Introduction
Colorectal cancer (CRC) is the third most common
type of cancer worldwide, and the fourth leading cause of
cancer-related deaths, with an incidence of more than one million
new cases per year (1–5). In addition, a significant increase of
the incidence rates in CRC in both sexes has been reported, in
Korea from 1999 to 2010 (4). The
development and progression of CRC occur over several years with
distinct molecular and cytological characteristics, eventually
leading to a carcinoma with a higher risk of invasion and
metastasis (3,6). The prevalent metastasis of CRC is to
the lymph node and the liver, which influence approximately half of
CRC patients, with a poor survival rate after the diagnosis
(7–9). It has been demonstrated that one of
the important factors causing progression and poor prognosis of
cancer is metastasis (10–12). Though tumor/node/metastasis (TNM)
classification and staging systems are widely used to predict
prognosis of CRC patients, it remains difficult to precisely
predict prognosis (13). Thus, it
has been suggested that it is urgently required to identify
specific biomarkers to improve the outcomes of patients with CRC
(14).
The cell cycle is tightly regulated to prevent
premature cell cycle progression, leading to faithful gene
replication and segregation (15).
Cyclin-dependent kinase 1 also known as CDK1 or cell division cycle
2 (CDC2) is an essential enzyme in cell cycle progression,
particularly in the regulation of the G2/M cell cycle transition
(16). The activity of CDK1 is
regulated by cyclin binding and phosphorylation (17). The phosphorylation of CDK1 at Tyr15
and Thr14 residues leads to the inactivation of CDK1, resulting in
a blockade of the transition from G2 to the mitotic phase (17). The protein kinase,
membrane-associated tyrosine/threonine 1 (PKMYT1) was originally
identified from Xenopus laevis as Myt1, a
membrane-associated inhibitory kinase that phosphorylates CDK1
efficiently on both threonine-14 and tyrosine-15 (18). Subsequently, it was cloned from
human with 46% identification to Xenopus laevis PKMYT1 and
localization to the endoplasmic reticulum and Golgi complex
(19). Thus, PKMYT1 was
demonstrated to inhibit the progression of the cell cycle through
the phosphorylation of CDK1. In addition, it was revealed that
PKMYT1 was required to induce apoptosis, leading to the suppression
of skin cancer development induced by UVA (20). Recently, however, several studies
reported that although PKMYT1 is dispensable for the normal cell
cycle, it has a rate-limiting function during checkpoint recovery
after DNA damage or it is an essential molecule in glioblastoma
stem-like cells, thus highlighting a potential role of PKMYT1 as a
therapeutic target (21,22). Since the functional importance of
PKMYT1 in CRC remains unknown, it is important to comprehend
whether PKYMT1 is indispensable in CRC. Thus, using CRC cell lines
and patient-derived samples, we addressed these issues and reported
that PKMYT1 was essential in the progression of CRC.
Materials and methods
Cell lines
Human CRC cell lines SW480, SW620, HCT116 and HT29
were purchased from the Korean Cell Line Bank (KCLB; Seoul, Korea).
Cells were grown in RPMI-1640 medium (Welgene, Geyongsan, Korea)
supplemented with 10% fetal bovine serum (FBS; Corning Inc.,
Corning, NY, USA) and 1% penicillin/streptomycin (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) at 37°C in a humidified
atmosphere containing 5% CO2.
siRNA and transfection
The siRNAs were obtained from Bioneer Corporation
(Daejeon, Korea) (RNAi nos. 1117350, 1117347 and 1117348) and
transfected using HiPerFect Transfection reagent (Qiagen, Inc.,
Valencia, CA, USA) according to the manufacturer's
instructions.
RNA extraction and realtime qPCR
RNA was isolated using RiboEx solution (GeneAll,
Seoul, Korea), and converted to cDNA using ReverTra Ace®
qPCR kit (Toyobo Life Science, Osaka, Japan) according to the
manufacturer's instructions. To determine the level of gene
expression, RT-qPCR was performed using the qPCR Master Mix kit
(Toyobo Life Science). The primer sequences for RT-qPCR are listed
in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Name | Sequence | Assay |
|---|
| PKMYT1 primer
forward |
5′-GGAGAACTGGGTTAAGATGC-3′ | RT-PCR |
| PKMYT1 primer
reverse |
5′-TCCACTTCCAGAATGGCAGT-3′ | RT-PCR |
| GAPDH primer
forward |
5′-CTTAGCACCCCTGGCCAAG-3′ | RT-PCR |
| GAPDH primer
reverse |
5′-GATGTTCTGGAGAGCCCCG-3′ | RT-PCR |
Western blot analysis
Cell lysates were harvested using Pro-Prep (Intron
Biotechnology Inc., Seongnam, Korea) and centrifuged at 16,000 × g
for 10 min at 4°C. Protein concentration of the supernatant was
determined by Bio-Rad Protein assay (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The protein extract was resolved using 10%
polyacrylamide gel and electro-transferred onto a 0.45-µm
hybridization nitrocellulose filter (HATF) membrane (Millipore,
Billerica, MA, USA) using Trans-Blot Turbo (Bio-Rad Laboratories,
Inc.). Membranes were immunoblotted with either mouse anti-PKMYT1
primary antibody (cat. no. NBP2-2275; Novus Biologicals, LLC,
Littleton, CO, USA) diluted 1:500 in TBS-T or rabbit polyclonal
anti-actin antibody (cat. no. ab179467; Abcam, Cambridge, MA, USA)
overnight at 4°C. The membranes were incubated with either
HRP-conjugated anti-rabbit immunoglobulin (cat. no. 7071; Cell
Signaling Technology, Inc., Danvers, MA, USA) or HRP-linked
anti-mouse immunoglobulin (cat. no. 7076; Cell Signaling
Technology, Inc.) for 1 h at room temperature. The protein signal
was detected by enhanced chemiluminescence (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) using the Amersham Imager 600
(GE Healthcare Life Sciences, Chalfont, UK).
Invasion and migration
Cell migration and invasion were analyzed in
vitro using the Transwell insert system (Corning Incorporated)
without or with coating by 20 µl of Matrigel (BD Biosciences,
Franklin Lakes, USA). The culture insert was attached on the bottom
of a 24-well plate and 100 µl of serum-free medium containing
1.0×105 cells was seeded into each well. Six-hundred
microliters of media containing 10% FBS were added outside the
Transwell culture insert. Cells were incubated at 37°C for 48 h in
a humidified atmosphere with 5% CO2. Transwells were
washed twice with phosphate-buffered saline (PBS) and cleaned using
cotton swaps. The cells were fixed with 1% formaldehyde for 15 min,
washed twice with PBS, stained with 0.1% of crystal violet for 15
min and then observed using an inverted microscope (Leica
Microsystems GmbH, Wetzlar, Germany).
Semi-solid agar colony forming
assay
Cell culture media containing 20% FBS was mixed with
1% agarose at 1:1 ratio and was solidified after dropping a few
drops. After mixing the cell-containing media with agarose to make
0.35% agarose, cells were spread and incubated for 17 days at 37°C
in 5% CO2 incubator.
Human colorectal carcinoma
specimens
A total of 179 colorectal carcinoma tissue specimens
were obtained from Soonchunhyang University Cheonan Hospital
(Cheonan, Korea), where samples were collected from patients who
underwent surgery between March 2002 and May 2006. This study was
conducted in accordance with the ethical standard of the Ethics
Committee of Soonchunhyang University Cheonan Hospital. These
tissues were formalin-fixed and paraffin-embedded (FFPE).
Clinicopathological data including age, sex, TNM classification,
invasion of blood vessel and lymphatic vessel are listed at
Table II. Tumor stage was
identified according to the American Joint Committee on Cancer's
TNM classification system.
| Table II.Clinicopathological features in
patients with PKMYT1 expression. |
Table II.
Clinicopathological features in
patients with PKMYT1 expression.
|
| PKMYT1
expression |
|
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
factors | Low (N=94) | High (N=85) | Total (N=179) | P-value |
|---|
| Age, years |
|
|
| 0.256 |
|
<60 | 25 (26.6) | 30 (25.3) | 55 (30.7) |
|
|
≥60 | 69 (73.4) | 55 (64.7) | 124 (69.3) |
|
| Sex |
|
|
| 0.449 |
|
Male | 37 (39.4) | 39 (45.9) | 103 (57.5) |
|
|
Female | 57 (60.6) | 46 (54.1) | 76 (42.5) |
|
| Vascular
invasion |
|
|
| 1.000 |
| No | 75 (79.8) | 68 (80.0) | 143 (79.9) |
|
|
Yes | 19 (20.2) | 17 (20.0) | 36 (20.1) |
|
| Lymphatic
invasion |
|
|
| 0.180 |
| No | 64 (68.1) | 66 (77.6) | 130 (72.6) |
|
|
Yes | 30 (31.9) | 19 (22.4) | 49 (27.4) |
|
| Perineural
invasion |
|
|
| 0.542 |
| No | 87 (82.6) | 81 (95.3) | 168 (93.9) |
|
|
Yes | 7 (7.4) | 4 (4.7) | 11 (6.1) |
|
| Stage |
|
|
| 0.455 |
| I,
II | 45 (47.9) | 46 (54.1) | 91 (50.8) |
|
| III,
IV | 49 (52.1) | 39 (45.9) | 88 (49.2) |
|
Tissue microarray (TMA) and
immunohistochemistry (IHC) assay
Immunohistochemical staining was performed using
tissue microarray (TMA) block sections to determine the expression
of PKMYT1 in CRC patient samples. Each TMA block contained 60 cores
(2 mm of size) from 30 samples. For immunohistochemistry, 4-µm
sections were obtained using a microtome, deparaffinized in xylene
and rehydrated in 100–70% alcohol series. Antigen retrieval was
achieved in citrate buffer (pH 6.0) using a microwave for 15 min.
To eliminate endogenous peroxidase activity, the sections were
incubated in peroxidase blocking solution (Dako; Agilent
Technologies GmbH, Waldbronn, Germany) for 30 min and then washed
with PBS containing 0.1% Tween-20 (PBST). The sections were
incubated with anti-PKMYT1 antibody (1:10 dilution; cat. no.
NBP2-2275; Novus Biologicals, LLC) for 2 h at room temperature,
followed by a secondary antibody for 1 h 30 min at room
temperature. After being washed with PBST, the sections were
incubated with DAB and observed under a light microscope.
IHC data analysis
The PKMYT1-stained tissue cores were examined and a
consensus score was determined for each specimen. A positive
reaction was scored into 4 grades, according to the intensity of
the staining as follows: 0, 1+, 2+ and 3+. The percentages of
PKMYT1-positive cells were also scored into 4 categories as
follows: 0, 0; 1, 1–30; 2, 31–70; and 3, 71–100%. The final score,
calculated as the product of the intensity score multiplied by the
percentage score, was classified as follows: 0 for negative; 1–3
for weak; 4–6 for moderate; and 7–9 for strong. Samples with a
final score ≤3 were grouped together as PKMYT1-expression negative
while those with a score ≥4 were grouped together as
PKMYT1-expression positive.
Statistical analysis
Statistical analysis was conducted using SPSS 19.0
(IBM Corp., Inc., Armonk, NY, USA) program. The results of RT-qPCR,
migration and invasion were analyzed with Student's t-test. Hazard
ratio and 95% confidence interval of clinicopathological data were
evaluated using Cox regression models. Kaplan-Meier method was used
to analyse disease-free survival rate using the log-rank test. A
P-value of <0.05 was considered to indicate statistically
significant differences.
Results
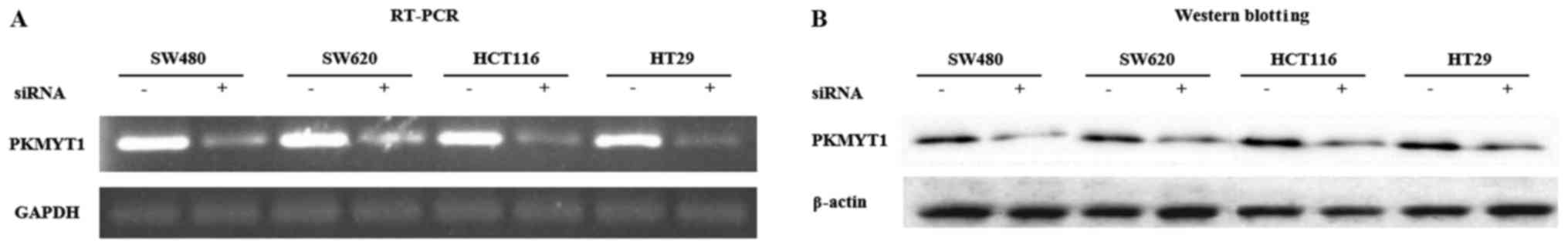
siRNA-mediated knockdown of
PKMYT1
Since it was revealed that PKYMT1 may be an
interesting target in an anticancer therapy (21,22),
we tried to determine whether PKMYT1 was functionally important in
CRC using siRNA. CRC cell lines including SW480, SW620, HCT116 and
HT29 were transduced with either control or PKMYT1 siRNA. To
determine the expression level of PKMYT1, RNA was extracted from
transduced cells. The expression level of PKMYT1 was significantly
lower in PKMYT1-inhibited cells by siRNA compared to
control-transduced cells (Fig. 1A).
Since mRNA expression is not always consistent with protein level,
we analyzed the level of PKMYT1 protein after siRNA-mediated PKMYT1
knockdown. Immunoblot analysis using cell lysates obtained from
siRNA-transduced cells revealed that PKMYT1 siRNA-transduced CRC
cell lines expressed much lower level of PKMYT1 protein compared to
the control cells (Fig. 1B). From
these data, we confirmed that the expression of PKMYT1 was
significantly inhibited by siRNA-mediated depletion at both the RNA
and protein level.
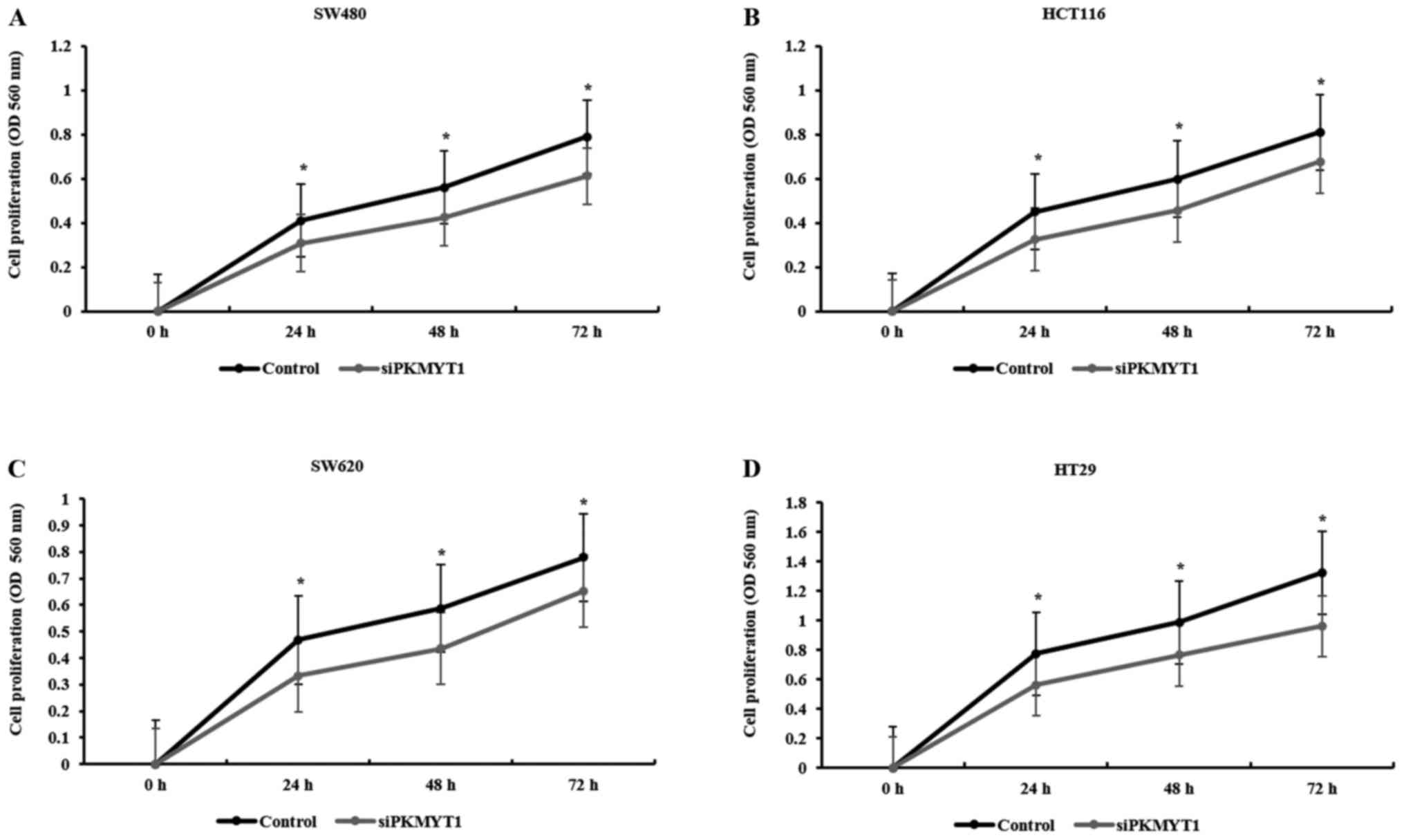
The proliferation and colony-forming
ability of CRC cell lines are defective by the knockdown of
PKMYT1
We tested whether PKMYT1 was implicated in the
growth of CRC cells using MTT assay after PKMYT1 knockdown. The
proliferation rate of control or PKMYT1 siRNA-transfected SW480,
HCT116, SW620 and HT29 cell lines was analyzed every 24 h for 3
days. The proliferation rate of PKMYT1-depleted CRC cells was
significantly decreased starting from 24 h up to 72 h in the tested
CRC cell lines (Fig. 2). To check
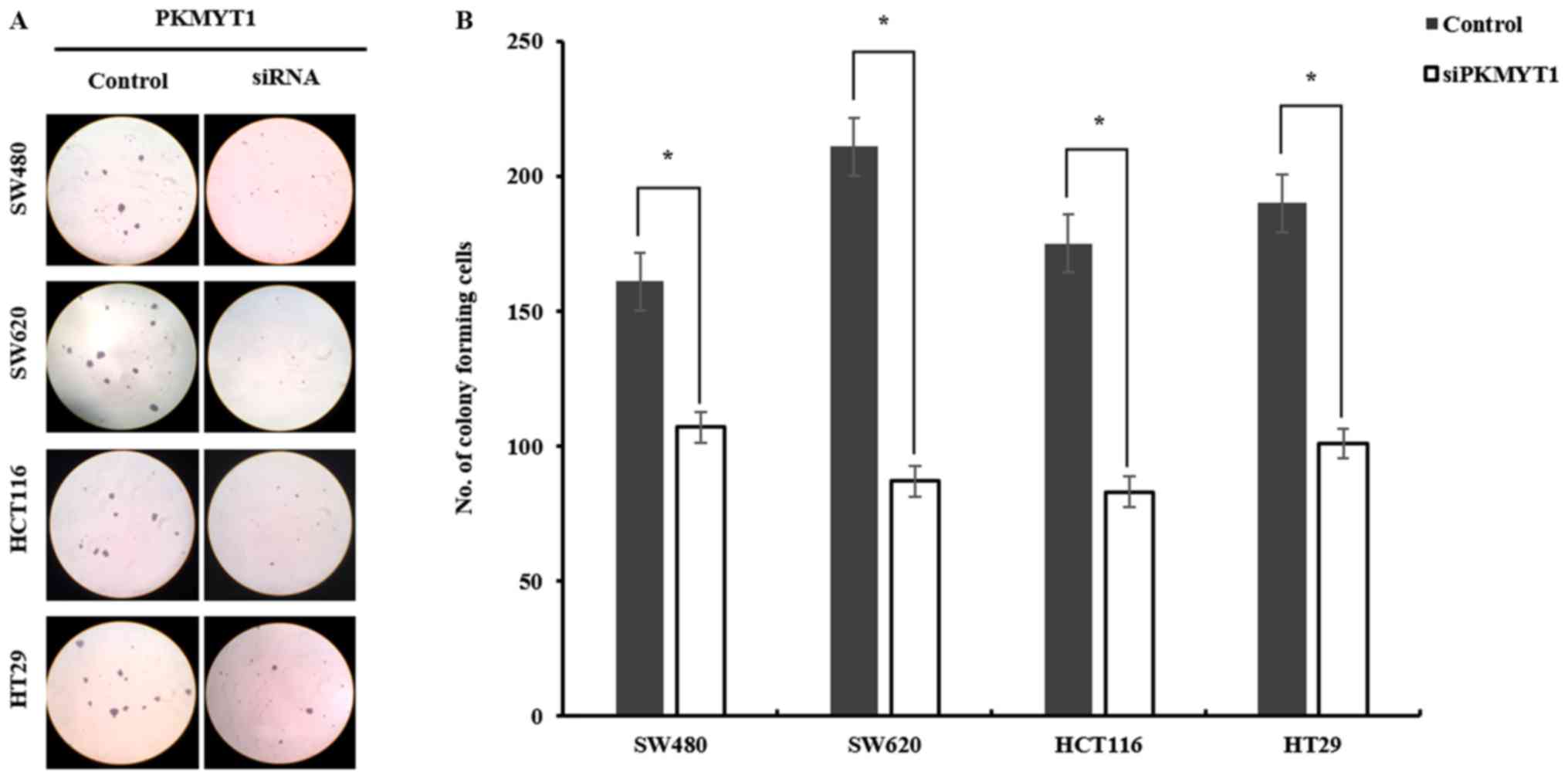
the anchorage-independent growth of CRC cell lines after knocking
down PKMYT1, semi-solid soft-agar colony forming ability was
tested. Control or PKMYT1 siRNA-transfected CRC cell lines were
seeded on soft-agar, and colony number and size were determined.
The colony size in PKMYT1-knockdown cells was much smaller than
that of control cells (Fig. 3).
Notably, as displayed in Fig. 3,
the number of colonies was significantly decreased in PKMYT1-siRNA
transfected CRC cell lines compared to the control (SW480, 161 vs.
106; SW620, 211 vs. 87; HCT116, 175 vs. 83; HT29, 190 vs. 101).
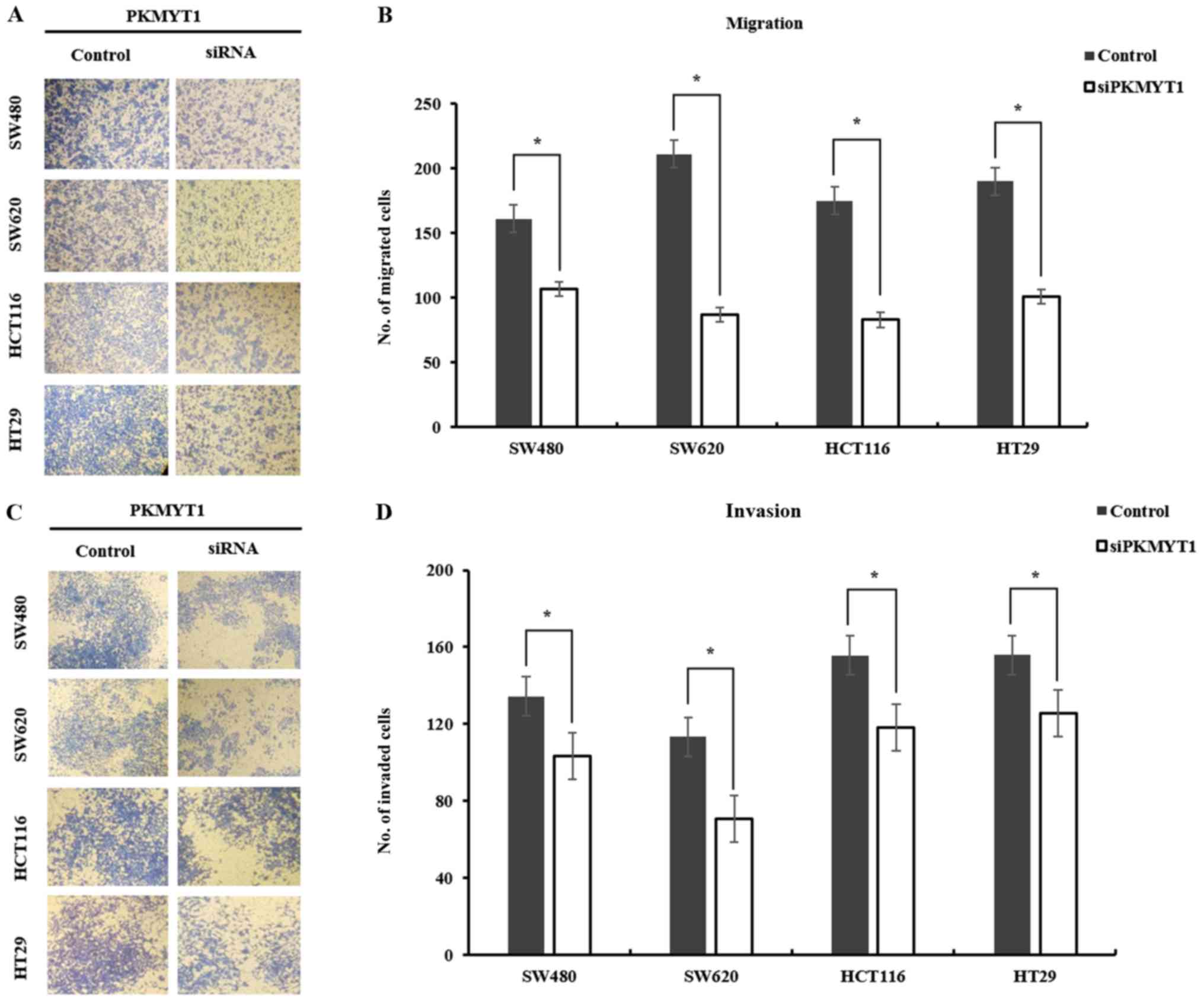
PKMYT1 is required for the mobility of
CRC cell lines
We examined the mobility change after knocking down
PKMYT1 in CRC cells to determine whether PKMYT1 was essential in
the progression of CRC. CRC cell lines SW480, SW620, HCT116 and
HT29 were transfected with either control or PKMYT1 siRNA.
Transduced cells were seeded on the Transwell system and incubated
for 48 h to assess the migratory ability. The migration of cancer
cells was evaluated by counting the cells on the inserts of the
Transwell system after staining. PKMYT1 siRNA-transfected cells
migrated ~14–38% less than control-transduced cells (Fig. 4A and B). To examine whether PKMYT1
was essential for the invasion of cancer cells in vitro,
control or PKMYT1-knockdown cells were plated on a Matrigel-coated
insert of a Transwell system and incubated for 48 h. PKMYT1
siRNA-transfected cells exhibited an invasion ability ~25–40% less
than the control-transduced cells (Fig.
4C and D). These results indicated that PKMYT1 is indispensable
for the mobility of CRC cells, thus possibly contributing to the
progression of CRC.
The association of PKMYT1 with a poor
prognosis of CRC patients
Following the characterization of the pivotal
function of PKMYT1 in CRC cell lines in vitro, we determined
the biological relevance of PKMYT1 to a patient prognosis model.
The demographic data of the patients analyzed in the present study
are provided in Table II. When the
correlation of PKMYT1 expression to overall survival (OS) rate was
analyzed, we observed that PKMYT1 was closely associated with the
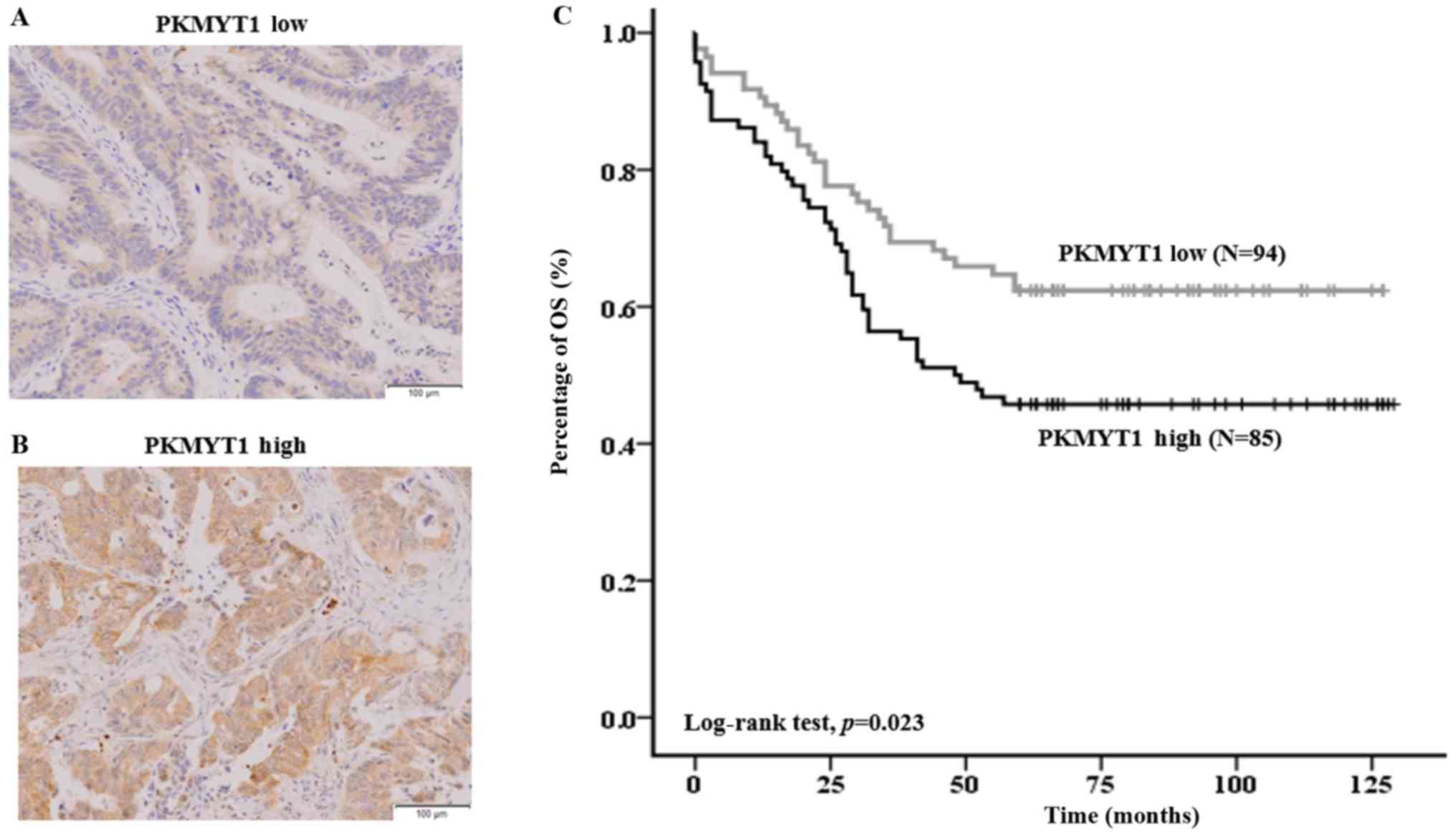
OS rate of CRC patients (Fig. 5).
Specifically, patients with a high level of PKMYT1 expression had
significantly lower overall survival (OS) rate than patients with
low expression of PKMYT1 (45.7 vs. 62.4%, P=0.023; Fig. 5C). In addition, PKMYT1 positivity
was correlated with other clinicopathological variables by Cox
regression analysis including lymphatic invasion, perineural
invasion and clinical stage (Table
III). These results indicated the close association of PKMYT1
with poor prognosis of CRC patients.
| Table III.Univariate and multivariate Cox
regression analysis of the relative risk of death according to the
expression of PKMYT1. |
Table III.
Univariate and multivariate Cox
regression analysis of the relative risk of death according to the
expression of PKMYT1.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Clinicopathological
factors | Hazard ratio (95%
Cl) | P-value | Hazard ratio (95%
Cl) | P-value |
|---|
| Age, years (<60,
≥60) | 1.158
(0.721–1.862) | 0.544 | 1.261
(0.768–2.070) | 0.359 |
| Sex | 1.154
(0.749–1.777) | 0.516 | 1.095
(0.702–1.708) | 0.688 |
| Vascular
invasion | 1.503
(0.908–2.488) | 0.113 | 0.817
(0.396–1.686) | 0.584 |
| Lymphatic
invasion | 2.055
(1.311–3.221) | 0.002 | 1.821
(0.923–3.590) | 0.084 |
| Perineural
invasion | 4.219
(2.158–8.245) | 0.000 | 3.496
(1.701–7.186) | 0.001 |
| Clinical stage (I,
II, III, IV) | 1.817
(1.172–2.817) | 0.008 | 1.720
(1.084–2.727) | 0.021 |
| PKMYT1 | 1.657
(1.064–2.579) | 0.025 | 1.594
(1.020–2.492) | 0.041 |
Discussion
In the present study, we found that PKMYT1 was
essential for the proliferation and mobility of CRC cells in
vitro. Depletion of the expression of PKMYT1 by siRNA resulted
in impaired proliferation, anchorage-independent colony forming
ability and mobility of CRC cells. In addition, using
patient-derived samples, we demonstrated that patients expressing a
high level of PKMYT1 displayed a worse overall survival rate than
those with a low level of PKMYT1. Thus, these results indicated
that PKMYT1 was a biomarker for the prediction of the prognosis of
the disease. In agreement with our findings, several other studies
have also recently reported that PKMYT1 was essential in several
cancers such as glioblastoma, however not in normal cells and a
promising target in developing chemotherapeutics (21,22).
It is well established that CDK1 is a key molecule
in the progression of mitosis and becomes inactivated when
phosphorylated by WEE1 (23,24).
Although CDK1 is also phosphorylated by PKMYT1, the contribution of
PKMYT1 in normal or cancer cells without DNA damage appears
relatively minor. Chow et al (21) revealed that the cell cycle or
mitosis timing in HeLa cells was not affected by the depletion of
PKMYT1 without DNA damage. However, PKMYT1 knockdown with DNA
damaging agents such as adriamycin or irradiation was able to
significantly inhibit cell survival (21). In human neural stem cells, it was
demonstrated that PKMYT1 and WEE1 are redundant. However, in
glioblastoma cells, depletion of PKMYT1 led to a dramatic increase
in cell death and cytokinesis failure (22). Previously, only WEE1 had been
studied as a potential therapeutic target in chemotherapy since
PKMYT was considered not essential or redundant. However, our data
along with other studies indicated that PKMYT1 was essentially
required in several cancers including CRC. Furthermore, it is the
first study, to the best of our knowledge, to demonstrate that
PKMYT1 was implicated in the progression of CRC.
It has been suggested that metastatic spread of CRC
is one of the pivotal factors leading to a high level of mortality
in CRC patients. Thus, to improve the survival rates of CRC
patients, it is required to identify essential signaling pathways
implicated in metastasis (25,26).
Epithelial-mesenchymal transition (EMT), an important process in
tumor progression, is involved in several aspects including
invasion, metastasis and drug resistance (27–34).
In hepatocellular carcinoma cells, PKMYT1 has been shown to
regulate EMT by activating beta-catenin/TCF signaling (35). In the present study, we also found
that depletion of PKMYT1 impaired the mobility of CRC cells, a
function that is related to EMT, proposing that PKMYT1 was an
essential molecule in the progression of CRC. Thus, developing
small molecules that block the function of PKMYT1 could be an
effective way to inhibit the progression of the disease.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Soonchunhyang
University Research Fund and the grant of the Korea Health
Technology R&D Project through the Korea Health Industry
Development Institute (KHID), funded by the Ministry of Health
& Welfare, Republic of Korea (grant no. HI15C1647).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
DJ, HYK and MJB conceived and designed the study.
DJ, HK, DK, SB, SO, HL, DK, TSA, SBB, MSL and SJ performed the
experiments. HYK and MJB wrote the paper. HJK and CJK reviewed and
edited the manuscript. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Ethics Committee of Soonchunhyang University Cheonan Hospital and
Soonchunhyang University Institutional Animal Care and Use
Committee.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of
interest.
References
|
1
|
Van Roosbroeck S, Hoeck S and Van Hal G:
Population-based screening for colorectal cancer using an
immunochemical faecal occult blood test: A comparison of two
invitation strategies. Cancer Epidemiol. 36:e317–e324. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Elias E, Mukherji D, Faraj W, Khalife M,
Dimassi H, Eloubeidi M, Hattoum H, Abou-Alfa GK, Saleh A and
Shamseddine A: Lymph-node ratio is an independent prognostic factor
in patients with stage III colorectal cancer: A retrospective study
from the Middle East. World J Surg Oncol. 10:632012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Deschoolmeester V, Baay M, Specenier P,
Lardon F and Vermorken JB: A review of the most promising
biomarkers in colorectal cancer: One step closer to targeted
therapy. Oncologist. 15:699–731. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jung KW, Won YJ, Kong HJ, Oh CM, Seo HG
and Lee JS: Cancer statistics in Korea: Incidence, mortality,
survival and prevalence in 2010. Cancer Res Treat. 45:1–14. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Michor F, Iwasa Y, Lengauer C and Nowak
MA: Dynamics of colorectal cancer. Semin Cancer Biol. 15:484–493.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Satram-Hoang S, Lee L, Yu S, Guduru SR,
Gunuganti AR, Reyes C and McKenna E: Comparative effectiveness of
chemotherapy in elderly patients with metastatic colorectal cancer.
J Gastrointest Cancer. 44:79–88. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tol J and Punt CJ: Monoclonal antibodies
in the treatment of metastatic colorectal cancer: A review. Clin
Ther. 32:437–453. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang L, Chen X, Li W and Sheng Z:
Antiepidermal growth factor receptor monoclonal antibody improves
survival outcomes in the treatment of patients with metastatic
colorectal cancer. Anticancer Drugs. 23:155–160. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wilke HJ and Van Cutsem E: Current
treatments and future perspectives in colorectal and gastric
cancer. Ann Oncol. 14 Suppl 2:ii49–ii55. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chan KM, Wu TH, Cheng CH, Lee WC, Chiang
JM, Chen JS and Wang JY: Prognostic significance of the number of
tumors and aggressive surgical approach in colorectal cancer
hepatic metastasis. World J Surg Oncol. 12:1552014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Swiderska M, Choromanska B, Dabrowska E,
Konarzewska-Duchnowska E, Choromańska K, Szczurko G, Myśliwiec P,
Dadan J, Ladny JR and Zwierz K: The diagnostics of colorectal
cancer. Contemp Oncol. 18:1–6. 2014.
|
|
13
|
Feigelson HS, Zeng C, Pawloski PA, Onitilo
AA, Richards CS, Johnson MA, Kauffman TL, Webster J, Nyirenda C,
Alexander GL, et al: Does KRAS testing in metastatic
colorectal cancer impact overall survival? A comparative
effectiveness study in a population-based sample. PLoS One.
9:e949772014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schee K, Lorenz S, Worren MM, Günther CC,
Holden M, Hovig E, Fodstad O, Meza-Zepeda LA and Flatmark K: Deep
sequencing the MicroRNA transcriptome in colorectal cancer. PLoS
One. 8:e661652013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elledge SJ: Cell cycle checkpoints:
Preventing an identity crisis. Science. 274:1664–1672. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rhind N and Russell P: Signaling pathways
that regulate cell division. Cold Spring Harb Perspect Biol.
4:pii:a0059422012. View Article : Google Scholar
|
|
17
|
Norbury C, Blow J and Nurse P: Regulatory
phosphorylation of the p34cdc2 protein kinase in vertebrates. EMBO
J. 10:3321–3329. 1991.PubMed/NCBI
|
|
18
|
Mueller PR, Coleman TR, Kumagai A and
Dunphy WG: Myt1: A membrane-associated inhibitory kinase that
phosphorylates Cdc2 on both threonine-14 and tyrosine-15. Science.
270:86–90. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu F, Stanton JJ, Wu Z and Piwnica-Worms
H: The human Myt1 kinase preferentially phosphorylates Cdc2 on
threonine 14 and localizes to the endoplasmic reticulum and Golgi
complex. Mol Cell Biol. 17:571–583. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Choi HS, Bode AM, Shim JH, Lee SY and Dong
Z: c-Jun N-terminal kinase 1 phosphorylates Myt1 to prevent
UVA-induced skin cancer. Mol Cell Biol. 29:2168–2180. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chow JP and Poon RY: The CDK1 inhibitory
kinase MYT1 in DNA damage checkpoint recovery. Oncogene.
32:4778–4788. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Toledo CM, Ding Y, Hoellerbauer P, Davis
RJ, Basom R, Girard EJ, Lee E, Corrin P, Hart T, Bolouri H, et al:
Genome-wide CRISPR-Cas9 screens reveal loss of redundancy between
PKMYT1 and WEE1 in glioblastoma stem-like cells. Cell Rep.
13:2425–2439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Parker LL and Piwnica-Worms H:
Inactivation of the p34cdc2-cyclin B complex by the human WEE1
tyrosine kinase. Science. 257:1955–1957. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Watanabe N, Broome M and Hunter T:
Regulation of the human WEE1Hu CDK tyrosine 15-kinase during the
cell cycle. EMBO J. 14:1878–1891. 1995.PubMed/NCBI
|
|
25
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee JJ and Lotze MT: Molecular basis of
metastasis. N Engl J Med. 360:1679–1680. 2009.PubMed/NCBI
|
|
27
|
Frisch SM, Schaller M and Cieply B:
Mechanisms that link the oncogenic epithelial-mesenchymal
transition to suppression of anoikis. J Cell Sci. 126:21–29. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang RY, Chung VY and Thiery JP:
Targeting pathways contributing to epithelial-mesenchymal
transition (EMT) in epithelial ovarian cancer. Curr Drug Targets.
13:1649–1653. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jordan NV, Johnson GL and Abell AN:
Tracking the intermediate stages of epithelial-mesenchymal
transition in epithelial stem cells and cancer. Cell Cycle.
10:2865–2873. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee K and Nelson CM: New insights into the
regulation of epithelial-mesenchymal transition and tissue
fibrosis. Int Rev Cell Mol Biol. 294:171–221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tam WL and Weinberg RA: The epigenetics of
epithelial-mesenchymal plasticity in cancer. Nat Med. 19:1438–1449.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: New insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu L, Wu J, Wang S, Luo X, Du Y, Huang D,
Gu D and Zhang F: PKMYT1 promoted the growth and motility of
hepatocellular carcinoma cells by activating beta-catenin/TCF
signaling. Exp Cell Res. 358:209–216. 2017. View Article : Google Scholar : PubMed/NCBI
|