Introduction
Hepatocellular carcinoma (HCC) is the third leading
cause of cancer-related death cancer worldwide. The incidence and
mortality of HCC are increasing in Asian countries as a result of
an ageing cohort infected with hepatitis B virus (HBV), and these
parameters are expected to continue to rise as a consequence of the
obesity epidemic or exposure to a contaminated environment
(1). Most HCC patients typically
present in advanced and incurable stages, and are usually
unsuitable for surgery. However, this situation is improving with a
better understanding of the molecular mechanisms involved in the
progression of HCC (2). Recently,
researchers have focused on plant-derived compounds that have the
potential to target the molecules which are critical for the
progression of HCC (3).
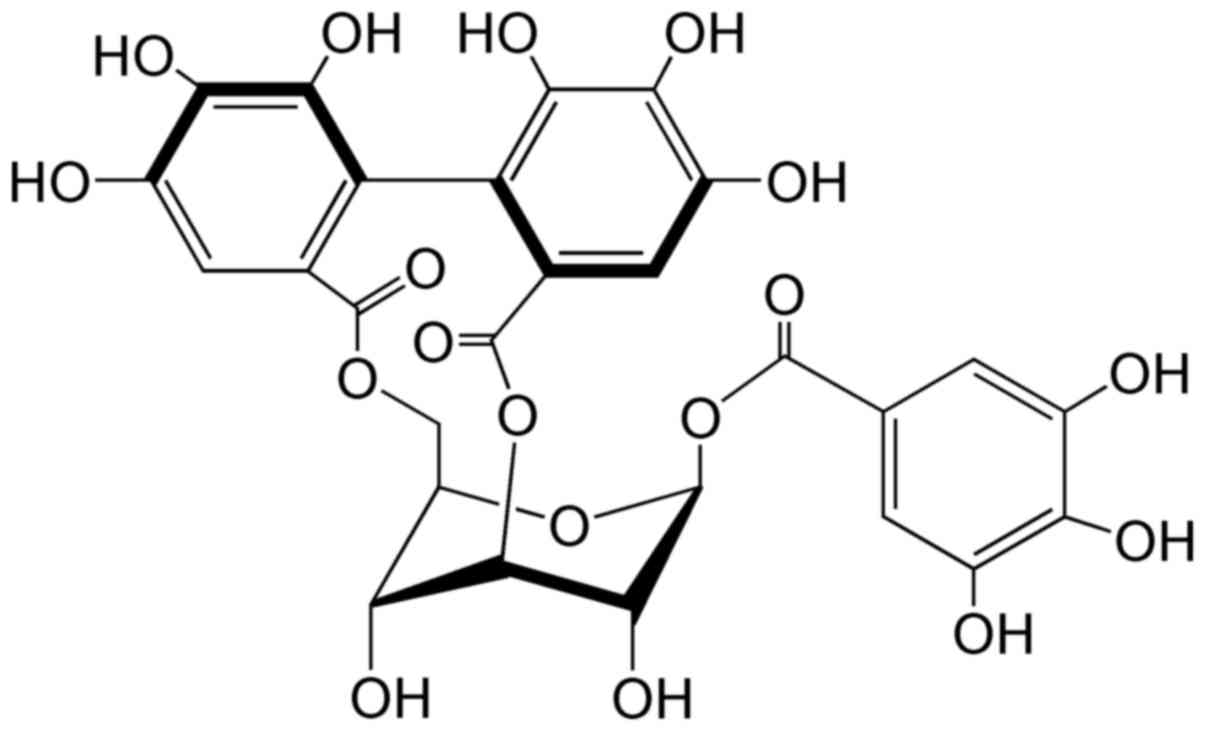
Corilagin (Fig. 1),
a natural plant polyphenol tannic acid, is a major active component
of many ethnopharmacological plants such as Phyllanthus (P.)
niruri L., P. emblica L. and P. urinaria L. It
was first isolated from Caesalpinia coriaria (Jacq.) Willd.
(Divi-divi) by Schmidt in 1951 (4).
In recent years, corilagin has provoked much attention due to its
antitumor, hepatoprotective, and anti-inflammatory activities, but
particularly as a candidate antitumor agent. In the past few
decades, corilagin has been reported to display various
pharmacological activities, including antioxidant activity
(5), hepatoprotective effect
(6), anti-inflammatory activity
(7), neuroprotective effect
(8), cardiovascular protective
activity (9) and anti-diabetic
activity (10). Recently, the
antitumor effect of corilagin (2,3,11,12)
has been a focus of increased research.
Research on the antitumor activity of corilagin
began in 1985. Corilagin was reported to inhibit the reverse
transcriptase activity of RNA tumor virus (13), to inhibit the topoisomerase
I-mediated relaxation of DNA, which inhibits the growth of tumor
cells (14,15), and to reduce the release of tumor
necrosis factor-α (TNF-α) in cancer cells. TNF-α is thought to
stimulate the growth and progression of early malignant tumor cells
(16,17). In recent years, corilagin has been
demonstrated to suppress human colon cancer, gastric adenocarcinoma
and metrocarcinoma (18–20), to inhibit the growth of tumor cells
in vivo, to suppress the proliferation of HCC Hep3B cells in
an athymic nude mouse xenograft model (11), to suppress the growth of HCC cells
by inducing G2/M phase arrest and to inhibit the proliferation of
ovarian cancer cells via the TGF-β/AKT/ERK signaling pathways
(2,3). In addition, corilagin was found to
inhibit the proliferation of cholangiocarcinoma (CCA) cells by
regulating the Notch signaling pathway. In vitro, corilagin
inhibited CCA cell proliferation, migration and invasion, and
induced the apoptosis of CCA cells by inhibiting the Notch
signaling pathway. Moreover, corilagin has shown the potential to
promote the antitumor activity of cisplatin and doxorubicin in HCC
cells (11,21) and to sensitize ovarian cancer cells
to paclitaxel and carboplatin by inhibiting the Snail-glycolysis
pathways (22).
Based on the above findings, corilagin induces the
apoptosis of HCC cells, yet the mechanism remains unclear.
Therefore, the present study aimed to explore the molecular
mechanisms involved in the pro-apoptotic effect of corilagin and to
provide evidence that corilagin may have the potential to become a
complementary anticancer herbal drug for HCC therapy.
Materials and methods
Chemicals and reagents
Corilagin (purity ≥98%) was prepared at the Xiamen
Overseas Chinese Subtropical Plant Introduction Garden (Xiamen,
China) as previously described (21). Three HCC cell lines (SMMC-7721,
Bel-l7402 and MHCC97-H) were used and all were obtained from the
Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
RPMI-1640 medium and Dulbecco's modified Eagle's medium (DMEM) were
obtained from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Bisbenzimide (Hoechst 33258), acridine orange (AO), JC-1,
ethidium bromide (EB), methyl thiazolyl tetrazolium (MTT) and
cisplatin (CDDP; purity ≥99.9%) were obtained from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). Annexin V-FITC/PI Apoptosis
Detection kit was obtained from Roche (Mannheim, Germany). The
antibodies against p53 (1:1,000; mouse mAb; cat. no. 48818), Bcl-2
(1:1,000; mouse mAb; cat. no. 15071), cytochrome c (1:1,000;
rabbit mAb; cat. no. 11940), caspase-9 (1:1,000; mouse mAb; cat.
no. 9508), caspase-8 (1:1,000; mouse mAb; cat. no. 9746), caspase-3
(1:1,000; rabbit mAb; cat. no. 9662), PARP (1:1,000; rabbit mAb;
cat. no. 9746), surviving (1:1,000; mouse mAb, cat. no. 2802), Fas
(1:1,000; mouse mAb; cat. no. 8023) and FasL (1:1,000; rabbit mAb;
cat. no. 4273) were purchased from Cell Signaling Technology
(Danvers, MA, USA).
Cell culture
Cells were cultured in RPMI-1640 or DMEM
supplemented with 10% FBS, penicillin (100 U/ml) and streptomycin
(100 mg/ml) in a humidified incubator aerated with 5%
CO2 and 95% air at 37°C. When the cells reached 70–80%
confluency, they were trypsinized and counted, and 5,000 cells were
seeded into each 96-well microtiter plate for overnight incubation
and then treated with corilagin complete cell medium for 48 h. The
control group was treated with DMSO (0.1%, w/v) for the same
duration.
MTT assay
The effect of the constituents on the growth of the
HCC cells was determined using the MTT assay. Briefly, the cells
were seeded overnight and the medium was removed. The cells were
then treated with 180 ml corilagin dissolved in medium at 6.25,
12.5, 25, 50, 75 and 100 µM or with DMSO (0.1%, w/v) as a control
for 24 h. After 24 h, 20 µl MTT (5 mg/ml) was added to each well,
and the plates were incubated for an additional 4 h at 37°C. The
supernatants were replaced with 150 ml of DMSO to dissolve the
formazan produced from the MTT by the metabolizing cells.
Absorbance at 492 nm was proportional to the live metabolizing cell
count, and cell survival was expressed as the absorbance of the
MTT-treated cells relative to that of the DMSO-treated controls
(absorbance was detected using Thermo Scientific Microplate Reader
MK3).
Colony formation assay
Cells were seeded at 800 cells per well in a 6-well
plate (9.5 cm2) and allowed to attach overnight prior to
treatment with DMSO or varying concentrations of corilagin. The
cells were treated for 24 h, then recovery medium was added and the
cells were allowed to continue to grow for 10 days. Colonies were
stained using 0.5% crystal violet solution in methanol for 30 min.
Staining solution was removed, wells were washed with deionized
H2O, and the stained colonies containing more than 50
cells were imaged and counted under microscopy (Leica Microsystems
GmbH, Wetzlar, Germany).
Morphological studies using
fluorescence microscopy
The HCC cells seeded into a 6-well plate were
treated with 37.5 µM corilagin or DMSO (0.1%, w/v) as a negative
control and 30 µM CDDP as a positive control for 24 h. The
supernatants were then replaced by PBS containing 100 µg/ml of
Hoechst 33258 or AO/EB in PBS for 10 min in the dark. Cell
morphology was visualized immediately using fluorescence microscopy
(Leica Microsystems GmbH).
Mitochondrial membrane potential
assay
Cells were treated with 12.5, 25, 37.5 and 50 µM
corilagin or DMSO (0.1%, w/v) as a negative control and 30 µM CDDP
as a positive control for 24 h. The cells were then incubated with
JC-1 (0.1 µg/ml) for 15 min. After washing with PBS, the cells were
suspended in PBS and observed with a fluorescence microscope as
previously described (23). The
green fluorescence from the JC-1 monomer and the red fluorescence
from the aggregated form of JC-1 were visualized.
Flow cytometry assay
SMMC-7721 cells were seeded into 60-mm plates
(1–2×105/plate) and incubated with corilagin (25, 37.5
and 50 µM) or DMSO (0.1%, w/v) as a control on the next day.
Control and treated cells were trypsinized after treatments for 24
h. Cells were collected in PBS and stained using the Annexin
V-FITC/PI Apoptosis Detection kit. The stained cells were analyzed
by flow cytometry.
Western blot analyses
Cells were seeded into 60-mm plates
(1–2×105/plate) and incubated with corilagin (12.5, 25
and 37.5 µM, which were below or approximately the IC50)
or with DMSO (0.1%, w/v) as a control on the next day for 24 h.
Cells were washed in PBS, suspended in ice-cold lysis buffer
containing 50 mM Tris-HCl (pH 7.4), 5 mM EDTA, 150 mM NaCl, 1%
Triton X-100, 1% sodium deoxycholate, 1 mM sodium, 20 mg/ml
aprotinin, 20 mg/ml leupeptin and 1 mM phenylmethylsulfonyl
fluoride (PMSF) and placed on ice for 5 min. The solution was
harvested then centrifugation was carried out at 12,000 × g for 20
min (4°C), after which the supernatants were collected and the
protein concentrations in the cell lysates were determined using
the BCA assay. Whole cell lysates were resolved by SDS
polyacrylamide gel electrophoresis and transferred onto
nitrocellulose membranes, which were probed with anti-β-actin.
Blots were then developed using ECL (Thermo Fisher Scientific,
Inc.).
Statistical analysis
All data subjected to statistical analysis was from
at least three independent experiments and are reported as the mean
± standard deviation. The criterion for statistical significance
was taken as P<0.05 using a two-tailed t-test, and the count
data were tested using Chi-square criterion comparing the frequency
of parameters. The analyses were performed using SPSS 15.0 software
(SPSS, Inc., Chicago, IL, USA).
Results
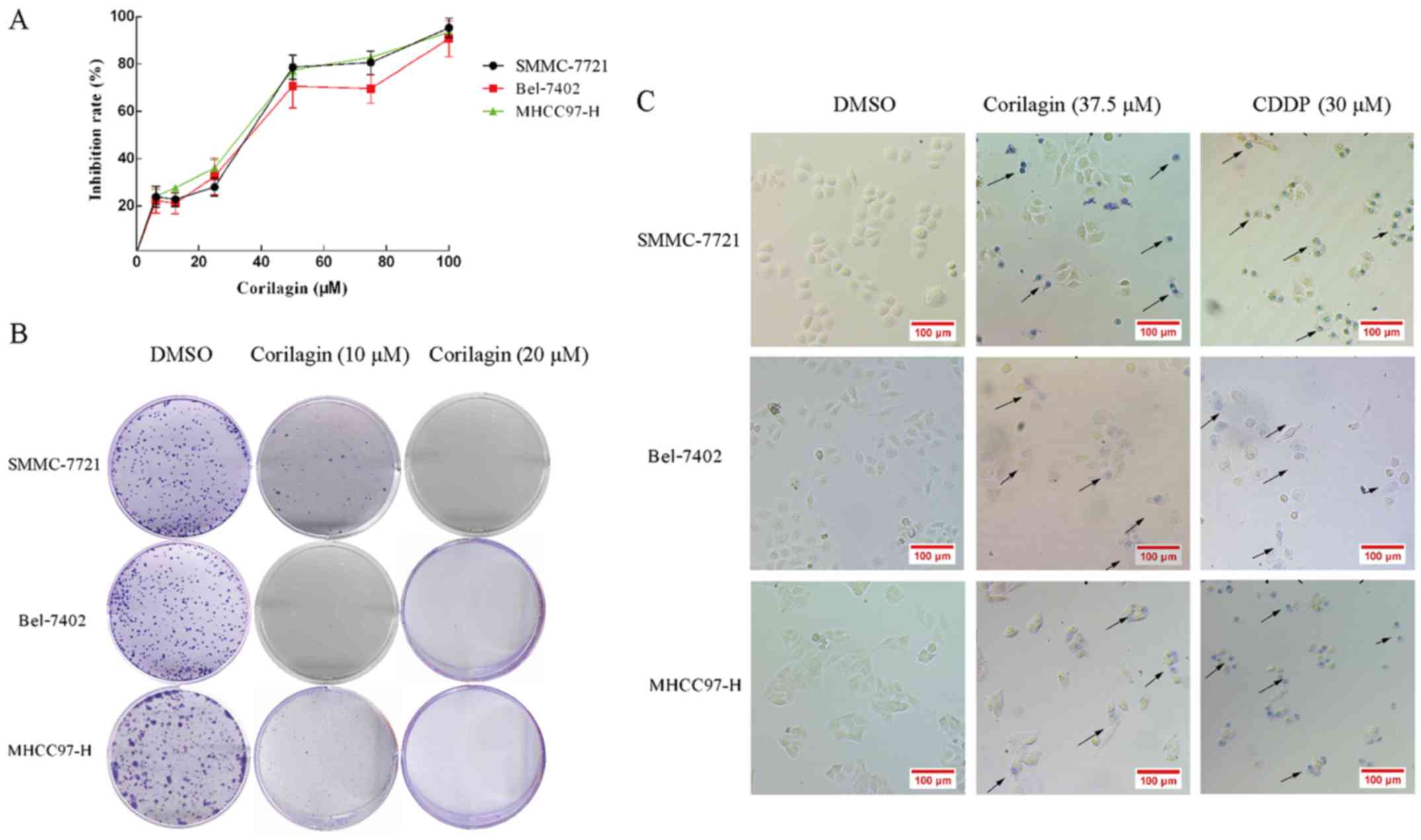
Corilagin inhibits the proliferation
and colony formation of hepatocellular carcinoma cells
The cytotoxicity of corilagin in HCC cells was
analyzed by the MTT assay. HCC cell lines, SMMC-7721, Bel-7402 and
MHCC97-H, were treated with increasing concentrations of corilagin
(6.25, 12.5, 25, 50, 75 and 100 µM) or with DMSO (0.1%, w/v) as a
control for 24 h. As shown in Fig.
2A, corilagin inhibited the proliferation of the three HCC cell
lines and the inhibitory rate exhibited a dose-dependent trend. The
IC50 values of corilagin against the SMMC-7721, Bel-7402
and MHCC97-H cells were 38.12±1.2, 39.7±1.4 and 37.05±0.9 µM,
respectively. In the colony formation assay, corilagin showed a
strong inhibitory effect on the HCC SMMC-7721 cells and HCC cell
lines Bel-7402 and MHCC97-H (Fig.
2B). Corilagin presented s similar inhibitory rate on the three
HCC cell lines. Based on the trypan blue staining assay (Fig. 2C), we observed trypan blue-stained
cells in the corilagin-treated group indicating that corilagin can
lead to the death of HCC SMMC-7721, Bel-7402 and MHCC97-H cells.
These results showed that corilagin significantly suppressed the
proliferation of the HCC cells.
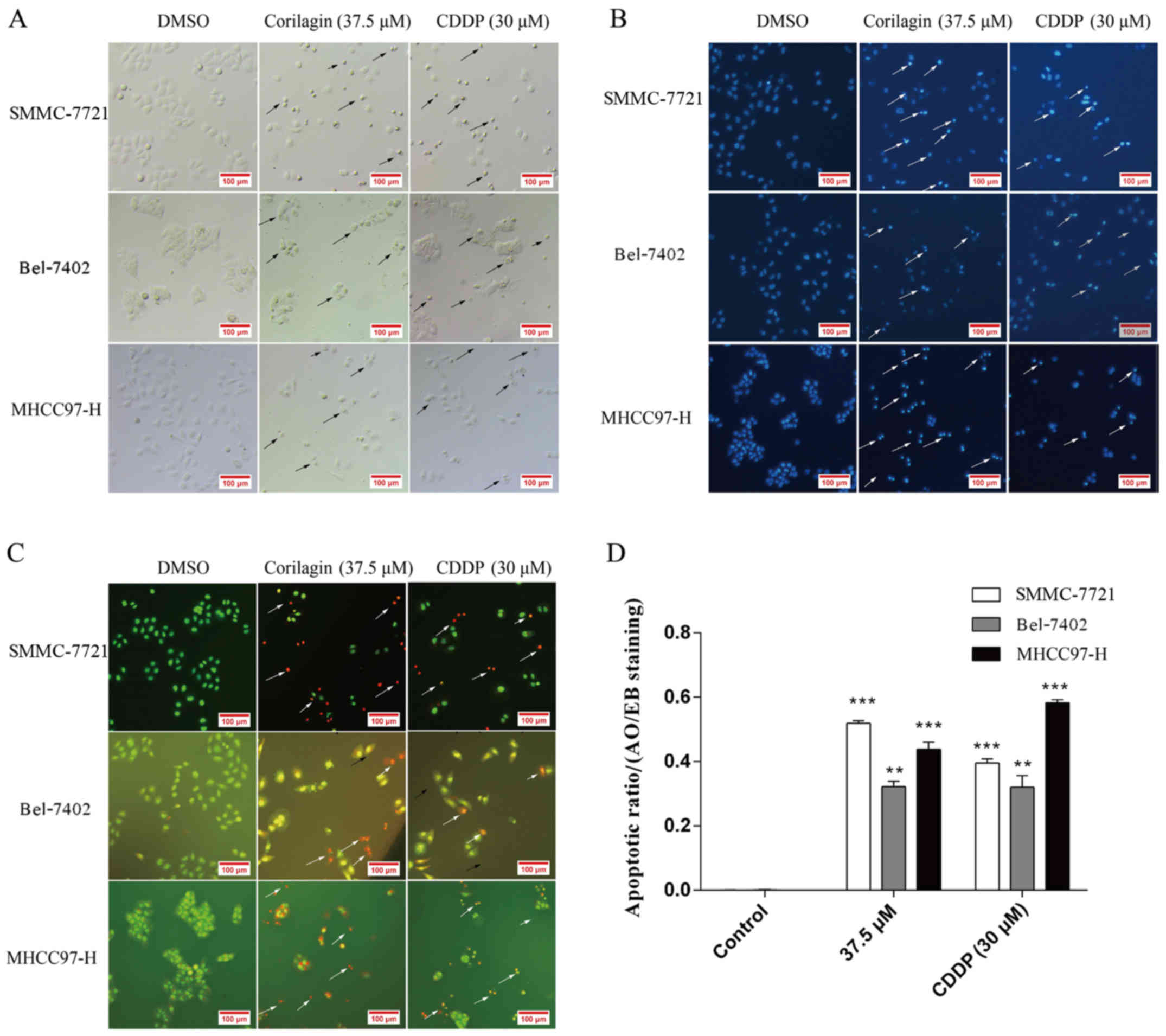
Observation of the morphological
changes in HCC cells after exposure to corilagin
Cell morphological changes after corilagin exposure
were observed under a microscope. As shown in Fig. 3A, cells treated with corilagin for
24 h showed typical features of apoptotic cells which were similar
to the features exhibited by the inhibitory effect of CDDP on the
HCC cells, such as nuclear fragmentation, cell shrinkage and
apoptotic bodies under the microscope. In addition, based on the
Hoechst 33258 staining assay, we found that the blue emission light
in apoptotic cells was much brighter than that in the control cells
(Fig. 3B). Upon AO/EB staining
assay, different cells (alive, apoptotic or necrotic) were clearly
differentiated by the different colors (Fig. 3C). Fig.
3D shows the quantification of 20 different images and
indicates the apoptotic ratios between the control and
corilagin-treated cells. These results indicated that the
corilagin-treated cells exhibited morphological changes indicating
apoptosis, including chromatin condensation and nuclear
fragmentation.
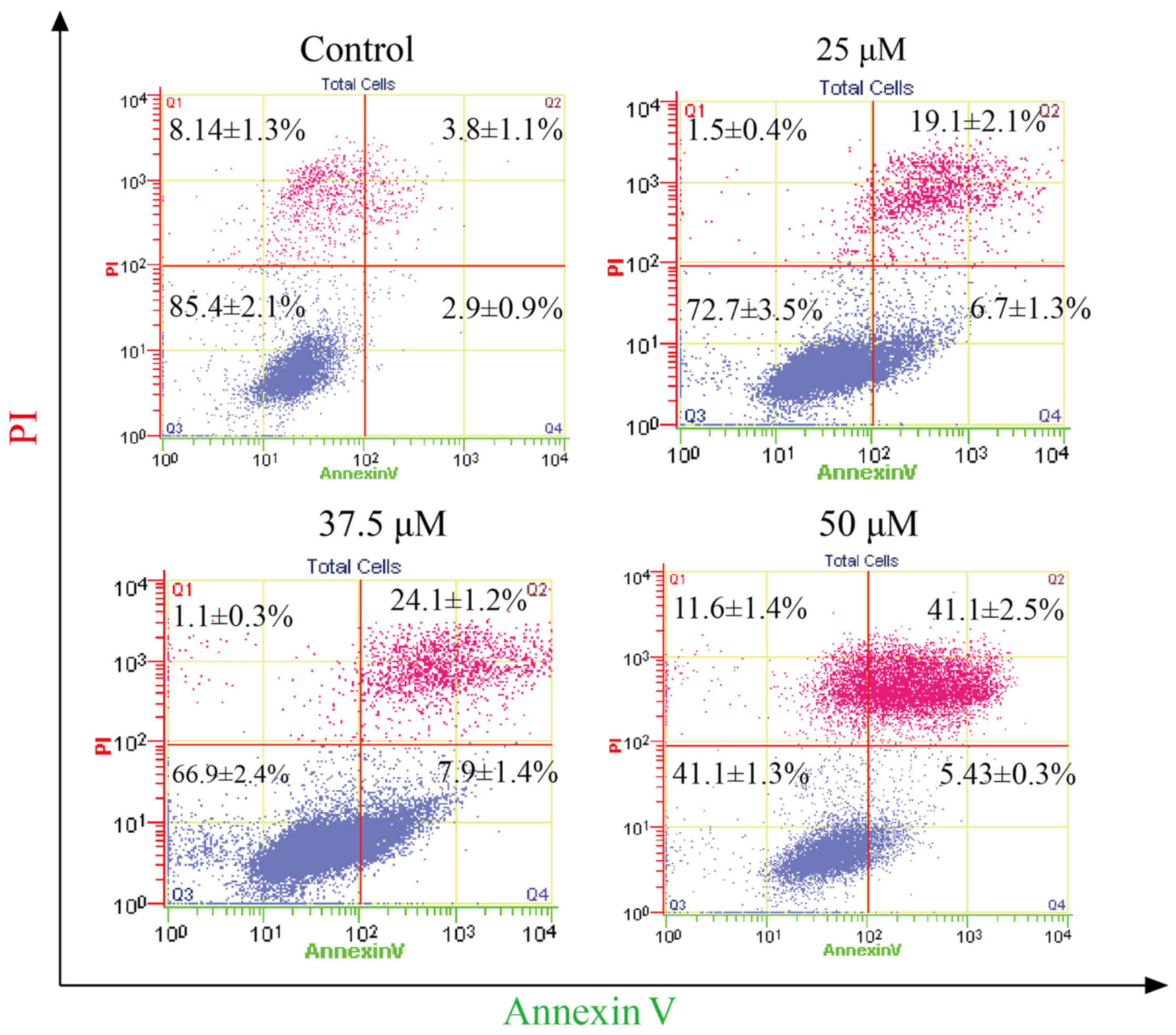
Effect of corilagin on the apoptosis
of HCC cells
To determine the apoptotic effect of corilagin on
HCC cells, SMMC-7721 cells were treated with different
concentrations of corilagin (25, 37.5 and 50 µM) for 24 h. Cells
were evaluated by flow cytometry through Annexin V/PI staining. As
shown in Fig. 4, increasing
percentages of apoptotic cells were observed when compared to the
control group. The apoptotic percentage of the HCC cells was 41.1%
following treatment with corilagin of 50 µM.
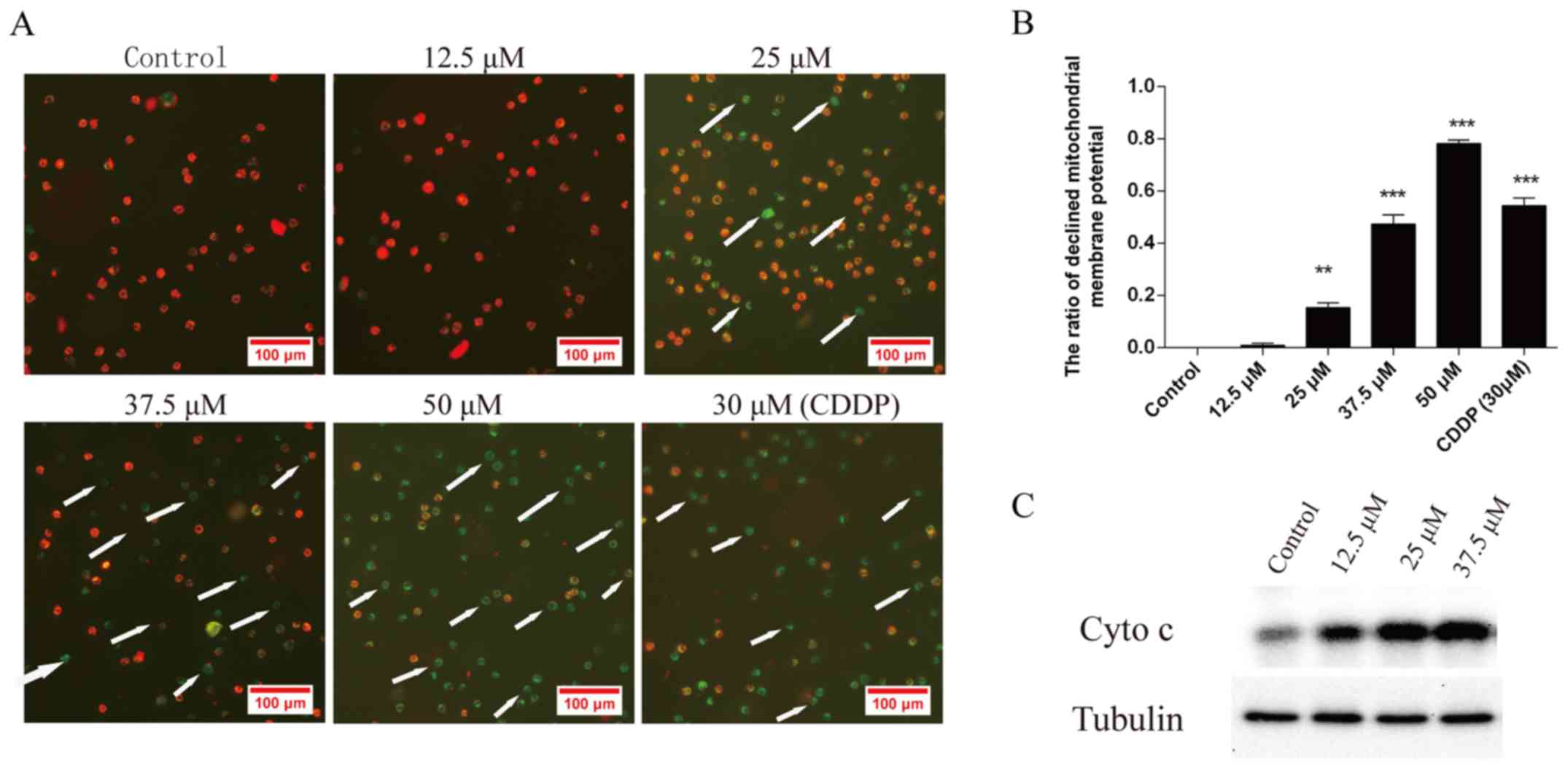
Effect of corilagin on the
mitochondrial transmembrane potential of HCC cells
To investigate the loss of Δψm during apoptosis
induced by corilagin, cells were treated with 12.5, 25, 37.5 and 50
µM corilagin or DMSO as a negative control and 30 µM CDDP as a
positive control for 24 h and then stained with JC-1 and monitored
with a fluorescence microscope. JC-1 forms monomer and emits green
fluorescence when Δψm is depolarized (common in apoptosis), while
JC-1 aggregates and emits red fluorescence at a highly polarized
Δψm. As shown in Fig. 5A, JC-1 was
accumulated in the control cells which presented red fluorescence
indicating a high Δψm. However, JC-1 was poorly accumulated in the
corilagin-treated cells, which displayed only green or weak red
fluorescence, indicating low Δψm in the mitochondria of the
corilagin-treated cells. Fig. 5B
shows the quantification of the ratio of decreased mitochondrial
membrane potential in the cells treated with corilagin. Moreover,
Fig. 5C shows that the protein
level of cytochrome c (Cyto c) in the cytoplasm was
increased following the treatment of corilagin which can lead to
the activation of caspase-9 and caspase-3. This result indicates
activation of the intrinsic apoptotic signaling pathways.
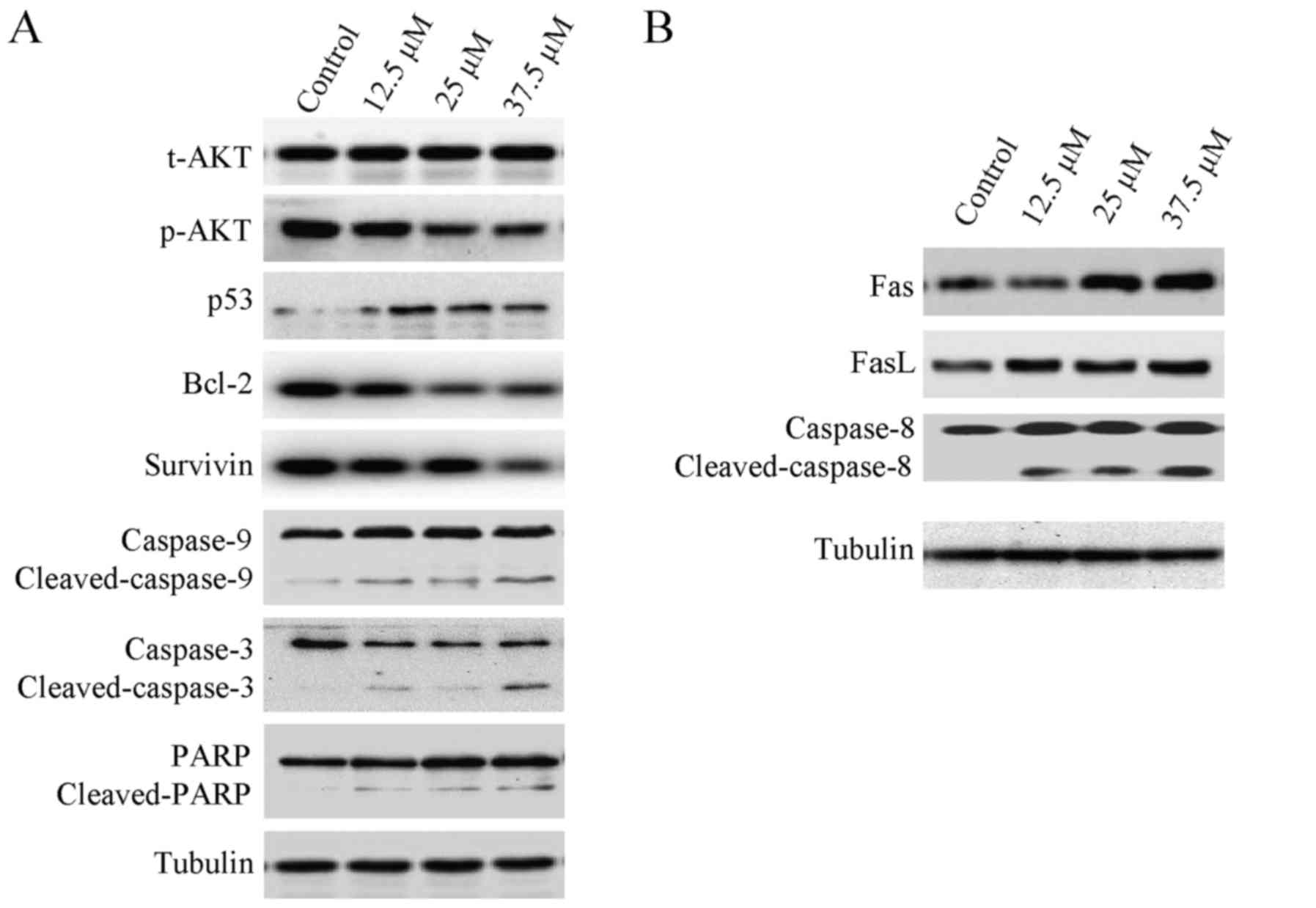
Effect of corilagin on the levels of
apoptosis-related proteins in HCC cells
Apoptosis is often relevant to a well-organized
sequence of cellular events, resulting in the alteration of
intracellular signaling pathways. HCC cells were treated with
different concentrations of corilagin (0, 12.5, 25 and 37.5 µM) for
24 h, and cytochrome c was translocated from the
mitochondria to the cytoplasm which confirmed the result of the
mitochondrial membrane potential assay. As shown in Fig. 6A, the expression of p-AKT was
downregulated, while the p53 protein level was upregulated
following treatment with corilagin. Cleavage of caspase-9,
caspase-3 and PARP was observed, indicating activation of the
intrinsic apoptotic signaling pathways (Fig. 6A). In addition, upregulation of Fas
and FasL was noted which led to the activation of caspase-8
(Fig. 6B); caspase-8 is an
important protein of the extrinsic apoptotic signaling pathways,
namely the Fas/FasL signaling pathway. These results indicate that
corilagin also activates the extrinsic signaling pathways. We also
observed downregulation of Bcl-2 and survivin which are important
anti-apoptotic proteins and may facilitate the inhibitory activity
of corilagin on HCC cells (Fig.
6A).
Discussion
Corilagin is the main bioactive component of many
traditional herbal plants (e.g., P. niruri L. and P.
emblica L.). Corilagin was found to inhibit the growth of many
types of cancer cells including nasopharyngeal carcinoma (19), HCC (2), gastric adenocarcinoma (19), lung adenocarcinoma (20), metrocarcinoma (18), colon cancer (19), ovarian cancer (3) and osteosarcoma cells (3) of human origin and murine melanoma
cells (18).
In a previous study, our group found that corilagin
inhibited the growth of HCC cells by inducing G2/M phase arrest by
downregulating p-Akt and cyclin B1/cdc2 and upregulating p-p53 and
p21 (2) and corilagin inhibited
ovarian cancer cells via the TGF-β/Akt/ERK signaling pathways
(3). Moreover, corilagin was
reported to enhance the antitumor activity of CDDP in HCC cells
(21). In the present study, we
found that corilagin-treated cells exhibited morphological changes
indicative of apoptosis, including chromatin condensation and
nuclear fragmentation through Hoechst 33258 and AO/EB staining
assays demonstrating that corilagin can induce the apoptosis of HCC
cells. The result of the flow cytometry assay was in accordance
with the observation of morphological change; corilagin does induce
the apoptosis of HCC cells.
At the molecular level, we also observed a change in
the mitochondrial membrane potential in the corilagin-treated HCC
cells. This result confirmed the work of Wang who found that
corilagin induced membrane potential change in the mitochondria of
human gastric cancer SGC-7901 cells (12). In the present study, cytochrome
c was translocated from mitochondria to the cytoplasm which
confirmed the result of the mitochondrial membrane potential assay.
Cleavage of caspase-9, caspase-3 and PARP was observed
demonstrating that corilagin can activate the intrinsic apoptotic
signaling pathways, namely the mitochondrial pathway. In the
western blot assay of corilagin-treated HCC cells, we observed
activation of caspase-8, which is downstream of the death receptor
pathway (Fas/FasL signaling pathway) and upregulation of Fas/FasL
(Fig. 6B). Activation of caspase-8
indicated that corilagin can also activate the death receptor
pathway. In addition, we also observed downregulation of Bcl-2 and
survivin and upregulation of p53. Bcl-2 is an important
anti-apoptotic protein which can block the release of cytochrome
c from mitochondria to the cytoplasm (24). Survivin is also an important
anti-apoptotic protein which can inhibit the activity of caspase-3
(25). p53 is an important
pro-apoptotic protein which can promote the release of cytochrome
c and activation of the intrinsic apoptotic signaling
pathways (26).
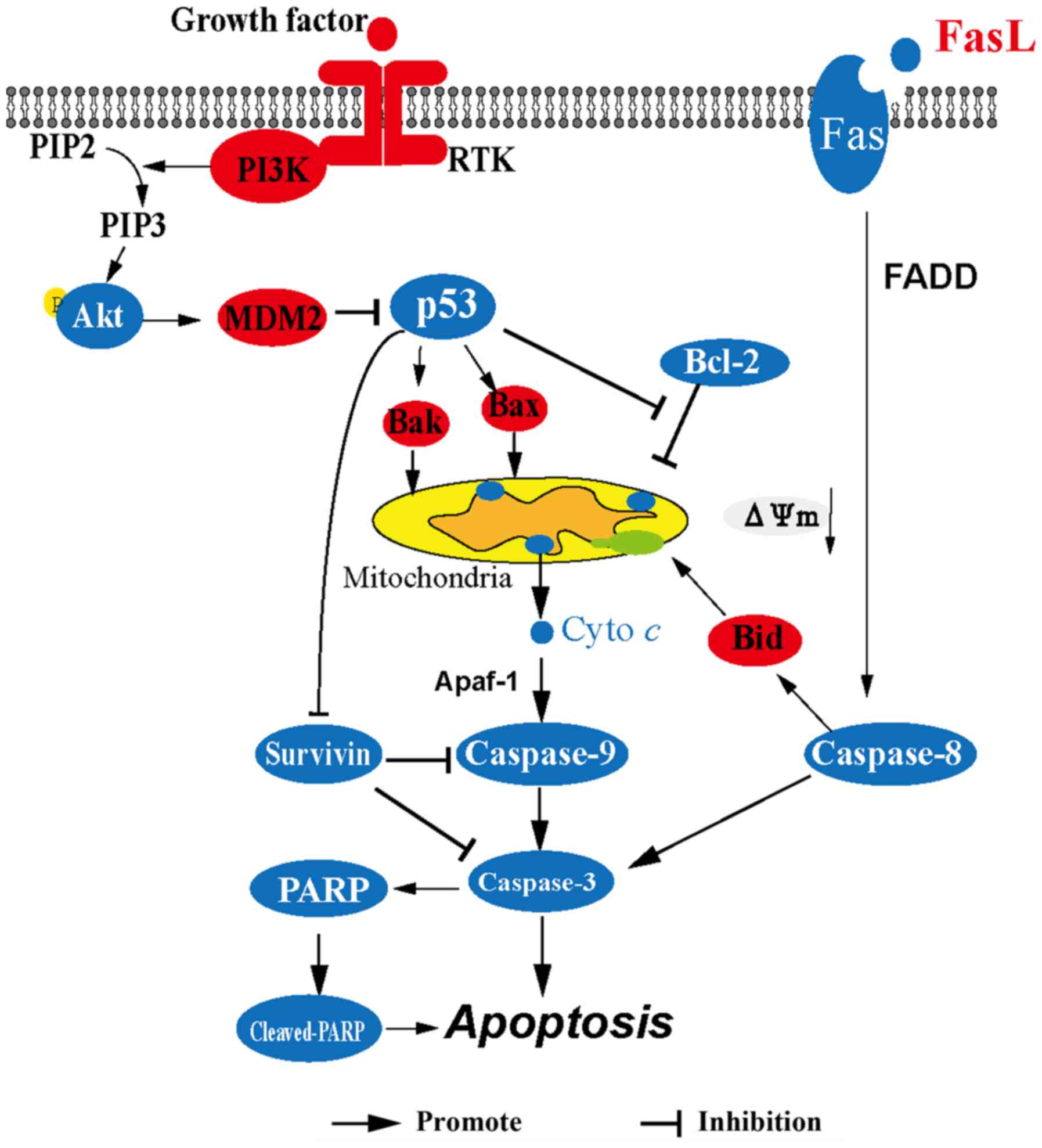
In summary (Fig. 7),
corilagin upregulates the expression of p53 leading to the release
of cytochrome c from the mitochondria to the cytoplasm and
then cytochrome c activates caspase-9, caspase-3 and
cleavage of PARP consequently resulting in the apoptosis of HCC
cells. In addition, corilagin activates the Fas/FasL/caspase-8
signaling pathway which can lead to the activation of caspase-3 and
the cleavage of Bid which contributes to the release of cytochrome
c. Downregulation of survivin also facilitates the activity
of caspase-9 and caspase-3. These results suggest that corilagin
significantly induces the apoptosis of HCC cells through both the
mitochondrial apoptotic pathway and the death receptor pathway. Our
findings are beneficial to the ongoing research on corilagin, and
provide evidence for the potential use of corilagin as an antitumor
drug.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81274149), the
Xiamen Municipal Science and Technology Innovation Fund Project of
China (grant no. 3502Z20130038), the Construction Projects of the
Top University at Fujian Agriculture and Forestry University of
China (grant no. 612014042), and the Science and Technology Plan
Projects of Xiamen (grant no. 3502Z20142003).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YM conceived and designed the experiments; YD, XL,
XL, ZZ, WH and LC performed the experiments; YD and YM analyzed the
data and wrote the paper; YM and QT revised the paper.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
P.
|
Phyllanthus
|
|
HCC
|
hepatocellular carcinoma
|
|
HBV
|
hepatitis B virus
|
|
CDDP
|
cis-diamminedichloroplatinum
(cisplatin)
|
|
IC50
|
50% inhibitory concentration
|
|
PI
|
propidium iodide
|
|
PARP
|
poly(ADP-ribose) polymerase
|
|
AO/EB
|
acridine orange/ethidium bromide
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(methyl thiazolyl tetrazolium)
|
|
PMSF
|
phenylmethylsulfonyl fluoride
|
References
|
1
|
Singh S, Singh PP, Roberts LR and Sanchez
W: Chemopreventive strategies in hepatocellular carcinoma. Nat Rev
Gastroenterol Hepatol. 11:45–54. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ming Y, Zheng Z, Chen L, Zheng G, Liu S,
Yu Y and Tong Q: Corilagin inhibits hepatocellular carcinoma cell
proliferation by inducing G2/M phase arrest. Cell Biol Int.
37:1046–1054. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jia L, Jin H, Zhou J, Chen L, Lu Y, Ming Y
and Yu Y: A potential anti-tumor herbal medicine, Corilagin,
inhibits ovarian cancer cell growth through blocking the TGF-β
signaling pathways. BMC Complement Altern Med. 13:332013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schmidt OT and Lademann R: Corilagin, ein
weiterer kristallisierter Gerbstoff aus Dividivi. X. Mitteilung
über natürliche Gerbstoffe. Justus Liebigs Ann Chem. 571:232–237.
1951.(In German). View Article : Google Scholar
|
|
5
|
Pham AT, Malterud KE, Paulsen BS, Diallo D
and Wangensteen H: DPPH radical scavenging and xanthine oxidase
inhibitory activity of Terminalia macroptera leaves. Nat
Prod Commun. 6:1125–1128. 2011.PubMed/NCBI
|
|
6
|
Yang F, Wang Y, Xue J, Ma Q, Zhang J, Chen
YF, Shang ZZ, Li QQ, Zhang SL and Zhao L: Effect of Corilagin on
the miR-21/smad7/ERK signaling pathway in a schistosomiasis-induced
hepatic fibrosis mouse model. Parasitol Int. 65:308–315. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong XR, Luo M, Fan L, Zhang T, Liu L,
Dong JH and Wu G: Corilagin inhibits the double strand
break-triggered NF-kappaB pathway in irradiated microglial cells.
Int J Mol Med. 25:531–536. 2010.PubMed/NCBI
|
|
8
|
Park JH, Joo HS, Yoo KY, Shin BN, Kim IH,
Lee CH, Choi JH, Byun K, Lee B, Lim SS, et al: Extract from
Terminalia chebula seeds protect against experimental
ischemic neuronal damage via maintaining SODs and BDNF levels.
Neurochem Res. 36:2043–2050. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duan W, Yu Y and Zhang L: Antiatherogenic
effects of phyllanthus emblica associated with corilagin and its
analogue. Yakugaku Zasshi. 125:587–591. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang MH, Vasquez Y, Ali Z, Khan IA and
Khan SI: Constituents from Terminalia species increase PPARα
and PPARγ levels and stimulate glucose uptake without enhancing
adipocyte differentiation. J Ethnopharmacol. 149:490–498. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gambari R, Hau DK, Wong WY and Chui CH:
Sensitization of Hep3B hepatoma cells to cisplatin and doxorubicin
by corilagin. Phytother Res. 28:781–783. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang BQ: Corilagin nanoparticle-induced
apoptosis in human gastric cancer SGC-7901 cells via the
mitochondrial pathway. Acta Pharmacol Sin. 34:14. 2013.
|
|
13
|
Kakiuchi N, Hattori M, Namba T, Nishizawa
M, Yamagishi T and Okuda T: Inhibitory effect of tannins on reverse
transcriptase from RNA tumor virus. J Nat Prod. 48:614–621. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Berry DE, MacKenzie L, Shultis EA, Chan JA
and Hecht SM: Naturally occurring inhibitors of topoisomerase I
mediated DNA relaxation. J Org Chem. 57:420–422. 1992. View Article : Google Scholar
|
|
15
|
Hecht SM, Berry DE, MacKenzie LJ, Busby RW
and Nasuti CA: A strategy for identifying novel, mechanistically
unique inhibitors of topoisomerase I. J Nat Prod. 55:401–413. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Komori A, Yatsunami J, Suganuma M, Okabe
S, Abe S, Sakai A, Sasaki K and Fujiki H: Tumor necrosis factor
acts as a tumor promoter in BALB/3T3 cell transformation. Cancer
Res. 53:1982–1985. 1993.PubMed/NCBI
|
|
17
|
Okabe S, Suganuma M, Imayoshi Y, Taniguchi
S, Yoshida T and Fujiki H: New TNF-alpha releasing inhibitors,
geraniin and corilagin, in leaves of Acer nikoense,
Megusurino-ki. Biol Pharm Bull. 24:1145–1148. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang YJ, Nagao T, Tanaka T, Yang CR,
Okabe H and Kouno I: Antiproliferative activity of the main
constituents from Phyllanthus emblica. Biol Pharm Bull.
27:251–255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Z, Wang D, Chen Y, Ren L, Li K and
Zhang W: Experiment studies on the pharmacodynamics experiment by
corilagin. Zhongliu Fangzhi Yanjiu. 29:356–358. 2002.
|
|
20
|
Chen Y and Ren L: Studies on the
anti-cancer active constituents of matsumura leafflower
(Phyllanthus matsumarae) II. Isolation and identification of
polyphenolic compounds. Chin Tradit Herbal Drugs. 28:198–202.
1997.
|
|
21
|
Zheng ZZ, Chen LH, Liu SS, Deng Y, Zheng
GH, Gu Y and Ming YL: Bioguided fraction and isolation of the
antitumor components from Phyllanthus niruri L. Biomed Res
Int. 2016:97292752016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jia L, Zhou J, Zhao H, Jin H, Lv M, Zhao
N, Zheng Z, Lu Y, Ming Y and Yu Y: Corilagin sensitizes epithelial
ovarian cancer to chemotherapy by inhibiting Snail glycolysis
pathways. Oncol Rep. 38:2464–2470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chazotte B: Labeling mitochondria with
JC-1. Cold Spring Harb Protoc. Sep 1–2011.(Epub ahead of print).
doi: 10.1101/pdb.prot065490. View Article : Google Scholar
|
|
24
|
Lampson BL and Davids MS: The development
and current use of BCL-2 inhibitors for the treatment of chronic
lymphocytic leukemia. Curr Hematol Malig Rep. 12:11–19. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shamsabadi FT, Eidgahi MR, Mehrbod P,
Daneshvar N, Allaudin ZN, Yamchi A and Shahbazi M: Survivin, a
promising gene for targeted cancer treatment. Asian Pac J Cancer
Prev. 17:3711–3719. 2016.PubMed/NCBI
|
|
26
|
Coutts AS, Adams CJ and La Thangue NB: p53
ubiquitination by Mdm2: A never ending tail? DNA Repair (Amst).
8:483–490. 2009. View Article : Google Scholar : PubMed/NCBI
|