Introduction
Angiogenesis represents a crucial event under both
physiological and pathological conditions (1). In tumors, in addition to traditional
angiogenesis, vasculogenic mimicry (VM) provides another mechanism
by which tumor cells can obtain nutrients to survive (2). VM defines the ability of highly
invasive tumor cells to form matrix-rich networks containing
channels; these vascular channels lack an endothelial cell (EC)
lining and have a basement membrane that stains positive with
periodic acid-Schiff (PAS) reagent (3). It has been recognized that VM is not
only involved in proliferation and metastatic potential but is also
associated with a poor patient prognosis in numerous types of
tumors, including melanoma, hepatoma, breast carcinoma, prostatic
carcinoma, lung carcinoma, colorectal cancer and ovarian carcinoma
(4,5).
Since the introduction of VM in 1999 as a novel
paradigm for tumor perfusion, many studies have contributed new
insights into the underlying molecular pathways supporting VM. It
has been established that vascular endothelial growth factor A
(VEGFA) plays a crucial role in the formation of VM (6–8). VEGFA
binds and activates two tyrosine kinase receptors, VEGF receptor 1
and 2 (VEGFR1 and VEGFR2). Some reports have shown that VEGFR2
combines with VEGFA in an autocrine or paracrine manner and
possesses many signaling capacities in the formation of VM
(9–11). In contrast with these findings,
other studies have shown that VEGFR1 kinase influences VM formation
independent of VEGFR2 (12,13). These studies suggest that there are
different molecular mechanisms involved in different tumor cell
types.
Nasopharyngeal carcinoma (NPC) is endemic in
Southern China and Southeast Asia. Approximately 80% of NPC
patients are infected with the Epstein-Barr virus (EBV); EBV
encodes latent membrane protein 1 (LMP1) which is believed to play
a key role in the pathogenesis of NPC (14). LMP1 activates various downstream
oncogenic signaling pathways and induces various downstream
pathological changes in the processes of cell proliferation,
anti-apoptosis and metastasis (15). In addition, a clear correlation
between LMP1 and angiogenesis has been observed in NPC, and this
effect is attributed to VEGF secretion via a mechanism involving
the induction of cyclooxygenase 2 (COX-2) and hypoxia inducible
factor 1α (HIF-1α) (16,17). Our previous studies also
demonstrated that LMP1 increased VEGFA expression and upregulated
angiogenesis in NPC (18,19). These reports suggest the possibility
that LMP1 is involved in VM formation in NPC.
Although VM has been observed in numerous types of
tumors, whether VM occurs in NPC and the mechanisms involved in its
generation have not been well defined. In this study, we tested the
roles of LMP1 and VEGFA/VEGFR signaling in VM formation and
investigated the correlation between the LMP1-mediated upregulation
of VM and the prognosis of NPC patients.
Materials and methods
Materials
The materials and reagents used in this study were
purchased from the following providers. The Periodic Acid-Schiff
(PAS) kit was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany); Matrigel was obtained from BD Biosciences (Franklin
Lakes, NJ, USA). Primary anti-VEGFA (sc-152), anti-VEGFR1 (sc-316),
anti-VEGFR2 (sc-505) and anti-β-actin (sc-8432) polyclonal
antibodies; VEGFA small interfering RNA (siRNA); VEGFR1 siRNA; and
VEGFR2 siRNA were purchased from Santa Cruz Biotechnology Inc.
(Santa Cruz, CA, USA). Anti-LMP1 antibody was purchased from
Dako/Agilent (M0897; Agilent Technologies, Inc., Santa Clara, CA,
USA); anti-CD34 antibody was purchased from Abcam Corp. (cat. no.
ab81289; Cambridge, UK); and the HistoMouse-SP Broad Spectrum DAB
kit was purchased from Invitrogen-Zymed (Thermo Fisher Scientific,
Inc., Waltham, MA, USA).
Cell culture and small interfering RNA
(siRNA) transfection
HK1 is an LMP1-negative NPC cell line, and HK1-LMP1
is a stable LMP1-integrated cell line (20). Cells were grown in Gibco™ RPMI-1640
medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS). Cells at 60–70%
confluence were transfected with siRNA using Invitrogen™
Lipofectamine™ 2000 (Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions.
In vitro tube formation assay
A Matrigel tube formation assay was developed for
testing tubular structure formation. Briefly, we transfected
HK1-LMP1 cells with targeted siRNA (LMP1, VEGFA, VEGFR1, or VEGFR2)
and control siRNA (CON). Cells were trypsinized after 24 h and
centrifuged at 600 × g for 5 min. Approximately 1×105
cells were seeded in each well of a 24-well plate with 200 µl of
embedded Matrigel. Next, the cells were incubated for 8 h, and the
extent of tubular structure formation was examined using an
inverted microscope (CKX41SF; Olympus Corp., Tokyo, Japan).
Western blot analysis
Cells were harvested and lysed at 4°C for 15 min in
lysis buffer, and the protein concentration was determined by the
Bradford protein assay (Bio-Rad Laboratories, Hercules, CA, USA)
according to the manufacturer's instructions. Proteins were then
separated by SDS-PAGE (4–20% Mini-PROTEAN® TGX™ Precast
Protein Gels (Bio-Rad Laboratories) and transferred to a
nitrocellulose membrane (GE Healthcare, Piscataway, NJ, USA).
Membranes were blocked with TBS-T (20 mM Tris-HCl, pH 7.4, 137 mM
NaCl and 0.1% Tween-20) containing 5% non-fat milk for 1 h at room
temperature (RT). Then, the membranes were incubated with primary
antibodies (LMP1, dilution 1:250; VEGFA, dilution 1:250; VEGFR1,
dilution 1:500; VEGFR2, dilution 1:500), followed by incubation
with horseradish peroxidase-conjugated secondary antibody (cat.
nos. sc-2005 or sc-2004; Santa Cruz Biotechnology) for 1 h at RT
and visualization with an enhanced chemiluminescence detection kit
(Pierce ECL; Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA).
The relative protein levels were quantified using ImageJ software
(National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence analysis
The cells were fixed and permeabilized with cold
methyl alcohol (−20°C) for 10 min, and then blocked in 5% donkey
serum in PBS for 1 h and incubated with the primary antibody in PBS
containing 1% BSA at 4°C overnight. The cells were washed 3 times
with PBS, and incubated for 30 min with fluorochrome-conjugated
secondary antibody (cat. nos. A-11001 or A-21207; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) for 30 min. For fluorescence
analysis, cell samples were visualized on a laser scanning confocal
microscopy with appropriate emission filters (LSM 510 META; Carl
Zeiss, Oberkochen, Germany).
Immunohistochemical analysis and
CD34/PAS double staining
The NPC tissue array was purchased from Pantomics
(Richmond, CA, USA), and the NPC paraffin-embedded tumor tissue
samples, clinical details, and follow-up data were obtained from
the Pathology Department of Xiangya Hospital from 2009 to 2015. The
institutional review board of the Xiangya Hospital Ethics Committee
approved the use of human samples in this study. IHC staining was
performed using a HistoMouse-SP Broad Spectrum DAB kit (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to standard
protocols. The signal was detected using a diaminobenzidine
solution. A semi-quantitative evaluation of the positivity of each
protein by IHC was performed using a previously defined method
(21). The percentage of positive
cells was divided into five grades (percentage scores): 0, ≤10%; 1,
11–25%; 2, 26–50%; 3, 51–75%; and 4, >75%. The intensity of
staining was divided into four grades (intensity scores): 0, no
staining; 1, light brown; 2, brown; and 3, dark brown. Staining
positivity was determined by the following formula: Overall score =
percentage score × intensity score. The stained sections were
independently examined by two of the authors (Z.Z. and B.L.).
Scores of 3 to 12 were considered positive for LMP1 and VEGFR1
expression. For CD34/PAS double staining, after IHC staining for
CD34 as described above, the sections were washed with running
water for 6 min, incubated with PAS for 15 min, and counterstained
with hematoxylin. To quantify the differences in the density of VM,
we calculated the total number of VMs in five fields for each tumor
dot or section.
Statistical analysis
Statistical analyses were performed using the
Student's t-test. The Kaplan-Meier method was used to estimate
progression-free survival, and the log-rank test was used to
evaluate differences between survival curves. A P-value of <0.05
was considered statistically significant.
Results
EBV-LMP1 is involved in the tubular
structure formation in NPC cells and is related to VEGFA
expression
Highly aggressive tumor cells form patterned
networks of matrix-rich tubular structures when cultured on
Matrigel (4). In this experiment,
we used an in vitro tube formation assay to evaluate the
tubular structure formation ability of different NPC tumor cells.
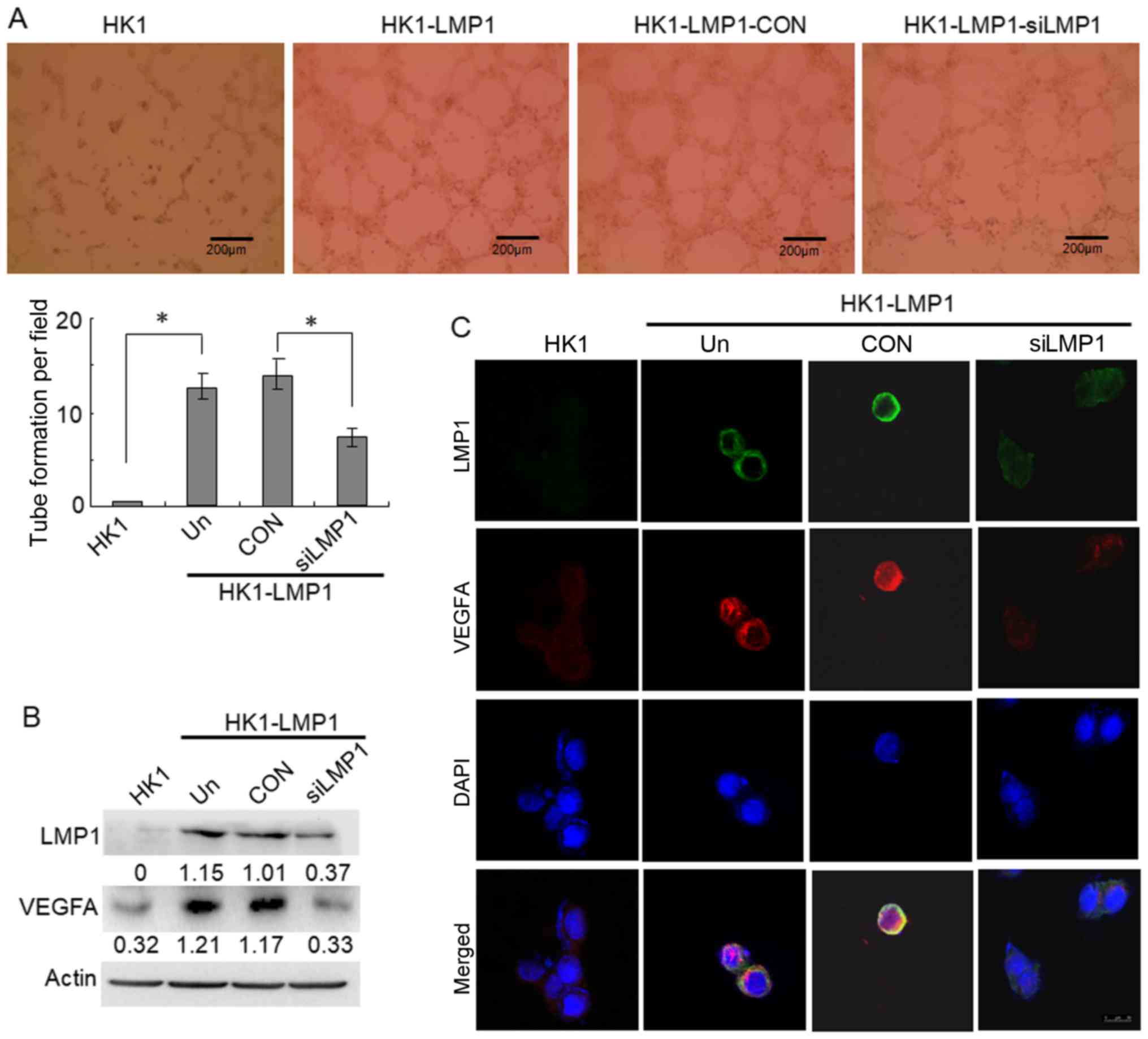
The results showed that it was almost impossible for the
LMP1-negative NPC cells, HK1, to form tubular structures in
Matrigel, whereas the stable LMP1-integrated cells, HK1-LMP1, were
able to form tubular structures. The tubular structure forming
ability of the HK1-LMP1 cells decreased by approximately one-half
after treatment with LMP1 siRNA compared with the control siRNA
(CON) group (P<0.05) (Fig. 1A).
Thus, the data indicated that LMP1 might contribute to tumor VM
formation in NPC.
Moreover, the western blotting results showed that
VEGFA was highly expressed in the HK1-LMP1 cells compared to the
level in the HK1 cells (Fig. 1B).
Furthermore, when HK1-LMP1 cells were treated with LMP1 siRNA, LMP1
expression was reduced compared to that noted in the CON group, and
this reduction was accompanied by decreased VEGFA expression.
Meanwhile, the data of the immunofluorescence assay showed that,
there was a higher VEGFA protein expression in HK1-LMP1 cells
compared to that in the HK1 cells, and the co-localization of LMP1
and VEGFA was in the cytoplasm of NPC cells. Following inhibition
of LMP1 expression by siRNA, both LMP1 and VEGFA expression was
reduced compared to the CON group, and the co-localization of LMP1
and VEGF become very weak (Fig.
1C). These results indicated that EBV-LMP1 is involved in the
tubular structure formation in NPC cells and is related to VEGFA
expression.
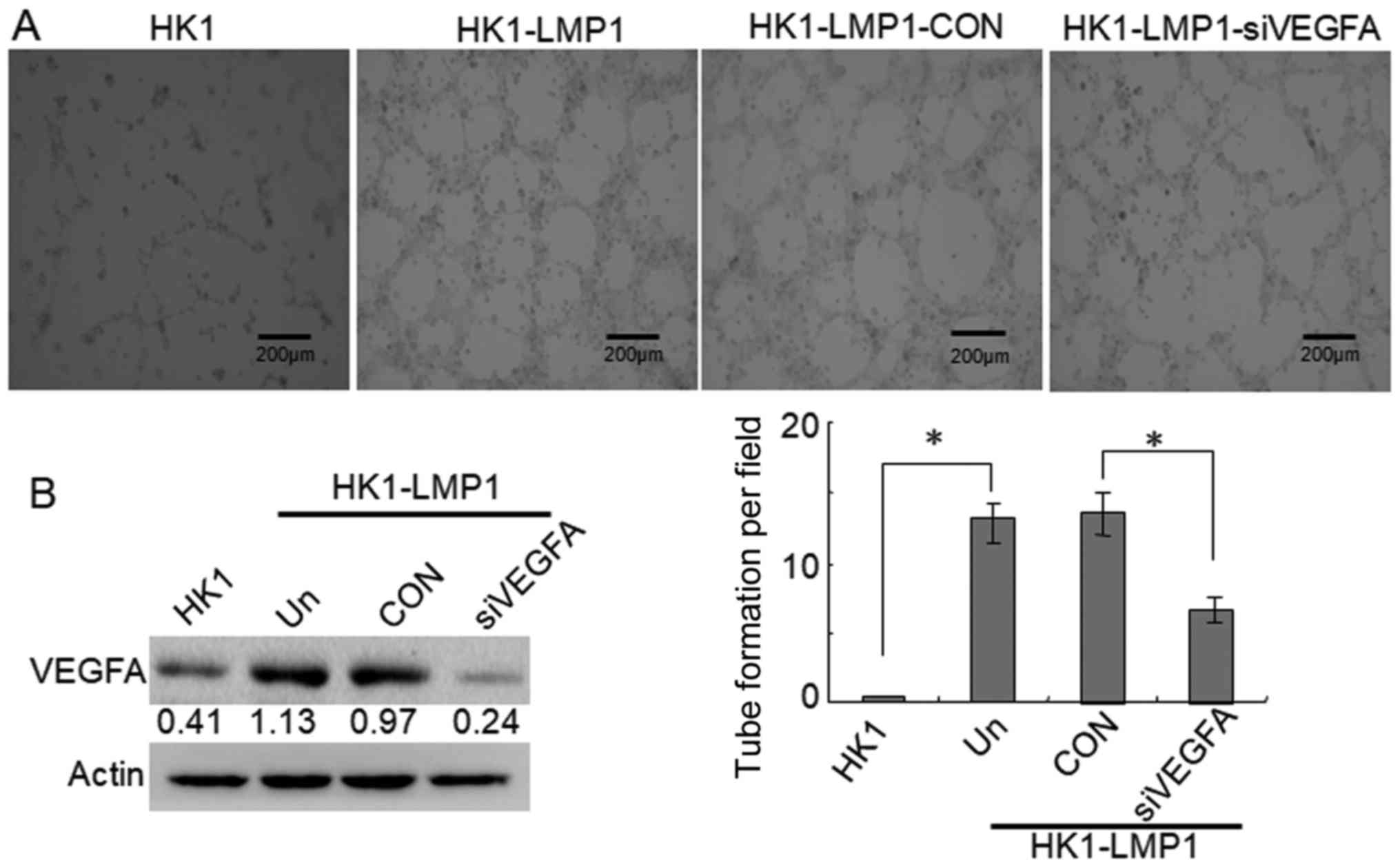
Furthermore, we investigated the contribution of
VEGFA to tubular structure formation in LMP1-positive NPC cells.
The data showed that VEGFA siRNA also strongly inhibited the
formation of tubular structures compared with the CON group in the
in vitro tube formation assays (P<0.05) (Fig. 2A). Meanwhile, VEGFA was highly
expressed in HK1-LMP1 cells compared to that noted in the HK1
cells, and VEGFA siRNA significantly reduced VEGFA expression
compared with that noted in the CON group (Fig. 2B). These findings indicated that
VEGFA is involved in VM formation in LMP1-positive NPC cells.
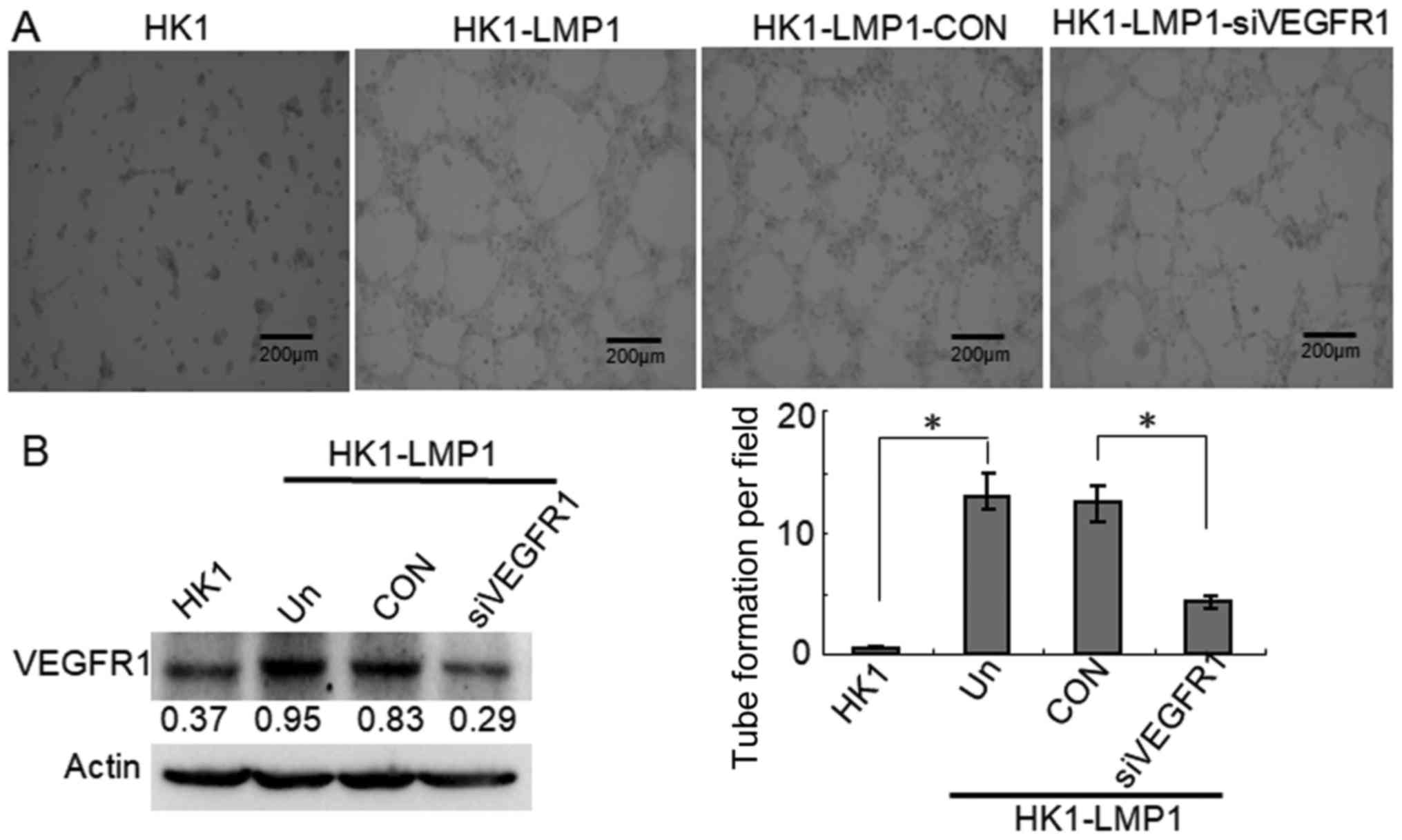
VEGFR1 but not VEGFR2 is required for
LMP1-mediated tubular structure formation in NPC cells
VEGFA performs its biological function mainly
through binding and activating its receptor VEGFRs, VEGFR1 and
VEGFR2, which have been shown to be involved in VM formation in
different tumor cells (6,12). Thus, we determined which VEGFR is
required for the tubular structure formation in human LMP1-positive
NPC cells. The data showed that VEGFR1 siRNA also strongly
inhibited the formation of tubular structures compared with the CON
group in in vitro tube formation assays (P<0.05)
(Fig. 3A). Furthermore, VEGFR1 was
highly expressed in HK1-LMP1 cells compared to that observed in the
HK1 cells. Moreover, VEGFR1 siRNA significantly reduced VEGFR1
expression compared with the CON group (Fig. 3B). These findings indicated that
VEGRA/VEGFR1 signaling was involved in VM formation in
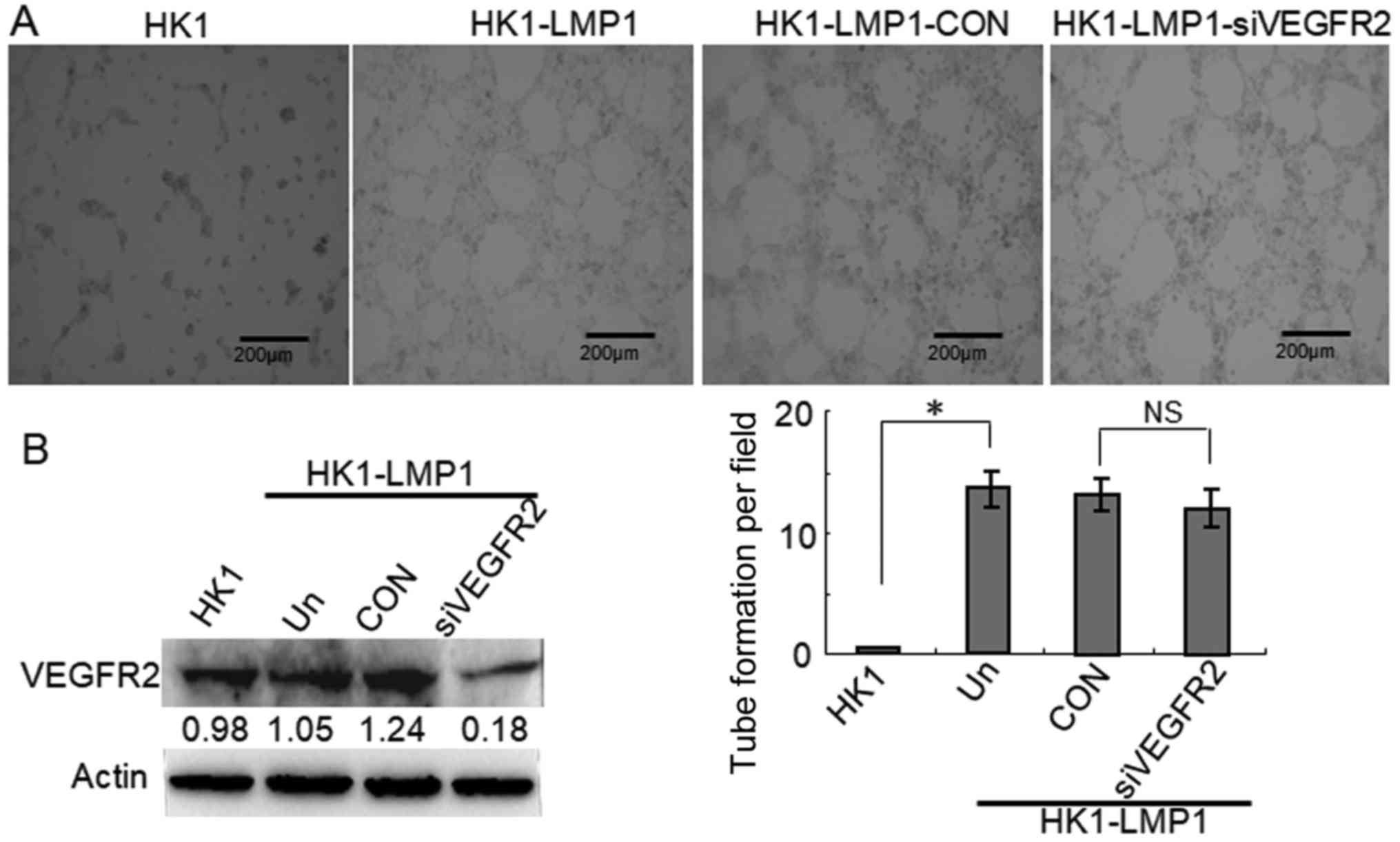
LMP1-positive NPC cells. In contrast, HK1-LMP1 cells were
transfected with VEGFR2 siRNA, which decreased the expression of
VEGFR2. However, we did not observe any changes in the main
geometrical features of the tubular structures that formed compared
with the CON group (Fig. 4). These
findings indicated that VEGFA/VEGFR2 signaling was not involved in
the tubular structure formation in LMP1-positive NPC cells.
According to the above western blotting results, VEGFA and VEGFR1
were significantly increased in LMP1-positive cells compared with
LMP1-negative cells, whereas VEGFR2 did not change substantially.
Therefore, we hypothesized that LMP1-VEGFA functions in the
formation of VM through VEGFR1 rather than VEGFR2, which may be
related to the proteins that are regulated by LMP1.
LMP1 upregulates VEGFR1 expression and
VM formation in primary NPCs
The VM channels consist of a basement membrane with
a lining of tumor cells on the external wall and do not contain
endothelial cells (ECs) on the inner wall. Thus, PAS-positive and
CD34-negative mature tumor vessels form a patterned network in the
VM morphology (2). In further
support of the preferential association of EBV-LMP1 and its
downregulation of VEGFA/VEGFR1 signaling with VM in NPC, we
examined the expression levels of LMP1 and VEGFR1 in a commercial
NPC tissue array, and VM was detected in the tumor tissue array
using CD34/PAS double staining. As shown in Fig. 5A, the VM channels were composed of
NPC cells and were PAS-positive, CD34-negative (indicated by black
arrows), while the endothelial-dependent vessels were positive for
both CD34 and PAS (indicated by red arrows). These results showed
that VM occurred in EBV-LMP1-positive NPCs and that hardly any VM
occurred in LMP1-negative NPCs. There was a positive correlation
between LMP1 expression and VM formation according to the Pearson
correlation coefficient results (r=0.397, P=0.002). Meanwhile, the
data showed a significant correlation between LMP1 and VEGFR1
expression (correlation coefficient=0.481, P=0.000) (Fig. 5B). These results indicated that LMP1
expression was associated with VEGFR1 and VM formation in NPCs.
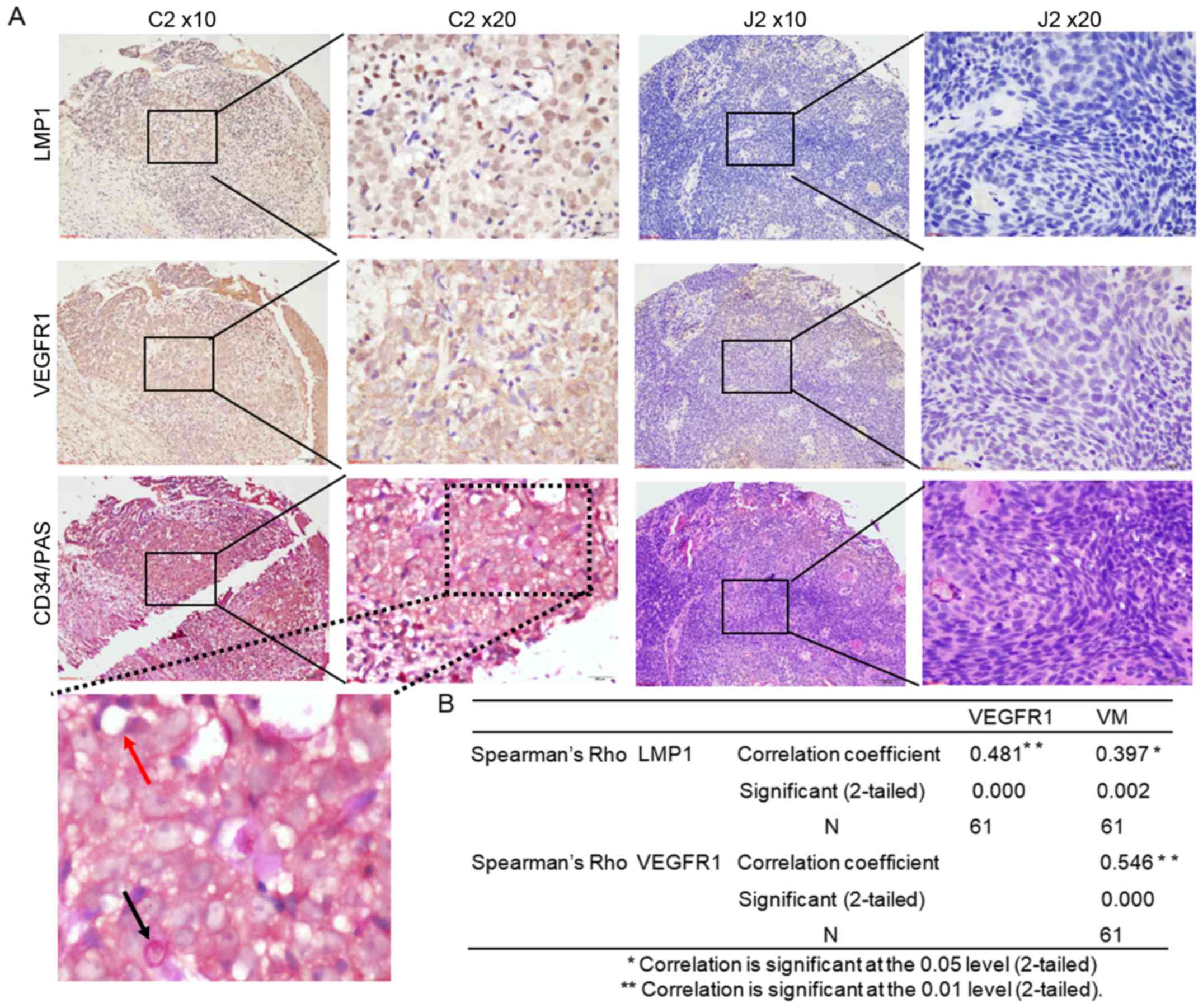 | Figure 5.LMP1 upregulates VEGFR1 expression and
VM formation in an NPC tissue array. (A) Immunohistochemical
analysis was performed to examine the levels of LMP1 and VEGFR1
expression, and CD34/PAS double staining was used to detect VM
formation in an NPC tissue array (magnification, ×100 and ×200).
The presence of PAS-positive and CD34-negative cells indicated VM
channels, which were lined with tumor cells (black arrows). The
endothelial-dependent vessels were positive for both CD34 and PAS
(red arrows). The tissue dot C2 exhibited high expression of LMP1
and VEGFR1 and was positive for VM, whereas tissue dot J2 exhibited
low levels of LMP1 and VEGFR1 and was negative for VM. (B)
Correlations between LMP1, VEGFR1, and VM formation in the NPC
tissue array. Asterisks (* or **) indicates a significant (0.05,
2-tailed or 0.01, 2-tailed, respectively) correlation. LMP1, latent
membrane protein 1; VEGFR1, vascular endothelial growth factor
receptor 1; VM, vascular mimicry; NPC, nasopharyngeal carcinoma;
PAS, periodic acid-Schiff. |
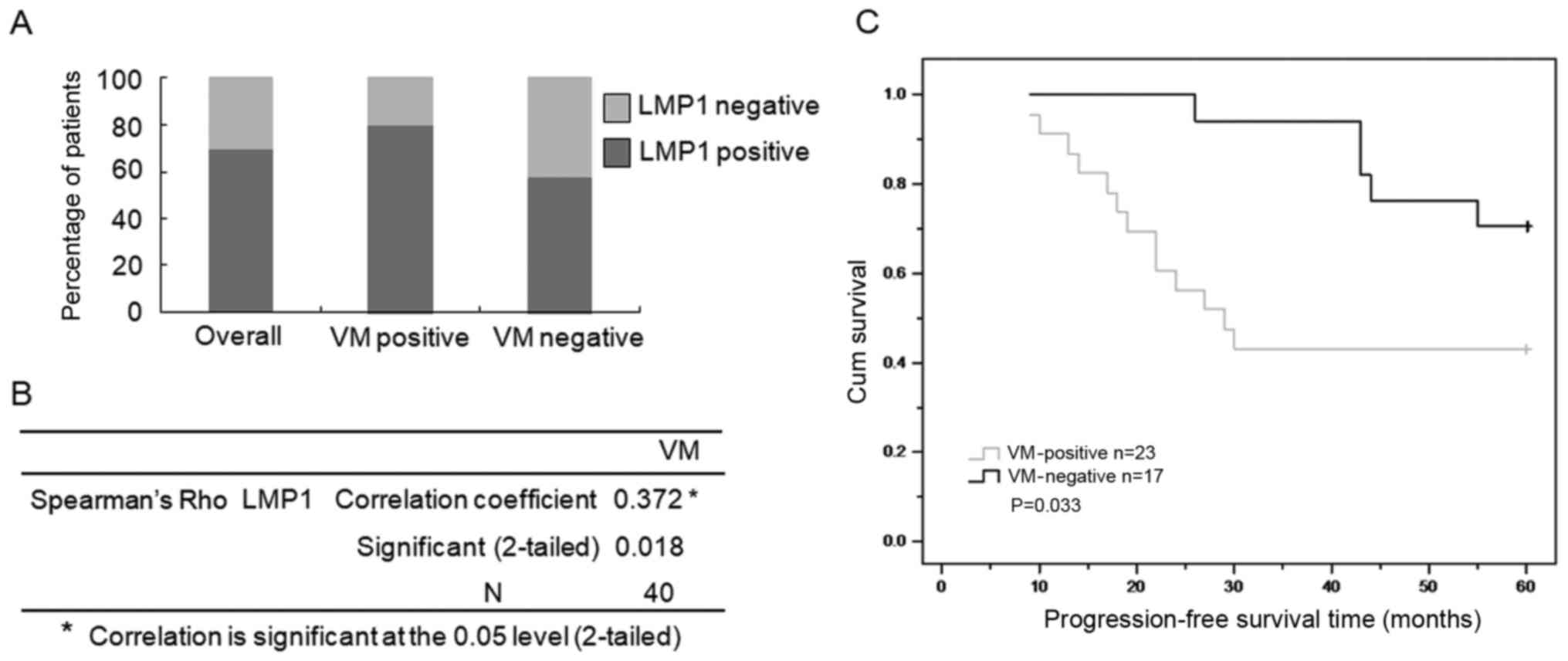
Presence of VM in tumor tissues is
associated with a poor clinical outcome
The presence of VM in malignant tumors is associated
with increased patient mortality (3). Thus, we determined whether the
LMP1-mediated upregulation of VM is associated with the prognosis
of NPC patients. We examined the expression levels of LMP1 and VM
formation in tumor tissue samples from 40 NPC patients. These
patients were successfully followed up (a median follow-up period
of 5.61 years) (Table I). The
positive staining rate of LMP1 in NPC tissues was 70% (28/40). The
positive staining rate of VM in NPC tissues samples was 57.5%
(23/40). There was a significant correlation between LMP1
expression and VM formation according to the Pearson correlation
coefficient (r=0.372, P=0.018). These results indicated that LMP1
expression is associated with VM formation in NPC tumor tissues,
consistent with the results from the commercial NPC tissue array
(Fig. 6A and B).
 | Table I.Clinical characteristics of the NPC
patients (N=40). |
Table I.
Clinical characteristics of the NPC
patients (N=40).
| Characteristics | No. of patients
(%) |
|---|
| Sex |
|
| Male | 29 (72.5) |
|
Female | 11 (27.5) |
| Age (years) |
|
|
>45 | 18 (45.0) |
| ≤45 | 22 (55.0) |
| Clinical stage |
|
| I | 1
(2.5) |
|
II–III | 23 (57.5) |
| IV | 16 (40) |
| Therapeutic
modality |
|
| Radiation
therapy | 31 (77.5) |
|
Concomitant
chemoradiotherapy | 7
(17.5) |
| No
treatment | 2
(5.0) |
| Histological
classification |
|
|
NKCC | 38 (95) |
|
KSCC | 2
(5.0) |
| Follow-up time
(years) |
|
|
Average |
5.61 |
| PFS
rate (%) 5-year |
55% |
Using the clinical follow-up data, we
retrospectively analyzed the prognostic significance of VM
formation on the progression-free survival time (PFS) of 40 NPC
patients, who received radiation therapy or concomitant
chemoradiotherapy. As shown in Fig.
6C, the results of the Kaplan-Meier method analysis with
log-rank test revealed a statistically significant difference in
PFS between the 23 patients in the VM-positive group (78.3% with
expression of LMP1, median survival time of 37.12 months) and the
17 patients in the VM-negative group (58.9% with expression of
LMP1, median survival time of 54.81 months). These results not only
indicated that VM was upregulated along with LMP1, but also
suggested that VM formation was associated with a worse clinical
outcome following therapy.
Discussion
Tumor growth depends strongly on the formation of
new blood vessels for their supply of oxygen and nutrients through
not only angiogenesis but also VM (4). Although VM has been reported in many
types of tumors, no data were available in the literature regarding
VM and the precise signaling mechanisms involved in VM formation in
NPC. In the present study, we used the HK1 cell line and a
corresponding cell line stably expressing LMP1 to carry out the
experiment. The results revealed several novel insights. First, VM
was shown to occur in LMP1-positive NPC. Second, VEGFA/VEGFR1
signaling was required for LMP1-mediated VM formation, and this
pathway was not dependent on VEGFR2 kinase. Finally, VM formation
was associated with a poor prognosis in NPC cases. Taken together,
our results provide novel insights into the regulatory mechanisms
underlying VM and specifically identify LMP1 and VEGFR1 as
potential targets in NPC.
VEGFA is secreted by almost all tumor cells and
belongs to the angiogenic growth factor family associated with
tumor angiogenesis. The binding of VEGFA to its ligand and the
activation of VEGFR1 and/or VEGFR2 results in cell proliferation,
angiogenesis, and VM formation in tumors (6). The role of VEGFA/VEGFR signaling in
regulating VM has been widely studied; however, it remains
controversial. It is generally believed that VEGFA binds to VEGFR2
to trigger angiogenesis and VM formation in tumors (9–13,22).
In contrast with these findings, several reports identified the
requirement of VEGFR1 for VM (12,13).
These discrepancies regarding the role of VEGFA/VEGFR signaling in
modulating VM formation may reflect cell type- and/or cell
line-specific differences. To gain a better understanding of the
biological relevance of VEGFR subtypes on tubular network formation
induced by EBV-LMP1, in our study, we used western blot assay, and
the results showed that VEGFA and VEGFR1 were significantly
increased in LMP1-positive cells compared with LMP1-negative cells,
whereas VEGFR2 did not change substantially. We further used an
in vitro tube formation assay and siRNA experiments, and the
results suggested that VEGFR1 was the sole VEGFA receptor involved
in signaling during VM formation in NPC. Therefore, we propose that
LMP1-VEGFA functioned in the formation of VM through VEGFR1 rather
than VEGFR2, which may be related to the proteins that are
regulated by LMP1.
VM provides a sufficient blood supply for various
malignant tumors to support their proliferation, invasion and
metastasis (23). Some studies have
shown that VM formation is significantly linked with a poor
prognosis for patients with aggressive tumors, including melanoma,
colorectal cancer, lung cancer, sarcomas and hepatic cancer
(24–26). However, several studies have
demonstrated that VM has no significant association with tumor
prognosis, although a shorter survival time was observed in
VM-positive patients (27,28). Therefore, the influence of VM on
cancer patient survival remains controversial. The results of our
experiments not only indicated that VM was upregulated along with
EBV-LMP1, but also suggested that VM formation was associated with
a worse clinical outcome following therapy.
LMP1 has been shown to induce VEGFA production and
increase angiogenesis in NPC tumors (18,29).
We previously reported that knockdown of LMP1 inhibited the
expression and secretion of VEGFA through the JNK/c-Jun signaling
pathway in NPC cells and enhanced the radiosensitivity of NPC cells
by inhibiting HIF-1/VEGF activity (19). Collectively, EBV-LMP1 promotes
angiogenesis in nasopharyngeal carcinoma, and also mediates the
formation of VM, thereby providing nutrients for rapidly growing
tumors. Further research of the molecular events underlying VM will
offer new insights into the development of therapeutic strategies
for EBV-LMP1-positive NPC.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the
National Natural Science Foundation of China (nos. 81372182 and
81672761) and the Fundamental Research Funds for the Central
Universities of Central South University (no. 1053320170401).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
LY designed the research. SX, JB and ZZ performed
the research and wrote the manuscript. All authors analysed the
data and were involved in writing the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of the Xiangya Hospital Ethics Committee (Changsha,
China).
Consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Priya Krishna S, Nagare RP, Sneha VS,
Sidhanth C, Bindhya S, Manasa P and Ganesan TS: Tumour
angiogenesis-Origin of blood vessels. Int J Cancer. 139:729–735.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang S, Zhang D and Sun B: Vasculogenic
mimicry: Current status and future prospects. Cancer Lett.
254:157–164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maniotis AJ, Folberg R, Hess A, Seftor EA,
Gardner LM, Pe'er J, Trent JM, Meltzer PS and Hendrix MJ: Vascular
channel formation by human melanoma cells in vivo and in vitro:
Vasculogenic mimicry. Am J Pathol. 155:739–752. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qiao L, Liang N, Zhang J, Xie J, Liu F, Xu
D, Yu X and Tian Y: Advanced research on vasculogenic mimicry in
cancer. J Cell Mol Med. 19:315–326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Williamson SC, Metcalf RL, Trapani F,
Mohan S, Antonello J, Abbott B, Leong HS, Chester CP, Simms N,
Polanski R, et al: Vasculogenic mimicry in small cell lung cancer.
Nat Commun. 7:133222016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kirschmann DA, Seftor EA, Hardy KM, Seftor
RE and Hendrix MJ: Molecular pathways: vasculogenic mimicry in
tumor cells: diagnostic and therapeutic implications. Clin Cancer
Res. 18:2726–2732. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schnegg CI, Yang MH, Ghosh SK and Hsu MY:
Induction of vasculogenic mimicry overrides VEGF-A silencing and
enriches stem-like cancer cells in melanoma. Cancer Res.
75:1682–1690. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang JY, Sun T, Zhao XL, Zhang SW, Zhang
DF, Gu Q, Wang XH, Zhao N, Qie S and Sun BC: Functional
significance of VEGF-a in human ovarian carcinoma: Role in
vasculogenic mimicry. Cancer Biol Ther. 7:758–766. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Karroum A, Mirshahi P, Faussat AM,
Therwath A, Mirshahi M and Hatmi M: Tubular network formation by
adriamycin-resistant MCF-7 breast cancer cells is closely linked to
MMP-9 and VEGFR-2/VEGFR-3 over-expressions. Eur J Pharmacol.
685:1–7. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yao X, Ping Y, Liu Y, Chen K, Yoshimura T,
Liu M, Gong W, Chen C, Niu Q, Guo D, et al: Vascular endothelial
growth factor receptor 2 (VEGFR-2) plays a key role in vasculogenic
mimicry formation, neovascularization and tumor initiation by
glioma stem-like cells. PLoS One. 8:e571882013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang S, Fu Z, Wei J, Guo J, Liu M and Du
K: Peroxiredoxin 2 is involved in vasculogenic mimicry formation by
targeting VEGFR2 activation in colorectal cancer. Med Oncol.
32:4142015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Frank NY, Schatton T, Kim S, Zhan Q,
Wilson BJ, Ma J, Saab KR, Osherov V, Widlund HR, Gasser M, et al:
VEGFR-1 expressed by malignant melanoma-initiating cells is
required for tumor growth. Cancer Res. 71:1474–1485. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vartanian A, Stepanova E, Grigorieva I,
Solomko E, Baryshnikov A and Lichinitser M: VEGFR1 and PKCα
signaling control melanoma vasculogenic mimicry in a VEGFR2
kinase-independent manner. Melanoma Res. 21:91–98. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Young LS, Yap LF and Murray PG:
Epstein-Barr virus: More than 50 years old and still providing
surprises. Nat Rev Cancer. 16:789–802. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang LW, Jiang S and Gewurz BE:
Epstein-Barr virus LMP1-mediated oncogenicity. J Virol. 91:912017.
View Article : Google Scholar
|
|
16
|
Kondo S, Seo SY, Yoshizaki T, Wakisaka N,
Furukawa M, Joab I, Jang KL and Pagano JS: EBV latent membrane
protein 1 up-regulates hypoxia-inducible factor 1alpha through
Siah1-mediated down-regulation of prolyl hydroxylases 1 and 3 in
nasopharyngeal epithelial cells. Cancer Res. 66:9870–9877. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Murono S, Inoue H, Tanabe T, Joab I,
Yoshizaki T, Furukawa M and Pagano JS: Induction of
cyclooxygenase-2 by Epstein-Barr virus latent membrane protein 1 is
involved in vascular endothelial growth factor production in
nasopharyngeal carcinoma cells. Proc Natl Acad Sci USA.
98:6905–6910. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z, Luo F, Li L, Yang L, Hu D, Ma X,
Lu Z, Sun L and Cao Y: STAT3 activation induced by Epstein-Barr
virus latent membrane protein1 causes vascular endothelial growth
factor expression and cellular invasiveness via JAK3 And ERK
signaling. Eur J Cancer. 46:2996–3006. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang L, Liu L, Xu Z, Liao W, Feng D, Dong
X, Xu S, Xiao L, Lu J, Luo X, et al: EBV-LMP1 targeted DNAzyme
enhances radiosensitivity by inhibiting tumor angiogenesis via the
JNKs/HIF-1 pathway in nasopharyngeal carcinoma. Oncotarget.
6:5804–5817. 2015.PubMed/NCBI
|
|
20
|
Xiao L, Hu ZY, Dong X, Tan Z, Li W, Tang
M, Chen L, Yang L, Tao Y, Jiang Y, et al: Targeting Epstein-Barr
virus oncoprotein LMP1-mediated glycolysis sensitizes
nasopharyngeal carcinoma to radiation therapy. Oncogene.
33:4568–4578. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu J, Wang N and Wang YJ: XRCC3 and RAD51
expression are associated with clinical factors in breast cancer.
PLoS One. 8:e721042013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Holmes K, Roberts OL, Thomas AM and Cross
MJ: Vascular endothelial growth factor receptor-2: Structure,
function, intracellular signalling and therapeutic inhibition. Cell
Signal. 19:2003–2012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hendrix MJ, Seftor EA, Seftor RE, Chao JT,
Chien DS and Chu YW: Tumor cell vascular mimicry: Novel targeting
opportunity in melanoma. Pharmacol Ther. 159:83–92. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tan LY, Mintoff C, Johan MZ, Ebert BW,
Fedele C, Zhang YF, Szeto P, Sheppard KE, McArthur GA, Foster-Smith
E, et al: Desmoglein 2 promotes vasculogenic mimicry in melanoma
and is associated with poor clinical outcome. Oncotarget.
7:46492–46508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bissanum R, Lirdprapamongkol K, Svasti J,
Navakanitworakul R and Kanokwiroon K: The role of WT1 isoforms in
vasculogenic mimicry and metastatic potential of human triple
negative breast cancer cells. Biochem Biophys Res Commun.
494:256–262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun J, Sun B, Sun R, Zhu D, Zhao X, Zhang
Y, Dong X, Che N, Li J, Liu F, et al: HMGA2 promotes vasculogenic
mimicry and tumor aggressiveness by upregulating Twist1 in gastric
carcinoma. Sci Rep. 7:22292017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Massi D, Franchi A, Paglierani M, Ketabchi
S, Borgognoni L, Reali UM and Santucci M: Vasculogenic mimicry has
no prognostic significance in pT3 and pT4 cutaneous melanoma. Hum
Pathol. 35:496–502. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shirakawa K, Wakasugi H, Heike Y, Watanabe
I, Yamada S, Saito K and Konishi F: Vasculogenic mimicry and
pseudo-comedo formation in breast cancer. Int J Cancer. 99:821–828.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoshizaki T, Kondo S, Wakisaka N, Murono
S, Endo K, Sugimoto H, Nakanishi S, Tsuji A and Ito M: Pathogenic
role of Epstein-Barr virus latent membrane protein-1 in the
development of nasopharyngeal carcinoma. Cancer Lett. 337:1–7.
2013. View Article : Google Scholar : PubMed/NCBI
|