR, Qin X, Lin XP, Wang JF,
Kaliyaperumal K, Tian YQ, Liu J, Liu F, Tu Z, Xu SH, et al: New
phenyl derivatives from endophytic fungus Botryosphaeria sp.
SCSIO KcF6 derived of mangrove plant Kandelia candel. Nat
Prod Res. 30:192–198. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zheng CJ, Huang GL, Xu Y, Song XM, Yao J,
Liu H, Wang RP and Sun XP: A new benzopyrans derivatives from a
mangrove-derived fungus Penicillium citrinum from the South
China Sea. Nat Prod Res. 30:821–825. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
El-Gendy MM, El-Bondkly AM and Yahya SM:
Production and evaluation of antimycotic and antihepatitis C virus
potential of fusant MERV6270 derived from mangrove endophytic fungi
using novel substrates of agroindustrial wastes. Appl Biochem
Biotechnol. 174:2674–2701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cai R, Chen S, Liu Z, Tan C, Huang X and
She Z: A new α-pyrone from the mangrove endophytic fungus
Phomopsis sp. HNY29-2B. Nat Prod Res. 31:124–130. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu X, Wu X, Ma Y, Zhang W, Hu L, Feng X,
Li X and Tang X: Endophytic fungi from mangrove inhibit lung cancer
cell growth and angiogenesis in vitro. Oncol Rep. 37:1793–1803.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
El-Hawary SS, Sayed AM, Rateb ME, Bakeer
W, AbouZid SF and Mohammed R: Secondary metabolites from fungal
endophytes of Solanum nigrum. Nat Prod Res. 31:2568–2571.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yahara I, Harada F, Sekita S, Yoshihira K
and Natori S: Correlation between effects of 24 different
cytochalasins on cellular structures and cellular events and those
on actin in vitro. J Cell Biol. 92:69–78. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chapla VM, Zeraik ML, Ximenes VF, Zanardi
LM, Lopes MN, Cavalheiro AJ, Silva DH, Young MC, Fonseca LM,
Bolzani VS, et al: Bioactive secondary metabolites from
Phomopsis sp., an endophytic fungus from Senna
spectabilis. Molecules. 19:6597–6608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yi JM, Kim J, Park JS, Lee J, Lee YJ, Hong
JT, Bang OS and Kim NS: In vivo anti-tumor effects of the ethanol
extract of gleditsia sinensis thorns and its active
constituent, cytochalasin h. Biol Pharm Bull. 38:909–912. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tao YW, Lin YC, She ZG, Lin MT, Chen PX
and Zhang JY: Anticancer activity and mechanism investigation of
beauvericin isolated from secondary metabolites of the mangrove
endophytic fungi. Anticancer Agents Med Chem. 15:258–266. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang JY, Tao LY, Liang YJ, Yan YY, Dai
CL, Xia XK, She ZG, Lin YC and Fu LW: Secalonic acid D induced
leukemia cell apoptosis and cell cycle arrest of G(1) with
involvement of GSK-3beta/beta-catenin/c-Myc pathway. Cell Cycle.
8:2444–2450. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
D'cruz A, Lin T, Anand AK, Atmakusuma D,
Calaguas MJ, Chitapanarux I, Cho BC, Goh BC, Guo Y, Hsieh WS, et
al: Consensus recommendations for management of head and neck
cancer in Asian countries: A review of international guidelines.
Oral Oncol. 49:872–877. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang YH, Zhang Y, Li XY, Feng XD, Jian W
and Li RQ: Antitumor activity of the pachymic acid in
nasopharyngeal carcinoma cells. Ultrastruct Pathol. 41:245–251.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chandar N, Billig B, McMaster J and Novak
J: Inactivation of p53 gene in human and murine osteosarcoma cells.
Br J Cancer. 65:208–214. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoo YD, Park JK, Choi JY, Lee KH, Kang YK,
Kim CS, Shin SW, Kim YH and Kim JS: CDK4 down-regulation induced by
paclitaxel is associated with G1 arrest in gastric
cancer cells. Clin Cancer Res. 4:3063–3068. 1998.PubMed/NCBI
|
|
25
|
Mullan PB, Quinn JE, Gilmore PM,
McWilliams S, Andrews H, Gervin C, McCabe N, McKenna S, White P,
Song YH, et al: BRCA1 and GADD45 mediated G2/M cell
cycle arrest in response to antimicrotubule agents. Oncogene.
20:6123–6131. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bai L and Wang S: Targeting apoptosis
pathways for new cancer therapeutics. Annu Rev Med. 65:139–155.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fisher DE: Apoptosis in cancer therapy:
Crossing the threshold. Cell. 78:539–542. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yi JM, Kim MS, Lee EH, Wi DH, Lee JK, Cho
KH, Hong SH and Kim HM: Induction of apoptosis by Paljin-Hangahmdan
on human leukemia cells. J Ethnopharmacol. 88:79–83. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saadat Rahbar Y, Saeidi N, Vahed Zununi S,
Barzegari A and Barar J: An update to DNA ladder assay for
apoptosis detection. Bioimpacts. 5:25–28. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen C, Hu SY, Luo DQ, Zhu SY and Zhou CQ:
Potential antitumor agent from the endophytic fungus
Pestalotiopsis photiniae induces apoptosis via the
mitochondrial pathway in HeLa cells. Oncol Rep. 30:1773–1781. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kralj M, Husnjak K, Körbler T and Pavelić
J: Endogenous p21WAF1/CIP1 status predicts the response of human
tumor cells to wild-type p53 and p21WAF1/CIP1 overexpression.
Cancer Gene Ther. 10:457–467. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chipuk JE and Green DR: Dissecting
p53-dependent apoptosis. Cell Death Differ. 13:994–1002. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chipuk JE and Green DR: How do BCL-2
proteins induce mitochondrial outer membrane permeabilization?
Trends Cell Biol. 18:157–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Besbes S, Mirshahi M, Pocard M and Billard
C: New dimension in therapeutic targeting of BCL-2 family proteins.
Oncotarget. 6:12862–12871. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Boatright KM and Salvesen GS: Mechanisms
of caspase activation. Curr Opin Cell Biol. 15:725–731. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tait SW and Green DR: Mitochondria and
cell death: Outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Erşahin M, Çevik Ö, Akakın D, Şener A,
Özbay L, Yegen BC and Şener G: Montelukast inhibits caspase-3
activity and ameliorates oxidative damage in the spinal cord and
urinary bladder of rats with spinal cord injury. Prostaglandins
Other Lipid Mediat. 99:131–139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang X, Tan T, Mao ZG, Lei N, Wang ZM, Hu
B, Chen ZY, She ZG, Zhu YH and Wang HJ: The marine metabolite
SZ-685C induces apoptosis in primary human nonfunctioning pituitary
adenoma cells by inhibition of the Akt pathway in vitro. Mar Drugs.
13:1569–1580. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gulhati P, Bowen KA, Liu J, Stevens PD,
Rychahou PG, Chen M, Lee EY, Weiss HL, O'Connor KL, Gao T, et al:
mTORC1 and mTORC2 regulate EMT, motility, and metastasis of
colorectal cancer via RhoA and Rac1 signaling pathways. Cancer Res.
71:3246–3256. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: A convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yarrow JC, Perlman ZE, Westwood NJ and
Mitchison TJ: A high-throughput cell migration assay using scratch
wound healing, a comparison of image-based readout methods. BMC
Biotechnol. 4:212004. View Article : Google Scholar : PubMed/NCBI
|